
Effect of Fe and EDTA on Freshwater Cyanobacteria Bloom Formation


National Engineering Laboratory for Circular Economy, Department of Environmental Science and Engineering, Fudan University, Shanghai 200433, China
*
Author to whom correspondence should be addressed.
Water 2017, 9(5), 326; https://doi.org/10.3390/w9050326
Submission received: 19 February 2017
/
Revised: 13 April 2017
/
Accepted: 2 May 2017
/
Published: 5 May 2017
Abstract
:Due to the fact that not all eutrophic lakes have cyanobacteria blooms, we hypothesized Fe may be another important limiting factor which regulates cyanobacteria bloom formation. We tested the hypothesis by batch cultures of bloom-forming Cyanobacterium, Microcystis aeruginosa with different ethylenediaminetetraacetic acid (EDTA)-Fe concentrations (0.5–6.0 mg/L), three levels of initial biomass, and excessive N and P (N = 4.2 mg/L, P = 0.186 mg/L) to simulate dynamically a cyanobacteria bloom in eutrophic conditions. The effect of EDTA and Fe uptake kinetics by M. aeruginosa were also examined. Results showed M. aeruginosa growth rate positively correlated with EDTA-Fe concentration and negatively correlated with biomass. Maximal biomass of M. aeruginosa was determined by Fe availability and initial biomass. EDTA could decrease both Fe availability and toxicity. Based on experimental results, a conceptual model of how Fe availability regulates cyanobacterial biomass in eutrophic lakes was developed. This study demonstrated bioavailable Fe is a potential limiting factor in eutrophic lakes that should be included in eutrophication management strategies.
1. Introduction
The presence of cyanobacteria blooms in freshwater is one of the most serious ecological problems in the world [1]. It causes water discoloration, foul odors and tastes, deoxygenation of bottom waters (hypoxia and anoxia), toxicity, fish kills, and food web alterations. Toxins produced by blooms can adversely affect animal (including human) health in waters used for recreational and drinking purposes [2]. In China, more than half of the freshwater systems have experienced algal blooms, and Microcystis aeruginosa is one of the dominant species that causes such blooms [3]. Despite extensive research in the past five decades, effective management of eutrophic lakes remains constrained by lack of mechanistic understanding of causal factors for cyanobacteria bloom formation [4].
The knowledge of the causal factors for cyanobacteria blooms is incomplete. Admittedly, N and P are strong risk factors since total nitrogen (TN) and total phosphorus (TP) are positively correlated with the frequency and intensity of cyanobacteria blooms [5]. However, eutrophic conditions do not ensure cyanobacteria bloom [6]. The failure of P or N reduction strategies in many systems also indicates the unsatisfactory explanation only by N and/or P [7,8]. With increasing anthropogenic nutrient loading (cultural eutrophication), phytoplankton become P and N saturated, i.e., their growth rate and biomass are insensitive to changes in P or N concentration. The work by Auer verified that the eutrophic waters of southern Green Bay, western Lake Erie, and nearshore Lake Ontario are nutrient saturated and insensitive to P reduction [9]. Clearly, something other than P or N regulates phytoplankton biomass in eutrophic lakes.
Fe limitation of phytoplankton has aroused an increasing concern since Martin’s Fe hypothesis [10,11]. About 30% of the world’s ocean surface area is limited by Fe availability where subsurface nutrient supply is enhanced (high-nutrient low-chlorophyll (HNLC) regions), including main oceanic upwelling regions of the Southern Ocean and eastern equatorial Pacific Ocean [12]. Since Fe is a limiting factor in HNLC regions, it could also be the causal factor for cyanobacteria blooms in eutrophic lakes. They are two sides to this mechanism. Fe is an essential micronutrient for all phytoplankton and plays an important role in photosynthesis and respiration [13]. Fe-containing compounds are ubiquitous in cells, specifically in electron transfer reactions and in enzymes [14]. Particularly, cyanobacteria need much more Fe than eukaryotic algae due to their evolutionary origins in Fe-rich anoxic oceans [15,16]. N2-fixing cyanobacteria have a specific need for Fe in nitrogenase [17]. Fe additions have been found to increase growth rates, photosynthesis, and N fixation in cyanobacterial populations [18,19].
Fe is the second most abundant metal and fourth most abundant element in the Earth’s crust, but its concentration in water is quite low because of its low solubility [20]. Complexation with siderophores and other uncharacterized organic ligands increases Fe solubility but reduces its availability [21]. In eutrophic lakes, especially P-rich water, bioavailable Fe is always poor because of Fe-P mineral formation [22]. Present knowledge on the limitation of phytoplankton productivity by bioavailable Fe is mainly based on marine phytoplankton research [23,24]. The extent to which bioavailable Fe regulates cyanobacterial biomass in fresh water remains unclear. Recently, a few enclosure experiments showed Fe is a potential limiting nutrient for cyanobacterial growth in eutrophic lakes [25,26]. Although the physiological effects of Fe on cyanobacteria and cyanobacteria’s Fe uptake mechanisms have been well studied [27,28], the effect of Fe on cyanobacteria bloom formation is not clear. Clarifying this effect is very important for understanding the cyanobacteria bloom mechanism and developing effective eutrophication management strategies.
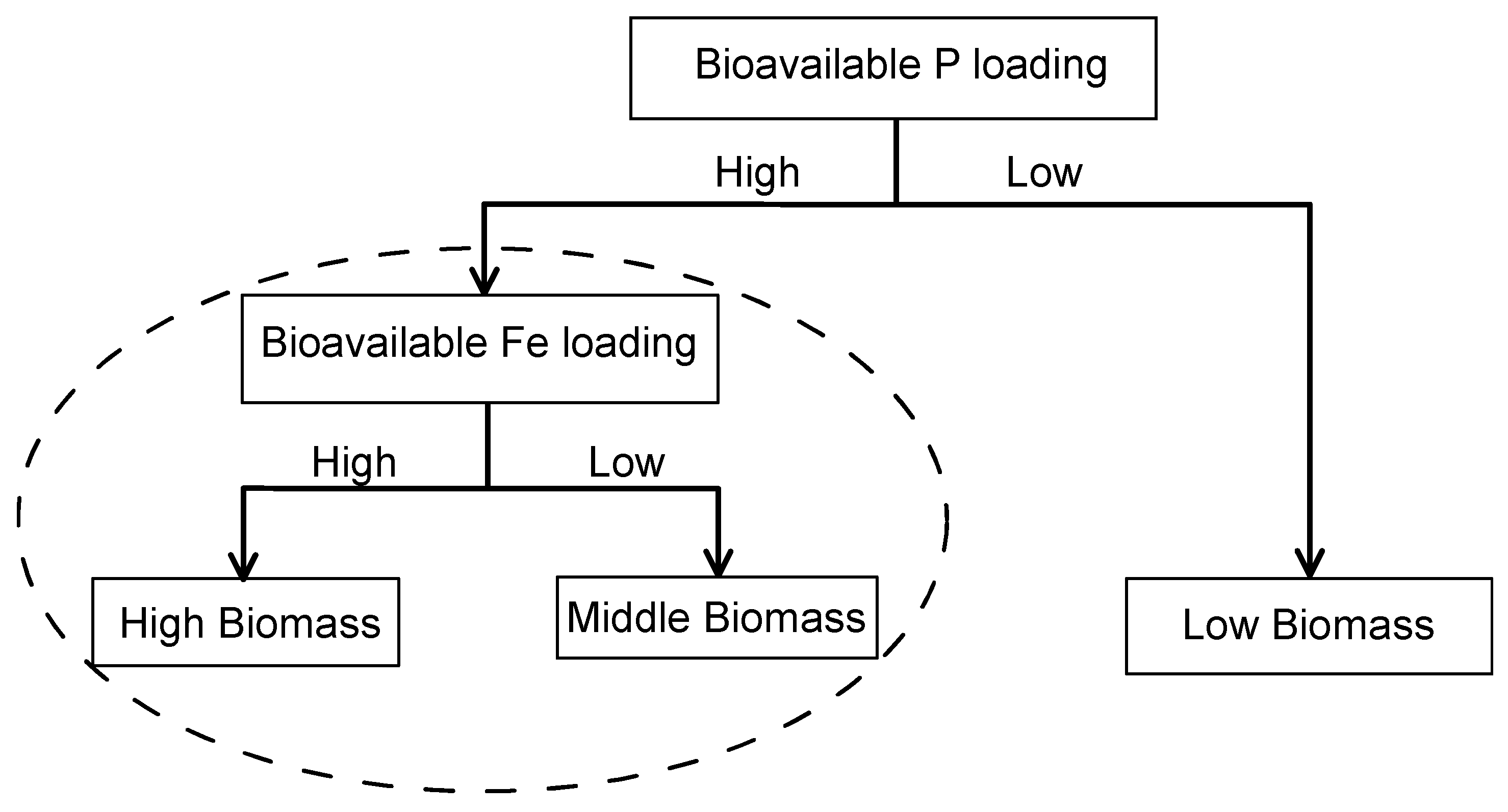
To probe the effects of bioavailable Fe on cyanobacteria bloom formation in eutrophic conditions, three bottle-scale experiments with bloom-forming cyanobacterium M. aeruginosa were conducted. In this paper, we admitted the importance of N or P. However, we hypothesized Fe is an important limiting factor for cyanobacteria growth in eutrophic lakes (excessive N and P) (Figure 1). This hypothesis follows Liebig's law of minimum, which suggests primary productivity is limited by the nutrient in shortest supply. The ultimate goal of this study is to develop a conceptual model of how Fe availability regulates cyanobacterial biomass in eutrophic lakes, elucidating qualitative relationships among Fe concentrations, growth rate, and cyanobacterial biomass dynamically based on our experimental results.
2. Materials and Methods
2.1. Cultures
The FACHB-905 strain of M. aeruginosa was isolated from eutrophic Lake Dianchi, China, in 1998 (bought from the Institute of Hydrobiology, Chinese Academy of Science, Wuhan, China). Stock cultures were maintained in 500 mL Erlenmeyer flasks containing 100 mL Algal Growth Potential (AGP) media [29]. According to the standard of trophic state classification of lakes in China [29] (Table S1), the AGP medium supplies nutrients in hyper-eutrophic status (N = 4.2 mg/L, P = 0.186 mg/L, C = 2.14 mg/L, Fe= 33 µg/L, Table S2). All flasks were closed with four-layer gauze and one-layer aluminum foil to prevent contamination. Cells were harvested for experiments during the exponential growth phase. Cultures were cultivated in Fe-free media for 72 h to lower the side effects of intracellular Fe. Prior to experiments, algae were concentrated by centrifugation (5 min, 3500 rpm) and washed three times with 15 ppm NaCO3 solution to reduce interference effects of extracellular nutrients. Strict trace metal cleaning techniques were applied in all culturing and experimental manipulations. All materials for stock cultures and experiments were cleaned by 10% HCl and sterilized by autoclaving before assembly and operation. Media were prepared with ultrapure (Milli-Q Advantage A10, Millipore, Billerica, MA, USA) and 99.99% metal basis chemicals (Aladdin, Shanghai, China). To provide maximal sterility, the procedures were conducted on a biological clean bench (BCM-1300A, Anda, Suzhou, China) and media were purified with high-pressure steam sterilization technology. Experiments were performed in 250 mL Erlenmeyer flasks with 150 mL designed AGP media. All flasks were placed in climate incubators (MGC-400B, Yiheng, Shanghai, China) at 25 °C and a constant light intensity in a 12 h:12 h light-dark cycle. During the experimental period, pH values were maintained at 7.0 0.1 by adding 0.1 mol/L KCl and 0.1 mol/L HCl solutions.
2.2. Growth Experiments
Fe3+, the common form in fresh water, was used in experiments. Experiment (Exp) 1 was based on a two-factor design with six Fe (EDTA-Fe) concentrations (0.5, 1.0, 1.5, 2.0, 4.0, and 6.0 mg/L) and three levels of initial biomass (low, OD665 = 0.079 0.002; middle, OD665 = 0.099 0.002; high, OD665 = 0.192 0.002) to test the effect of Fe concentrations and initial biomass on M. aeruginosa growth. Fe concentrations used here exceeded average Fe concentrations (0.5 mg/L) in eutrophic Lake Dianchi; this was done to test M. aeruginosa grow potential with higher Fe concentrations [30]. Each treatment was conducted in triplicate. Hence, a total of 54 flasks were used in Exp 1. Exp 1 was performed for 10 days.
Exp 2 was designed to test the effects of EDTA and Fe3+ availability on M. aeruginosa growth. Fe3+ (FeCl3) was added to reach the same target concentrations in Exp 1 (0.5, 1.0, 1.5, 2.0, 4.0, and 6.0 mg/L) with a constant EDTA background (Na2EDTA·2H2O, 3.0 mg/L). Initial biomass in Exp 2 was constant (OD665 = 0.135 0.002). P-free media with 0.5 mg/L Fe3+ (FeCl3) and a constant EDTA background (3.0 mg/L) were used as a negative control group. Each treatment was conducted in triplicate. Hence, a total of 21 flasks were used in Exp 2. Exp 2 was performed for 4 days.
2.3. Fe Uptake Tests
Exp 3 was designed to test Fe3+ uptake kinetics of M. aeruginosa in an exponential concentration gradient. Fe3+ (FeCl3) was added to reach target concentrations of 0.000, 0.112, 0.224, 0.448, 0.896, 1.792, 3.584, and 7.168 mg/L with a constant EDTA background (Na2EDTA·2H2O, 0.3 mg/L). Each treatment was conducted in triplicate. Hence, a total of 24 flasks were used in Exp 3. Exp 3 was performed for 12 days. Extracellular dissolved total Fe (DTFe) concentration remaining in the medium was measured every 2 h for 12 h and every 2 day for 10 days using a flame atomic absorption spectrometer (ContrAA 300, Analytic Jena, Jena, Germany). The samples were concentrated by centrifugation (5 min, 8000 rpm) and supernatant were used to test DTFe.
2.4. Measurement of Growth
Algal biomass was monitored using an optical density of 665 nm (OD665, UV2800SPC spectrophotometer; Sunny Hengping, Shanghai, China). A cell suspension with an optical density of 1.0 contained 4.2 × 107 7 × 105 cells per mL, as determined by viable counts.
2.5. Data Analysis
Growth rate was computed based on phytoplankton biomass
where μ (d−1) is the growth rate, and ti − t0 is the time interval during which phytoplankton biomass increases from initial biomass (N0) to Ni. Maximal growth rate during the experimental period was chosen as growth rate (d−1) for each experimental unit.
Change curves of growth rate with Fe concentrations were fitted with the Monod model:
where µ (d−1) is the growth rate, µmax (d−1) is the maximal growth rate at infinite Fe, Km (mg/L) is the half-saturation constant, and C (mg/L) is the initial Fe concentration added in medium. Growth affinities were defined as µmax/Km.
Fe uptake kinetic constants were obtained by fitting the Michaelis-Menten equation, which has the same form as the Monod Model:
where is the Fe uptake rate (mg/(L∙h)), is the maximal Fe uptake rate (mg/(L∙h)), Km (mg/L) is the half-saturation constant, and C (mg/L) is the initial Fe concentration added in medium. Fe uptake rate (mg/(L∙h)) in each treatment is defined as the average change rate of extracellular DTFe concentration (h−1) between 2 h and 4 h.
Statistical tests were performed using R, Version 3.2.3 (R Core Team 2015, Auckland, New Zealand). ANOVA with Tukey’s Honest Significant Difference (HSD) test was used in Exp 1. The differences were considered significant at P < 0.05. All graphs in this study were created with Origin 9.0.
3. Results
3.1. Exp 1: Effect of EDTA-Fe and Initial Biomass on M. aeruginosa Growth
In Exp 1, growth curves of M. aeruginosa for all treatments showed the same S-shaped patterns. Biomass and growth rate of M. aeruginosa in almost all treatments positively correlated with EDTA-Fe concentration (0.5–6.0 mg/L) (Figure 2A and Figure S1, Table S3). This result indicates great M. aeruginosa growth potential with 6.0 mg/L EDTA-Fe and probably even with higher Fe levels. Maximal biomass of M. aeruginosa positively correlated with initial biomass and Fe concentration (Figure 2B).
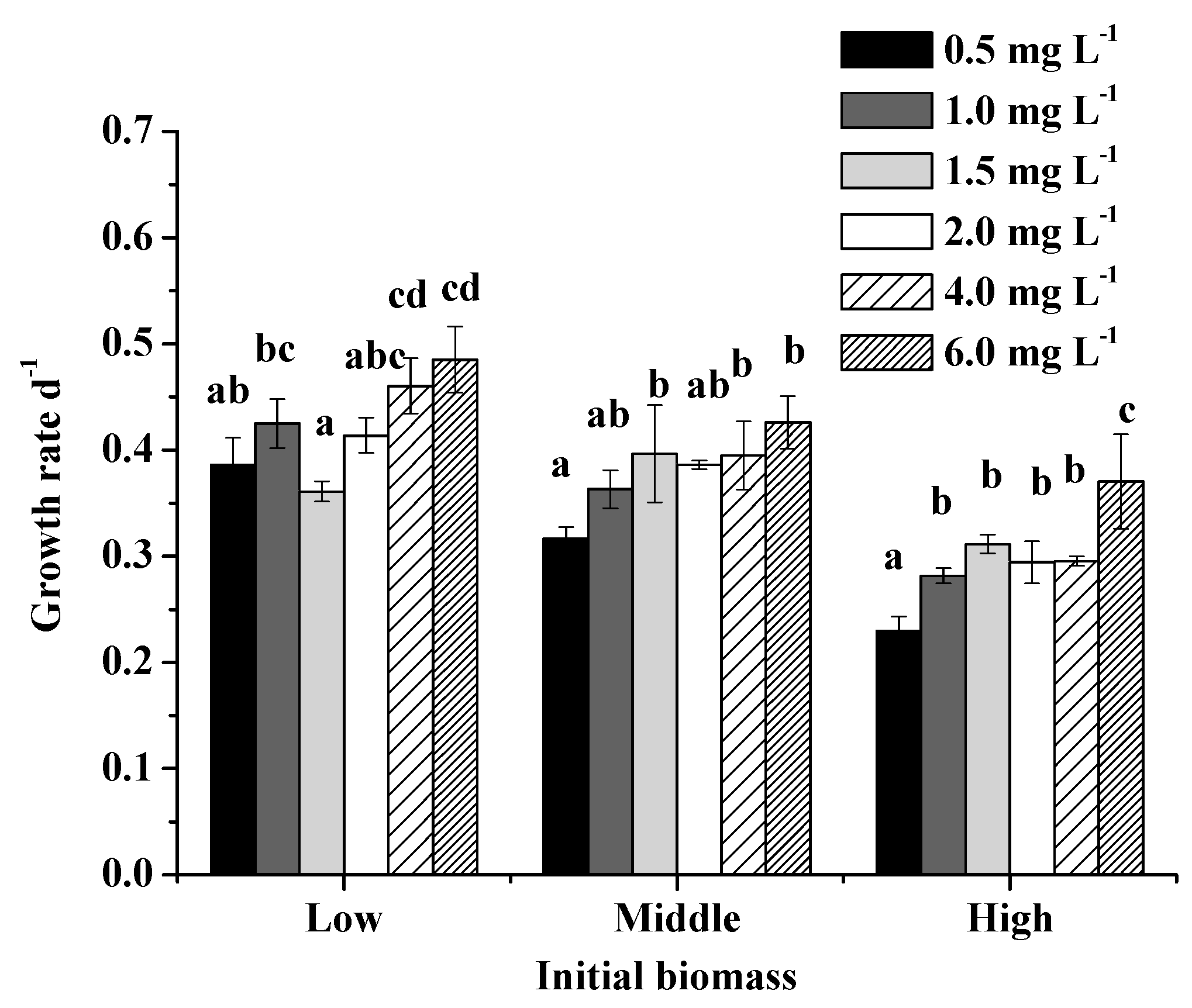
Global ANOVA test results of the effects of EDTA-Fe concentrations on M. aeruginosa growth rate were significant (P < 0.01) at all initial biomass levels. Subsequent pairwise tests showed significant differences between low and high concentrations (e.g., P < 0.01 for 0.5 and 6.0 mg/L, Figure 3, Table S3) and no significant differences within intermediate concentrations. M. aeruginosa growth rate at high initial biomass with 6.0 mg/L EDTA-Fe significantly continued to increase compared with 4.0 mg/L EDTA-Fe (P < 0.01). Global ANOVA test showed significant differences between different initial biomass treatments (P < 0.01). Significant negative effects of initial biomass on M. aeruginosa growth rate were found at all Fe levels (Figure S2, Table S4, P < 0.05).
Change curves of growth rate with Fe concentrations conformed to the Monod model. Maximal growth rate (µmax) of M. aeruginosa at infinite Fe negatively correlated with initial biomass (Table 1). The half-saturation constant for M. aeruginosa growth with respect to Fe positively correlated with initial biomass. A negative correlation was found between growth affinity (µmax/Km) of M. aeruginosa to EDTA-Fe and initial biomass (Table 1).
3.2. Exp 2: Effect of EDTA and Fe3+ on M. Aeruginosa Growth
Growth affinity (µmax/Km) of M. aeruginosa to Fe3+ was three times stronger than its affinity to EDTA-Fe, indicating Fe3+ was more available to M. aeruginosa than EDTA-Fe (Table 1). Growth curves with Fe3+ addition were incompatible with the S-shape, which showed a different pattern from EDTA-Fe treatments (Figure 2C). This could be because the running time of Exp 2 was much shorter (4 days) than that of Exp 1 (10 days). All algae died within 2 h after addition of the high Fe3+ concentration (6.0 mg/L), which was accompanied with precipitant. Small substances in the medium were observed under microscope and no large algal particles were found. Algae in P-free media were not grown and started to die slowly above 2 days. M. aeruginosa biomass decreased with increasing Fe3+ concentrations after 1.0 mg/L.
3.3. Exp 3: Effect of Fe3+ on M. Aeruginosa Growth and Fe3+ Uptake
The same growth pattern was observed in Exp 2 and Exp 3 (Figure 2C,D). At day 6, M. aeruginosa biomass increased with increasing Fe3+ concentrations within 0–0.896 mg/L and reached its highest at 0.896 mg/L. However, it decreased with increasing Fe3+ concentration within 0.896–7.168 mg/L. After day 6, M. aeruginosa biomass with an Fe3+ concentration of 0.896 mg/L started to decline whereas M. aeruginosa biomass with an Fe3+ concentration of 1.792 mg/L continued to grow until day 10. M. aeruginosa growth rate positively correlated with Fe concentration (0–3.584 mg/L) (Figure S3).
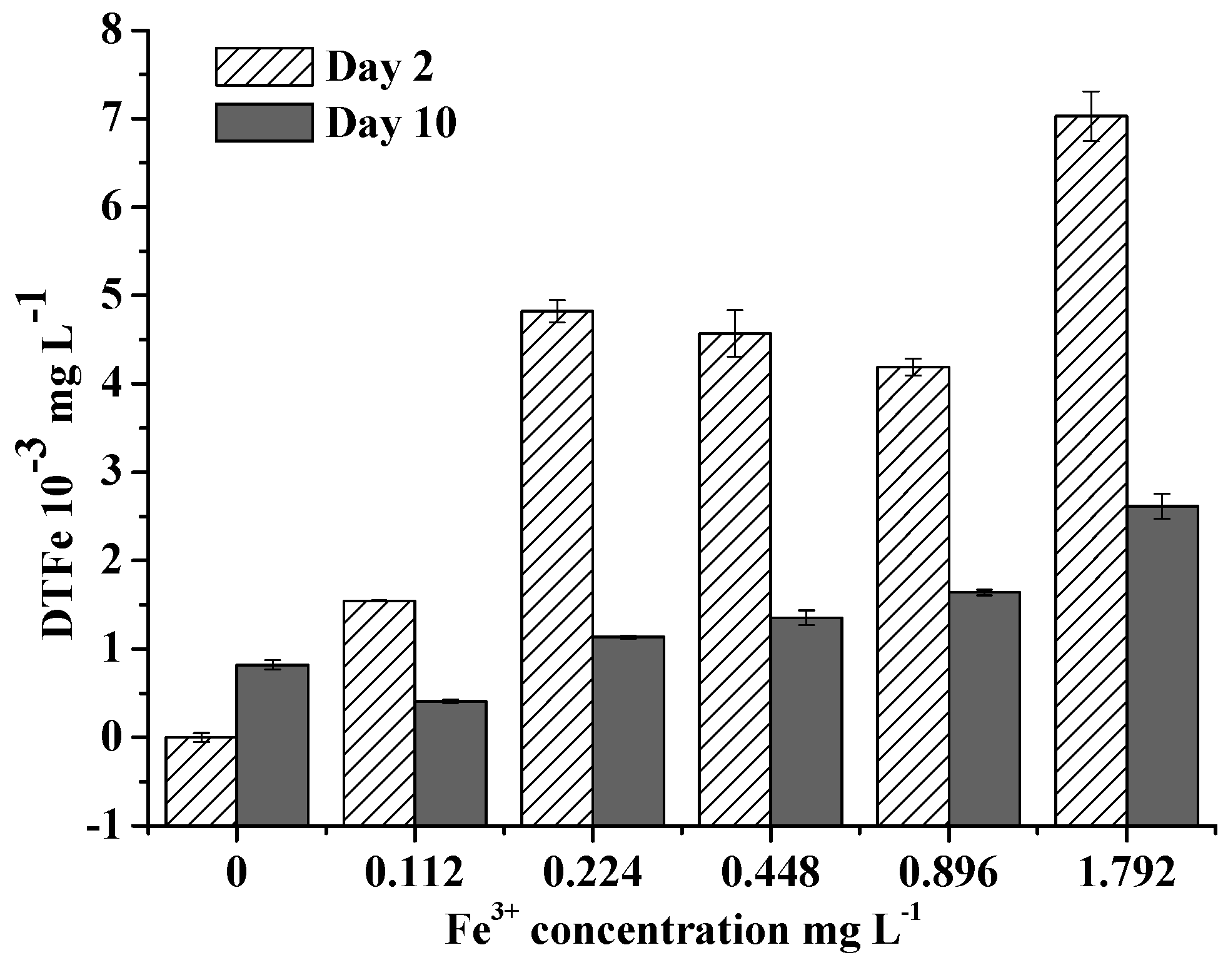
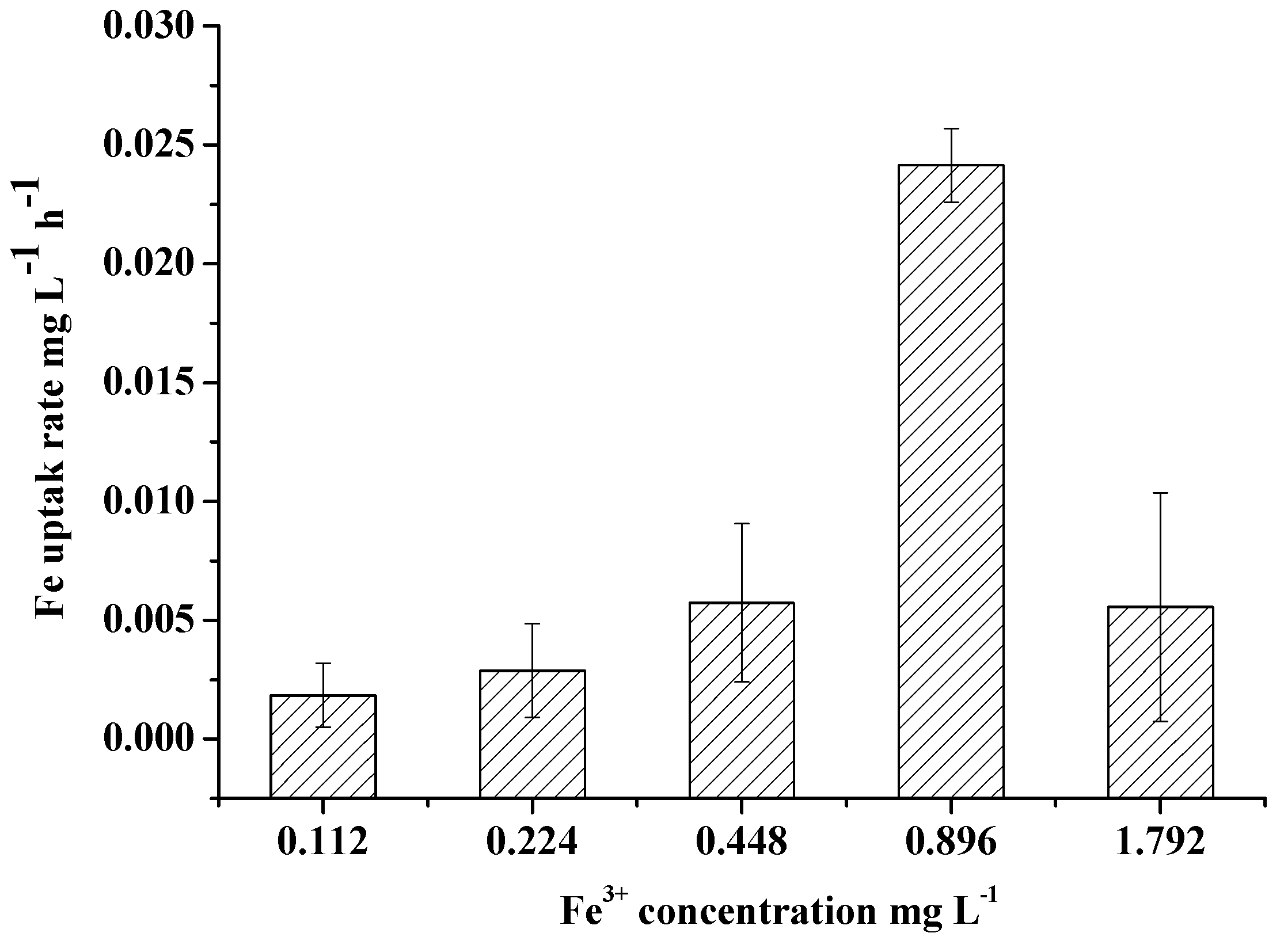
Extracellular DTFe concentration decreased over time, indicating Fe had been absorbed by algae. Extracellular DTFe concentration at day 10 was lower than that at day 2 (Figure 4). Fe uptake rate of M. aeruginosa positively correlated with Fe3+ concentration within 0–0.896 mg/L, suggesting M. aeruginosa has a good ability to absorb and utilize Fe3+ within this concentration range (Figure 5). This result also supported the hypothesis that M. aeruginosa growth is limited by Fe. At Fe3+ concentrations higher than 0.896 mg/L, the Fe uptake rate of M. aeruginosa was very low, suggesting Fe3+ with high concentrations could not be absorbed by algae. This result is consistent with findings in growth experiments. When Fe uptake rate was plotted as a function of Fe3+ concentration, the data merged and exhibited Michaelis–Menten kinetics with a Km of 3.26 0.01 mg/L and a maximal Fe uptake rate () of 0.053 mg/(L∙h).
4. Discussion
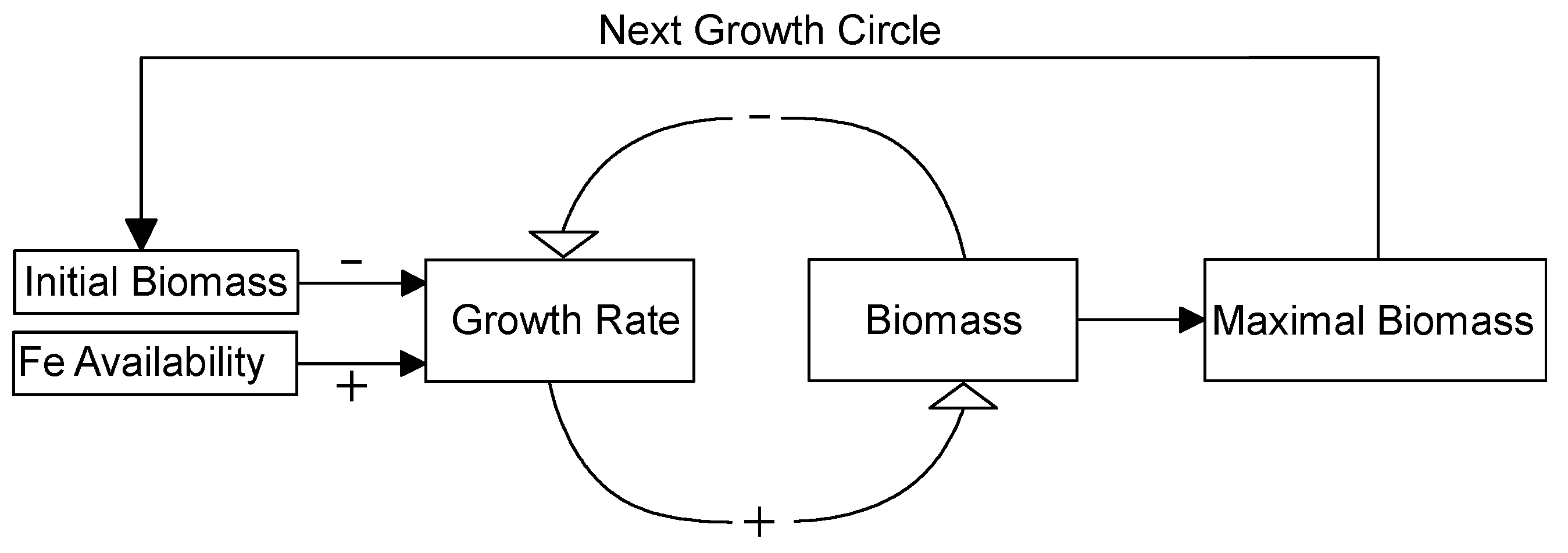
Our results revealed the positive effect of bioavailable Fe on cyanobacteria biomass and indicated Fe could be a potential limiting factor in eutrophic lakes. Main results from three experiments were that: (i) M. aeruginosa growth rate positively correlated with EDTA-Fe concentration and negatively correlated with biomass, (ii) maximal biomass of M. aeruginosa was determined by Fe availability and initial biomass, and (iii) EDTA could decrease both Fe availability and toxicity. Based on experimental results, a conceptual model of how Fe availability regulates cyanobacterial biomass in eutrophic lakes (Figure 6) was developed. Below, we talk about the conceptual model and main results, as well as discuss implications for eutrophication management.
4.1. A Conceptual Model of How Fe Availability Regulates Cyanobacterial Biomass in Eutrophic Lakes
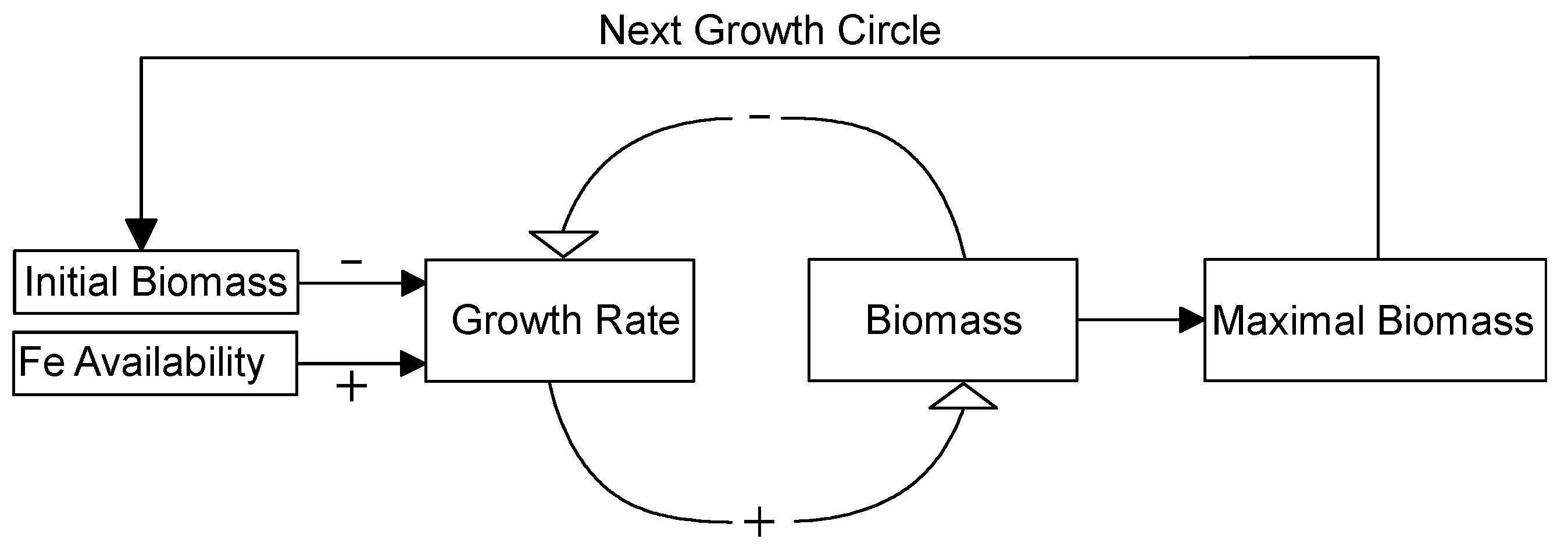
This conceptual model considers qualitative relationships among Fe concentration, growth rate, and phytoplankton biomass within one population growth cycle and over multiple population growth cycles. Within one population growth cycle, Fe availability exerts a positive effect on the population growth rate. Moreover, algal biomass, including initial biomass and biomass at any time, negatively affects population growth rate, which leads to a final balanced maximal biomass. For the next population growth cycle, the maximal biomass in last cycle serves as the initial biomass for this cycle and the process repeats. Therefore, Fe availability plays an important role in cyanobacterial population dynamics. This model could explain why cyanobacteria outbreaks occur over a short time, because cyanobacteria growth rate is relatively high when initial biomass is low. This model could also explain why cyanobacteria blooms are becoming much more serious year after year, because initial biomass increases for the next growth cycle.
4.2. Effect of Fe Concentration and Initial Biomass
Exp 1 showed M. aeruginosa growth rate positively correlated with EDTA-Fe concentration and negatively correlated with biomass. Maximal biomass of M. aeruginosa was determined by Fe availability and initial biomass. Results of Exp 1 indicated great M. aeruginosa grow potential with 6.0 mg/L EDTA-Fe and probably even with higher concentrations of Fe. Examination of Fe concentrations reported in water basins revealed Fe concentrations in freshwater is much less than 6 mg/L. For example, the soluble Fe concentration in Lake Dianchi (China) where the M. aeruginosa was isolated is estimated to be 0.3–1.0 mg/L [30]. According to Liebig’s law of minimum, M. aeruginosa could be limited by Fe in many eutrophic lakes. From the results, we can infer M. aeruginosa had a very high Fe requirement. The Fe threshold concentration in this result was much higher than 10−8–10−7 M for the growth of Anabaena circinalis (cyanobacterium) [31]. Two main reasons for this phenomenon are different Fe speciations and the testing concentration range used in the experiments. Sun et al used FeCl3 with a series of testing concentrations ranging from 2.2 × 10−5 M to 2.2 × 10−9 M, which is much lower than the concentration range in Exp 1. Results of Exp 2 with the same Fe form, FeCl3, were highly consistent with their reports. We infer the reason for this gap is related to Fe bioavailability and toxicity.
Results showed M. aeruginosa growth rate negatively correlated with biomass in a certain nutrient availability indicating Fe is a negative density-dependent regulator of cyanobacterial population growth. Whether or not nutrient availability is density-dependent is always a controversial topic in ecological literature [31,32,33,34,35,36]. Our experiments under controlled environments and replicated conditions suggested nutrient availability is density-dependent. Possible explanations for this phenomenon are rooted in intraspecific competition and constant nutrient demand of individual algal cells. On one hand, resource availability could cause density dependence by affecting intraspecific competition among individuals. An increased density means an increase in intraspecific competition [37]. On the other hand, individual nutrient demand is relatively constant. When the population reaches a certain density, the amount of available nutrients to each cell becomes insufficient and cannot support cell growth, which causes cell division reduction [38]. Density regulation has been observed in a broad range of taxa. The negative density-dependent (restriction) processes of Fe availability contribute to resilience of the cyanobacterial population.
4.3. Effect of EDTA and Fe3+
In Exp 2, all algae died within 2 h after addition of high Fe3+ concentration (6.0 mg/L). Several alternative explanations can be considered for this phenomenon. First, Fe3+ is very unstable and can be precipitated with P; thus, algae are attached and precipitated with an Fe-P complex. Second, algae died without P. Third, excess Fe3+ is toxic to cyanobacteria, leading to cyanobacteria autolysis. Results of the negative control group of P-free media excluded the second reason in that algae did not die without P and merely stopped growing. We observed small substances in the medium under microscope and found no large algal particles, which indicated cyanobacteria were autolyzed because of Fe3+ toxicity. This explanation is supported by the low Fe uptake rate of M. aeruginosa when Fe3+ concentration was higher than 0.896 mg/L in Exp 3. Therefore, high Fe3+ could be toxic to cyanobacteria and EDTA could reduce Fe toxicity.
Exp 2 showed Fe3+ is more available to M. aeruginosa than EDTA-Fe, which is consistent with most results in the literature [39]. The reason is that only ionic Fe (Fe3+ and Fe2+) can be absorbed by cyanobacteria. Fe3+ can be transported across cell membranes mainly through two strategies: strategy I (Fe reduction at cell membrane followed by Fe2+ transport) and strategy II (siderophore-based Fe3+ acquisition) [40]. In strategy II, siderophores are organic Fe3+ or other metal-chelating molecules that solubilize and scavenge Fe3+ from the ambient environment. Most freshwater phytoplankton, especially cyanobacteria, can secrete siderophores under an Fe-limited environment [41,42,43]. Fe3+ is not stable in fresh water despite its availability, which is higher than that of EDTA-Fe. EDTA as organic ligands could enhance Fe solubility and decrease both Fe availability and toxicity. EDTA-Fe is the normal form for Fe fertilization [44]. Cyanobacteria can release siderophore to compete and utilize Fe3+ from EDTA-Fe [45]. Although EDTA could decrease the Fe3+ uptake rate of M. aeruginosa, additions of EDTA-Fe could increase bioavailable Fe concentration and boost plankton growth.
In natural water, dissolved organic matters (DOMs) possess chelation properties such as EDTA that enhance phytoplankton growth [46]. For example, humic substances, which are common in water environments, can form complexes with metals and alter their availability. Presence of humic substances stimulates photosynthesis in the green alga Pseudokirchneriella subcapitata by reducing toxicity of heavy metals [47]. Humic substances also inhibit M. aeruginosa growth by reducing bioavailable Fe concentration [48]. Fe concentration negatively correlates with P concentration in fresh water because Fe can precipitate with P [49]. Therefore, the outbreak of cyanobacteria often occurs in polymictic and high-organic-matter lakes because of sufficient Fe and P [50]. In addition, soluble bioavailable Fe can easily be released from endogenous compounds in an anoxic environment if H2S concentration is low [51]. Cyanobacteria blooms are always accompanied with anoxic environments, leading to increased bioavailable Fe and increased cyanobacterial biomass [52] and showing a vicious cycle.
4.4. Implications
This study demonstrated bioavailable Fe is an important limiting factor that regulates cyanobacterial population growth in eutrophic lakes. We do not deny the importance of N and P. However, we hope our work will draw other investigators’ attention to the effects of Fe on cyanobacteria blooms. This work has significant potential to improve eutrophication management. An Fe-reduction management strategy along with a macronutrient-reduction (N and P) strategy should be considered to control cyanobacteria blooms. Over the last 50 years, Fe salts have been applied to 16 lakes and reservoirs to control harmful algal blooms by inhibiting P release [40]. While this approach has been successful in the short term, it did not remove internal sources of nutrients, but increased Fe concentrations in the lake. This may increase the magnitude of cyanobacteria blooms after Fe and P recycling in the long term. This work suggests that cyanobacterial growth in eutrophic lakes can be suppressed by controlling Fe availability. Fluvial inputs, internal recycling, and seepage of Fe-rich groundwater are main sources of Fe to lakes [53]. The growing disposal of sulfur wastes or changes in riverine discharges and groundwater supply due to climate change would affect the bioavailable Fe pool in fresh water [54]. The pathway by which anthropogenic activities over the last century and climate change impact natural Fe cycling in lakes is still unknown. Additional theoretical and practical studies about Fe availability and its effect on cyanobacteria are urgently needed.
Supplementary Materials
Supplementary materials are available online at www.mdpi.com/2073-4441/9/5/326/s1.
Acknowledgments
This study was funded by National Major Science and Technology Program of China for Water Pollution Control and Treatment (Grant No. 2012ZX07104-004).
Author Contributions
Ting Zhang and Xingzhang Luo designed and carried out experiments and prepared the manuscript. Jian He partially contributed to the experiment and analysis of the data.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Heisler, J.; Glibert, P.M.; Burkholder, J.M.; Anderson, D.M.; Cochlan, W.; Dennison, W.C.; Lewitus, A. Eutrophication and harmful algal blooms: A scientific consensus. Harmful Algae 2008, 8, 3–13. [Google Scholar] [CrossRef]
- Paerl, H.W.; Fulton, R.S.; Moisander, P.H.; Dyble, J. Harmful freshwater algal blooms, with an emphasis on cyanobacteria. Sci. World J. 2001, 1, 76–113. [Google Scholar] [CrossRef] [PubMed]
- Pan, G.; Zhang, M.M.; Chen, H. Removal of cyanobacterial blooms in Taihu Lake using local soils. I. Equilibrium and kinetic screening on the flocculation of Microcystis aeruginosa using commercially available clays and minerals. Environ. Pollut. 2006, 141, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Smith, V.H.; Schindler, D.W. Eutrophication science: Where do we go from here? Trends Ecol. Evol. 2009, 24, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Lu, X.; Chen, Y. The effects of temperature and nutrient ratios on Microcystis blooms in Lake Taihu, China: An 11-year investigation. Harmful Algae 2011, 10, 337–343. [Google Scholar] [CrossRef]
- Schindler, D.W. The dilemma of controlling cultural eutrophication of lakes. Proc. R. Soc. Lond. B Biol. Sci. 2012. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Xu, H.; McCarthy, M.J.; Zhu, G.; Qin, B.; Li, Y.; Gardner, W.S. Controlling harmful cyanobacterial blooms in a hyper-eutrophic lake (Lake Taihu, China): The need for a dual nutrient (N & P) management strategy. Water Res. 2011, 45, 1973–1983. [Google Scholar] [PubMed]
- Conley, D.J.; Paerl, H.W.; Howarth, R.W.; Boesch, D.F.; Seitzinger, S.P.; Havens, K.E.; Likens, G.E. Controlling eutrophication: Nitrogen and phosphorus. Science 2009, 323, 1014–1015. [Google Scholar] [CrossRef] [PubMed]
- Auer, M.T.; Kieser, M.S.; Canale, R.P. Identification of critical nutrient levels through field verification of models for phosphorus and phytoplankton growth. Can. J. Fish. Aquat. Sci. 1986, 43, 379–388. [Google Scholar] [CrossRef]
- Chappell, P.D.; Whitney, L.P.; Wallace, J.R.; Darer, A.I.; Jean-Charles, S.; Jenkins, B.D. Genetic indicators of iron limitation in wild populations of Thalassiosira oceanica from the northeast Pacific Ocean. ISME J. 2015, 9, 592–602. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Legendre, L.; Jiao, N. Phytoplankton responses to nitrogen and iron limitation in the tropical and subtropical Pacific Ocean. J. Plankton Res. 2015. [Google Scholar] [CrossRef]
- Moore, C.M.; Mills, M.M.; Arrigo, K.R.; Berman-Frank, I.; Bopp, L.; Boyd, P.W.; Jickells, T.D. Processes and patterns of oceanic nutrient limitation. Nat. Geosci. 2013, 6, 701–710. [Google Scholar] [CrossRef]
- Flynn, K.J.; Hipkin, C.R. Interactions between iron, light, ammonium, and nitrate: Insights from the construction of a dynamic model of algal physiology. J. Phycol. 1999, 35, 1171–1190. [Google Scholar] [CrossRef]
- Briat, J.F.; Dubos, C.; Gaymard, F. Iron nutrition, biomass production, and plant product quality. Trends Plant Sci. 2015, 20, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Sunda, W.G.; Huntsman, S.A. High iron requirement for growth, photosynthesis, and low-light acclimation in the coastal cyanobacterium Synechococcus bacillaris. Front. Microbiol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Sorichetti, R.J.; Creed, I.F.; Trick, C.G. Iron and iron-binding ligands as cofactors that limit cyanobacterial biomass across a lake trophic gradient. Freshw. Biol. 2016, 61, 146–157. [Google Scholar] [CrossRef]
- Garcia, N.S.; Fu, F.; Sedwick, P.N. Iron deficiency increases growth and nitrogen-fixation rates of phosphorus-deficient marine cyanobacteria. ISME J. 2015, 9, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Boyd, P.W.; Abraham, E.R. Iron-mediated changes in phytoplankton photosynthetic competence during SOIREE. Deep Sea Res. Part II Top. Stud. Oceanogr. 2001, 48, 2529–2550. [Google Scholar] [CrossRef]
- Shi, D.; Kranz, S.A.; Kim, J.M. Ocean acidification slows nitrogen fixation and growth in the dominant diazotroph Trichodesmium under low-iron conditions. Proc. Natl. Acad. Sci. USA 2012, 109, E3094–E3100. [Google Scholar] [CrossRef] [PubMed]
- Lewin, R. How microgranisms transport iron. Science 1984, 225, 401–402. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Wang, W.X.; Guo, L. Phase partitioning and solubility of iron in natural seawater controlling by dissolved organic matter. Glob. Biogeochem. Cycles 2004. [Google Scholar] [CrossRef]
- Orihel, D.M.; Schindler, D.W.; Ballard, N.C.; Wilson, L.R.; Vinebrooke, R.D. Experimental iron amendment suppresses toxic cyanobacteria in a hypereutrophic lake. Ecol. Appl. 2016. [Google Scholar] [CrossRef] [PubMed]
- Greene, R.M.; Geider, R.J.; Kolber, Z.; Falkowski, P.G. Iron-induced changes in light harvesting and photochemical energy conversion processes in eukaryotic marine algae. Plant Physiol. 1992, 100, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Sunda, W.G.; Huntsman, S.A. Interrelated influence of iron, light and cell size on marine phytoplankton growth. Nature 1997, 390, 389–392. [Google Scholar] [CrossRef]
- Hyenstrand, P.; Rydin, E.; Gunnerhed, M.; Linder, J.; Blomqvist, P. Response of the cyanobacterium Gloeotrichia echinulata to iron and boron additions—An experiment from Lake Erken. Freshw. Biol. 2001, 46, 735–741. [Google Scholar] [CrossRef]
- Molot, L.A.; Li, G.; Findlay, D. L; Watson, S.B. Iron-mediated suppression of bloom-forming cyanobacteria by oxine in a eutrophic lake. Freshw. Biol. 2010, 55, 1102–1117. [Google Scholar] [CrossRef]
- Sorichetti, R.J.; Creed, I.F.; Trick, C.G. The influence of iron, siderophores and refractory DOM on cyanobacterial biomass in oligotrophic lakes. Freshw. Biol. 2014, 59, 1423–1436. [Google Scholar] [CrossRef]
- Lis, H.; Kranzler, C.; Keren, N. A comparative study of iron uptake rates and mechanisms amongst marine and fresh water cyanobacteria: Prevalence of reductive iron uptake. Life 2015, 5, 841–860. [Google Scholar] [CrossRef] [PubMed]
- Jinhua, S. The Assessment Methodology for Eutrophication Level of Lakes in China. Environ. Pollut. Control 1990, 12, 2–7. [Google Scholar]
- Xing, W.; Huang, W.M.; Liu, Y.D.; Li, D.H.; Shen, Y.W.; Li, G.B. Environmental mechanism of change in cyanobacterial species composition in the Northeastern part of Lake Dianchi (China). Fresenius Environ. Bull. 2007, 16, 82–90. [Google Scholar]
- Sun, B.K.; Tanji, Y.; Unno, H. Influences of iron and humic acid on the growth of the cyanobacterium Anabaena circinalis. Biochem. Eng. J. 2005, 24, 195–201. [Google Scholar] [CrossRef]
- Clark, F.; Brook, B.W.; Delean, S.; Reşit Akçakaya, H.; Bradshaw, C.J. The theta-logistic is unreliable for modelling most census data. Methods Ecol. Evol. 2010, 1, 253–262. [Google Scholar] [CrossRef]
- Reynolds, S.A.; Brassil, C.E. When can a single-species, density-dependent model capture the dynamics of a consumer-resource system? J. Theor. Biol. 2013, 339, 70–83. [Google Scholar] [CrossRef] [PubMed]
- Sæther, B.E.; Engen, S. Pattern of variation in avian population growth rates. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2002, 357, 1185–1195. [Google Scholar]
- Bull, J.C.; Bonsall, M.B. Overcompensatory population dynamic responses to environmental stochasticity. J. Anim. Ecol. 2008, 77, 1296–1305. [Google Scholar] [CrossRef] [PubMed]
- Murray, D.L.; Anderson, M.G.; Steury, T.D. Temporal shift in density dependence among North American breeding duck populations. Ecology 2010, 91, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Hixon, M.A.; Johnson, D.W. Density dependence and independence. Encycl. Life Sci. 2009. [Google Scholar] [CrossRef]
- Borlestean, A.; Frost, P.C.; Murray, D.L. A mechanistic analysis of density dependence in algal population dynamics. Front. Ecol. Evol. 2015, 3, 37. [Google Scholar] [CrossRef]
- Hassler, C.S.; Twiss, M.R. Bioavailability of iron sensed by a phytoplanktonic Fe-bioreporter. Environ. Sci. Technol. 2006, 40, 2544–2551. [Google Scholar] [CrossRef] [PubMed]
- Xing, W.; Liu, G. Iron biogeochemistry and its environmental impacts in freshwater lakes. Fresenius Environ. Bull. 2011, 20, 1339–1345. [Google Scholar]
- Wilhelm, S.W. Ecology of iron-limited cyanobacteria: A review of physiological responses and implications for aquatic systems. Aquat. Microb. Ecol. 1995, 9, 295–303. [Google Scholar] [CrossRef]
- Ferreira, F.; Straus, N.A. Iron deprivation in cyanobacteria. J. Appl. Phycol. 1994, 6, 199–210. [Google Scholar] [CrossRef]
- Gress, C.D.; Treble, R.G.; Matz, C.J.; Weger, H.G. Biologicalavailability of iron to the freshwater cyanobacterium Anabaena flos-aquae1. J. Phycol. 2004, 40, 879–886. [Google Scholar] [CrossRef]
- Chen, Y.; Barak, P. Iron nutrition of plants in calcareous soils. Adv. Agron. 1982, 35, 217–240. [Google Scholar]
- Wilhelm, S.W.; Trick, C.G. Iron-limited growth of cyanobacteria: Multiple siderophore production is a common response. Limnol. Oceanogr. 1994, 39, 1979–1984. [Google Scholar] [CrossRef]
- Church, M.J.; Hutchins, D.A.; Ducklow, H.W. Limitation of bacterial growth by dissolved organic matter and iron in the Southern Ocean. Appl. Environ. Microbiol. 2000, 66, 455–466. [Google Scholar] [CrossRef] [PubMed]
- Koukal, B.; Gueguen, C.; Pardos, M.; Dominik, J. Influence of humic substances on the toxic effects of cadmium and zinc to the green alga Pseudokirchneriella subcapitata. Chemosphere 2003, 53, 953–961. [Google Scholar] [CrossRef]
- Imai, A.; Fukushima, T.; Matsushige, K. Effects of iron limitation and aquatic humic substances on the growth of Microcystis aeruginosa. Can. J. Fish. Aquat. Sci. 1999, 56, 1929–1937. [Google Scholar] [CrossRef]
- Elbaz-Poulichet, F.; Dupuy, C.; Cruzado, A.; Velasquez, Z.; Achterberg, E.P.; Braungardt, C.B. Influence of sorption processes by iron oxides and algae fixation on arsenic and phosphate cycle in an acidic estuary (Tinto river, Spain). Water Res. 2000, 34, 3222–3230. [Google Scholar] [CrossRef]
- Wang, W.X.; Dei, R.C. Bioavailability of iron complexed with organic colloids to the cyanobacteria Synechococcus and Trichodesmium. Aquat. Microb. Ecol. 2003, 33, 247–259. [Google Scholar] [CrossRef]
- Carey, E.; Taillefert, M. The role of soluble Fe (III) in the cycling of iron and sulfur in coastal marine sediments. Limnol. Oceanogr. 2005, 50, 1129. [Google Scholar] [CrossRef]
- Martynova, M.V. Iron compound occurrence forms in freshwater deposits: Analytical review. Water Res. 2010, 37, 488–496. [Google Scholar] [CrossRef]
- Nürnberg, G.K.; Dillon, P.J. Iron budgets in temperate lakes. Can. J. Fish. Aquat. Sci. 1993, 50, 1728–1737. [Google Scholar]
- Schindler, D.W.; Donahue, W.F. An impending water crisis in Canada's western prairie provinces. Proc. Natl. Acad. Sci. USA 2006, 103, 7210–7216. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Scheme for cyanobacteria bloom formation including both Fe and P factors. We admitted the importance of P. However, we hypothesized, in eutrophic lakes (excessive N and P), the limiting factor is bioavailable Fe. Both P and Fe control cyanobacterial biomass.
Figure 1.
Scheme for cyanobacteria bloom formation including both Fe and P factors. We admitted the importance of P. However, we hypothesized, in eutrophic lakes (excessive N and P), the limiting factor is bioavailable Fe. Both P and Fe control cyanobacterial biomass.
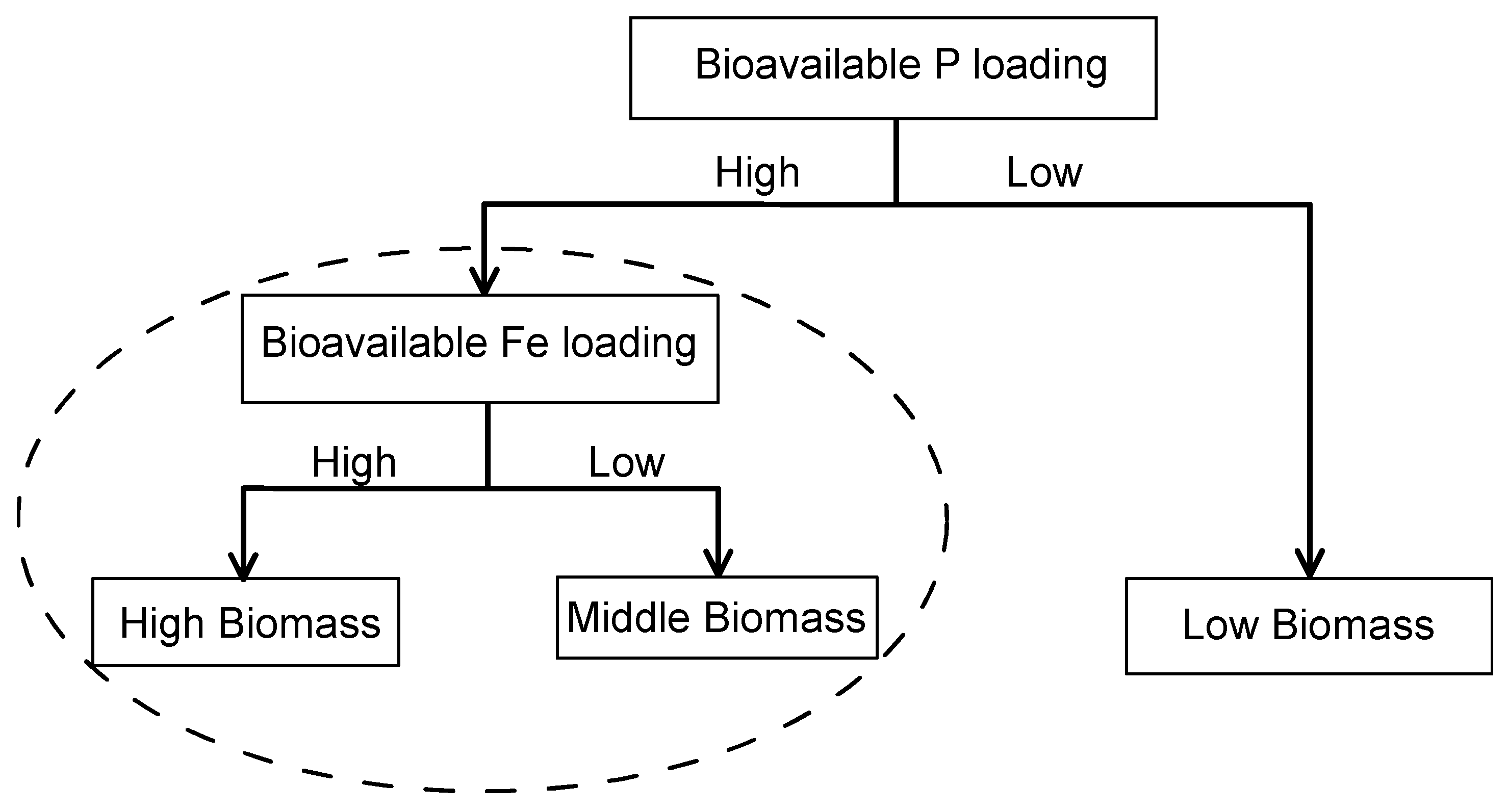
Figure 2.
Growth curves of M. aeruginosa with different concentrations of EDTA-Fe with middle initial biomass (A). Selected curves (B) are shown for the following initial biomass and EDTA-Fe concentrations: low initial biomass with 0.5 mg/L; low initial biomass with 6.0 mg/L; high initial biomass with 0.5 mg/L; high initial biomass with 6.0 mg/L. Growth curves of M. aeruginosa with different Fe3+ concentrations (0.5–6.0 mg/L) and 3.0 mg/L EDTA (C). P-free media with 0.5 mg/L Fe3+ (FeCl3) and a constant EDTA background (3.0 mg/L) were used as negative control group. Growth curves of M. aeruginosa with different Fe3+ concentrations (0–7.168 mg/L) and 0.3 mg/L EDTA (D). Error bars show standard deviation ().
Figure 2.
Growth curves of M. aeruginosa with different concentrations of EDTA-Fe with middle initial biomass (A). Selected curves (B) are shown for the following initial biomass and EDTA-Fe concentrations: low initial biomass with 0.5 mg/L; low initial biomass with 6.0 mg/L; high initial biomass with 0.5 mg/L; high initial biomass with 6.0 mg/L. Growth curves of M. aeruginosa with different Fe3+ concentrations (0.5–6.0 mg/L) and 3.0 mg/L EDTA (C). P-free media with 0.5 mg/L Fe3+ (FeCl3) and a constant EDTA background (3.0 mg/L) were used as negative control group. Growth curves of M. aeruginosa with different Fe3+ concentrations (0–7.168 mg/L) and 0.3 mg/L EDTA (D). Error bars show standard deviation ().
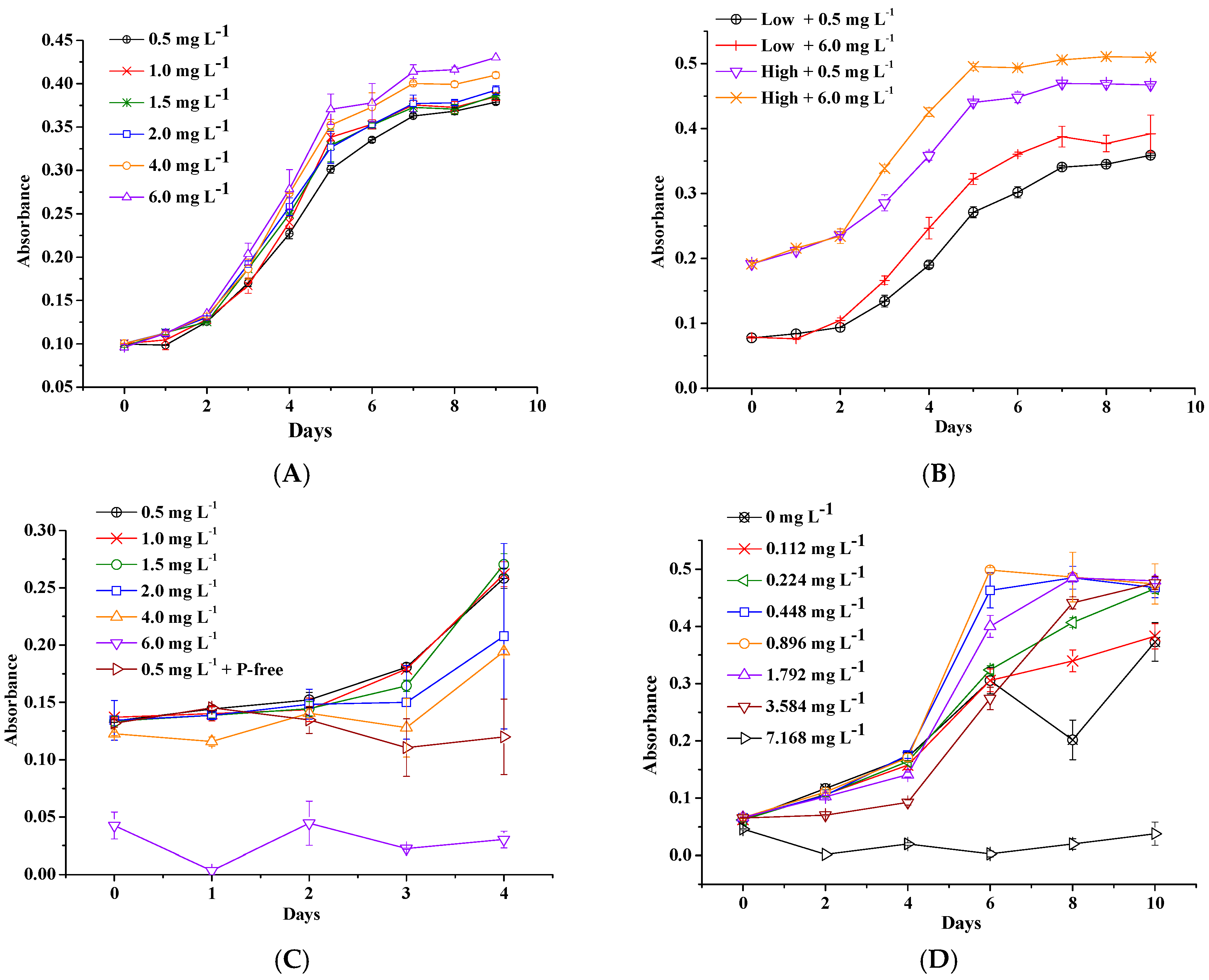
Figure 3.
Effect of EDTA-Fe concentrations on growth rate with low, middle, and high initial biomass. Small letters (a, b, c, d) indicate significantly different effects of EDTA-Fe concentrations on growth rate (P < 0.05, Tukey, HSD). The global effects of concentrations are significant for all three initial biomasses (P < 0.05). Error bars show standard deviation ().
Figure 3.
Effect of EDTA-Fe concentrations on growth rate with low, middle, and high initial biomass. Small letters (a, b, c, d) indicate significantly different effects of EDTA-Fe concentrations on growth rate (P < 0.05, Tukey, HSD). The global effects of concentrations are significant for all three initial biomasses (P < 0.05). Error bars show standard deviation ().
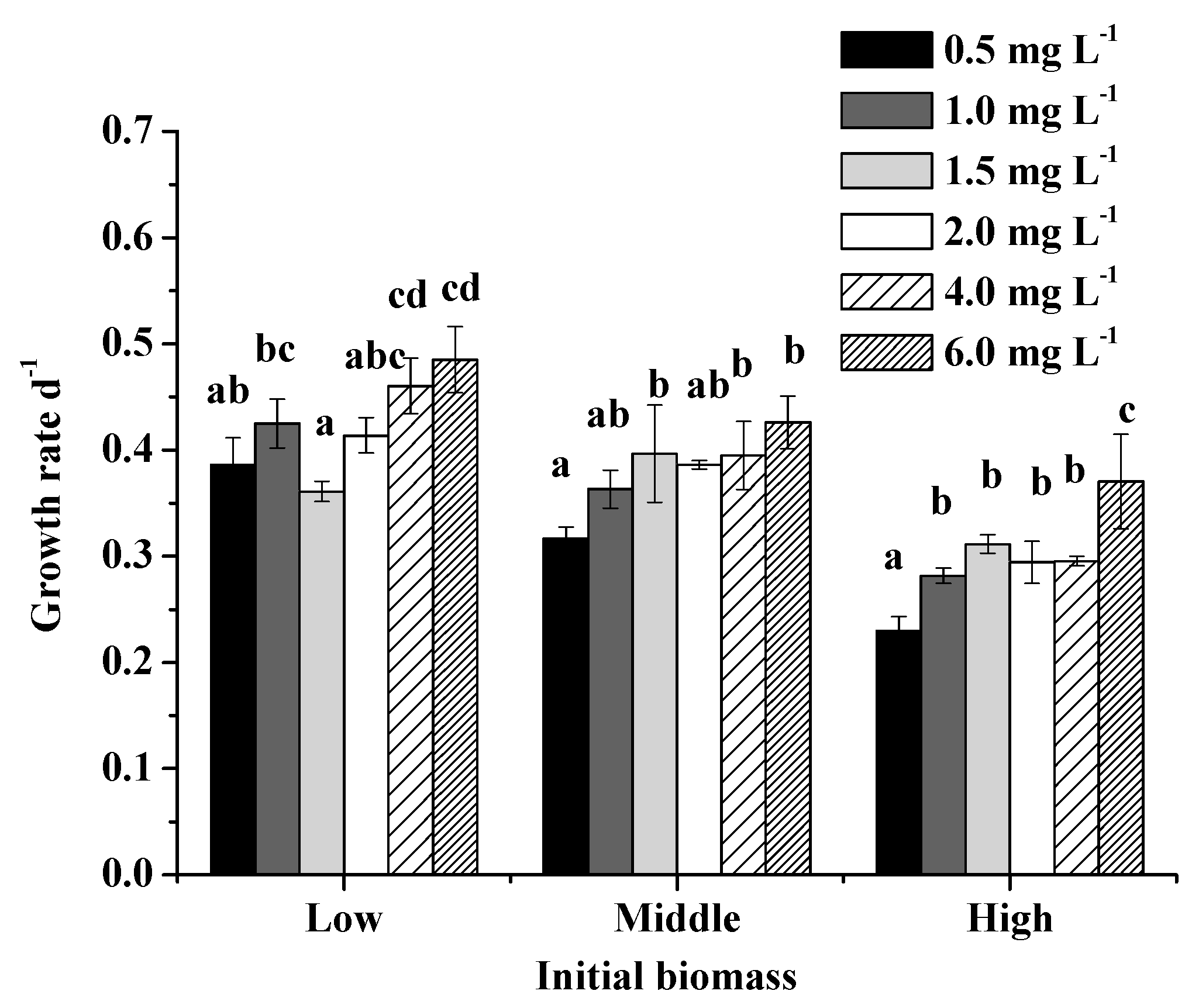
Figure 4.
Extracellular dissolved total Fe (DTFe) concentrations at day 2 and day 10 in different Fe3+ concentration treatments in Exp 3. Error bars show standard deviation ().
Figure 4.
Extracellular dissolved total Fe (DTFe) concentrations at day 2 and day 10 in different Fe3+ concentration treatments in Exp 3. Error bars show standard deviation ().
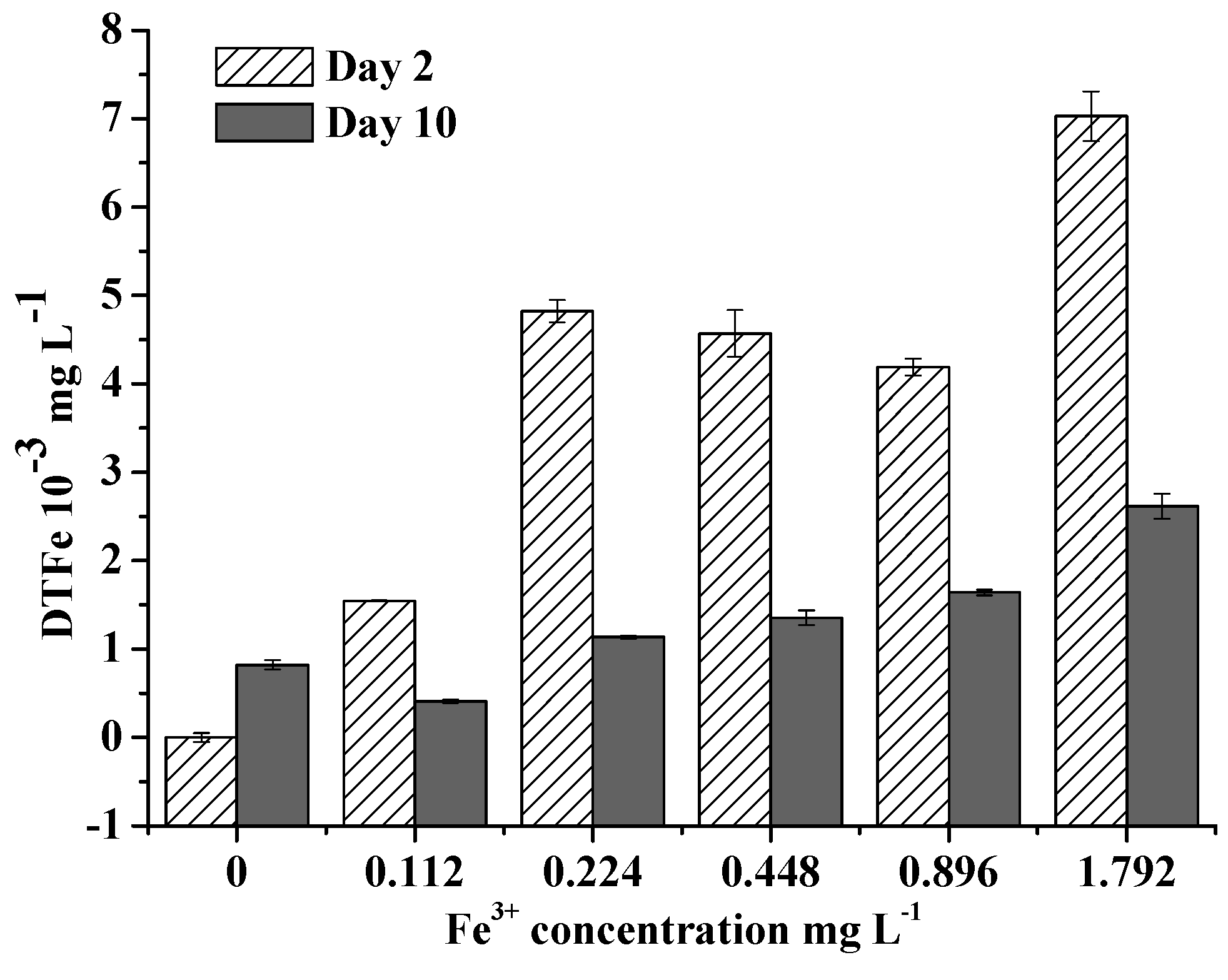
Figure 5.
Effects of Fe3+ concentration on the Fe uptake rate of M. aeruginosa. Error bars show standard deviation ().
Figure 5.
Effects of Fe3+ concentration on the Fe uptake rate of M. aeruginosa. Error bars show standard deviation ().

Figure 6.
A conceptual model of how Fe availability regulates cyanobacterial biomass in eutrophic lakes. “+” indicates a positive effect and “−” indicates a negative effect. Details can be found in the Discussion section.
Figure 6.
A conceptual model of how Fe availability regulates cyanobacterial biomass in eutrophic lakes. “+” indicates a positive effect and “−” indicates a negative effect. Details can be found in the Discussion section.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Monod Kinetic parameters calculated in Origin 9.0. Change curves of growth rate with Fe concentrations were fitted with the Monod model. Km: half-saturation constant for M. aeruginosa growth with respect to the Fe form. µmax: maximal growth rate with respect to the Fe form.
Table 1.
Monod Kinetic parameters calculated in Origin 9.0. Change curves of growth rate with Fe concentrations were fitted with the Monod model. Km: half-saturation constant for M. aeruginosa growth with respect to the Fe form. µmax: maximal growth rate with respect to the Fe form.
| Parameters | µmax | Km | µmax/Km | Adj. R-Square |
|---|---|---|---|---|
| Fe3+ | 0.5628 | 1.7040 | 0.330 | 0.8668 |
| EDTA-Fe | - | - | - | - |
| Low initial biomass | 0.4609 | 2.9981 | 0.154 | 0.5278 |
| Middle initial biomass | 0.4203 | 3.0543 | 0.138 | 0.8666 |
| High initial biomass | 0.3528 | 4.9090 | 0.072 | 0.6830 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhang, T.; He, J.; Luo, X. Effect of Fe and EDTA on Freshwater Cyanobacteria Bloom Formation. Water 2017, 9, 326. https://doi.org/10.3390/w9050326
AMA Style
Zhang T, He J, Luo X. Effect of Fe and EDTA on Freshwater Cyanobacteria Bloom Formation. Water. 2017; 9(5):326. https://doi.org/10.3390/w9050326
Chicago/Turabian StyleZhang, Ting, Jian He, and Xingzhang Luo. 2017. "Effect of Fe and EDTA on Freshwater Cyanobacteria Bloom Formation" Water 9, no. 5: 326. https://doi.org/10.3390/w9050326
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.