Inactivation Effect of Antibiotic-Resistant Gene Using Chlorine Disinfection

1
Department of Health Science, Kitasato University, 1-15-1 Kitasato, Sagamihara-Minami, Kanagawa 252-0373, Japan
2
Environmental Systems Course, Graduate School of Engineering, University of Miyazaki, 1-1 Gakuen Kibanadai-Nishi, Miyazaki 889-2192, Japan
3
Department of Electrical and Electronic Engineering, National Institute of Technology, Oita College, 1666 Maki, Oita 870-0152, Japan
*
Author to whom correspondence should be addressed.
Water 2017, 9(7), 547; https://doi.org/10.3390/w9070547
Submission received: 19 June 2017
/
Revised: 14 July 2017
/
Accepted: 17 July 2017
/
Published: 21 July 2017
(This article belongs to the Special Issue New Advances in Disinfection of Wastewater)
Abstract
:The aim of this study was to elucidate the inactivation effects on the antibiotic-resistance gene (vanA) of vancomycin-resistant enterococci (VRE) using chlorination, a disinfection method widely used in various water treatment facilities. Suspensions of VRE were prepared by adding VRE to phosphate-buffered saline, or the sterilized secondary effluent of a wastewater treatment plant. The inactivation experiments were carried out at several chlorine concentrations and stirring time. Enterococci concentration and presence of vanA were determined. The enterococci concentration decreased as chlorine concentrations and stirring times increased, with more than 7.0 log reduction occurring under the following conditions: 40 min stirring at 0.5 mg Cl2/L, 20 min stirring at 1.0 mg Cl2/L, and 3 min stirring at 3.0 mg Cl2/L. In the inactivation experiment using VRE suspended in secondary effluent, the culturable enterococci required much higher chlorine concentration and longer treatment time for complete disinfection than the cases of suspension of VRE. However, vanA was detected in all chlorinated suspensions of VRE, even in samples where no enterococcal colonies were present on the medium agar plate. The chlorine disinfection was not able to destroy antibiotic-resistance genes, though it can inactivate and decrease bacterial counts of antibiotic-resistant bacteria (ARB). Therefore, it was suggested that remaining ARB and/or antibiotic-resistance gene in inactivated bacterial cells after chlorine disinfection tank could be discharged into water environments.
1. Introduction
Antibiotics are mainly applied to treat bacterial infections, which are a major public health issue, and have received great attention [1]. Although antibiotic-resistant bacteria (ARB) have been regarded as serious causes of nosocomial infections thus far, there have been recent reports of their infection and detection in water environments such as rivers and coastal areas [2,3,4,5]. Concerns over water polluted by ARB have surfaced as a new water environmental issue. Given the widespread use and misuse of antibiotics in livestock and aquaculture industries as well as human health management, ARB have been frequently detected in water environments [6]. In fact, over 250 million antibiotic prescriptions are written annually in the United States of America [7]. Furthermore, many kinds of antibiotics are used as veterinary medicine, biocides in production, and feed additives in agricultural industries [8]. According to the World Health Organization (WHO), antibiotic resistance will become a critical global public health problem in the near future. As sewers are systems that effectively aggregate ARB and pathogenic microbes from urban areas, incorporating a process that would inactivate ARB in sewage treatment plants may considerably reduce their spread into aquatic environments [9,10]. However, a number of bacterial pathogens resistant to antibiotics including methicillin-resistant Staphylococcus aureus, Escherichia coli, Salmonella, and enterococci can survive wastewater treatment [11,12,13,14]. In particular, vancomycin-resistant enterococci (VRE) have recently been isolated from wastewater effluent [12,14,15], and reclaimed water used for spray irrigation [16]. Additionally, we conducted an investigation on a sewage treatment plant with the purpose of identifying the presence of VRE and their resistance genes, vanA and vanB. Although VRE were hardly detected in each sewage treatment process, there were no reports on the reduction in the number of vanA and vanB, and their presence was detected even in the chlorination tanks [17]. As VRE are listed in the ARB rankings at Priority 2: High in a priority pathogens list for R&D of new antibiotics released by WHO [18], VRE are an important ARB that pose the greatest threat to human health [18,19].
As chlorine destroys target microorganisms by oxidizing cellular materials, and is more cost-effectively than either UV or ozone, it is the most widely used disinfectant for municipal wastewater [20]. Previous studies have shown that a high concentration of chlorine could inactivate antibiotic-resistant heterotrophic bacteria in drinking water and wastewater [21]; however, how the resistance ARB gene behaves during chlorination remains unclear. Hence, to inhibit the spreading of ARB to water environments, understanding the inactivation effects on antibiotic-resistance genes along with ARB by chlorination is vital.
In this study, we aimed to elucidate the chlorine inactivation effects on VRE—considered particularly dangerous even among ARB—and their vanA resistance gene. A suspension of VRE was prepared by suspending a vanA-possessing Enterococcus faecium in phosphate-buffered saline (PBS), followed by measuring enterococci concentration changes with several chlorine concentration treatments and stirring times. DNA was then extracted from chlorinated samples to confirm the presence of vanA using PCR. The secondary effluent was collected from a wastewater treatment plant. Next, pre-cultured vanA-possessing E. faecium was suspended in a water sample after sterilization. Furthermore, this VRE suspended in secondary effluent water sample was used in the inactivation experiment with the same procedures as the suspension of VRE.
2. Materials and Methods
2.1. Inactivation of Suspensions of Vancomycin-Resistant Enterococci (VRE)
E. faecium ATCC 51559 (possessing vanA) was used as the standard VRE strain. E. faecium pre-cultured in 30 mL of Todd–Hewitt broth (Becton and Dickinson, Franklin Lakes, NJ, USA) was placed into 50-mL centrifuge tubes and centrifuged for 10 min at 17,534× g rcf. The supernatant was removed, and the E. faecium pellet was washed twice using 20 mL of phosphate-buffered saline (PBS, pH: 7.4). The pellet was subsequently resuspended in PBS, and its absorbance was measured at OD600, followed by adjusting the bacterial concentration to 108 CFU/mL. A 10-mL sample of this solution was mixed with 90 mL of PBS to make suspensions of VRE (bacterial concentration: 107 CFU/mL), which was then used in the inactivation experiments.
Sodium hypochlorite (Wako Pure Chemical, Osaka, Japan) was added to 100 mL of the VRE suspension in a 300 mL beaker to have initial chlorine concentrations of 0.0, 0.1, 0.3, 0.5, 1.0, and 3.0 mg Cl2/L. Subsequently, the solutions were mixed with magnetic stirrers at 150 rpm for a set amount of time (3, 5, 10, 20, 40, and 60 min). After stirring, 700 μL of 0.025 M of sodium thiosulfate (Wako Pure Chemical) was added, and the solution was stirred for another 3 min to neutralize the chlorine. These inactivation experiments were carried out at room temperature (around 25 °C). Enterococci concentration measurement and DNA extraction were then performed on the neutralized suspension of VRE.
2.2. Measurement of Enterococci Concentration
The enterococci concentration was measured in the chlorine-inactivated suspension of VRE. A 100 or 1000 μL sample was inoculated on m-Enterococcus agar (Becton and Dickinson, containing 1.5% agar) in triplicate and incubated at 35 ± 1.0 °C for 48 h, after which the red enterococci colonies were counted. An adequate amount of a high-density sample was diluted (1–105 time dilution series), and the diluted sample was inoculated. The limit of detection for enterococci was 1 CFU/mL, and samples with colony counts less than that were considered as below detection limit (<DL).
2.3. DNA Extraction and Resistance Gene Detection
Chlorine-inactivated suspensions of VRE (90 mL) were centrifuged for 10 min at 17,534× g rcf before the supernatant was removed. The pellets were resuspended in 1.5 mL of sterile distilled water and transferred into 1.5-mL tubes. Subsequently, DNA extraction was performed using the InstaGene Matrix (BioRad, Hercules, CA, USA) as per the instruction manual. The DNA extracts were stored at −20 °C until they were ready for use in vanA detection using PCR.
vanA detection using PCR was performed using the GoTaq® Green Master Mix (Promega, Madison, WI, USA). The PCR reaction solution was made up of 10 μL of 2 × GoTaq® Green Master Mix, 1.0 μL of forward and reverse primers (each at a 0.5 μM final concentration), 1.0 μL of template DNA, and 7.0 μL of nuclease-free water to a total volume of 20 μL. The nucleotide sequence for the forward primer vanAfw was 5′-GGGAAAACGACAATTGC-3′, and the reverse primer vanArv was 5′-GTACAATGCGGCCGTTA-3′ [22]. The PCR reaction conditions were as follows: Initial denaturation at 94 °C for 2 min, followed by another 1 min at 94 °C, annealing at 54 °C for 1 min, extension at 72 °C for 1 min for 30 cycles, with the final extension performed at 72 °C for 5 min. A 1.5% agarose gel electrophoresis was subsequently performed on the PCR products to confirm the presence of vanA. The amplicon size of vanA was 732 bp.
2.4. Inactivation of VRE in Secondary Effluent Samples
Secondary effluent samples were collected from a wastewater treatment plant (conventional activated sludge process, separate sewer system) in Oita prefecture, Japan. The collected water samples were stored in sterilized polyethylene bottles, transported to the laboratory, and used for inactivation experiments. Electrical conductivity, pH, turbidity, and the temperature of the water sample were 79.3 mS/m, 6.6, 2.6 degree (kaolin), and 17.7 °C, respectively. In the preliminary experiment, DNA in the secondary effluent was extracted using a DNeasy Blood & Tissue Kit (Qiagen, Hilden, Germany) as per Furukawa et al. [17]. vanA was not detected in the water sample of the secondary effluent. The secondary effluent sample was sterilized by autoclaving (121 °C, 15 min), and precultured E. faecium ATCC 51559 was suspended in the sterilized secondary effluent (bacterial concentration: 107 CFU/mL). Next, the inactivation experiment was carried out using the same procedures as above-mentioned. Sodium hypochlorite was added to a 100 mL of the VRE suspended in secondary effluent water sample in a 300-mL beaker to give it initial chlorine concentrations of 0.0, 0.5, 3.0, 5.0, and 10.0 mg Cl2/L. These inactivation experiments were carried out at room temperature (around 25 °C).
3. Results and Discussion
3.1. Inactivation Ratio of VRE and Detection of vanA in Suspensions of VRE (Suspended in PBS Solution)
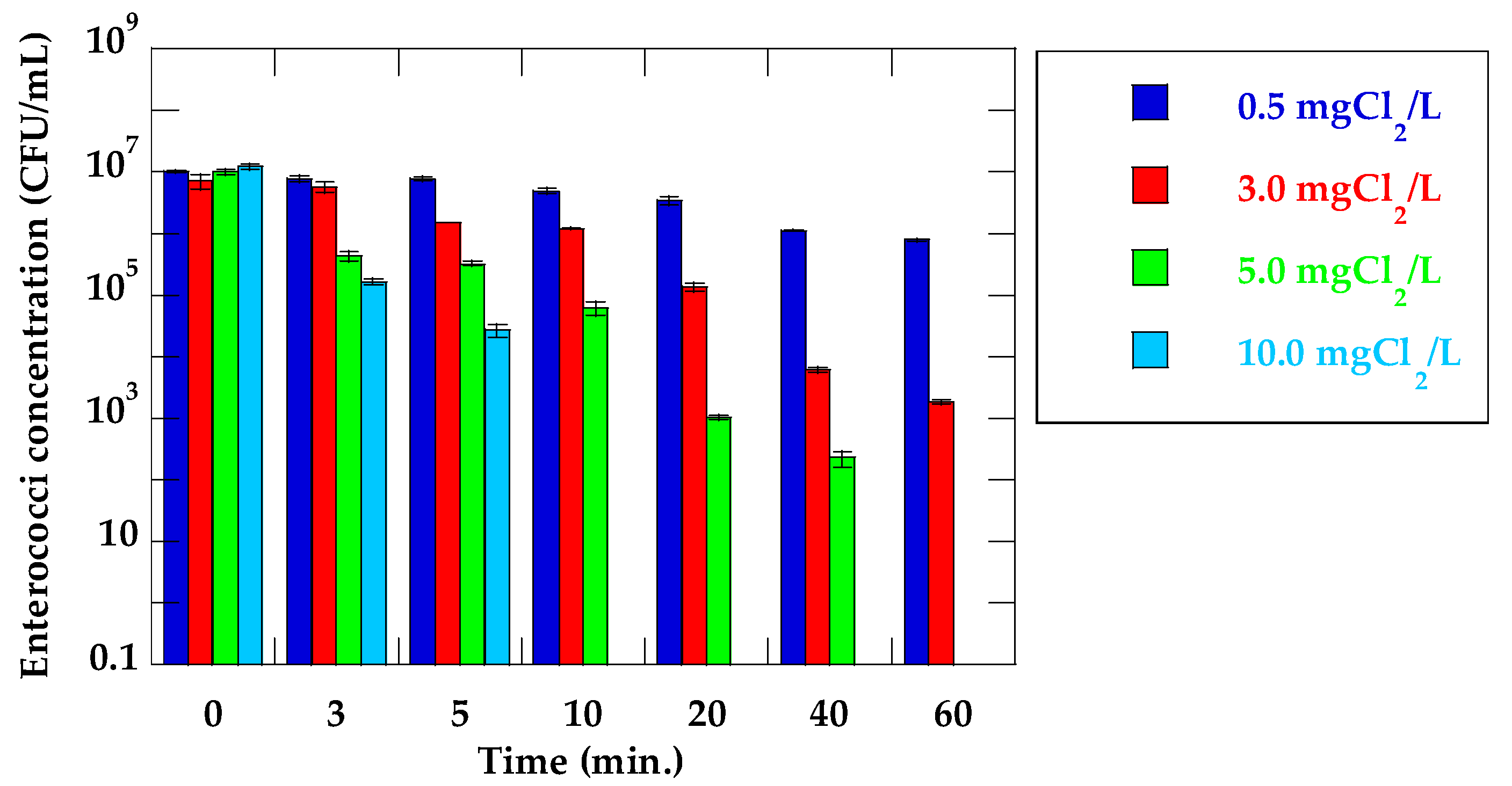
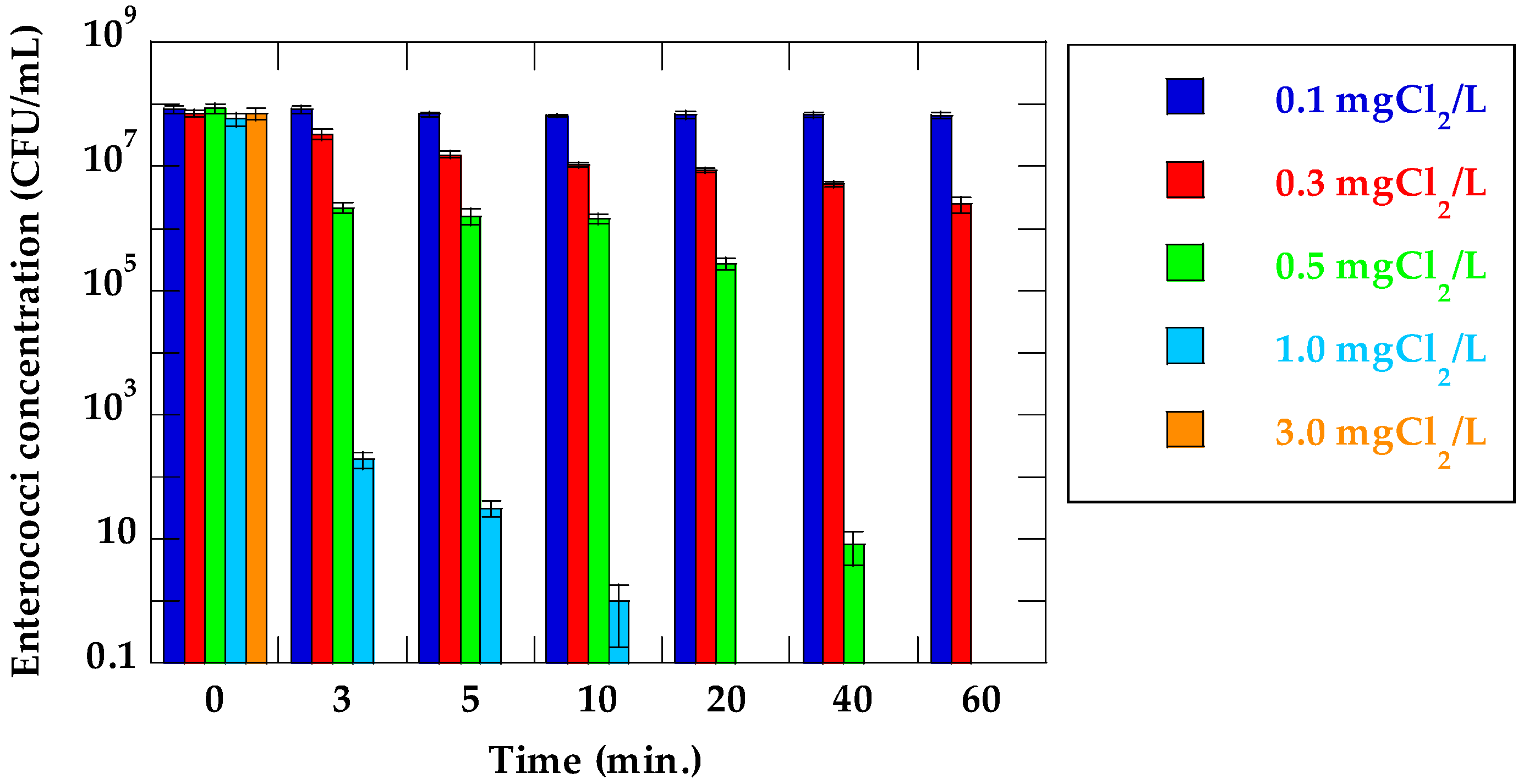
Changes in enterococci concentration and the log reduction value of suspensions of VRE in their respective chlorine concentrations and stirring times are shown in Figure 1 and Table 1, respectively. In cases where initial chlorine concentration was 0.1 mg Cl2/L, there were mostly no reductions of enterococci. A slight reduction in enterococci concentration was observed at 0.3 mg Cl2/L, but enterococci at an order of magnitude of 106 CFU/mL survived in the treated suspension, even after 60 min stirring time. At 0.5–3.0 mg Cl2/L, enterococci concentration decreased significantly with increasing initial chlorine concentrations and stirring times. Enterococci were <DL at initial chlorine concentrations and stirring times of 0.5 mg Cl2/L for 60 min and at 1.0 mg Cl2/L for 20 min, respectively. Furthermore, enterococci were <DL at 3.0 mg Cl2/L for all stirring times.
The inactivation ratio was calculated from the enterococci concentration of the untreated suspension of VRE (C0) and that of each treated sample (C). Enterococci inactivation increased as initial chlorine concentration and stirring time increased. The maximum inactivation ratio at 0.1 and 0.3 mg Cl2/L were 0.1log and 1.46log, respectively. In contrast, the maximum inactivation ratio at 0.5, 1.0, and 3.0 mg Cl2/L were above 7.5log. Like these results, fecal bacteria (fecal coliform and fecal streptococci) were destroyed at more than 99.999% in primary sewage effluent by chlorine disinfection [23].
Bacterial DNA was extracted from each chlorinated suspension of VRE. The presence of vanA in each suspension was determined by PCR (Table 2). The vanA was detected from all chlorinated suspensions of VRE by PCR, even in the sample where no enterococcal colonies were detected on m-Enterococcus agar plate.
3.2. Inactivation Ratio of VRE and Detection of vanA in Secondary Effluent Water Sample
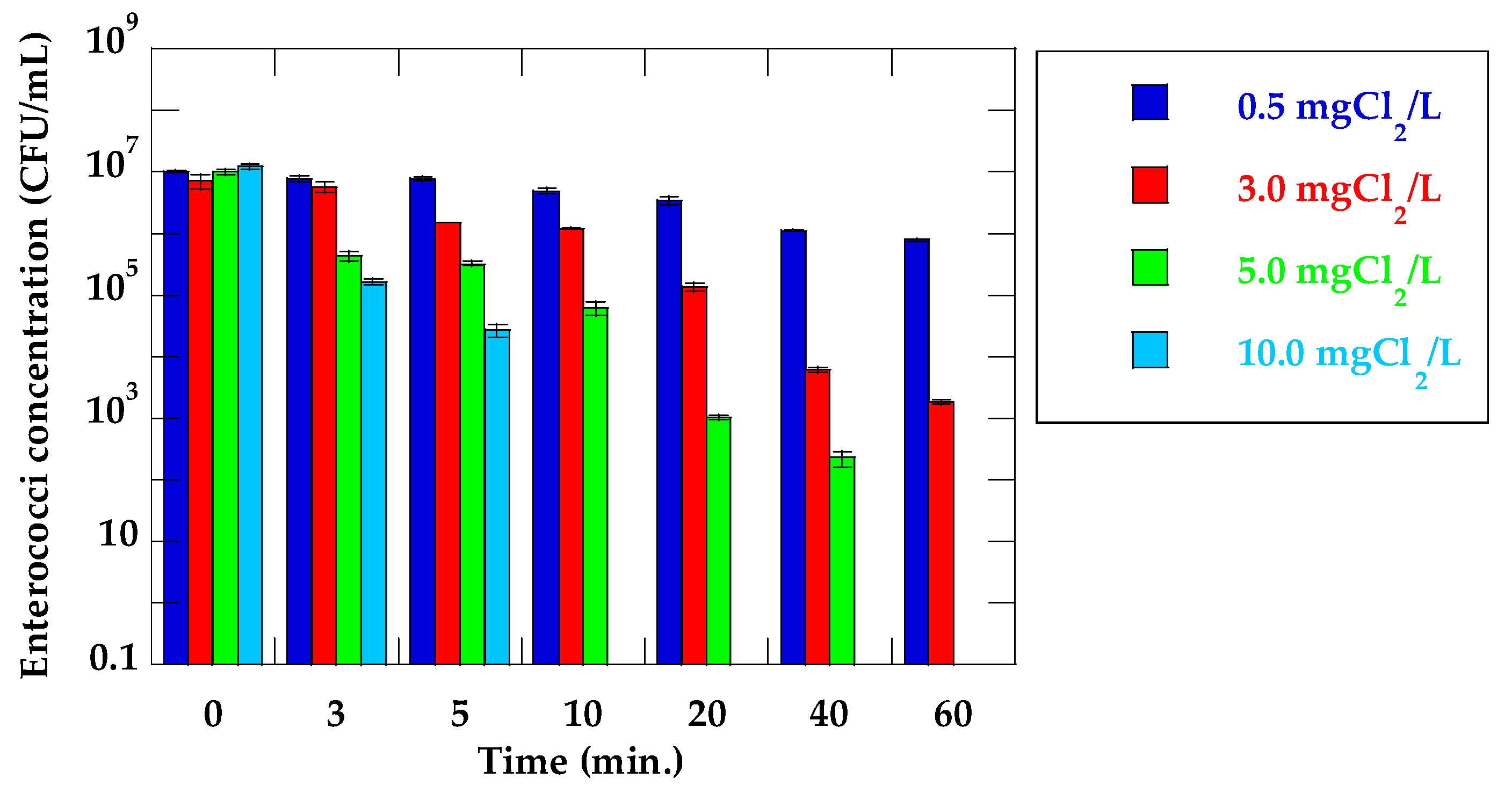
Changes in the enterococci concentration and log reduction value of VRE suspended in secondary effluent water samples in the respective initial chlorine concentrations and stirring times are shown in Figure 2 and Table 3, respectively. A high concentration of enterococci remained culturable in PBS at 0.5 mg Cl2/L and 60 min stirring time (7.9 × 105 CFU/mL). No enterococci was detected from VRE suspension at the condition in 5.0 mg Cl2/L and 60 min stirring time. Contrary to this result, with respect to VRE suspended in secondary effluent, enterococci that could not be inactivated remained in the chlorinated water sample even at the condition in 5.0 mg Cl2/L and 60 min stirring time (removal rate: 3.5log). No enterococci were detected in the chlorinated water samples at 5.0 mg Cl2/L and 60 min stirring time, and 10.0 mg Cl2/L and more than 10 min stirring time. High initial chlorine concentration and long treatment time were required to inactivate enterococci in secondary effluents when compared with those in VRE suspensions. Previous study has shown that chlorine dosage varies based on wastewater characteristics, and there are several other factors that ensure optimum conditions for disinfection [20]. Organic materials, ammonium-nitrogen, and organic nitrogen all consume chlorine. Due to the presence of these chlorine consumption materials in the secondary effluents, the large amount of chlorine was required to inactivate enterococci. Table 4 shows the presence of vanA in the chlorinated secondary effluent determined by PCR. vanA was detected in all chlorinated water samples as with the case of inactivation experiment using suspensions of VRE. Some studies on the inactivation of ARB and resistance genes by chlorination have been carried out. Pang et al. [24] showed that chlorination could inactivate ampicillin-resistant E.coli, but was not an efficient disinfection method for disrupting resistance genes. Yuan et al. [21] showed that all bacteria, except erythromycin- and sulfadiazine-resistant bacteria, were inactivated fully by 15 mg Cl2 min/L, and a chlorine dose of > 60 mg Cl2 min/L was needed to inactivate erythromycin- and sulfadiazine-resistant bacteria. Additionally, the proportion of multidrug-resistant bacteria isolated from the chlorinated effluent increases compared to the influent, and antibiotic-resistance genes remain detectable in the chlorinated effluent [25]. Huang et al. [26] also indicated that tetracycline-resistant E. coli showed a higher resistance to chlorine than tetracycline-sensitive E. coli. These results suggested that the tolerance to chlorine may vary depending on the type of ARB. Other previous studies have reported that higher doses of chlorine (25, 30 mg Cl2/L) compared with this study (max concentration: 10 mg Cl2/L) could decrease antibiotic resistance genes (sul1, tetX, tetG, and intI1) [27]. However, as the dose of chlorine usually ranges from 5–20 mg Cl2/L in actual wastewater treatment plants [20], these antibiotic-resistance genes and ARB cannot be decreased in the disinfection system. Considering that vanA was not quantified in this experiment, the question of whether vanA decreased through chlorination remains unclear. As vanA was detected in all water samples, including water samples in which no VRE colony was detected, it was clear that vanA could not be inactivated sufficiently, and persisted in water samples after chlorine disinfection. Yuan et al. [21] showed that erythromycin- and tetracycline-resistance genes were detected in the chlorination disinfection tank in a wastewater treatment plant, indicating that antibiotic-resistance genes may not be inactivated by chlorination. Other studies have demonstrated that chlorination contributes to resistant strain selection. Armstrong et al. [28] noted that the proportion of multiple antibiotic-resistant phenotypes increased during chlorination for Gram-positive cocci and Gram-negative, non-fermentative rods. In addition, Jia et al. [29] revealed that a total of 151 antibiotic-resistance genes were detectable in the drinking water, and chlorination evidently increase their total relative abundance, while reducing their diversity in the opportunistic bacteria.
In chlorine disinfection, compounds such as HOCl have a strong oxidizability that can inhibit enzyme functions across the cell membrane of microorganisms, and its inactivation effects have been shown to destroy bacterial cell walls [20]. Thus, although chlorine inhibits ARB survival and growth, it may not be able to damage antibiotic-resistance genes such as vanA. Therefore, our results suggested that any ARB survivors and/or antibiotic-resistance genes in inactivated bacterial cells after chlorine disinfection could be discharged into the water environment. Another possible scenario is that enterococci stressed by chlorination might be viable, but at a non-culturable (VBNC) stage. Although enterococci in a VBNC stage could not grow on an m-Enterococcus agar plate, the detectable vanA genes remained in bacterial cells by PCR after chlorine disinfection. As the VBNC bacteria can be alive, show very low levels of metabolic activity, and can become active again in certain conditions [30], the infection risk caused by the reactivation of VRE in a VBNC stage should not be ignored in aquatic environments.
The resistance gene of VRE, vanA, is localized in the plasmid, and such plasmids allow the horizontal propagation of genes between bacterial cells. The results acquired in this study implied that even if the VRE cells themselves were inactivated by chlorine, it cannot be denied that persisting antibiotic-resistance genes may horizontally transfer to other microbial cells, giving rise to new ARB. The mechanisms of acquiring antibiotic-resistance genes in bacteria are mutation, horizontal transfer, and the inducing of genes [31], as horizontal gene transfer plays an important role in the adaptation of microorganisms to the environmental changes. The types of horizontal gene transfer are transformation, conjugation, and transduction [32,33,34]. Furthermore, in certain environments, extracellular DNA is present in 25 times greater than the intercellular DNA of bacteria [35,36], indicating that a significant amount of DNA (including plasmid DNA possessing antibiotic-resistance genes) might be present in wastewater treatment systems. A wastewater treatment system contains enormous amounts of microorganisms, enrich nutrients, and various antibiotics. A wastewater treatment system may become a suitable environment not only for the survival of microorganisms , but also for the occurrence of new ARB caused by mutation and gene inducing, as well as horizontal gene transfer. Consequently, it is likely that a wastewater treatment system pools ARB and/or antibiotic-resistance genes, given that heterotrophic bacteria at an order of magnitude of 105 CFU/100 mL survived in the chlorine disinfection tank [37], could serve as a carrier of antibiotic- resistance genes. Our results showed that the effect of chlorine disinfection could not destroy antibiotic-resistance genes, though it could inactivate and decrease bacterial counts of ARB. On the other hand, it has been reported that UV can decrease ARB more effectively than chlorination [21,26]. Sequential UV/chlorination, in particular, enhanced the inactivation of antibiotic resistance genes [27]. Furthermore, ozone and TiO2-photocatalysis can induce damage in the plasmid DNA structure, and are expected to help reduce the risk of antibiotic resistance pollution and antibiotic transfer [38]. On the contrary, Dunlop et al. [39] showed that although antibiotic-resistant E. coli was inactivated by photocatalysis using a TiO2-UVA lamp, a full recovery of E. coli was observed during post treatment incubation for 24 h at 37 °C. As shown above, it has been suggested that these disinfectants have a certain inactivating efficiency for ARB along with antibiotic-resistance genes, but could not inactivate them completely. In addition, these inactivation techniques are far less costly than chlorine disinfection. To prevent the spread of ARB to environmental waters, it might be necessary to introduce new disinfection technologies that can eliminate ARB together with antibiotic-resistance genes in wastewater treatment facilities.
4. Conclusions
The aim of this study was to clarify the inactivation effects of chlorination on VRE, a type of ARB, as well as on its antibiotic-resistance gene, vanA.
(1) In the inactivation experiment of varying initial chlorine concentrations and stirring times, the inactivation ratios of VRE suspended in PBS increased as both chlorine concentrations and stirring times increased above 0.5 mg Cl2/L. The maximum inactivation ratio for 0.5, 1.0, and 3.0 mg Cl2/L was shown to be more than 7.5 log reduction.
(2) In the inactivation experiment using VRE suspended in secondary effluent, inactivating enterococci required much higher chlorine concentration and longer treatment time than the cases of suspensions of VRE (10 mg Cl2/L, more than 10 min stirring time).
(3) The presence of vanA was confirmed in the extracted DNA of each chlorinated sample using PCR. The vanA was detected in all samples, including samples where enterococci were not detected by plating methods, suggesting that the resistance gene remained in chlorinated water. In other words, despite achieving the inactivation of VRE with chlorine disinfection, the damage did not extend to the resistance gene.
Acknowledgment
This work was supported by Grants-in-Aid for Scientific Research, Japan Society for the Promotion of Science, Grant Number 15K21596, and the Maeda Engineering Foundation, Japan.
Author Contributions
Takashi Furukawa and Takahisa Ueno conceived and designed the experiments; Takashi Furukawa and Atsushi Jikumaru performed experiments and analyzed the data; Takashi Furukawa wrote the paper; Takashi Furukawa, Takahisa Ueno, and Kazunari Sei revised the paper.
Conflicts of Interest
The authors declare no conflict of interest. The founding sponsors had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, and in the decision to publish the results.
References
- Sharma, V.K.; Johnson, N.; Cizmas, L.; McDonald, T.J.; Kim, H. A review of the influence of treatment strategies on antibiotic resistant bacteria and antibiotic resistance genes. Chemosphere 2016, 150, 702–714. [Google Scholar] [CrossRef] [PubMed]
- Ham, Y.S.; Kobori, H.; Kang, J.H.; Matsuzaki, T.; Iino, M.; Nomura, H. Distribution of antibiotic resistance in urban watershed in Japan. Environ. Pollut. 2012, 162, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Sidrach-Cardona, R.; Bècares, E. Fecal indicator bacteria resistance to antibiotics in experimental constructed wetlands. Ecol. Eng. 2013, 50, 107–111. [Google Scholar] [CrossRef]
- Suzuki, Y.; Kajii, S.; Nishiyama, M.; Iguchi, A. Susceptibility of Pseudomonas aeruginosa isolates collected from river water in Japan to antipseudomonal agents. Sci. Total Environ. 2013, 450–451, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Urase, T.; Sato, T. Quantitative monitoring of resistance in Escherichia coli to clinically important antimicrobials in urban watershed. J. Water Environ. Technol. 2016, 14, 341–349. [Google Scholar] [CrossRef]
- Ash, R.J.; Mauck, B.; Morgan, M. Antibiotic resistance of gram-negative bacteria in rivers, United States. Emerg. Infect. Dis. 2002, 8, 713–716. [Google Scholar] [CrossRef] [PubMed]
- Rosi-Marshall, E.J.; Kelly, J.J. Antibiotic stewardship should consider environmental fate of antibiotics. Environ. Sci. Technol. 2015, 49, 5257–5258. [Google Scholar] [CrossRef] [PubMed]
- Silbergeld, E.K.; Graham, J.; Price, L.B. Industrial food animal production, antimicrobial resistance, and human health. Annu. Rev. Public Health 2008, 29, 151–169. [Google Scholar] [CrossRef] [PubMed]
- Novo, A.; André, S.; Viana, P.; Nunes, O.C.; Manaia, C.M. Antibiotic resistance, antimicrobial residues and bacterial community composition in urban wastewater. Water Res. 2013, 47, 1875–1887. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, L.; Manaia, C.; Merlin, C.; Schwartz, T.; Dagot, C.; Ploy, M.C.; Michael, I.; Fatta-Kassinos, D. Urban wastewater treatment plants as hotspots for antibiotic resistant bacteria and genes spread into the environment: A review. Sci. Total Environ. 2013, 447, 345–360. [Google Scholar] [CrossRef] [PubMed]
- Levantesi, C.; La Mantia, R.; Masciopinto, C.; Böckelmann, U.; Ayuso-Gabella, M.N.; Salgot, M.; Tandoi, V.; Van Houtte, E.; Wintgens, T.; Grohmann, E. Quantification of pathogenic microorganisms and microbial indicators in three wastewater reclamation and managed aquifer recharge facilities in Europe. Sci. Total Environ. 2010, 408, 4923–4930. [Google Scholar] [CrossRef] [PubMed]
- Nagulapally, S.R.; Ahmad, A.; Henry, A.; Marchin, G.L.; Zurek, L.; Bhandari, A. Occurrence of ciprofloxacin-, trimethoprim-sulfamethoxazole-, and vancomycin-resistant bacteria in a municipal wastewater treatment plant. Water Environ. Res. 2009, 81, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg Goldstein, R.E.; Micallef, S.A.; Gibbs, S.G.; Davis, J.A.; He, X.; George, A.; Kleinfelter, L.M.; Schreiber, N.A.; Mukherjee, S.; Sapkota, A.; et al. Methicillin-resistant Staphylococcus aureus (MRSA) Detected at Four U.S. wastewater treatment plants. Environ. Health Perspect. 2012, 120, 1551–1558. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg Goldstein, R.E.; Micallef, S.A.; Gibbs, S.G.; George, A.; Claye, E.; Sapkota, A.; Joseph, S.W.; Sapkota, A.R. Detection of vancomycin-resistant enterococci (VRE) at four U.S. wastewater treatment plants that provide effluent for reuse. Sci. Total Environ. 2014, 466–467, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Garcia, S.; Wade, B.; Bauer, C.; Craig, C.; Nakaoka, K.; Lorowitz, W. The effect of wastewater treatment on antibiotic resistance in Escherichia coli and Enterococcus sp. Water Environ. Res. 2007, 79, 2387–2395. [Google Scholar] [CrossRef] [PubMed]
- Carey, S.A.; Goldstein, R.E.R.; Gibbs, S.G.; Claye, E.; He, X.; Sapkota, A.R. Occurrence of vancomycin-resistant and -susceptible Enterococcus spp. in reclaimed water used for spray irrigation. Environ. Res. 2016, 147, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, T.; Hashimoto, R.; Mekata, T. Quantification of vancomycin-resistant enterococci and corresponding resistance genes in a sewage treatment plant. J. Environ. Sci. Health A Tox. Hazard. Subst. Environ. Eng. 2015, 50, 989–995. [Google Scholar] [CrossRef] [PubMed]
- World Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics. Available online: http://www.who.int/medicines/publications/WHO-PPL-Short_Summary_25Feb-ET_NM_WHO.pdf?ua=1 (accessed on 19 June 2017).
- Gold, H.S.; Moellering, R.C., Jr. Antimicrobial-drug resistance. N. Eng. J. Med. 1996, 335, 1445–1453. [Google Scholar] [CrossRef]
- U.S. Environmental Protection Agency. Wastewater Technology Fact Sheet: Chlorine Disinfection, EPA 832-F-99-062; U.S. EPA: Washington, DC, USA, 1999.
- Yuan, Q.; Guo, M.; Yang, J. Fate of antibiotic resistant bacteria and genes during wastewater chlorination: Implication for antibiotic resistance control. PLoS ONE 2015, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Dutka-Malen, S.; Evers, S.; Courvalin, P. Detection of glycopeptide resistance genotypes and identification to the species level of clinically relevant enterococci by PCR. J. Clin. Microbiol. 1995, 33, 24–27, PMCID:PMC227872. [Google Scholar] [PubMed]
- Tree, J.A.; Adams, M.R.; Lees, D.N. Chlorination of indicator bacteria and viruses in primary sewage effluent. Appl. Environ. Microbiol. 2003, 69, 2038–2043. [Google Scholar] [CrossRef] [PubMed]
- Pang, Y.; Huang, J.; Xi, J.; Hu, H.; Zhu, Y. Effect of ultraviolet irradiation and chlorination on ampicillin-resistant Escherichia coli and its ampicillin resistance gene. Front. Environ. Sci. Eng. 2016, 10, 522–530. [Google Scholar] [CrossRef]
- Al-Jassim, N.; Ansari, M.; Harb, M.; Hong, P.Y. Removal of bacterial contaminants and antibiotic resistance genes by conventional wastewater treatment processes in Saudi Arabia: Is the treated wastewater safe to reuse for agricultural irrigation? Water Res. 2015, 73, 277–290. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Hu, H.; Wu, Y.; Wei, B.; Lu, Y. Effect of chlorination and ultraviolet disinfection on tetA-mediated tetracycline resistance of Escherichia coli. Chemosphere 2013, 90, 2247–2253. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhuang, Y.; Geng, J.; Ren, H.; Zhang, Y.; Ding, L.; Xu, K. Inactivation of antibiotic resistance genes in municipal wastewater effluent by chlorination and sequential UV/chlorination disinfection. Sci. Total Environ. 2015, 512–513, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, J.L.; Shigeno, D.S.; Calomiris, J.J.; Seidler, R.J. Antibiotic-resistant bacteria in drinking water. Appl. Environ. Microbiol. 1981, 42, 277–283, PMCID:PMC244002. [Google Scholar]
- Jia, S.; Shi, P.; Hu, Q.; Li, B.; Zhang, T.; Zhang, X.X. Bacterial community shift drives antibiotic resistance promotion during drinking water chlorination. Environ. Sci. Technol. 2015, 49, 12271–12279. [Google Scholar] [CrossRef] [PubMed]
- Oliver, J.D. The viable but nonculturable state in bacteria. J. Microbiol. 2005, 43, 93–100. [Google Scholar] [PubMed]
- Hashimoto, H.; Murayama, S. Emergence and evolution of microbial drug-resistance. J. Jpn. Soc. Clin. Microbiol. 2013, 23, 1–11. [Google Scholar]
- Lederberg, J. Genetic recombination in bacteria: A discovery account. Annu. Rev. Genet. 1987, 21, 23–46. [Google Scholar] [CrossRef] [PubMed]
- Lederberg, J.; Tatum, E.L. Gene recombination in Escherichia coli. Nature 1946, 158, 558, PMCID:PMC518375. [Google Scholar] [CrossRef]
- Zinder, N.D.; Lederberg, J. Genetic exchange in Salmonella. J. Bacteriol. 1952, 64, 679–699, PMCID:PMC169409. [Google Scholar]
- Dell’anno, A.; Faviano, M.; Mei, M.L.; Danovaro, R. Pelagic-benthic coupling of nucleic acids in an abyssal location of the northeastern Atlantic Ocean. Appl. Environ. Microbiol. 1999, 65, 4451–4457, PMCID:PMC91592. [Google Scholar]
- Lorenz, M.G.; Wackernagel, W. Bacterial gene transfer by natural genetic transformation in the environment. Microbiol. Rev. 1994, 58, 563–602, PMCID:PMC372978. [Google Scholar]
- Hashimoto, R.; Furukawa, T.; Suzuki, Y. Fate of vancomycin-resistant bacteria and corresponding resistance genes in a sewage treatment plant. In Proceedings of the Water and Environment Technology Conference 2016, Tokyo, Japan, 27–28 August 2016. [Google Scholar]
- Öncü, N.B.; Menceloğlu, Y.Z.; Balcıoğlu, I.A. Comparison of the effectiveness of chlorine, ozone, and photocatalytic disinfection in reducing the risk of antibiotic resistance pollution. J. Adv. Oxid. Technol. 2011, 14, 196–203. [Google Scholar] [CrossRef]
- Dunlop, P.S.M.; Ciavola, M.; Rizzo, L.; McDowell, D.A.; Byrne, J.A. Effect of photocatalysis on the transfer of antibiotic resistance genes in urban wastewater. Catal. Today 2015, 240, 55–60. [Google Scholar] [CrossRef]
Figure 1.
Changes of enterococci concentration (suspension of VRE).

Figure 2.
Changes of enterococci concentration (VRE suspended in secondary effluent).

{kind=link}
{kind=link}
Table 1.
Log reduction value for each initial chlorine concentration and stirring time (suspension of vancomycin-resistant enterococci in PBS solution).
Table 1.
Log reduction value for each initial chlorine concentration and stirring time (suspension of vancomycin-resistant enterococci in PBS solution).
| Time (min) | Log Reduction (log C/C0) | ||||
|---|---|---|---|---|---|
| 0.1 mg Cl2/L | 0.3 mg Cl2/L | 0.5 mg Cl2/L | 1.0 mg Cl2/L | 3.0 mg Cl2/L | |
| 3 | 0.00 | 0.33 | 1.59 | 5.48 | >7.85 |
| 5 | 0.08 | 0.66 | 1.73 | 6.27 | >7.85 |
| 10 | 0.10 | 0.83 | 1.78 | 7.76 | >7.85 |
| 20 | 0.09 | 0.91 | 2.49 | >7.77 | >7.85 |
| 40 | 0.09 | 1.14 | 7.01 | >7.77 | >7.85 |
| 60 | 0.10 | 1.46 | >7.93 | >7.77 | >7.85 |
Table 2.
vanA detection for each initial chlorine concentration and treatment time (suspensions of VRE in PBS solution).
Table 2.
vanA detection for each initial chlorine concentration and treatment time (suspensions of VRE in PBS solution).
| Time (min) | vanA Detection (-) | ||||
|---|---|---|---|---|---|
| 0.1 mg Cl2/L | 0.3 mg Cl2/L | 0.5 mg Cl2/L | 1.0 mg Cl2/L | 3.0 mg Cl2/L | |
| 0 | + | + | + | + | + |
| 3 | + | + | + | + | + |
| 5 | + | + | + | + | + |
| 10 | + | + | + | + | + |
| 20 | + | + | + | + | + |
| 40 | + | + | + | + | + |
| 60 | + | + | + | + | + |
Notes: + means vanA was amplified by PCR. Shaded cell means that enterococci concentration was <DL.
Table 3.
Log reduction value in each initial chlorine concentration and stirring time (VRE suspended in secondary effluent).
Table 3.
Log reduction value in each initial chlorine concentration and stirring time (VRE suspended in secondary effluent).
| Time (min) | Log Reduction (log C/C0) | |||
|---|---|---|---|---|
| 0.5 mg Cl2/L | 3.0 mg Cl2/L | 5.0 mg Cl2/L | 10 mg Cl2/L | |
| 3 | 0.12 | 0.33 | 1.37 | 1.86 |
| 5 | 0.11 | 0.90 | 1.50 | 2.65 |
| 10 | 0.32 | 0.77 | 2.21 | >7.09 |
| 20 | 0.47 | 1.73 | 3.99 | >7.09 |
| 40 | 0.96 | 3.07 | 4.65 | >7.09 |
| 60 | 1.12 | 3.59 | >7.01 | >7.09 |
Table 4.
vanA detection in each initial chlorine concentration and treatment time (VRE suspended in secondary effluent).
Table 4.
vanA detection in each initial chlorine concentration and treatment time (VRE suspended in secondary effluent).
| Time (min) | vanA Detection (-) | |||
|---|---|---|---|---|
| 0.5 mg Cl2/L | 3.0 mg Cl2/L | 5.0 mg Cl2/L | 10 mg Cl2/L | |
| 0 | + | + | + | + |
| 3 | + | + | + | + |
| 5 | + | + | + | + |
| 10 | + | + | + | + |
| 20 | + | + | + | + |
| 40 | + | + | + | + |
| 60 | + | + | + | + |
Notes: + means vanA was amplified by PCR. Shaded cell means that enterococci concentration was <DL.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Furukawa, T.; Jikumaru, A.; Ueno, T.; Sei, K. Inactivation Effect of Antibiotic-Resistant Gene Using Chlorine Disinfection. Water 2017, 9, 547. https://doi.org/10.3390/w9070547
AMA Style
Furukawa T, Jikumaru A, Ueno T, Sei K. Inactivation Effect of Antibiotic-Resistant Gene Using Chlorine Disinfection. Water. 2017; 9(7):547. https://doi.org/10.3390/w9070547
Chicago/Turabian StyleFurukawa, Takashi, Atsushi Jikumaru, Takahisa Ueno, and Kazunari Sei. 2017. "Inactivation Effect of Antibiotic-Resistant Gene Using Chlorine Disinfection" Water 9, no. 7: 547. https://doi.org/10.3390/w9070547
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.