
Rangelands: Where Anthromes Meet Their Limits

1
Department of Geography, University of California-Berkeley, Berkeley, CA 94720, USA
2
Department of History, University of California-Davis, Davis, CA 95616, USA
3
USDA-ARS-Jornada Experimental Range, New Mexico State University, Las Cruces, NM 88003, USA
*
Author to whom correspondence should be addressed.
Land 2017, 6(2), 31; https://doi.org/10.3390/land6020031
Submission received: 18 January 2017
/
Revised: 27 March 2017
/
Accepted: 26 April 2017
/
Published: 1 May 2017
(This article belongs to the Special Issue Anthropogenic Biomes)
Abstract
:Defining rangelands as anthromes enabled Ellis and Ramankutty (2008) to conclude that more than three-quarters of Earth’s land is anthropogenic; without rangelands, this figure would have been less than half. They classified all lands grazed by domestic livestock as rangelands, provided that human population densities were low; similar areas without livestock were excluded and classified instead as ‘wildlands’. This paper examines the empirical basis and conceptual assumptions of defining and categorizing rangelands in this fashion.
Empirically, we conclude that a large proportion of rangelands, although used to varying degrees by domesticated livestock, are not altered significantly by this use, especially in arid, highly variable environments and in settings with long evolutionary histories of herbivory by wild animals. Even where changes have occurred, the dynamics and components of many rangelands remain structurally and functionally equivalent to those that preceded domestic livestock grazing or would be found in its absence. In much of Africa and Asia, grazing is so longstanding as to be inextricable from ‘natural’ or reference conditions for those sites. Thus, the extent of anthropogenic biomes is significantly overstated.
Conceptually, rangelands reveal the dependence of the anthromes thesis on outdated assumptions of ecological climax and equilibrium. Coming to terms with rangelands—how they can be classified, understood, and managed sustainably—thus offers important lessons for understanding anthromes and the Anthropocene as a whole. At the root of these lessons, we argue, is not the question of human impacts on ecosystems but property relations among humans.
Edges, whether of thought or of country, are revealing places for the inquirer.—F. Fraser Darling (1956, 779 [1])
1. Introduction
Ellis and Ramankutty (2008, 439 [2]) contend that Earth’s terrestrial ecosystems should be conceived as anthromes (or anthropogenic biomes) because “More than 75% of Earth’s ice-free land showed evidence of alteration as a result of human residence and land use, with less than a quarter remaining as wildlands.” Their anthromes thesis has unquestionably struck a chord in and beyond scientific circles. According to Google Scholar, the paper had been cited nearly 800 times as of early 2017, and over 50 of those sources had themselves been cited 100 times or more. Some of the world’s most renowned ecologists are among those who have cited the 2008 paper (e.g., Terry Chapin, Peter Vitousek, Monica Turner, Richard Hobbs, Carl Folke, and Steward Pickett), and several follow-up papers (Ellis et al. 2010, Ellis 2011, Ellis et al. 2013, Ellis 2015 [3,4,5,6]) have likewise been heavily cited. Maps of global anthromes have circulated widely in the blogosphere and in popular media such as National Geographic.
In its substance, the anthromes thesis has antecedents reaching back many hundreds of years. The idea of human alteration of ecosystems at large spatial and temporal scales was expressed by ancient Greek thinkers such as Plato and Theophrastus (Marsh 1864 [7]; Glacken 1967 [8]), and across most of the last century, the notion that humanity was surpassing Earth’s biophysical limits found expression in lexicons of soil erosion, over-population, and resource depletion, often hitched to apocalyptic prognoses of national, civilizational, or planetary collapse (Bennett and Chapline 1928, Sears 1935, Vogt 1948, Hardin 1968, Meadows et al. 1972 [9,10,11,12,13]). In a more scholarly vein, the early ecologist A. G. Tansley (1923, 48; [14]) gave the term “anthropogenic” its current meaning while rebutting F. E. Clements’ (1916, [15]) contention that human-altered sites were not real or true “climax” communities—exactly the contrast that Ellis and Ramankutty draw between anthromes and biomes. A quarter-century later, geographer Carl Sauer (1950, [16]) expanded Tansley’s thesis to embrace all of the world’s grasslands, which he argued were not climatic climaxes but rather had coevolved with humans and fire for some 200,000 years. The famous 1955 conference, Man’s Role in Changing the Face of the Earth (Thomas 1956, [17]), which Sauer co-organized, produced two volumes of papers that foreshadowed today’s debates about the Anthropocene (Crutzen and Stoermer 2000, [18]) in remarkable detail. These antecedents are often overlooked, however. For example, the arguments and evidence marshaled by Vitousek et al. (1997, [19]) closely resemble those of Holdren and Ehrlich (1974, 287 [20]), who reached effectively the same conclusion: “As a global geological and biological force, mankind is today becoming comparable to and even exceeding many natural processes.”
If the content was not new, the form in which Ellis and Ramankutty (2008 [2]) presented their anthromes thesis was: a quantitative metric of anthropogenic biomes at a global scale. Remote sensing, GIS, and computational algorithms enabled them to convert large and diverse datasets into a single, spatially explicit model output. Ostensibly, the model and the resulting maps integrated all of the varieties of human activities and biophysical settings, such that the measurement of anthromes obtained a simplicity and precision that Holdren and Ehrlich, let alone Tansley, could have only imagined. We would postulate that the output also surpassed an unstated threshold of qualitative significance that was critical to the paper’s reception. By this we mean that “more than three-quarters” made the notion of an “anthropogenic biosphere” (Ellis 2015 [6]) seem irrefutable, whereas “nearly half” would have left the case ambiguous. Ellis and Ramankutty’s classification of rangelands as anthromes was necessary to achieve this effect, because the model output would indeed have been less than 50% if rangelands—which make up 30.4% of all ice-free lands, according to the paper—were not included as anthromes.
In this sense, the anthromes thesis can be said to depend on the answer to the question: To what extent does livestock grazing alter the ecological characteristics and attributes of rangelands? No simple answer to this question exists, however. Different rangeland ecosystems respond differently to livestock grazing, for various reasons, and grazing itself is a complex, highly variable disturbance. In many rangelands, the effects of domestic livestock may be ecologically similar to those of wild animals, other organisms, or abiotic drivers such as fire or drought (Vavra et al. 1994 [21], Nicholson 2011 [22], Herrmann and Hutchinson 2006 [23], McNaughton 1985 [24], Mortimore 2009 [25], Whitford 2002 [26]). Moreover, there is no scientific consensus about the extent of rangeland degradation or even how such degradation ought to be defined (Ash et al. 2002 [27], Behnke 2002 [28], Gillson and Hoffman 2007 [29], Milchunas and Lauenroth 1993 [30], Sprugel 1991 [31], Véron et al. 2006 [32]). The most recent scholarship in rangeland ecology concludes that in rangelands with high (≥0.33) coefficients of variation of precipitation, ecosystem dynamics are driven by abiotic factors, with livestock typically having little or no large-scale, long-term impact (Behnke et al. 1993 [33], Ellis and Swift 1988 [34], Gillson and Hoffman 2007 [29], Kratli 2015 [35], Reynolds et al. 2007 [36], Scoones 1995 [37], Vetter 2005 [38], von Wehrden 2012 [39]). For all of these reasons, we conclude that the extent of anthropogenic transformation of Earth’s ice-free surface, as reported by Ellis and Ramankutty (2008 [2]), is substantially overstated.
This, in turn, raises the conceptual question: What specifically constitutes anthropogenic influence on ecosystems? Rangelands pose this question more profoundly than other types of land, and in more recent papers, Ellis (2011 [4]) and his colleagues (Ellis et al. 2010 [3]) have questioned the original anthromes thesis as it applies to rangelands, as we will see below. Ellis and Ramankutty (2008, 445 [2]) remained agnostic as to the positive or negative effects of human influences on ecosystems, preferring simply to say that “human and natural systems” are inextricably intermingled “almost everywhere on Earth’s terrestrial surface, demonstrating that interactions between these systems can no longer be avoided in any substantial way”. Defining rangelands by population density and land use, rather than ecological effects, was consistent with this stance. Such value neutrality is commendable, and for some scientists it may have lent the paper an attractive objectivity. It is not clear how one can avoid the question of impacts, however, at least not in the case of rangelands. Given the legacies of 20th century debates, alluded to already, as well as the title of one of the follow-up papers—“Used planet: a global history” (Ellis et al. 2013 [5])—the unmistakable connotation is that anthromes are at least disrupted and probably degraded in some fashion relative to non-anthropogenic wildlands. Few would dispute this for urban and cultivated lands, or for clear-cut forests and dammed rivers, and some grazed lands are indeed produced by clearing forests. However, a large majority of rangelands—87.8% by Ellis and Ramankutty’s (2008 [2]) estimation—are not. We propose here that the anthromes thesis should be understood not in terms of human impacts on ecosystems—a conceptualization that both presupposes and reinforces the very nature-culture dualism that the anthromes concept aspires to overcome—but in terms of social relations among humans with respect to the environment, specifically relations concerning land as property.
2. The Empirical Basis
Defining and measuring rangelands are not simple tasks. Ellis and Ramankutty (2008 [2]) arrived at an estimate of global rangelands that resembles other studies, but several important qualifications must be noted. Their definition of rangelands is unusual in its near-total dependence on land use: setting aside croplands and areas of dense human settlement or forest cover, they consider all remaining lands that are grazed by livestock to be rangelands, while excluding those without livestock. They estimate the extent of the world’s rangelands, defined in this way, to be 39.74 million km2, or 30.4% of all ice-free lands: ‘remote rangelands’, with a human population density of zero, make up 20.91 million km2; ‘populated rangelands’, with an average density of four persons km2, make up another 11.52 million km2; ‘residential rangelands’ make up the rest and have an average density of 32 persons km2, an order of magnitude lower than in ‘village’ anthromes with pastoral components.
Areas defined in this way are more commonly referred to as extensive grazing lands, a category that overlaps but is non-identical with current definitions of rangelands; a third category, drylands, overlaps but is non-identical with both. Other estimates of Earth’s rangelands vary significantly in both directions, in part due to different definitions. Ecologists and range scientists today use rangelands to encompass a number of land cover types or biomes, including savanna, grassland, steppe, desert, shrubland, and tundra. These aggregate to 74.4 million km2 (or 56.4% of all ice-free land), according to Asner et al. (2004, 266 [40]). Interestingly, the same study also found that only 26.0 million km2 of this area was actually grazed, or less than two-thirds of Ellis and Ramankutty’s estimate. On the other hand, Reid, Galvin, and Kruska (2008, [41]) estimated that ‘extensive grazing lands’ encompass 55.7 million km2 (42.2%), without asking whether they were in fact being grazed. Finally, drylands make up some 61.5 million km2 or 47.2% of ice-free land (Reynolds and Stafford Smith 2002, 2 [42]). ‘Drylands’ are defined as areas where the ratio of mean annual precipitation to mean annual potential evapo-transpiration is <0.65. Of this, 12.9 million km2 (9.9%) are dry subhumid (P/PET between 0.50 and 0.65). If one excludes hyper-arid areas (P/PET ratio < 0.05), where the lack of vegetation makes livestock grazing impossible in most years, the extent of drylands is about 51.7 million km2 or about 39.7% of ice-free lands (Reynolds and Stafford Smith 2002, 2 [42]; cf. Reynolds et al. 2007 [36]).
Unlike biomes, rangelands are not defined independently of land use, at least in the sense that areas converted to other uses and cover types cease to be rangelands. Approximately 35–50% of the world’s wetter, more fertile rangelands have been converted to crop agriculture, for example (Millennium Ecosystem Assessment 2005 [43]), including more than 96% of tall-grass prairie in the United States (Samson, Knopf, and Ostlie 1998 [44]). As a corollary, the rangelands that remain are, on average, more xeric and less fertile; in some contexts, rangelands and drylands are treated as near synonyms (Reynolds and Stafford Smith 2002 [42]). They also tend to be the lowest value lands (in money terms), where conversion to more intensive uses has been precluded either by biophysical constraints or by relative remoteness from infrastructure and development (Bestelmeyer et al. 2015 [45]). For this very reason, however, almost any other land use is likely to appear economically advantageous; today, land use-land cover change and fragmentation are widely considered to be the greatest threats facing most rangelands (Galvin et al. 2008 [46]). In effect, rangelands constitute a residual category, united by what they are not—namely, cultivated, forested, covered by ice, or built over (Sayre 2017 [47]).
Ellis and Ramankutty (2008 [2]) echo this notion of rangelands, if only by contrast with their other major anthrome types: urban, villages, croplands, and forested. However, they do not distinguish between pastures and rangelands, even though tame pastures are usually not considered rangelands. According to Grice and Hodgkinson (2002, 2 [48]) “rangelands occur in areas of relatively low rainfall or where winters are long and cold. The vegetation is always dominated by natural plant communities rather than by sown vegetation. Biotically, 34.9 million km2 (87.8%) of the lands categorized as rangelands by Ellis and Ramankutty (2008, WebTable 5 [2]) are found in grasslands, savannas, shrublands, tundra, and desert—areas conventionally deemed rangelands. Conversely, based on the absence of livestock, they excluded almost all of the tundra biome and about half of the desert biome, which others (e.g., Asner et al. 2004 [40]) classify as rangelands. Thus, the actual places classified as rangelands by Ellis and Ramankutty (2008 [2]) differ in part from those of standard accounts: they exclude 20.0 million km2 of tundra and desert areas without livestock (these made up more than half of Earth’s remaining ‘wildlands’), while they include 4.8 million km2 of non-rangelands, almost entirely in areas that others have classified as forested. Although the logic of each of these steps seems reasonable enough, and they partially offset each other in areal terms, the aggregate result contains two potentially significant flaws.
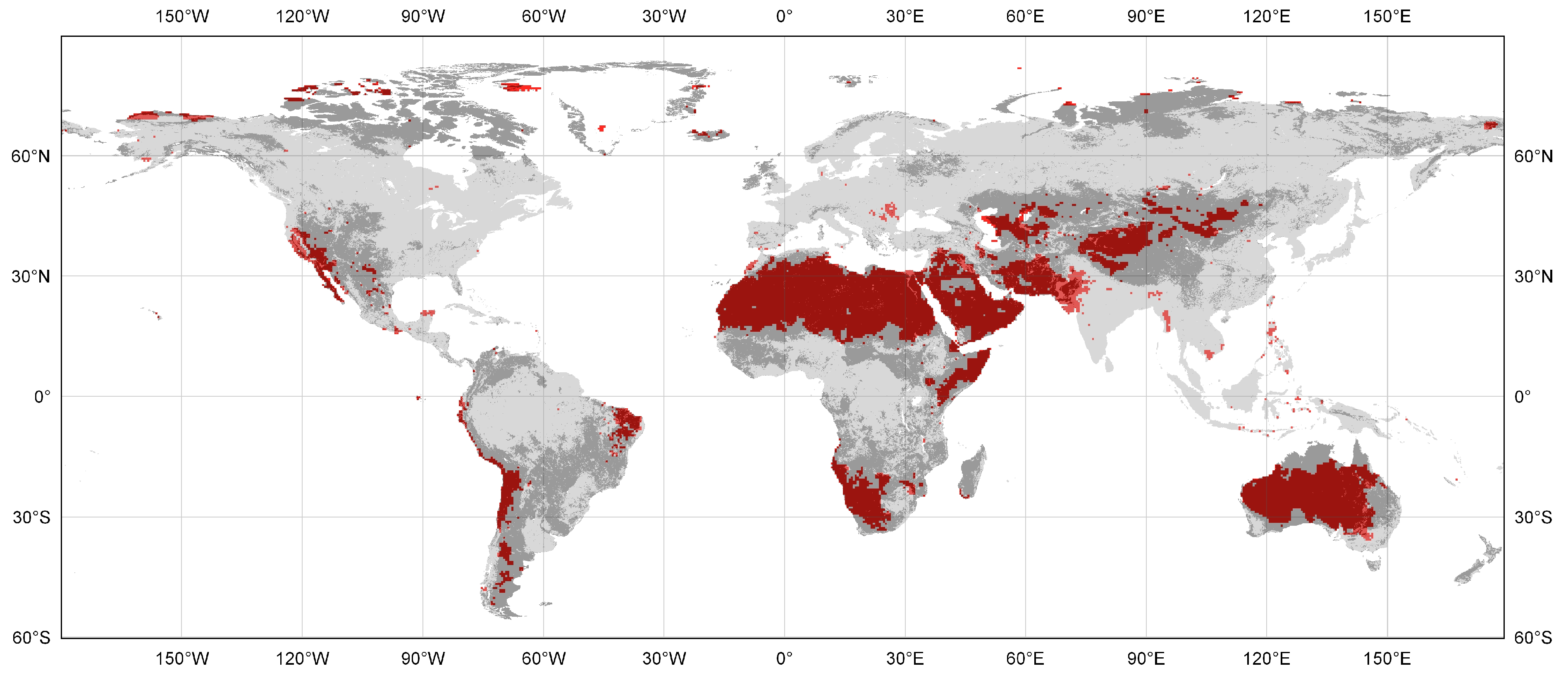
First, livestock grazing as a land use is converted into the definitive ecological criterion, as though it were a sufficient basis for distinguishing anthropogenic areas even in the complete absence of human inhabitants. This conflicts with the fact that a wide variety of organisms in rangelands—including invertebrates, birds, and small and large wild mammals—may exert ecological effects that resemble those of livestock (Vavra, Laycock, and Pieper 1994 [22]) and may be as great or greater in their ecosystemic significance (Whitford 2002 [26], McNaughton 1985 [24]). In other words, the presence of domesticated livestock per se may or may not be of unique or particular ecological significance. One must attend to contextual specifics, such as the presence or absence of grazing animals over evolutionary time scales (Milchunas and Lauenroth 1993 [30]). In some cases, an abiotic factor such as recurrent wildfire may exert an evolutionary effect analogous to grazing, creating a niche for a biotic factor such as livestock to occupy (Bond and Keeley 2005 [49], Fuhlendorf et al. 2008 [50]). Currently, the prevailing view among rangeland ecologists is that where the coefficient of variation of inter-annual precipitation is ≥0.33, ecosystem dynamics are controlled predominantly by abiotic factors, not biotic ones (whether livestock or other organisms) (Noy-Meir 1979 [51], Illius and O’Connor 1999 [52], Vetter 2005, [38]). Based on a meta-analysis of 58 published studies, von Wehrden et al. (2012, [39]) concluded that degradation is in fact absent from these areas. We estimate that 28.06% of all rangeland anthromes (residential, populated, and remote), and 42.78% of remote rangelands, as defined by Ellis and Ramankutty (2008 [2]), meet this criterion (Table 1 and Figure 1). It is interesting to note that highly variable precipitation (CoV ≥ 0.33) is even more prevalent in the areas that Ellis and Ramankutty (2008) designate as inhabited treeless and barren lands (60.2%) and wild treeless and barren lands (56.6%). For comparison, the figure for Earth’s ice-free lands as a whole is 19.9%. At a minimum, then, approximately 11.59 million km2 of these areas should not be deemed ecologically anthropogenic, even if livestock grazing occurs there.
Second, areas where humans have actively converted other vegetation types for grazing—by clearing forest and planting grasses, for example—are lumped together with areas where livestock consume naturally occurring plants. Most of these are presumably located within the 4.8 million km2 of rangelands that Ellis and Ramankutty (2008 [2]) found in biomes otherwise considered to be forested, and thus cannot represent more than 12.2% of global rangelands. However, they may bear disproportionate weight precisely because they seem obviously anthropogenic. By contrast, assessing the impact of people (or livestock) on the other 87.8% of rangelands is remarkably difficult. The effects of livestock grazing, like other disturbances, vary depending on timing, intensity, frequency and various contextual factors (Sayre 2001 [53]). It may or may not cause significant ecological changes; those changes may or may not represent degradation, and they may or may not be reversed if grazing ceases. Different studies have concluded that as little as 10–20% or as much as 80–90% of global drylands are degraded (Millennium Ecosystem Assessment 2005, 637 [43]). This divergence reflects not only limited, low-quality, or incommensurable data, but also disagreements about what constitutes degradation at all. In short, equating livestock grazing with anthropogenic impact raises thorny conceptual problems.
3. The Conceptual Basis
As mentioned above, “remote rangelands” for Ellis and Ramankutty (2008) have a residential human population density of zero; they were defined as “rangelands with inconsequential human populations” (2008, 442 Table 1 [2]). In their map, remote rangelands encompassed large portions of Australia, central Asia, southwestern Africa, western North America, and the southern cone of South America, as well as significant areas scattered across North Africa and the Middle East. If the human populations in such settings were “inconsequential”, then presumably it must have been their livestock whose effects were deemed significant enough to render these vast expanses anthromes, but the criteria behind this judgment were not elaborated. In two more recent papers, however, Ellis (2011 [4]) and his colleagues (Ellis et al., 2010 [3]) attempt to determine when the world’s biomes became anthromes, and these issues cannot be avoided.
Whereas scholars usually understand rangelands as a whole to have contracted over time due to conversion to croplands (Reid, Galvin, and Kruska 2008 [41]), the first follow-up paper portrays them as expanding dramatically. From 1700 to 2000, “the extent of rangelands increased rapidly in every century… As a result, the foremost global land-use change of the Industrial Revolution in terms of total area was the expansion of pastures from 3% of ice-free land in 1700 to 26% in 2000” (Ellis et al. 2010, 593 [3]). Among biomes, “grasslands, savannas and shrublands showed the greatest changes over time, with all of these experiencing >80% conversion to used anthromes from 1700 to 2000” (Ellis et al. 2010, 595-96 [3]). (It is worth noting that this passage continues: “Most of this was the result of converting both wildlands and seminatural anthromes to rangelands, though the conversion to croplands was substantial in grasslands (28%), savanna (23%) and dense shrublands (19%).”) Thus, circa 1700, many of the areas listed above as remote rangelands were not rangelands at all but “treeless and barren lands”, “without human populations or substantial land use” (Ellis et al. 2010, 591 Table 1 [3]) and therefore “wild”, corresponding to the “barren wildlands” category of the 2008 paper. Or they were “seminatural” anthromes that were (no longer) “wild” but also not (yet) “used”, and thus not rangelands. Such areas included the Sahel and Horn of Africa and large parts of central Asia, where pastoralists have grazed livestock since long before 1700. This in-between, seminatural category does not appear in Ellis and Ramankutty (2008 [2]). It seems to hinge on human population density and land use intensity, but specifics are lacking in the 2010 paper.
The second follow-up paper, which compared model runs for the period 6000 BCE to 2000 CE based on two different land use and population datasets, confronted these issues more directly. Ellis (2011; pp. 1020–1021 [4]) acknowledges the variable and uneven degree of human modification of rangelands. “Domesticated grazing animals are typically adapted to grasslands and savannas, so their ecological alteration of these biomes tends to be less novel, except when stocking rates are very high… In savannas, shrublands and grasslands, rangeland development may produce only minor alteration of land cover and NPP, depending on the extent and intensity of land clearing and grazing.” He proceeds to argue that livestock grazing tends nonetheless to favor exotic species invasions and altered fire regimes, suggesting that these, too, constitute grounds for classification as anthropogenic. However, the reference cited for plant invasions (DiTomaso 2000 [54]) is based solely on the case of US rangelands west of the Rocky Mountains—that is, areas without an evolutionary history of large ungulate grazing. In areas with such a history—such as the US Great Plains, central Asia, and sub-Saharan Africa—grazing effects can be quite different (McNaughton 1985 [24]; Milchunas and Lauenroth 1993 [30]). As for altered fire regimes, we have at this point come full circle back to Sauer (1950 [16]), who argued that grasslands have been anthromes for some 200,000 years. As Stephen Pyne (1997 [55]) has demonstrated in abundant detail, the idea of Homo sapiens without the alteration of fire regimes is nonsensical, not only in rangelands but in any terrestrial setting outside Antarctica.
By the end of the 2011 article, Ellis seems to concede that making sense of rangelands in terms of the anthromes thesis is nearly impossible. The two models of historical land use change that he compares “yield different trends in anthrome development”, in particular radically different estimates of when anthromes came to dominate (Ellis 2011; p. 1025 [4]). By one model, the critical shift occurred only in the past 2–3 centuries, whereas the other dates it to between 1000 and 0 BCE. And whereas the least conservative measure of anthrome development indicates 75% of terrestrial areas transformed by human activities in 2000—the conclusion of Ellis and Ramankutty (2008 [2])—the most conservative measure yields a result of just 29%. Crucially, this last calculation includes only those rangelands developed by clearing woodlands. The delineation of anthromes, he suggests, should be limited to areas “in which pre-existing ecosystem forms and processes have been shifted outside their native range and novel anthropogenic ecological processes predominate” (Ellis 2011; p. 1025 [4]). Significantly, Ellis (2011; p. 1027) closes by questioning “whether indisputable quantitative measurements of anthropogenic transformation could be made across the terrestrial biosphere”—precisely what Ellis and Ramankutty (2008) purportedly achieved—and he proposes, rather philosophically, that “the most important repercussions of embracing our anthropogenic biosphere will come from changes in social, political, and economic points of view.” It is to these issues that we now turn.
4. Conclusions: The Importance of Mobility, Property, & Variability
Ellis and Ramankutty’s (2008 [2]) classification of nearly 40 million km2 of rangelands as anthropogenic biomes can be challenged on both empirical and conceptual grounds. In characterizing rangelands as anthromes—as though they were categorically identical and ecologically equivalent with urban or cultivated lands—Ellis and Ramankutty (2008) overstate the degree to which rangelands, and by extension the Earth as a whole, have been transformed by human action. At a minimum, the roughly 11.6 million km2 with highly variable precipitation, plus a large but indefinite area with long evolutionary histories of grazing, should be deducted from their estimate. This does not in any way detract from the significance of human impacts on the rest of Earth’s terrestrial areas, for which Ellis and Ramankutty’s (2008) paper provides a novel metric. However, the conceptual issues raised by rangelands pose challenges for the anthromes thesis as a whole.
At the 1955 conference Man’s Role in Changing the Face of the Earth, F. Fraser Darling noted that movement and mobility were critical to the material success of pastoralists and the sustainable use of their arid and semi-arid environments. “[W]here pastoralism over a long period has not damaged the habitat, it is markedly nomadic in character” (Darling 1956; p. 781 [1]). Sedentary, commercial pastoralism, by contrast, had decidedly different outcomes: “Vegetational climaxes have been broken insidiously rather than by some grand traumatic act, and, just as cultivation of food plants involves setting back ecological succession to a primary stage, pastoralism deflects succession to the xeric, a profound and dangerous change” (Darling 1956; p. 781 [1]). Nomadic pastoralism and civilization were at odds: “A world of sedentary cultures impinges always on nomadic territory not held in fee simple, and the nomadic society is brittle. What is to be the answer?” (Darling 1956; p. 787 [1]).
This question is more urgent than ever today. Beginning in colonial times and accelerating since Darling spoke, indigenous rangeland management systems that hinge on various forms of mobility have been systematically suppressed and marginalized. Accused of overgrazing, desertification, and land degradation, pastoralists across Africa, Asia, the Middle East, and the Americas have been compelled to become more sedentary, to reduce or eliminate their herds, and often dispossessed of their lands (Awad 1962 [56], Bedrani 1983 [57], Ballais 1994 [58], Benjaminsen 2016 [59], Chatty 2006 [60], Lavauden 1927 [61], Davis 2007 [62], Jacoby 2001 [63], Monteil 1959 [64], Weisiger 2009 [65], Whyte 1956 [66]). These actions have been taken on the basis of “scientific” theories forged first in the context of colonial forestry (Davis 2016 [67]) and subsequently in the western US, strongly influenced by Clements’s (1916 [15]) theory of plant succession, equilibrium, and climatic climax. That livestock were always the cause of rangeland degradation, and that reducing or removing livestock would always heal or restore the damage, became hegemonic truths, institutionalized in laws, policies, textbooks and development programs (Sayre 2017 [46]). However, a large body of historical and ecological evidence now refutes both of these claims and suggests instead that in many rangelands, the ecological effects of pastoralism are insignificant (relative to abiotic drivers) and/or indistinguishable from those of wild or natural herbivores (Behnke, Scoones, and Kerven 1993 [33]; Behnke and Mortimore 2016 [68]; Kratli 2015 [35]). The land degradation that does exist in these rangelands more often results from the very programs that curtail mobility, ‘rationalize’ land tenure, and intensify commercial livestock production—a phenomenon Davis (2016 [67]) terms “accumulation by desertification.”
In light of this history, there are at least two reasons to be concerned about an exaggerated anthromes thesis. One is the risk that it will be used—especially but not only in rangelands—to legitimate socially unjust and ecologically counterproductive policies and programs, just as the desertification thesis was used to sedentarize and disenfranchise countless indigenous peoples around the world beginning in the nineteenth century (Davis 2016 [67]). The notion that livestock grazing alone—even without any resident human inhabitants—necessarily renders any and all rangelands anthropogenic, dominated by human uses to such a degree that “non-human nature” is insignificant, recapitulates the assumptions of Clementsian ecological theory and Euro-colonial forestry. Similarly, the old dogma that livestock removal necessarily triggers restorative succession to climax lurks just beneath the surface of the “rangeland anthrome” designation, and it is easy to imagine the idea of a “used planet” (Ellis et al. 2013 [5]) being invoked to rationalize crisis measures, such as removing people in order to turn an anthropogenic biome back into ‘wildland’. If the history of rangelands, the largest anthrome, is so fraught with misunderstandings and failed attempts at ‘improvement’, we need to proceed very cautiously indeed with applying the anthromes thesis to three quarters of the planet. It should be noted, in this connection, that Clements is also credited as the founder of the concept of biomes itself.
The second risk is that opportunities may be missed to understand human-environment relations in more sustainable and socially just ways. Rangelands provide a unique lens through which to analyze the idea of anthropogenic biomes, not only because they are the largest single type of land on Earth, but also because they are environments where human use does not necessarily cause significant, ecologically disruptive change. Rangelands challenge us to envision ways of living in the world that adapt to and fit within the constraints of complex, variable and often extreme ecological conditions (Corbett 2005 [69]). This requires more than simply new or better scientific understandings of ecosystems and their relative degrees of human alteration. As Darling’s 1955 lecture demonstrates—presciently and almost uniquely—Clementsian ideals of climax and succession are intimately bound up with ideas about property and civilization. Rangelands will not be conserved by protecting them from pastoralists, but by proactively supporting pastoral ways of living there, including forms of land tenure and inhabitation other than sedentary private property. Rather than nostalgic, romantic programs that would pretend to resurrect past conditions, such systems could embrace the latest technological innovations, as many remaining pastoral nomads already do (Roe, Huntsinger, and Labnow 1998 [70]).
A final and even more fundamental challenge is to consider, with Sauer, the notion that rangelands have always been anthropogenic, which is to say that the human/non-human distinction is a fallacious construct. If confronting this challenge is one objective of the anthromes thesis—as we believe may well be the case—then another theoretical framework is needed, one that does not reproduce the very dualism it sets out to transcend. Here, the lesson from rangelands is that where notable ecological changes have occurred, they have usually been the result of transformations in social, political, and economic relations among people. The primary driver of environmental change in rangelands over the last several centuries has been a political and economic system that prioritizes private property, sedentarism, and the generation of profit, and this, rather than “human use” in the abstract, is the relevant indicator and ultimate cause of all anthropogenic biomes (Sayre 2012 [71]).
Acknowledgments
We are grateful to Erle Ellis and the editors for the invitation to contribute to this special issue, and to three anonymous reviewers for their suggestions. Open access publication fees were covered from research funds provided to NFS by the University of California, Berkeley. B.B. and J.C.W. were supported by appropriated funds to the USDA-ARS.
Author Contributions
N.F.S. conceived the paper, recruited the co-authors, and wrote approximately 80% of the first submitted version of the paper. D.K.D. wrote the remaining 20%, and B.B. reviewed and edited the first version. B.B., D.K.D., and N.F.S. contributed equally to the formulating the argument in outline form prior to writing. B.B. and J.C.W. produced the map and did the underlying analysis from which it and Table 1 were produced. D.K.D. and N.F.S. contributed equally to revising the paper following peer review, in dialogue with B.B..
Conflicts of Interest
The authors declare no conflict of interest. Their institutions had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Darling, F.F. Man’s Ecological Dominance through Domesticated Animals on Wild Lands. In Man’s Role in Changing the Face of the Earth; Thomas, W.L., Jr., Ed.; University of Chicago Press: Chicago, IL, USA; London, UK, 1956; Volume 2, pp. 778–787. [Google Scholar]
- Ellis, E.C.; Ramankutty, N. Putting people in the map: Anthropogenic biomes of the world. Front. Ecol. Environ. 2008, 6, 439–447. [Google Scholar] [CrossRef]
- Ellis, E.C.; Goldewijk, K.K.; Siebert, S.; Lightman, D.; Ramankutty, N. Anthropogenic transformation of the biomes, 1700 to 2000. Glob. Ecol. Biogeogr. 2010, 19, 589–606. [Google Scholar] [CrossRef]
- Ellis, E.C. Anthropogenic transformation of the terrestrial biosphere. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2011, 369, 1010–1035. [Google Scholar] [CrossRef] [PubMed]
- Ellis, E.C.; Kaplan, J.O.; Fuller, D.Q.; Vavrus, S.; Goldewijk, K.K.; Verburg, P.H. Used planet: A global history. Proc. Natl. Acad. Sci. USA 2013, 110, 7978–7985. [Google Scholar] [CrossRef] [PubMed]
- Ellis, E.C. Ecology in an anthropogenic biosphere. Ecol. Monogr. 2015, 85, 287–331. [Google Scholar] [CrossRef]
- Marsh, G.P. Man and Nature; Charles Scribner: New York, NY, USA, 1865. [Google Scholar]
- Glacken, C. Traces on the Rhodian Shore: Nature and Culture in Western Thought from Ancient Times to the End of the Eighteenth Century; University of California Press: Berkeley, CA, USA, 1967. [Google Scholar]
- Bennett, H.H.; Chapline, W.R. Soil Erosion: A National Menace; Circular no. 33; USDA: Washington, DC, USA, 1928.
- Sears, P.B. Deserts on the March; University of Oklahoma Press: Norman, OK, USA, 1935. [Google Scholar]
- Vogt, W. Road to Survival; W. Sloane: New York, NY, USA, 1948. [Google Scholar]
- Hardin, G. The tragedy of the commons. Science 1968, 162, 1243–1248. [Google Scholar] [CrossRef] [PubMed]
- Meadows, D.H.; Meadows, D.L.; Randers, J.; Behrens, W.W., III. The Limits to Growth; Universe Books: New York, NY, USA, 1972. [Google Scholar]
- Tansley, A.G. Practical Plant Ecology: A Guide for Beginners in Field Study of Plant Communities; Allen & Unwin: London, UK, 1923. [Google Scholar]
- Clements, F.E. Plant Succession: An Analysis of the Development of Vegetation; Publication No. 242; Carnegie Institution of Washington: Washington, DC, USA, 1916. [Google Scholar]
- Sauer, C.O. Grassland climax, fire, and man. J. Range Manag. 1950, 3, 16–21. [Google Scholar] [CrossRef]
- Thomas, W.L., Jr. (Ed.) Man’s Role in Changing the Face of the Earth; University of Chicago Press: Chicago, IL, USA; London, UK, 1956; Volume 2. [Google Scholar]
- Crutzen, P.J.; Stoermer, E.F. The ‘Anthropocene’. IGBP Newsl. 2000, 41, 17–18. [Google Scholar]
- Vitousek, P.M.; Mooney, H.A.; Lubchenco, J.; Melillo, J.M. Human domination of Earth’s ecosystems. Science 1997, 277, 494–499. [Google Scholar] [CrossRef]
- Holdren, J.P.; Ehrlich, P.R. Human population and the global environment: Population growth, rising per capita material consumption, and disruptive technologies have made civilization a global ecological force. Am. Sci. 1974, 62, 282–292. [Google Scholar]
- Vavra, M.; Laycock, W.A.; Pieper, R.D. Ecological Implications of Livestock Herbivory in the West; Society for Range Management: Denver, CO, USA, 1994. [Google Scholar]
- Nicholson, S.E. Dryland Climatology; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Herrmann, S.; Hutchinson, C.F. The scientific basis: Linkages between land degradation, drought and desertification. In Governing Global Desertification: Linking Environmental Degradation, Poverty and Participation; Johnson, P., Mayrand, K., Paquin, M., Eds.; Ashgate Publishing: Hampshire, UK, 2006. [Google Scholar]
- McNaughton, S. J. Ecology of a grazing ecosystem: The Serengeti. Ecol. Monogr. 1985, 55, 259–294. [Google Scholar] [CrossRef]
- Mortimore, M. Dryland Opportunities: A New Paradigm for People, Ecosystems and Development; IUCN: Gland, Switzerland, 2009. [Google Scholar]
- Whitford, W.G. Ecology of Desert Systems; Academic Press: London, UK; San Diego, CA, USA, 2002. [Google Scholar]
- Ash, A.J.; Stafford Smith, D.M.; Abel, N. Land degradation and secondary production in semi-arid and arid grazing systems. In Global Desertification: Do Humans Cause Deserts? Reynolds, J.F., Smith, S., Eds.; Dahlem University Press: Berlin, Germany, 2002; pp. 111–134. [Google Scholar]
- Behnke, R.; Döll, P.; Ellis, J.; Harou, P.; Lynam, M.A.T.; Nicholson, S.; Thornes, J. Responding to desertification at the national scale. In Global Desertification: Do Humans Cause Deserts? Reynolds, J.F., Smith, S., Eds.; Dahlem University Press: Berlin, Germany, 2002; pp. 357–385. [Google Scholar]
- Gillson, L.; Hoffman, M.T. Rangeland ecology in a changing world. Science 2007, 315, 53–54. [Google Scholar] [CrossRef] [PubMed]
- Milchunas, D.G.; Lauenroth, W.K. Quantitative effects of grazing on vegetation and soils over a global range of environments. Ecol. Monogr. 1993, 63, 327–366. [Google Scholar] [CrossRef]
- Sprugel, D. Disturbance, equilibrium, and environmental variability: What is ‘natural’ vegetation in a changing environment? Biol. Conserv. 1991, 58, 1–18. [Google Scholar] [CrossRef]
- Véron, S.R.; Paruelo, J.M.; Osterheld, M. Assessing desertification. J. Arid Environ. 2006, 66, 751–763. [Google Scholar] [CrossRef]
- Behnke, R.H.; Scoones, I.; Kerven, C. (Eds.) Range Ecology at Disequilibrium: New Modes of Natural Variability and Pastoral Adaptation in African Savannas; Overseas Development Institute: London, UK, 1993. [Google Scholar]
- Ellis, J.E.; Swift, D.M. Stability of African pastoral ecosystems: Alternate paradigms and implications for development. J. Range Manag. 1988, 41, 450–459. [Google Scholar] [CrossRef]
- Kratli, S. Valuing Variability: New Perspectives on Climate-Resilient Drylands Development; International Institute for Environment and Development: London, UK, 2015. [Google Scholar]
- Reynolds, J.F.; Smith, D.M.S.; Lambin, E.F.; Turner, B.L., II; Mortimore, M.; Batterbury, S.P.J.; Downing, T.E.; Dowlatabadi, H.; Fernández, R.J.; Herrick, J.E.; et al. Global desertification: Building a science for dryland development. Science 2007, 316, 847–851. [Google Scholar] [CrossRef] [PubMed]
- Scoones, I. (Ed.) Living with Uncertainty: New Directions in Pastoral Development in Africa; Intermediate Technology Publications: London, UK, 1995. [Google Scholar]
- Vetter, S. Rangelands at equilibrium and non-equilibrium: Recent developments in the debate. J. Arid Environ. 2005, 62, 321–341. [Google Scholar] [CrossRef]
- Von Wehrden, H.; Hanspach, J.; Kaczensky, P.; Fischer, J.; Wesche, K. Global assessment of the non-equilibium concept in rangelands. Ecol. Appl. 2012, 22, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Asner, G.P.; Elmore, A.J.; Olander, L.P.; Martin, R.E.; Harris, A.T. Grazing systems, ecosystem responses, and global change. Ann. Rev. Environ. Resour. 2004, 29, 261–299. [Google Scholar] [CrossRef]
- Reid, R.S.; Galvin, K.A.; Kruska, R.S. Global Significance of Extensive Grazing Lands and Pastoral Societies: An Introduction. In Fragmentation in Semi-Arid and Arid Landscapes: Consequences for Human and Natural Systems; Galvin, K.A., Reid, R.S., Behnke, R.H., Hobbs, N.T., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 1–24. [Google Scholar]
- Reynolds, J.F.; Smith, D.M.S. (Eds.) Global Desertification: Do Humans Cause Deserts? Dahlem University Press: Berlin, Germany, 2002. [Google Scholar]
- Millennium Ecosystem Assessment (Ed.) Ecosystems and Human Well-Being; Island Press: Washington, DC, USA, 2005; Volume 5. [Google Scholar]
- Samson, F.B.; Knopf, F.L.; Ostlie, W.R. Grasslands. In Status and Trends of the Nation’s Biological Resources; Mac, M.J., Opler, P.A., Puckett Haecker, C.E., Doran, P.D., Eds.; US Department of Interior, US Geological Survey: Reston, VA, USA, 1998; Volume 2, pp. 437–472. [Google Scholar]
- Bestelmeyer, B.T.; Okin, G.S.; Duniway, M.C.; Archer, S.R.; Sayre, N.F.; Williamson, J.C.; Herrick, J.E. Desertification, land use, and the transformation of global drylands. Front. Ecol. Environ. 2015, 13, 28–36. [Google Scholar] [CrossRef]
- Galvin, K.A.; Reid, R.S.; Behnke, R.H.; Hobbs, N.T. (Eds.) Fragmentation in Semi-Arid and Arid Landscapes: Consequences for Human and Natural Systems; Springer: Dordrecht, The Netherlands, 2008. [Google Scholar]
- Sayre, N. The Politics of Scale: A History of Rangeland Science; University of Chicago Press: Chicago, IL, USA; London, UK, 2017. [Google Scholar]
- Grice, A.C.; Hodgkinson, K.C. Global Rangelands: Progress and Prospects; Centre for Agriculture and Bioscience International: Wallingford, UK, 2002. [Google Scholar]
- Bond, W.J.; Keeley, J.E. Fire as global ‘herbivore’: The ecology and evolution of flammable ecosystems. Trends Ecol. Environ. 2005, 20, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Fuhlendorf, S.D.; Engle, D.M.; Kerby, J.; Hamilton, R. Pyric herbivory: Rewilding landscapes through the recoupling of fire and grazing. Conserv. Biol. 2008, 23, 588–598. [Google Scholar] [CrossRef] [PubMed]
- Noy-Meir, I. Structure and function of desert ecosystems. Israel J. Bot. 1979, 28, 1–19. [Google Scholar]
- Illius, A.W.; O’Connor, T.G. On the relevance of nonequilibrium concepts to arid and semiarid grazing systems. Ecol. Appl. 1999, 9, 798–813. [Google Scholar] [CrossRef]
- Sayre, N.F. The New Ranch Handbook: A Guide to Restoring Western Rangelands; Quivira Coalition: Santa Fe, NM, USA, 2001. [Google Scholar]
- DiTomaso, J.M. Invasive weeds in rangelands: Species, impacts, and management. Weed Sci. 2000, 48, 255–265. [Google Scholar] [CrossRef]
- Pyne, S.J. World Fire: The Culture of Fire on Earth; University of Washington Press: Seattle, WA, USA, 1997. [Google Scholar]
- Awad, M. Nomadism in the Arab lands of the Middle East. In The Problems of the Arid Zone: Proceedings of the Paris Symposium; UNESCO: Paris, France, 1962; pp. 325–339. [Google Scholar]
- Bedrani, S. Going slow with pastoral cooperatives. Ceres 1983, 16, 16–21. [Google Scholar]
- Ballais, J.-L. Aeolian activity, desertification and the ‘Green Dam’ in the Ziban Range, Algeria. In Environmental Change in Drylands: Biogeographical and Geomorphological Perspectives; Millington, A.C., Pye, K., Eds.; Wiley: New York, NY, USA, 1994; pp. 177–198. [Google Scholar]
- Benjaminsen, T.A. Does climate change lead to conflicts in the Sahel? In The End of Desertification: Disputing Environmental Change in the Drylands; Behnke, R., Mortimore, M., Eds.; Springer: Dordrecht, The Netherlands, 2016. [Google Scholar]
- Chatty, D. (Ed.) Nomadic Societies in the Middle East and North Africa Entering the Twenty-first Century; Brill: Leiden, The Netherlands, 2006. [Google Scholar]
- Lavauden, L. Les Forêts du Sahara. Revue des Eaux et Forêts 1927, 65, 265–277, 329–341. [Google Scholar]
- Davis, D.K. Resurrecting the Granary of Rome: Environmental History and French Colonial Expansion in North Africa; Ohio University Press: Athens, OH, USA, 2007. [Google Scholar]
- Jacoby, K. Crimes against Nature: Squatters, Poachers, Thieves, and the Hidden History of American Conservation; University of California Press: Berkeley, CA, USA, 2001. [Google Scholar]
- Monteil, V. The evolution and settling of the Nomads of the Sahara. Int. Soc. Sci. J. 1959, 11, 572–585. [Google Scholar]
- Weisiger, M.L. Dreaming of Sheep in Navajo Country; University of Washington Press: Seattle, WA, USA, 2009. [Google Scholar]
- Whyte, R.O. Grazing Resources. In The Future of Arid Lands: Papers and Recommendations from the International Arid Lands; White, G.F., Ed.; American Association for the Advancement of Science: Washington, DC, USA, 1956; pp. 179–188. [Google Scholar]
- Davis, D.K. The Arid Lands: History, Power, Knowledge; The MIT University Press: Cambridge, MA, USA, 2016. [Google Scholar]
- Behnke, R.; Mortimore, M. (Eds.) The End of Desertification: Disputing Environmental Change in the Drylands; Springer: Dordrecht, The Netherlands, 2016. [Google Scholar]
- Corbett, J. Sanctuary for All Life: The Cowballah of Jim Corbett; Howling Dog Press: Berthoud, CO, USA, 2005. [Google Scholar]
- Roe, E.; Huntsinger, L.; Labnow, K. High Reliability Pastoralism. J. Arid Environ. 2005, 39, 39–55. [Google Scholar] [CrossRef]
- Sayre, N.F. The politics of the anthropogenic. Ann. Rev. Anthropol. 2012, 41, 57–70. [Google Scholar] [CrossRef]
Figure 1.
Map of global land area with an inter-annual precipitation coefficient of variation ≥33% (red), based on data from 1970 through 1999 (University of Delaware global gridded monthly precipitation dataset (http://www.esrl.noaa.gov/psd/)). Greyed background indicates rangeland and nonforested wildland anthromes (residential, populated, and remote rangelands, inhabited treeless and barren lands, and wild treeless and barren lands; http://ecotope.org/anthromes/v2/maps/).
Figure 1.
Map of global land area with an inter-annual precipitation coefficient of variation ≥33% (red), based on data from 1970 through 1999 (University of Delaware global gridded monthly precipitation dataset (http://www.esrl.noaa.gov/psd/)). Greyed background indicates rangeland and nonforested wildland anthromes (residential, populated, and remote rangelands, inhabited treeless and barren lands, and wild treeless and barren lands; http://ecotope.org/anthromes/v2/maps/).

{kind=link}
{kind=link}
Table 1.
Proportions of rangeland anthromes (Ellis and Ramankutty 2008 [2]) where the coefficient of variation (CV) of inter-annual precipitation is ≥0.33, based on the University of Delaware global gridded monthly precipitation dataset (http://www.esrl.noaa.gov/psd/).
Table 1.
Proportions of rangeland anthromes (Ellis and Ramankutty 2008 [2]) where the coefficient of variation (CV) of inter-annual precipitation is ≥0.33, based on the University of Delaware global gridded monthly precipitation dataset (http://www.esrl.noaa.gov/psd/).
| Anthrome | Total Area (km2) | Area CV ≥ 0.33 (km2) | Area CV ≥ 0.33 (%) |
|---|---|---|---|
| Residential rangelands | 7,911,700 | 836,679 | 10.58 |
| Populated rangelands | 13,757,100 | 2,354,100 | 17.11 |
| Remote rangelands | 19,623,900 | 8,394,510 | 42.78 |
| Totals | 41,292,700 | 11,585,289 | 28.06 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sayre, N.F.; Davis, D.K.; Bestelmeyer, B.; Williamson, J.C. Rangelands: Where Anthromes Meet Their Limits. Land 2017, 6, 31. https://doi.org/10.3390/land6020031
AMA Style
Sayre NF, Davis DK, Bestelmeyer B, Williamson JC. Rangelands: Where Anthromes Meet Their Limits. Land. 2017; 6(2):31. https://doi.org/10.3390/land6020031
Chicago/Turabian StyleSayre, Nathan F., Diana K. Davis, Brandon Bestelmeyer, and Jeb C. Williamson. 2017. "Rangelands: Where Anthromes Meet Their Limits" Land 6, no. 2: 31. https://doi.org/10.3390/land6020031
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.