Silicon-Rich, Iron Oxide Microtubular Sheath Produced by an Iron-Oxidizing Bacterium, Leptothrix sp. Strain OUMS1, in Culture

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Section
2.1. Culture Conditions
2.2. Transmission Electron Microscopy (TEM) Measurements
2.3. Qualitative and Quantitative Energy Dispersive X-Ray (EDX) Analysis of Si, Fe, O, and P in Sheaths
2.4. X-Ray Diffractometry (XRD) Measurements
3. Results and Discussion
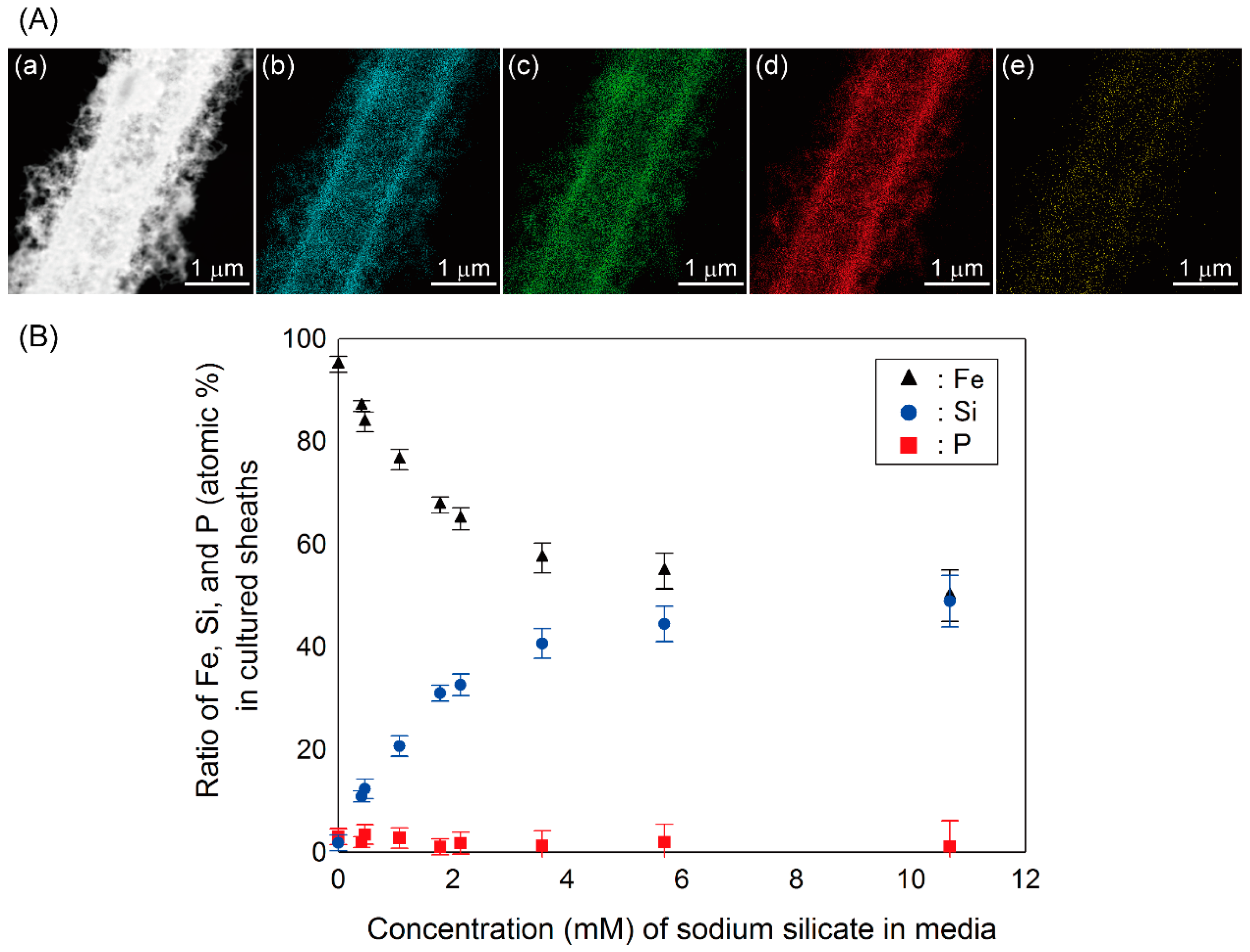
3.1. Morphology, Size and Electron Diffraction (ED)-Results of Sheaths


3.2. Elemental Analysis and Ratios of Sheaths

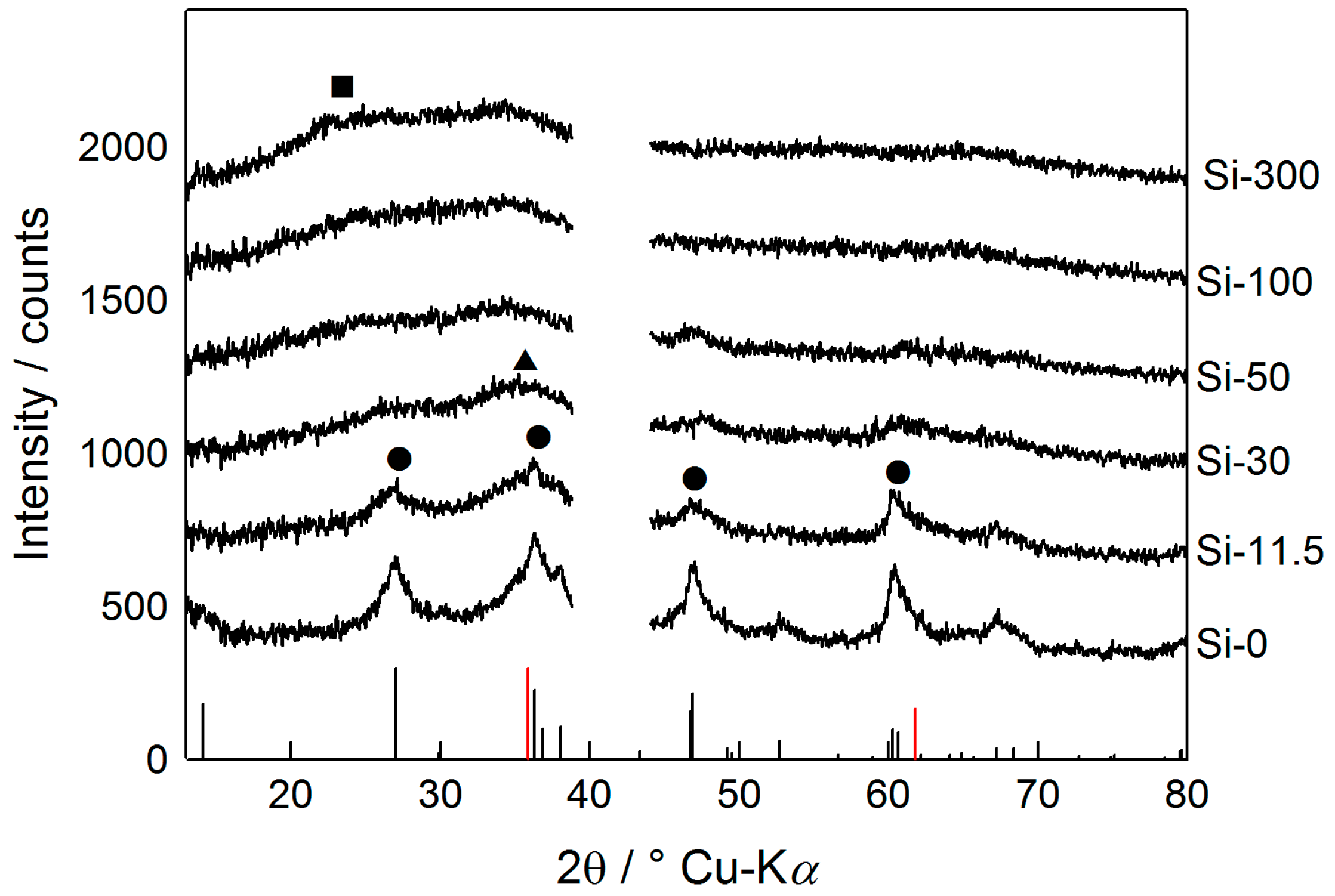
3.3. ED/XRD-Results and Crystallinity of Sheaths

4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ferris, F.G.; Fyfe, W.S.; Beveridge, T.J. Bacteria as nucleation sites for authigenic minerals in a metal-contaminated lake sediment. Chem. Geol. 1987, 63, 225–232. [Google Scholar] [CrossRef]
- Ghiorse, W.C.; Hirsch, P. An ultrastructural study of iron and manganese deposition associated with extracellular polymers of pedomicrobium-like budding bacteria. Arch. Microbiol. 1979, 123, 213–226. [Google Scholar] [CrossRef]
- Van Veen, W.L.; Mulder, E.G.; Deinema, M.H. The Sphaerotilus-Leptothrix group of bacteria. Microbiol. Rev. 1978, 42, 329–356. [Google Scholar]
- Ghiorse, W.C. Biology of iron- and manganese-depositing bacteria. Annu. Rev. Microbiol. 1984, 38, 515–550. [Google Scholar] [CrossRef]
- Emerson, D.; Fleming, E.J.; McBeth, J.M. Iron-oxidizing bacteria: An environmental and genomic perspective. Annu. Rev. Microbiol. 2010, 64, 561–583. [Google Scholar] [CrossRef]
- Fleming, E.J.; Langdon, A.E.; Martinez-Garcia, M.; Stepanauskas, R.; Poulton, N.J.; Masland, E.D.P.; Emerson, D. What’s new is old: Resolving the identity of Leptothrix ochracea using single cell genomics, pyrosequencing and FISH. PLoS One 2011, 6, 1–16. [Google Scholar]
- Spring, S. The genera Leptothrix and Sphaerotilus. Prokaryotes 2006, 5, 758–777. [Google Scholar] [CrossRef]
- Sawayama, M.; Suzuki, T.; Hashimoto, H.; Kasai, T.; Furutani, M.; Miyata, N.; Kunoh, H.; Takada, J. Isolation of a Leptothrix strain, OUMS1, from ocherous deposits in groundwate. Curr. Microbiol. 2011, 63, 173–180. [Google Scholar] [CrossRef]
- Furutani, M.; Suzuki, T.; Ishihara, H.; Hashimoto, H.; Kunoh, H.; Takada, J. Initial assemblage of bacterial saccharic fibrils and element deposition to form an immature sheath in cultured Leptothrix sp. strain OUMS1. Minerals 2011, 1, 157–166. [Google Scholar] [CrossRef]
- Hashimoto, H.; Fujii, T.; Kohara, S.; Asaoka, H.; Kusano, Y.; Ikeda, Y.; Nakanishi, M.; Benino, Y.; Nanba, T.; Takada, J. Amorphous structure of iron oxide of bacterial origin. Mater. Chem. Phys. 2012, 137, 571–575. [Google Scholar] [CrossRef]
- Furutani, M.; Suzuki, T.; Ishihara, H.; Hashimoto, H.; Kunoh, H.; Takada, J. Assemblage of bacterial saccharic microfibrils in sheath skeleton formed by cultured Leptothrix sp. strain OUMS1. J. Mar. Sci. Res. Dev. 2011, 5. [Google Scholar] [CrossRef]
- Suzuki, T.; Hashimoto, H.; Ishihara, H.; Kasai, T.; Kunoh, H.; Takada, J. Structural and spatial associations between Fe, O, and C in the network structure of the Leptothrix ochracea sheath surface. Appl. Environ. Microbiol. 2011, 77, 7873–7875. [Google Scholar]
- Chan, S.C.; Fakra, S.C.; Emerson, D.; Fleming, E.J.; Edwards, K.J. Lithotrophic iron-oxidizing bacteria produce organic stalks to control mineral growth: Implications for biosignature formation. ISME J. 2011, 5, 717–727. [Google Scholar] [CrossRef]
- Ferris, F.G.; Schultze, S.; Witten, T.C.; Fyfe, W.S.; Beveridge, T.J. Metal interactions with microbial biofilms in acidic and neutral pH environments. Appl. Environ. Microbiol. 1989, 55, 1249–1257. [Google Scholar]
- Hallberg, R.; Ferris, F.G. Biomineralization by Gallionella. Geomicrobiol. J. 2004, 21, 325–330. [Google Scholar] [CrossRef]
- Kennedy, C.B.; Scott, S.D.; Ferris, F.G. Characterization of bacteriogenic iron oxide deposits from axial volcano, Juan de Fuca Ridge, Northeast Pacific Ocean. Geomicrobiol. J. 2003, 20, 199–214. [Google Scholar] [CrossRef]
- Suzuki, T.; Ishihara, H.; Furutani, M.; Shiraishi, T.; Kunoh, H.; Takada, J. A novel method for culturing of Leptothrix sp. strain OUMS1 in natural conditions. Minerals 2012, 2, 118–128. [Google Scholar] [CrossRef]
- Hashimoto, H.; Hashimoto, H.; Kobayashi, G.; Sakuma, R.; Fujii, T.; Hayashi, N.; Kanno, R.; Takano, M.; Takada, J. Bacterial nanometric amorphous Fe-based oxide: A potential lithium-ion battery anode material. ACS Appl. Mater. Interfaces 2014, 6, 5374–5378. [Google Scholar] [CrossRef]
- Ema, T.; Miyazaki, Y.; Kozuki, I.; Sakai, T.; Hashimoto, H.; Takada, J. Highly active lipase immobilized on biogenous iron oxide via an organic bridging group: The dramatic effect of the immobilization support on enzymatic function. Green Chem. 2011, 13, 3187–3195. [Google Scholar] [CrossRef]
- Ema, T.; Miyazaki, Y.; Taniguchi, T.; Takada, J. Robust porphyrin catalysts immobilized on biogenous iron oxide for the repetitive conversions of epoxides and CO2 into cyclic carbonates. Green Chem. 2013, 15, 2485–2492. [Google Scholar]
- Mandai, K.; Korenaga, T.; Ema, T.; Sakai, T.; Furutani, M.; Hashimoto, H.; Takada, J. Biogenous iron oxide-immobilized palladium catalyst for the solvent-free Suzuki–Miyaura coupling reaction. Tetrahedron Lett. 2012, 53, 329–332. [Google Scholar] [CrossRef]
- Hashimoto, H.; Asaoka, H.; Nakano, T.; Kusano, Y.; Ishihara, H.; Ikeda, Y.; Nakanishi, M.; Fujii, T.; Yokoyama, T.; Horiishi, N.; Nanba, T.; Takada, J. Preparation, microstructue, and color tone of microtubule material composed of hematite/amorphous-silicate nonocomposite from iron oxide of bacterial origin. Dye. Pigment. 2012, 95, 639–643. [Google Scholar] [CrossRef]
- Rogers, S.R.; Anderson, J.J. Measurement of growth and iron deposition in Sphaerotilus discophorus. J. Bacteriol. 1976, 126, 257–263. [Google Scholar]
- Hashimoto, H.; Yokoyama, S.; Asaoka, H.; Kusano, Y.; Ikeda, Y.; Seno, M.; Takada, J.; Fujii, T.; Nakanishi, M.; Murakami, R. Characteristics of hollow microtubes consisting of amorphous iron oxide nanoparticles produced by iron oxidizing bacteria, Leptothrix ochracea. J. Magn. Magn. Mater. 2007, 310, 2405–2407. [Google Scholar] [CrossRef]
- Schwertmann, U.; Cornell, R.M. Iron Oxides in the Laboratory: Preparation and Characterization, 2nd ed.; Wiley-VCH: Weinheim, Germany, 2000; pp. 154–196. [Google Scholar]
- Eggleton, R.A.; Fitzpatrick, R.W. New data and a revised structural model for ferrihydrite. Clays Clay Miner. 1988, 36, 111–124. [Google Scholar]
- Châtellier, X.; West, M.M.; Rose, J.; Fortin, D.; Leppard, G.G.; Ferris, F.G. Characterizations of iron-oxides formed by oxidation of ferrous ions in the presence of various bacterial species and inorganic ligands. Geomicrobiol. J. 2004, 21, 99–112. [Google Scholar] [CrossRef]
- Karim, Z. Characteristics of ferrihydrites formed by oxidation of FeCl2 solutions containing different amounts of silica. Clays Clay Minerals 1984, 32, 181–184. [Google Scholar]
- Carlson, L.; Schwertmann, U. Natural ferrihydrites in surface deposits from Finland and their association with silica. Geochim. Cosmochim. Acta 1981, 45, 421–429. [Google Scholar] [CrossRef]
- Dyer, L.; Fawell, P.D.; Newman, O.M.G.; Richmond, W.R. Synthesis and characterization of ferrihydrite/silica co-precipitates. J. Colloid Interface Sci. 2010, 348, 65–70. [Google Scholar]
- Dyer, L.M.; Chapman, K.W.; English, P.; Saunders, M.; Richmond, W. Insights into the crystal and aggregate structure of Fe3+ oxide/silica co-precipitates. Am. Mineral. 2012, 97, 63–69. [Google Scholar] [CrossRef]
- Seehra, M.S.; Roy, P.; Raman, A.; Manivannan, A. Structural investigations of synthetic ferrihydrite nanoparticles doped with Si. Solid State Commun. 2004, 130, 597–601. [Google Scholar] [CrossRef]
- Toner, B.M.; Santelli, C.M.; Marcus, M.A.; Wirth, R.; Chan, C.S.; McCollon, T.; Bach, W. Biogenic iron oxyhydroxide formation at mid-ocean ridge hydrothermal vents: Juan de Fuca Ridge. Geochim. Cosmochim. Acta 2009, 73, 388–403. [Google Scholar] [CrossRef]
- Emerson, D.; Ghiorse, W.C. Ultrastructure and chemical composition of the sheath of Leptothrix discophora SP-6. J. Bacteriol. 1993, 175, 7808–7818. [Google Scholar]
- Emerson, D.; Ghiorse, W.C. Role of disulfide bonds in maintaining the structural integrity of the sheath of Leptothrix discophora SP-6. J. Bacteriol. 1993, 175, 7819–7827. [Google Scholar]
- Sakai, T.; Miyazaki, Y.; Murakami, A.; Sakamoto, N.; Ema, T.; Hashimoto, H.; Furutani, M.; Nakanishi, M.; Fujii, T.; Takada, J. Chemical modification of biogenous iron oxide to create an excellent enzyme scaffold. Org. Biomol. Chem. 2010, 8, 336–338. [Google Scholar] [CrossRef]
- Takeda, M.; Makita, H.; Ohno, K.; Nakahara, Y.; Koizumi, J. Structural analysis of the sheath of a sheathed bacterium, Leptothrix cholodnii. Int. J. Biol. Macromol. 2005, 37, 92–98. [Google Scholar] [CrossRef]
- Mann, S. Biomineralization: Principles and Concepts in Biogenic Materials Chemistry; Oxford University Press: Oxford, UK, 2001; pp. 154–196. [Google Scholar]
- Hashimoto, H.; Itadani, A.; Kudoh, T.; Kuroda, Y.; Seno, M.; Kusano, Y.; Ikeda, Y.; Nakanishi, M.; Fujii, T.; Takada, J. Acidic amorphous silica prepared from iron oxide of bacterial origin. ACS Appl. Mater. Interfaces 2013, 5, 518–523. [Google Scholar] [CrossRef]
- Hashimoto, H.; Itadani, A.; Fujii, T.; Nakanishi, M.; Asaoka, H.; Kusano, Y.; Ikeda, Y.; Kuroda, Y.; Takada, J. Nano–micro-architectural composites with acid properties: Magnetic iron oxides/amorphous silicate prepared from iron oxide produced by iron-oxidizing bacterium, Leptothrix ochracea. Mater. Res. Bull. 2013, 48, 1174–1177. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ishihara, H.; Hashimoto, H.; Taketa, E.; Suzuki, T.; Mandai, K.; Kunoh, H.; Takada, J. Silicon-Rich, Iron Oxide Microtubular Sheath Produced by an Iron-Oxidizing Bacterium, Leptothrix sp. Strain OUMS1, in Culture. Minerals 2014, 4, 565-577. https://doi.org/10.3390/min4030565
Ishihara H, Hashimoto H, Taketa E, Suzuki T, Mandai K, Kunoh H, Takada J. Silicon-Rich, Iron Oxide Microtubular Sheath Produced by an Iron-Oxidizing Bacterium, Leptothrix sp. Strain OUMS1, in Culture. Minerals. 2014; 4(3):565-577. https://doi.org/10.3390/min4030565
Chicago/Turabian StyleIshihara, Hiromichi, Hideki Hashimoto, Eisuke Taketa, Tomoko Suzuki, Kyoko Mandai, Hitoshi Kunoh, and Jun Takada. 2014. "Silicon-Rich, Iron Oxide Microtubular Sheath Produced by an Iron-Oxidizing Bacterium, Leptothrix sp. Strain OUMS1, in Culture" Minerals 4, no. 3: 565-577. https://doi.org/10.3390/min4030565