
Role of Fungi in the Biomineralization of Calcite

1
Laboratory of Microbiology, University of Neuchâtel, 2000 Neuchâtel, Switzerland
2
Institute of Earth Surface Dynamics, University of Lausanne, 1015 Lausanne, Switzerland
*
Author to whom correspondence should be addressed.
Minerals 2016, 6(2), 41; https://doi.org/10.3390/min6020041
Submission received: 29 February 2016
/
Revised: 25 April 2016
/
Accepted: 28 April 2016
/
Published: 5 May 2016
(This article belongs to the Special Issue Biomineralization: Towards a Unification of Concepts in Chemistry, Physics, Earth Sciences and Biology)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:In the field of microbial biomineralization, much of the scientific attention is focused on processes carried out by prokaryotes, in particular bacteria, even though fungi are also known to be involved in biogeochemical cycles in numerous ways. They are traditionally recognized as key players in organic matter recycling, as nutrient suppliers via mineral weathering, as well as large producers of organic acids such as oxalic acid for instance, an activity leading to the genesis of various metal complexes such as metal-oxalate. Their implications in the transformation of various mineral and metallic compounds has been widely acknowledged during the last decade, however, currently, their contribution to the genesis of a common biomineral, calcite, needs to be more thoroughly documented. Calcite is observed in many ecosystems and plays an essential role in the biogeochemical cycles of both carbon (C) and calcium (Ca). It may be physicochemical or biogenic in origin and numerous organisms have been recognized to control or induce its biomineralization. While fungi have often been suspected of being involved in this process in terrestrial environments, only scarce information supports this hypothesis in natural settings. As a result, calcite biomineralization by microbes is still largely attributed to bacteria at present. However, in some terrestrial environments there are particular calcitic habits that have been described as being fungal in origin. In addition to this, several studies dealing with axenic cultures of fungi have demonstrated the ability of fungi to produce calcite. Examples of fungal biomineralization range from induced to organomineralization processes. More examples of calcite biomineralization related to direct fungal activity, or at least to their presence, have been described within the last decade. However, the peculiar mechanisms leading to calcite biomineralization by fungi remain incompletely understood and more research is necessary, posing new exciting questions linked to microbial biomineralization processes.
1. Introduction
Biomineralization is defined as the formation of a mineral as a result of biological activity [1]. Three levels of biological mediation over the formation of biominerals can be defined [2,3]: (i) biologically controlled mineralization (BCM), when crystal nucleation, growth and morphology are under the tight genomic control of an organism; (ii) biologically induced mineralization (BIM), when biological activity induces physicochemical changes in the environment resulting in mineral nucleation and growth, also influencing mineral morphology; (iii) biologically influenced mineralization (or organomineralization sensu stricto; [2]) involving a biological matrix to initiate or enhance crystal nucleation and growth, with an influence on mineral morphology. Importantly, in this last type of biomineralization, living organisms are not directly required and only the organic fraction matters as a template for nucleation. For this reason, Trichet and Défarge [4] defined organomineralization as a “precipitation mediated by non-living organic substrates in soils and sediments” and this is the definition that will be used throughout this contribution.
The term microbe encompasses diverse organisms that are too small to be visible to the naked eye and belonging to both the Prokaryotes (bacteria and archaea) and the Eukaryotes (fungi, protists, and microscopic algae; [5]). All of them are involved in at least one type of biomineralization. For instance, BCM is common in algae and protists, but also very widespread in macroscopic pluricellular organisms (skeletons, shells, etc.; [6]). As a result, it is considered of minor importance in the microbial world except in the case of magnetotactic bacteria [7]. On the contrary, the two other types of biomineralization (BIM and organomineralization) are very common in microbes. Numerous examples exist in the scientific literature dealing with microbial BIM (e.g., [8,9,10,11]). Organomineralization studies published in the last decade demonstrate that this process might be of greater importance than previously thought (e.g., [2,3,12,13,14]). In particular, microbes with their minute sizes (1–20 µm) exhibit large surface to volume ratio and therefore microbial surfaces and residues represent a very large fraction of potential organic templates for mineral precipitation.
Among biominerals, calcite and aragonite (two polymorphs of calcium carbonate, CaCO3) are the most commonly encountered minerals [15]. As a matter of fact, CaCO3 biominerals precipitated under, or related to the influence of biological processes are widespread and represent a greater contribution to the CaCO3 pool on Earth than physicochemical CaCO3 [16,17]. As a result, CaCO3 biominerals are found in a myriad of different habits. Their recognition as biominerals, as well as the actual processes that led to their formation, are sometimes difficult to assess. In particular, biominerals from biologically induced or influenced mineralization often exhibit poorly defined shapes in contrast to biominerals formed under controlled conditions [18]. As a result, when observed in association with a given organism, attributing their formation to the activity of this organism is not always straightforward.
Biominerals and biomineralization are important fields in both geo- and bio-sciences because of their great influence in global biogeochemical cycles: CaCO3 minerals are of major importance in both the global carbon (C) and calcium (Ca) biogeochemical cycles. Therefore, processes related to their dissolution and precipitation are of major interest in our understanding of how microbes influence global biogeochemical cycles at their microscopic scale. In addition, biominerals represent critical indicators of past environmental conditions when observed in fossil records, as they offer testimony to biological activity [1]. Therefore their clear recognition as biominerals and the processes to which they might be related, are of great significance [1,2]. Finally, in an era of increased pressure to find sustainable solutions to our industrial way of life it has become more important than ever to utilize microbes as catalysts for some activities such as metal decontamination for example, and presses the need to more thoroughly characterize the diversity of biomineralization processes linked to microbes.
The aim of this contribution is to review the latest advances in the field of CaCO3 fungal biomineralization. In order to set the context, a brief definition of the fungal kingdom will be presented together with its role in global biogeochemical cycles. Then a thorough review of what is known currently about fungal biominerals will be presented, with an emphasis on fungal CaCO3 biominerals including both a literature review, as well as the work of the authors. Finally, the last part will be devoted to what the reasons might be for the lack of knowledge regarding the field of fungal biomineralization when compared to prokaryotic biomineralization.
2. Fungi in the Geobiosphere
Fungi are ubiquitous on the surface of the Earth, wherever oxygen is present; they are also able to live in the absence of oxygen, however, this is not their main lifestyle. Soils are believed to be their most characteristic habitat where their biomass can reach 75% of the soil microbial biomass [19]. They are also very common in more oligotrophic environments such as caves and rock surfaces [20,21,22]. Finally, they are also common inhabitants of aquatic ecosystems such as streams and lakes and are even observed down in deep-sea environments [23,24].
Fungi are chemo-organo-heterotrophic organisms and therefore depend on organic matter in order to sustain their metabolism. They obtain their carbon source either from dead organic matter (as saprotrophs) or from associations with living partners (either as mutualistic or parasitic symbionts). Mutualistic symbiosis may occur with plants (mycorrhizae), cyanobacteria or algae (lichens) and even with animals [25,26]. These mutualistic symbioses have a paramount importance in terms of global biogeochemical cycling of elements [20,25]. Fungi acquire their nutrients by absorption, meaning that they must first pre-digest their substrate using hydrolytic or oxidative enzymes secreted in the external environments. Solubilized nutrients are then transported inside their cells.
Most fungi are multicellular, exhibiting a branched filamentous growth habit called the mycelium. This vegetative structure consists of rigid tubular hyphae made of elongated cells arranged one after the other [26]. Hyphae have a very high mechanical resistance resulting from the presence of the fungal cell wall, an highly rigid structure and the turgor pressure exerted by the protoplast on the inner side of the cell wall [27,28]. Thanks to this high mechanical resistance, fungal hyphae have been observed actively drilling into mineral substrates [29,30].
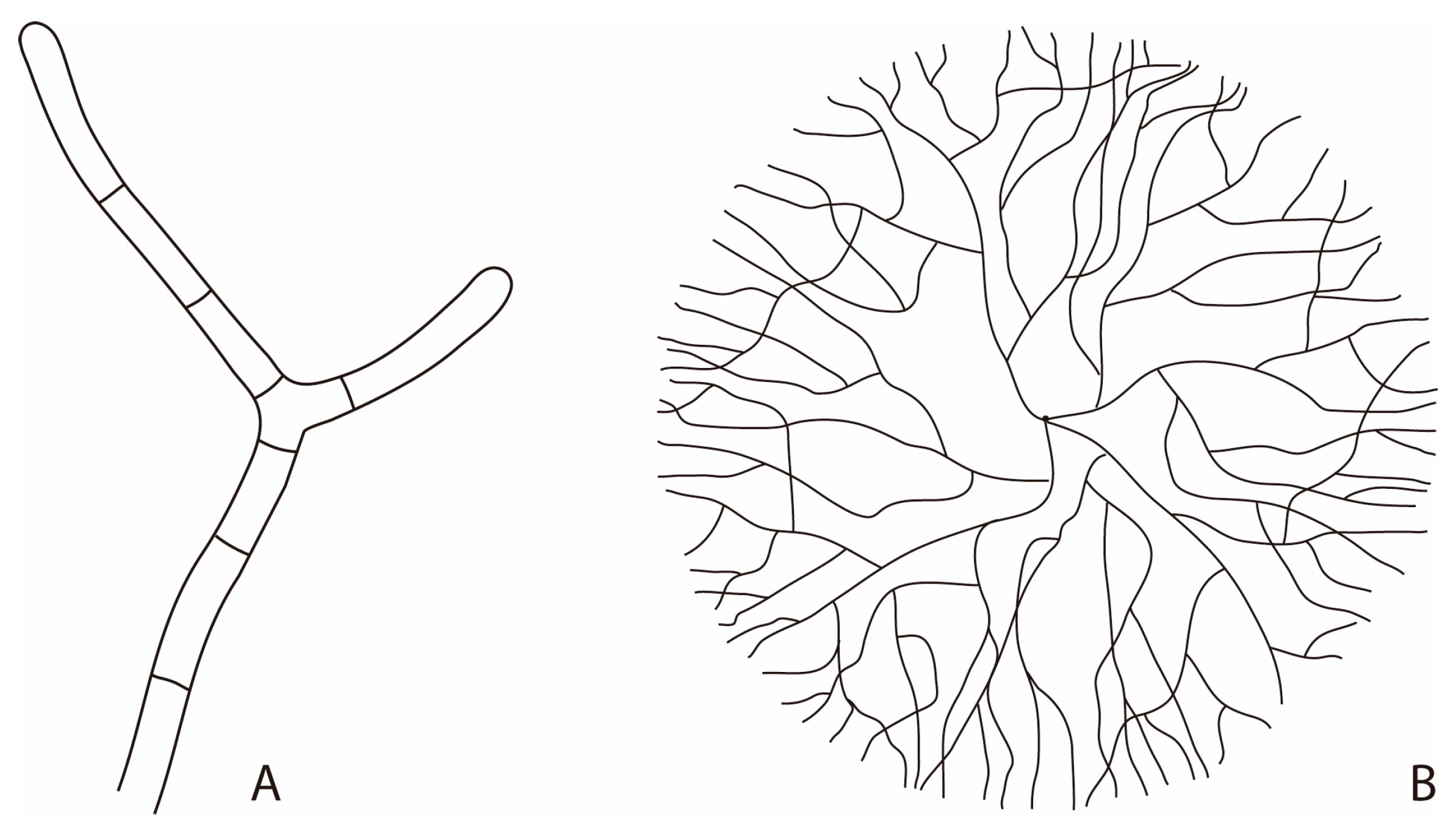
Hyphae grow apically with new apices arising from the creation of lateral branches forming a three-dimensional network (Figure 1). This particularity allows them to explore and exploit their environment while maintaining exponential growth by branching. This pattern is of great advantage to fungi as they usually live in environments displaying both nutritional and structural spatio-temporal heterogeneities [31]. Metabolically active cytoplasm usually move forward together with the expanding hyphal tips as a result of exploiting heterogeneous substrates, leaving the older parts of the mycelium as empty tubes [28,32]. In soils for instance, it has been shown that most of the hyphal network is actually made of empty hyphae [33], which are, however, important for the functionality of the whole mycelium.
Metabolic activity of a mycelial network is highly heterogeneous and depends mostly on the available nutrients [34,35]. Extensive translocation of nutrients and water occurs between metabolically active parts. As a result, the mycelial network can be described as a real logistic network [36,37].
Besides multicellular fungi, there are also unicellular fungi as well as polymorphic fungi, which are able to change from a uni- to a multi-cellular lifestyle as a result of abiotic or biotic stimuli. Finally, fungi reproduce both sexually and asexually, resulting in the formation of numerous types of spores [26]. Spores are used both for resistance to unfavourable conditions and for dispersion. As a result, the density of fungal spores in the aerial environment is high and allows the ubiquitous distribution of fungi in terrestrial environments to be explained. Spores are so common in the atmosphere that they contribute to cloud formation [38,39].
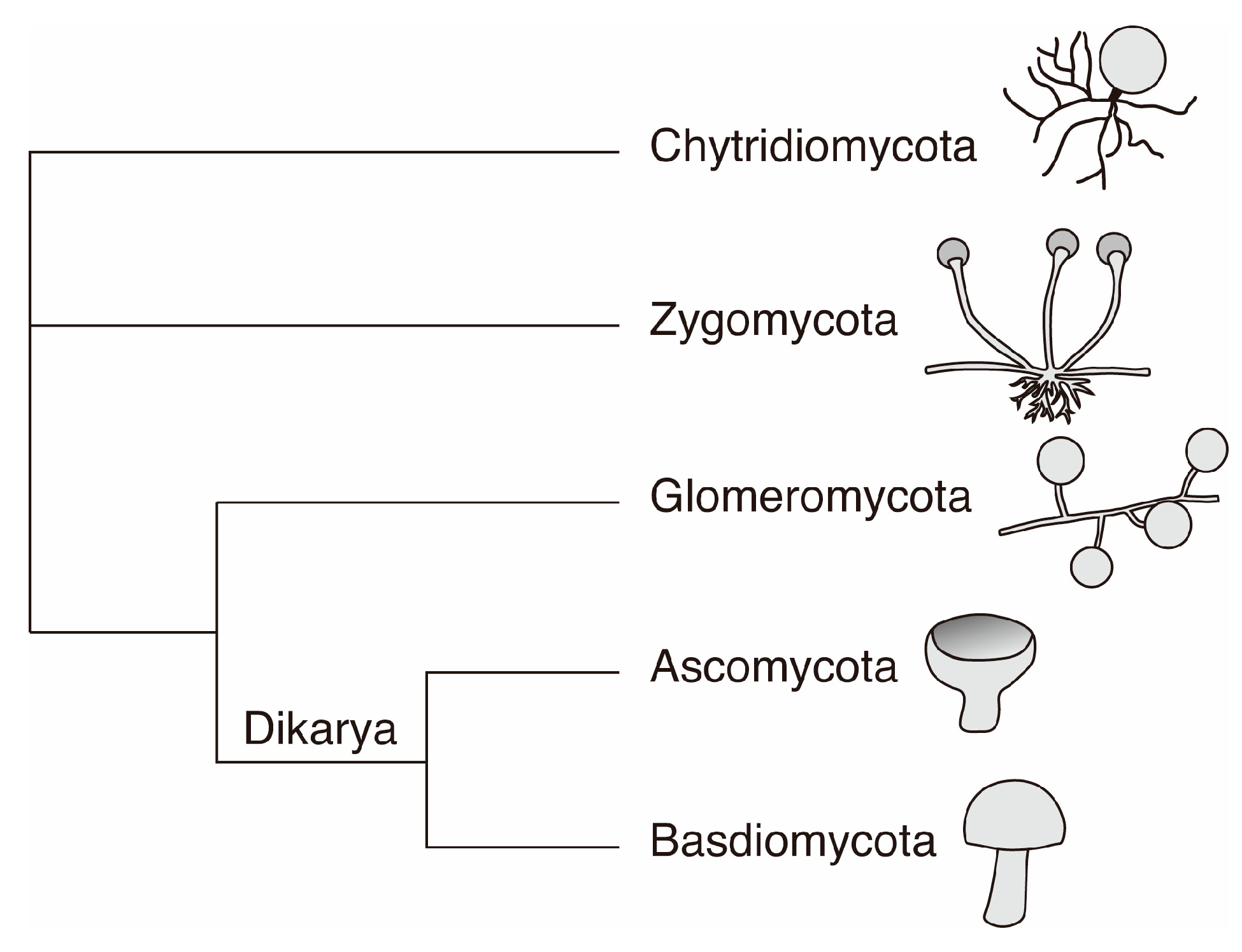
In phylogenetic terms, the fungal kingdom is a monophyletic group [40]. In a simplified way, the fungal tree of life can be described as containing five phyla: Chytridiomycota, Zygomycota, Glomeromycota, Ascomycota and Basidiomycota [26] (Figure 2).
Chytridiomycota is a primitive group of mostly unicellular fungi with both parasitic and saprophytic lifestyles. Zygomycota comprise of many opportunistic fungi, either parasitic or saprotrophic. Glomeromycota form characteristic associations with plants called endomycorrhizas or arbuscular mycrorhizal fungi (AMF). Over 80% of land plants are involved in a mutualistic association with AMF and therefore Glomeromycota represent a phyla of major importance for the functioning of most terrestrial ecosystems [41].
Ascomycota and Basidiomycota belong to the sub-kingdom Dikarya which represents 98% of the fungal diversity described currently [42]. Both uni- and multi-cellular forms are observed within the Dikarya. Ascomycota represent the largest fraction of fungi described currently with 64% of the described fungal species. They are ubiquitous in the environment and two of their most prominent lifestyles are related to either lichen symbioses or to parasitism of plants and animals. However, they are also common mycorrhizal fungi, as well as saprotrophs. Additionally, some have developed an extremophilic lifestyle by colonizing rock surfaces in extreme environments (microcolonial fungi [21]). Finally, Basidiomycota account for 34% of the total described fungal diversity. They are the common mushroom forming fungi and are well known as ectomycorrhizal mutualistic symbionts of trees in boreal and temperate forests [41]. However, they are also common saprotrophs and parasites of plants and animals. Ascomycota and Basidiomycota exploit highly varied ecological niches from organic rich to very oligotrophic mineral environments and are therefore important players in global biogeochemical cycles.
3. Importance of Fungi in the Biogeochemical Cycles of Elements
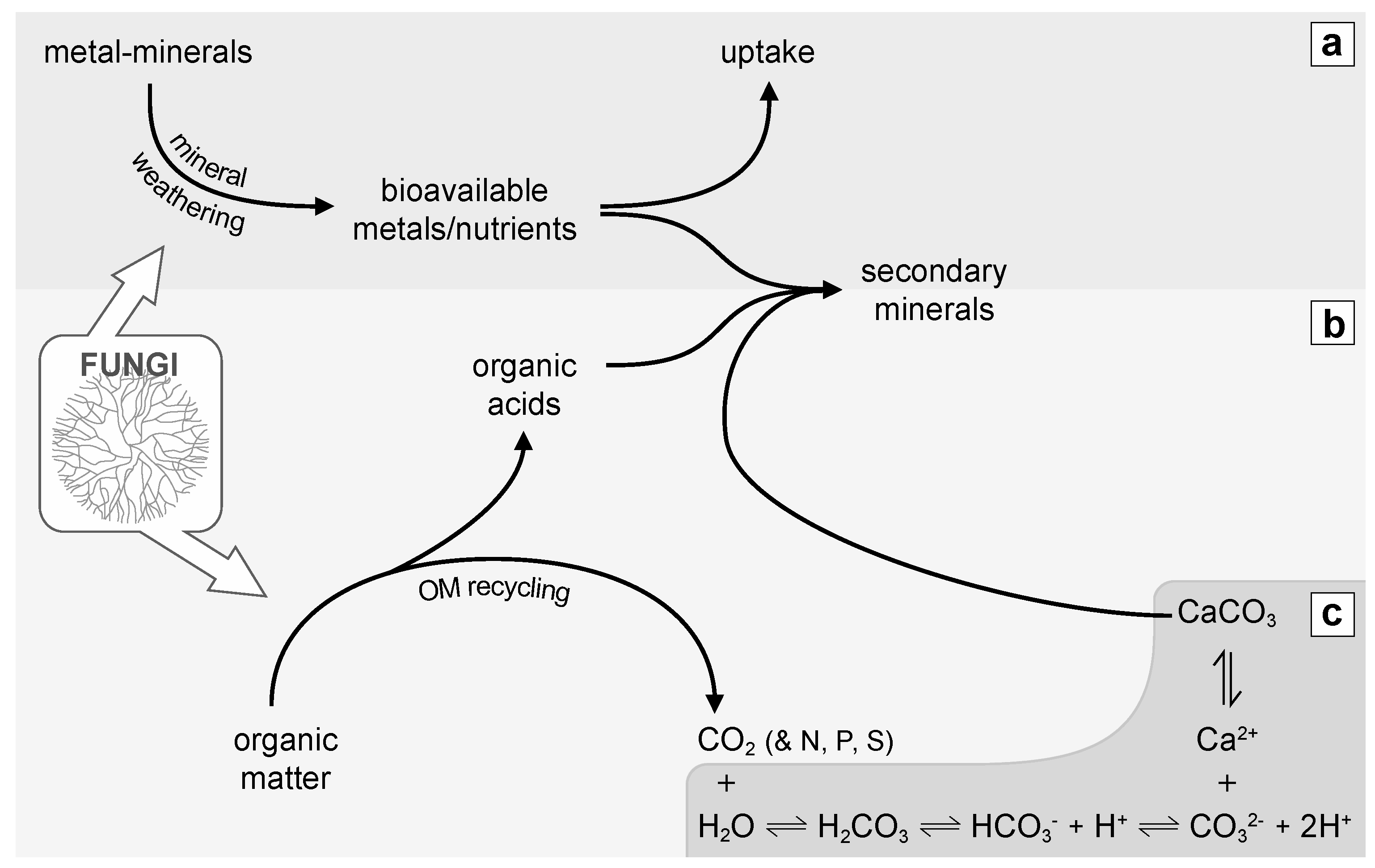
Geomycology, a term coined by Professor Geoffrey M. Gadd and his team, is the field that investigates the role of fungi in biogeochemical cycles [25,43,44]. Traditionally, fungi have been considered as key players in the global C biogeochemical cycle thanks to their ability to recycle organic matter, either through saprophytic or parasitic lifestyles. This activity typically leads to the release of CO2 and major nutrients such as N, P and S (Figure 3; [25]). However, fungi also interact with metals and other inorganic minerals and therefore are also involved in the biogeochemical cycling of compounds such as Fe, Ca, K, P, Mg, etc. Typically, fungi involved in lichen and mycorrhizal symbioses are important players of mineral turnover at global scales [26]. Similarly, microcolonial fungi colonizing rock surfaces in arid environments interact with their mineral substrate, influencing its chemical and physical stability [21,45].
The interaction of fungi with metals and minerals can be summed up by two groups of processes leading to either mobilization, or immobilization of metals and minerals in solution [46]. Mobilization involves all the mechanisms leading to the transfer of mineral constituents into solutions. These processes are crucial, for instance, for nutrient bioavailability in the biosphere but can also lead to the release of toxic compounds in the environment [32,47]. Mobilization of metals and minerals by fungi involves both mechanical and biochemical processes, with the latter considered as being more important in terms of the amounts of released material. Mechanical action of fungi is a result of their ability to grow into cracks, as well as to drill within mineral substrata [20,48,49]. Biochemical action involves several processes such as chelation, through siderophores and organic acids, redox transformation and methylation processes. Collectively these processes can also be referred to as bioweathering [43].
On the other hand, mechanisms leading to the immobilization of metals and minerals can be described by both active and passive mechanisms. Active mechanisms involve intracellular accumulation and redox transformations, whereas passive mechanisms involve sorption to cell surfaces and exopolymeric substances (EPS) secreted outside cells, as well as formation of insoluble complexes with conjugated bases of organic acids. Immobilization processes can eventually lead to biomineral formation [25,46].
Within the last few decades, numerous studies have been dedicated to the interaction of fungi with metals and minerals and have shown that fungi are able to resist high concentrations of toxic metals [50,51], are important players in mineral weathering [29,30,48,52,53] and are large producers of secondary mineral compounds, in particular of metal-oxalates [49,54,55,56,57,58] (Figure 3). As a result, fungi, similarly to prokaryotes, have to be considered as major players in biogeochemical cycles. Indeed, in terms of diversity of metabolic processes associated to elemental cycling, they have a profound influence on biogeochemical cycles of both organic and inorganic components of the Earth system [25,43,49,59].
4. Fungal Biominerals
When considering microbial biomineralization, the majority of studies and literature deals with prokaryotes. To the contrary, induced microbial biomineralization by eukaryotes is much less documented, particularly when considering the Fungal Kingdom [20,43,60,61]. As an example, a quick search in Web of ScienceTM for fung* AND biomineralization in comparison to bact* AND biomineralization retrieves 129 and 1238 hints respectively. As fungi are ubiquitous in both terrestrial and aquatic environments, this conflicts with their ecological importance.
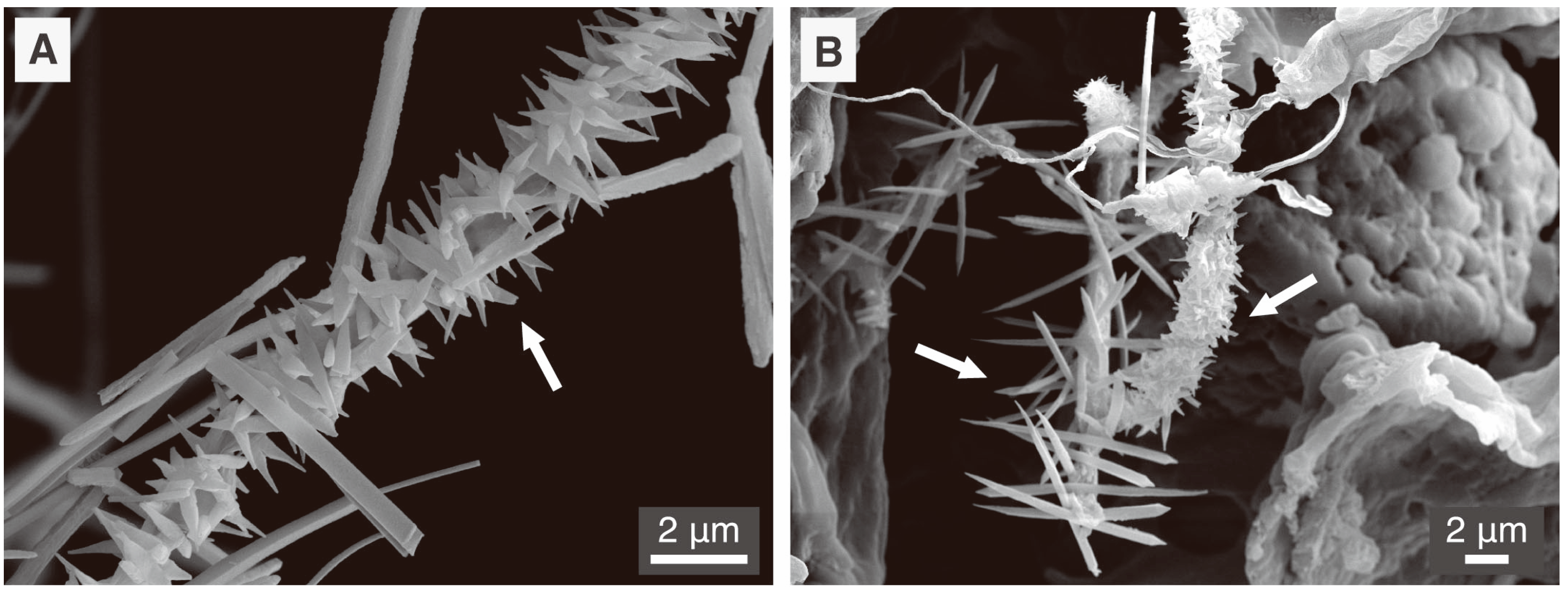
Fungi are well known to produce large amounts of various organic acids, also called low molecular weight organic acids (LMWOA) [20,25]. As reported by Sterflinger [20], acetic, citric, formic, fumaric, gluconic, glyoxylic and oxalic acids are commonly produced by a wide array of fungi observed in relationship with mineral substrata. Most of these acids are present as their conjugated bases at environmental pHs (4 to 9) and are therefore prone to precipitation with different metals depending on their corresponding salt solubility product [62]. Oxalic acid is one of the most important and ubiquitous organic acids produced by fungi as it serves several different functions in fungal metabolism. It is involved in pathogenicity, competition with other fungi, wood degradation, nutrient release from minerals and resistance to toxic metals [55,63]. Oxalic acid is a strong organic acid (pKa1 = 1.23 and pKa2 = 4.19) and is mainly present in the environment as the oxalate ion or as an oxalate salt. As a result of oxalate ubiquity in relationship to fungal metabolism, metal-oxalates are the most common and widespread biominerals of fungal origin described in present-day literature. Figure 4 depicts some typical calcium-oxalate crystals associated to fungal hyphae in samples from natural environments.
However, it is expected that fungi might be linked to the formation of other types of biominerals. Indeed, they are ubiquitous in most of the terrestrial environments where both their biomass and surface-to-volume ratio are enormous [19]. Additionally they have the ability to interact with various metals and minerals [25]. Despite this fact, their implication in the genesis of other types of minerals remains poorly documented to this day. A recent study mentions an Ascomycota, Purpureocillium lilacinum, as being involved in the biomineralization of jarosite, a peculiar type of sulphur and iron mineral formed in acidic conditions, through its metabolic activity [64]. Biomineralization of phosphate minerals as a result of fungal activity has also been documented recently [65,66,67,68,69]. All are typical examples of BIM, confirming the fact that fungi are also important players in microbial biomineralization.
5. Fungi and CaCO3
Fungi are involved in both CaCO3 bioweathering and biomineralization [20,48,52,70,71,72]. These two activities are actually strongly related, since products released as a result of CaCO3 dissolution can further re-precipitate as CaCO3 depending on physicochemical conditions [71]. Whether CaCO3 is stable or not in a given solution depends mainly on factors such as temperature, pH, pCO2 and/or carbonate alkalinity. Carbonate alkalinity is controlled mainly by the pH, which will define the amount of the various carbonate species present in a solution (CO2 gas, H2CO3, HCO32− and CO32−). As a result, in order to precipitate CaCO3 two factors are critical: carbonate alkalinity and calcium (Ca2+) concentration [2,73].
Fungal metabolism can influence both carbonate alkalinity and Ca2+ concentrations. Typical fungal activities that can decrease alkalinity are heterotrophic respiration leading to an increase in pCO2, production of organic acids and excretion of H+ during fungal thigmotropism [74]. In contrast, typical fungal activities that can increase carbonate alkalinity are water consumption (e.g., through translocation within the fungal network), physicochemical degassing of fungal respired CO2, organic acid oxidation [75,76], urea mineralization [77,78] and nitrate assimilation [79].
In addition to their influence on carbonate alkalinity, fungi can also influence Ca2+ concentrations, either directly or indirectly. Ca2+ is important for the apical growth of fungal hyphae as it should be highly concentrated at the apex for proper apical growth. Consequently, Ca2+ concentration within metabolically active fungal cells is under strict control: Ca2+ must be concentrated at the apex and instantly decreased in subapical regions [80]. Average concentrations of free cytoplasmic Ca2+ range from 100 to 350 nM, whereas at the tip, concentrations up to 2600 nM are observed. In order to keep this steep gradient, fungi have to efficiently regulate Ca2+. Ca2+ can enter the cytoplasm both actively and passively, so Ca2+ concentration in the cytoplasm is maintained at low levels by actively pumping it either out of the cell, or by sequestration in organelles (mitochondria, endoplasmic reticulum, and vacuoles), or by binding it onto cytoplasmic proteins (calmodulins) and within the cell wall [50,80,81].
Besides fungal metabolic activities that can influence alkalinity and Ca2+ concentrations potentially leading to induced CaCO3 biomineralization, fungi can also participate through organomineralization. Fungal cell walls, as well as secreted EPS, can adsorb various metals such as Ca2+. In environments with high metal concentrations, it represents a first rampart of passive metal immobilization [50]. Interestingly, among fungal cell wall polymers, chitin is known for its ability to bind Ca2+ [82]. Cell wall glycoproteins with available acidic groups may display a similar binding capacity [50]. In fact, overconcentration of Ca2+ on an organic matrix is a first prerequisite for CaCO3 nucleation and subsequent organomineralization [2]. Consequently, there are several factors in relationship to fungi that can directly or indirectly influence CaCO3 stability.
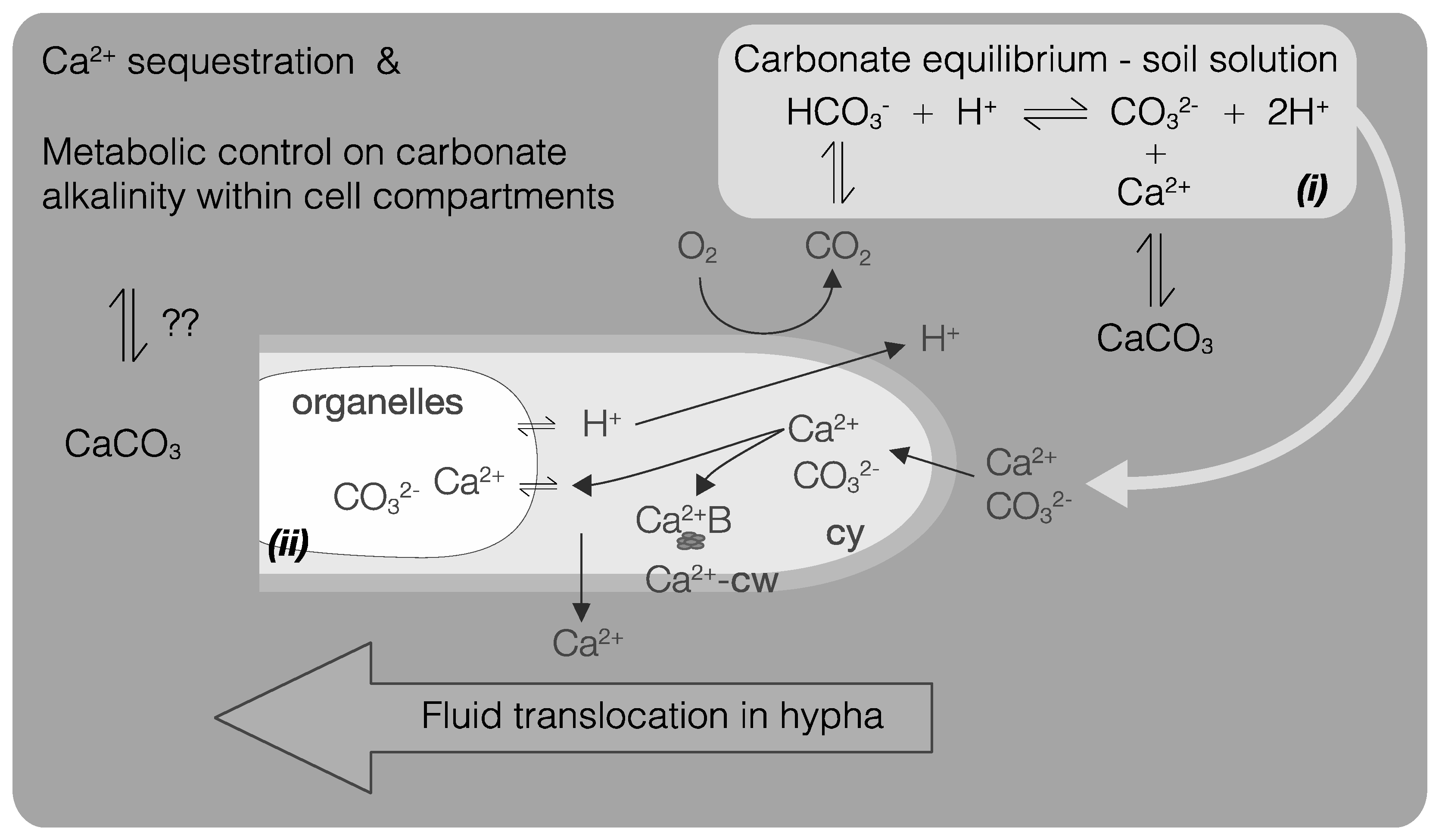
Fungi growing in Ca-rich environments are exposed to high concentrations of Ca2+ and CO32− in solution. These conditions likely represent a source of stress due to subsequent osmotic pressure and Ca2+ cytotoxicity. The formation of Ca-oxalates has been suggested as a means to immobilize excessive Ca2+ [54,55,83]. Precipitation of CaCO3 by fungi may represent a similar passive mechanism leading to a decrease of their internal Ca2+ content. This process is documented for bacteria [84,85,86], but remains a hypothesis regarding fungi. Similarly to osmotic pressure generated by Ca2+, excessive alkalinity can also represent a stress for fungal cells and precipitating metal-CO3, such as CaCO3, could represent a mean of intracellular protection (Figure 5).
5.1. Fungal CaCO3 Produced in the Laboratory
CaCO3 biominerals are regularly mentioned in typical fungal environments and as a result, a fungal implication in their genesis has often been suggested [43,48,70,71,87,88,89,90,91,92]. CaCO3 is a ubiquitous component in the environment and is the most common biomineral [16]. Up until recently, it was considered that mostly bacteria and microscopic algae were involved in its biomineralization at a microbial level [61,71]. However, within the last decade, several studies have started to unravel some of the specific mechanisms leading to fungal CaCO3 biomineralization.
Li et al. [78,93] demonstrated that urease-positive fungi similarly to ureolytic bacteria, can induce CaCO3 biomineralization. In particular, in one of their studies [93], they isolated fungi from calcareous soils, showing that in these environments ureolytic fungi are indeed present and may contribute to CaCO3 formation/stability. This is again a typical example of BIM since the main consequence of urea transformation into ammonium is a pH increase, one of the critical factors for CaCO3 precipitation.
In another study linked to nitrogen metabolism, Hou et al. [79] noted the precipitation of CaCO3 by Alternaria sp. as a consequence of nitrate removal from the medium. Nitrate uptake correlated with an increase in pH, leading to CaCO3 precipitation. Interestingly, they also mentioned that nucleation seems to occur around silica particles and that the initial stage of biomineralization involves an amorphous calcium carbonate (ACC) intermediate. This latter fact has already been discussed as a possible intermediate, yet rarely identified in experimental procedures for biominerals by several authors [94,95,96].
Rautaray et al. [97,98], Ahmad et al. [99] and Sanyal et al. [100] performed similar studies in which they demonstrated that fungal biomass exposed to significant concentration of Ca2+ (as CaCl2) leads to CaCO3 biomineralization. They observed very distinct morphologies of crystals depending on the fungal species involved and proposed that specific proteins were responsible for this. Moreover, this effect was also reproduced only from cultures filtrates, meaning that some soluble compounds were linked to this process. As a result, this is an example of both BIM and organomineralization.
Other authors performed experiments in less controlled conditions, i.e., in microcosms leading to the production of CaCO3 phases as a consequence of the presence of fungi. Burford et al. [71] grew Serpula himantioides (a brown rot Basidiomycota) and Cephalotrichum sp. (previously isolated from a limestone sample) on limestone powders with no organic C source. They observed extensive crystal production of both calcium oxalate and CaCO3, the ratio between the two chemical components being a consequence of the fungal species. Crystal production was observed with both dead and live biomass, with a more prominent effect using live biomass. This again points to the fact that both BIM and organomineralization processes are also related to fungal presence and activity.
In another microcosm study, Masaphy et al. [101] could demonstrate the production of CaCO3 as a result of the presence of Morchella sp. They observed calcification of quartz sands as well as coating of hyphae with nanostructures identified as CaCO3. They proposed that water uptake by the mycelium, evaporation and degassing of respired CO2 are responsible for adjusting the physicochemical conditions in favour of CaCO3 precipitation. This process corresponds to BIM. In addition, they observed fungal hyphae within cavities with CaCO3 precipitated at some distance. They proposed that LMWOA excreted by fungal hyphae led to dissolution of CaCO3 which will further re-precipitate in favourable physicochemical conditions. This is in agreement with the mechanism already discussed by Verrecchia et al. [102], Burford et al. [43], Kolo et al. [48], as well as Fomina et al. [49] (rock-building hypothesis).
As a conclusion, all these examples point to the fact that fungi are responsible, similarly to bacteria, for a fraction of the CaCO3 biomineralization observed in the natural environment. Moreover, they are capable of both induced biomineralization and organomineralization processes.
5.2. Fungal CaCO3 in the Natural Environment
There are many records of suspected fungal involvement in the production of secondary CaCO3 in the natural environment [43,48,70,71,87,88,89,90,91,92]. Owing to the knowledge gathered from laboratory experiments performed during the last ten years, it is now possible to support observations made by early authors. Indeed, results of some typical heterotrophic activity performed by fungi enhance saturation in CaCO3, e.g., (1) conversion of urea into ammonium [78,93]; (2) uptake of nitrate [79]; (3) degassing of respired CO2; (4) water translocation [101]. All these examples are typical of BIM. In addition to this, Bindschedler et al. [14] demonstrated that fungi can be a source of organic substrates that are likely to act as template for organomineralization in environments supersaturated in CaCO3.
Currently, no in situ studies exist to actually demonstrate the role of fungi in CaCO3 precipitation directly in the natural environment. Consequently, only a suspected link can be drawn. In order to discuss this aspect, we choose to present our own experience with a typical example of secondary CaCO3 crystals attributed to fungal activity but still under debate! These minerals are known as needle fibre calcite (NFC) and their associated nanofibres ([103] and references therein; [14] and references from their Table 1; [90,91,92,104,105]). Both features are distinct micro- to nano-crystalline shapes of calcite crystals [14] (Figure 6). Discussions among the scientific community attribute the origin of these features either to physicochemical [106,107,108] or biogenic [90,103,104,109,110,111,112] processes.
However, a fungal origin can be further supported based on what is presently known about the involvement of fungi in CaCO3 biomineralization. Indeed, both NFC and nanofibres are observed only in vadose calcareous environments. Both Li et al. [78] and Burford et al. [71] demonstrated that calcareous environments are efficient sources of calcifying fungi. Morever, as discussed in Castanier et al. [73], the heterotrophic pathway of CaCO3 precipitation presented for bacteria is also valid for fungi. As a result, fungi have the ability to induce the physicochemical conditions leading to carbonate supersaturation.
In addition, they also act on the Ca2+ component of the system. Energy Dispersive Spectroscopy (EDS) measurements under a Transmission Electron Microscope (TEM) of ultrathin sections of fungal rhizomorphs sampled in calcareous environments highlighted high concentrations of Ca within hyphal walls as well as in intrahyphal inclusions (Figure 7; for details of sampling site and experimental procedures, see reference [92]).
These results stress that two factors critical for CaCO3 precipitation in calcareous environment can be triggered by fungi: (1) carbonate alkalinity and (2) concentration of Ca2+ on an organic matrix. However, in situ studies actually linking CaCO3 biominerals and fungal activity are still lacking. This is mainly due to the access to advanced analytical technologies having only become available within the last decade.
6. Methodological Issues in the Study of Fungal Biominerals from Natural Environments
An approach to associate an organism to a mineral in order to demonstrate a causal link, is to look for biomarkers. A biomarker represents a measurable form of a given biological state. Depending of the research field using this term, it can be an organic or an inorganic chemical compound, an isotopic signature or even a morphological characteristic [113]. Indicators based on organic compounds are frequently used as tracers in geological and environmental processes [114]. Therefore, the search for organic molecules in mineral matter might be a way of emphasizing a biotic origin of a particular mineral. In the following section, we propose to use the terms “organic signature” or “organic biomarker” to illustrate this.
There are two ways in which an organic molecule may be trapped within the crystal lattice. First, the nucleation and/or the crystal growth are controlled by an organic matrix, which is then subsequently trapped in the crystal lattice [115]. Second, organic matter present in the environment is passively trapped between crystal planes of polycrystalline phases [116]. Therefore, this latter process has no influence on crystallogenesis sensu stricto. However, it may still be used as an environmental or biogeochemical proxy.
Retrieving an organic signature from environmental samples may be achieved in several manners. The analysis may be performed on a bulk sample by an “extraction-analysis” approach which is then most likely a mixture of several phases, as experienced in our own observations with NFC and nanofibres [91,105]. By this method, the presence of a given organic biomarker may indicate the presence of an organism or a metabolism (depending on the nature of the biomarker) but it does not necessarily indicate an implication of the targeted organism in the crystal genesis [117,118,119]. Alternatively, targeting the inside of the crystal lattice using fine microanalytical tools may be carried out. Yet this approach implies a critical size for the single crystal in order to be able to focus the analysis [120].
The “extraction-analysis” approach usually leads to poor results. Often no organic signatures are detected, a result which is related to either detection limits, which are frequently above biomarker compound concentrations within the object investigated, or the simple absence of the considered biomarker. However, such results tend to remain unpublished (personal observation by the authors).
In contrast, if organic biomarkers are detected and identified, their causal link to a given biomineral remains dubious [117,118,119]. Consequently, using a fine-scale analytical approach is necessary because of the low specificity and/or poor resolution of broad-spectrum methods [120,121,122]. Indeed, the study of organic molecules possibly trapped into the crystal lattice of sub-micrometric objects such as biominerals, require methods capable of very high spatial and chemical resolutions [123].
Many biominerals exhibit submicron sizes, which are often very near or under the resolution limits of most of the methods currently available in the field of in situ microscopy and microanalysis. The use of Scanning Electron Microscopy (SEM) and TEM coupled to EDS analyses has led to major advances in the field of biomineralization. However, it does not allow for the direct discrimination of organic vs. mineral matter, nor does it permit molecular characterization of the objects observed since EDS only analyses elemental composition.
Electron Energy Loss Spectroscopy (EELS) and Electron Spectroscopic Imaging (ESI) coupled to the TEM are very powerful microanalysis tools, as together they enable element and high spatial resolutions. The latter configuration corresponds well to the sizes of the objects to be analysed but a major problem subsists because of the high heterogeneity of natural samples. As a result, spectrograms obtained with such samples are almost impossible to interpret, as too many peaks are present. This problem was not encountered in lab experiments, as shown in a study by Cailleau et al. [104]. Therefore, it appears that this method is not adapted to the investigation of natural samples.
An accurate examination of the chemistry of biominerals at submicrometric scales requires a microanalytical tool of very high spatial and spectral resolution that is also able to focus on one element at a time, such as Scanning Transmission X-ray Microscopy (STXM). Looking at an object being analysed through X-ray images is a critical approach in biomineralization. The use of NEXAFS (Near edge X-ray absorption fine structure) spectrometry allows for very high mass resolution appropriate for detecting light elements such as C, N, O and Ca which are constitutive elements of fungal CaCO3 biominerals. Moreover, analysis with fine X-ray sources provide the opportunity to retrieve information regarding bond types between elements and is therefore the tool of choice when studying organic materials. Indeed, organic materials are usually often composed of the same major elements (C, H, O, N) but the difference lies on the bond types between these elements. A final advantage of this method is the possibility of analysing hydrated environmental samples [120]. By this means, it is possible to avoid the complex preparation of samples prior to observation, which can potentially create artefacts to the sample. Indeed, one major problem that remains in conventional electron microscopy studies with these kinds of samples, especially when containing a large fraction of biological material, is that they have to be fixed and dehydrated using chemical and/or physical methods. Each of these treatment steps can modify the structure of a sample, as well as the relationships between the different elements within a sample [124]. This fact has been suggested as an explanation for the observation of enigmatic or unidentified features in some studies [117,125].
To sum up, all the characteristics mentioned above make STXM a tool of very high analytic resolution, able to retrieve nanometer scale chemical information under in situ environmental conditions [120]. And as a matter of fact, STXM has proven to be an appropriate tool to study the biogenecity of some minerals [53,126,127,128,129].
NanoSIMS (Secondary Ion Mass Spectrometry) can also be a valuable option for investigating fungal biominerals as it also exhibits very high lateral (around 50 nm [130]) and mass resolutions. However, nanoSIMS does not have the capability of analysing bond types, which is critical when studying organic materials as explained above. Fine elemental mapping as well as isotopic discrimination are possible [130], which could lead to the recovery of either an isotopic or elemental pattern in biominerals, which may help to discuss their origin [131].
Finally, another approach often used in the field of geomycology are DNA-based methods (DNA extraction, PCR and sequencing). While crucial advances have been made in the last ten years, in particular regarding sequencing methods, fungal community ecology still lags behind prokaryotic community ecology. The term microbial has often been (and is still sometimes) used for studies only describing bacteria [60,132]. Additionally, besides exhibiting very diverse life forms, fungi spread across three ecological categories: saprophytes, mycorrhizae and pathogens/parasites, which have been targeted individually by researchers [133,134]. However, recent research clearly demonstrates that all three categories matter for soil functioning [133]. Therefore, a global overview of fungal community ecology (the different fungi present in an environment and the ecological functions they fulfil) requires the use of advanced molecular methods, which are currently being developed. Consequently, the poor results usually obtained with DNA-based approaches in the field of fungal biomineralization [135] are more likely due to poor primer specificity and under-representation of fungal sequences in databases [136,137,138], as well as peculiarities of fungal biology [134] in comparison to prokaryotes, rather than a lack of fungal involvement.
7. Conclusions
Microbial biomineralization is a rapidly expanding field, mainly due to the fast development of new methods to qualitatively and quantitatively analyse interactions at the scale of microbes. However, while microbe is a term that defines a wide range of organisms (from prokaryotes to eukaryotes), studies dealing with microbial biomineralization are still biased towards prokaryotes. This is probably the result of traditional microbial ecology that first developed in the field of bacteriology, as well as the difficulty in studying complex organisms such as fungi that usually exhibit very heterogeneous morphologies and metabolic activities in space and time. Bacteria are usually single-celled organisms whose morphology and related metabolic activity are much easier to define and to study accurately in laboratory conditions. In addition, fungi have long been investigated regarding their importance in organic matter degradation, either through saprotrophy or parasitic/pathogenic interactions. Due to this, their link to inorganic constituents was considered as being restricted to mineral nutrition through mycorrhizal symbiosis, mineral weathering of lichens as well as production of mycogenic oxalates. Presently this view appears as being rather minimalistic, since recent studies in the field of geomycology have demonstrated that fungi are widely present and active in relationship to mineral substrata on a global scale. They are, similarly to bacteria, major players in the mobilization and immobilization of various mineral and metal compounds. In particular, considering the immobilization of mineral compounds, their action goes far beyond producing oxalate minerals. Fungi have an important role in microbial CaCO3 biomineralization and participate to both biologically induced biomineralization and organomineralization processes. Several examples of induced biomineralization through fungal metabolic activity (i.e., through heterotrophic pathways) have been documented. The relationship of fungi to CaCO3 organomineralization is also mentioned regularly, however the exact mechanisms still remain to be more thoroughly assessed. Therefore, in order to adequately quantify the importance of microbes in CaCO3 biomineralization, the most common biomineral encountered in the biosphere, further efforts should be made to highlight the diversity of fungal processes, as well as their importance in global biogeochemical of both the C and Ca cycles. In addition, it also emphasizes their possible role in the biomineralization of other type of mineral of ecological importance such as phosphate and oxides.
Acknowledgments
The authors would like to thank the Subsidy tremplin at Unine for financial support. Some of the work presented was supported by the Swiss National Science Foundation, Grant Nos. FN 646 205320-109497/1 and FN 205320-122171.
Author Contributions
Conflicts of Interest
The authors declare no conflict of interest.
References
- Weiner, S.; Dove, P.M. An overview of biomineralization processes and the problem of the vital effect. In Biomineralization; Dove, P.M., de Yoreo, J.J., Weiner, S., Eds.; Mineralogical Society of America: Chantilly, VA, USA, 2003; Volume 54, pp. 1–29. [Google Scholar]
- Dupraz, C.; Reid, R.P.; Braissant, O.; Decho, A.W.; Norman, R.S.; Visscher, P.T. Processes of carbonate precipitation in modern microbial mats. Earth Sci. Rev. 2009, 96, 141–162. [Google Scholar]
- Défarge, C. Organomineralization. In Encyclopedia of Geobiology; Reitner, J., Thiel, V., Eds.; Springer: Dodrecht, The Netherlands, 2011; pp. 697–701. [Google Scholar] [Green Version]
- Trichet, J.; Défarge, C. Non-biologically supported organomineralization. Bull. Inst. Océanogr. Monaco Numéro Spec. 1995, 14, 203–236. [Google Scholar]
- Madigan, M.T.; Martinko, J.M.; Bender, K.S.; Buckley, D.H.; Stahl, D.H. Brock Biology of Microorganisms, 14th ed.; Pearson Education, Inc.: Boston, MA, USA, 2015. [Google Scholar]
- Mann, S. Biomineralization: Principles and Concepts in Bioinorganic Materials Chemistry; Oxford University Press, Inc.: New York, NY, USA, 2001. [Google Scholar]
- Bazylinski, D.A.; Frankel, R.B. Biologically controlled mineralization in prokaryotes. Rev. Mineral. Geochem. 2003, 54, 217–247. [Google Scholar]
- Badger, M.R.; Price, G.D. The role of carbonic-anhydrase in photosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1994, 45, 369–392. [Google Scholar] [CrossRef]
- Castanier, S.; le Metayer-Levrel, G.; Perthuisot, J.-P. Bacterial roles in the precipitation of carbonate minerals. In Microbial Sediments; Riding, R.E., Awramik, S.M., Eds.; Springer-Verlag: Berlin, Germany; Heidelberg, Germany, 2000; pp. 32–39. [Google Scholar]
- Braissant, O.; Cailleau, G.; Aragno, M.; Verrecchia, E.P. Biologically induced mineralization in the tree Milicia excelsa (Moraceae): Its causes and consequences to the environment. Geobiology 2004, 2, 59–66. [Google Scholar]
- Visscher, P.T.; Stolz, J.F. Microbial mats as bioreactors: Populations, processes, and products. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 219, 87–100. [Google Scholar]
- Dupraz, C.; Visscher, P.T. Microbial lithification in marine stromatolites and hypersaline mats. Trends Microbiol. 2005, 13, 429–438. [Google Scholar]
- Benzerara, K.; Morin, G.; Yoon, T.H.; Miot, J.; Tyliszczak, T.; Casiot, C.; Bruneel, O.; Farges, F.; Brown, G.E. Nanoscale study of As biomineralization in an acid mine drainage system. Geochim. Cosmochim. Acta 2008, 72, 3949–3963. [Google Scholar] [CrossRef]
- Bindschedler, S.; Cailleau, G.; Braissant, O.; Millière, L.; Job, D.; Verrecchia, E.P. Unravelling the enigmatic origin of calcitic nanofibres in soils and caves: Purely physicochemical or biogenic processes? Biogeosciences 2014, 11, 2809–2825. [Google Scholar] [CrossRef]
- Lowenstam, H.A.; Weiner, S. On Biomineralization; Oxford University Press, Inc.: New York, NY, USA, 1989. [Google Scholar]
- Ehrlich, H.I. Geomicrobiology, 4th ed.; CRC Press: Boca Raton, FL, USA, 2002. [Google Scholar]
- Skinner, H.C.W. Biominerals. Mineral. Mag. 2005, 69, 621–641. [Google Scholar] [CrossRef]
- Mann, S. The chemistry of form. Angew. Chem. Int. Ed. 2000, 39, 3393–3406. [Google Scholar] [CrossRef]
- Ritz, K.; Young, I.M. Interactions between soil structure and fungi. Mycologist 2004, 18, 52–59. [Google Scholar] [CrossRef]
- Sterflinger, K. Fungi as geologic agents. Geomicrobiol. J. 2000, 17, 97–124. [Google Scholar] [CrossRef]
- Gorbushina, A.A. Life on the rocks. Environ. Microbiol. 2007, 9, 1613–1631. [Google Scholar] [CrossRef] [PubMed]
- Vanderwolf, K.J.; Malloch, D.; McAlpine, D.F.; Forbes, G.J. A world review of fungi, yeasts, and slime molds in caves. Int. J. Speleol. 2013, 42, 77–96. [Google Scholar] [CrossRef]
- Shearer, C.A.; Descals, E.; Kohlmeyer, B.; Kohlmeyer, J.; Marvanovà, L.; Padgett, D.; Porter, D.; Raja, H.A.; Schmit, J.P.; Thorton, H.A. Fungal biodiversity in aquatic habitats. Biodivers. Conserv. 2007, 16, 49–67. [Google Scholar] [CrossRef]
- Nagahama, T.; Nagano, Y. Cultured and uncultured fungal diversity in deep-sea environments. In Biology of Marine Fungi; Springer-Verlag: Berlin, Germany; Heidelberg, Germany, 2012; pp. 173–187. [Google Scholar]
- Gadd, G.M. Geomycology: Biogeochemical transformations of rocks, minerals, metals and radionuclides by fungi, bioweathering and bioremediation. Mycol. Res. 2007, 111, 3–49. [Google Scholar] [CrossRef] [PubMed]
- Moore, D.; Robson, G.D.; Trinci, A.P.J. 21st Century Guidebook to Fungi; Cambridge University Press: New York, NY, USA, 2011. [Google Scholar]
- Deacon, J.W. Fungal Biology; Blackwell Publishing Ltd: Malden, MA, USA, 2007. [Google Scholar]
- Klein, D.A.; Paschke, M.W. Filamentous fungi: The indeterminate lifestyle and microbial ecology. Microb. Ecol. 2004, 47, 224–235. [Google Scholar] [CrossRef] [PubMed]
- Jongmans, A.G.; van Breemen, N.; Lundstrom, U.; van Hees, P.A.W.; Finlay, R.D.; Srinivasan, M.; Unestam, T.; Giesler, R.; Melkerud, P.A.; Olsson, M. Rock-eating fungi. Nature 1997, 389, 682–683. [Google Scholar] [CrossRef]
- Van Schöll, L.; Kuyper, T.W.; Smits, M.M.; Landeweert, R.; Hoffland, E.; van Breemen, N. Rock-eating mycorrhizas: Their role in plant nutrition and biogeochemical cycles. Plant Soil 2008, 303, 35–47. [Google Scholar] [CrossRef]
- Boswell, G.P.; Jacobs, H.; Ritz, K.; Gadd, G.M.; Davidson, F.A. The development of fungal networks in complex environments. Bull. Math. Biol. 2007, 69, 605–634. [Google Scholar] [CrossRef] [PubMed]
- Paul, E.A. Soil Microbiology, Ecology and Biochemistry; Academic Press: San Diego, CA, USA, 2007; p. 532. [Google Scholar]
- Jones, P.C.T.; Mollison, J.E.; Quenouille, M.H. A technique for the quantitative estimation of soil micro-organisms. Microbiology 1948, 2, 54–69. [Google Scholar]
- Ritz, K. Growth responses of some soil fungi to spatially heterogeneous nutrients. FEMS Microbiol. Ecol. 1995, 16, 269–279. [Google Scholar] [CrossRef]
- Wright, D.P.; Johansson, T.; Le Quéré, A.; Söderström, B.; Tunlid, A. Spatial patterns of gene expression in the extramatrical mycelium and mycorrhizal root tips formed by the ectomycorrhizal fungus Paxillus involutus in association with birch (Betula pendula) seedlings in soil microcosms. New Phytol. 2005, 167, 579–596. [Google Scholar] [CrossRef] [PubMed]
- Olsson, S. Mycelial density profiles of fungi on heterogeneous media and their interpretation in terms of nutrient reallocation patterns. Mycol. Res. 1995, 99, 143–153. [Google Scholar] [CrossRef]
- Harms, H.; Schlosser, D.; Wick, L.Y. Untapped potential: Exploiting fungi in bioremediation of hazardous chemicals. Nat. Rev. Microbiol. 2011, 9, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Fröhlich-Nowoisky, J.; Burrows, S.; Xie, Z.; Engling, G.; Solomon, P.; Fraser, M.; Mayol-Bracero, O.; Artaxo, P.; Begerow, D.; Conrad, R. Biogeography in the air: Fungal diversity over land and oceans. Biogeosciences 2012, 9, 1125–1136. [Google Scholar] [CrossRef] [Green Version]
- Womack, A.M.; Artaxo, P.E.; Ishida, F.Y.; Mueller, R.C.; Saleska, S.R.; Wiedemann, K.T.; Bohannan, B.J.M.; Green, J.L. Characterization of active and total fungal communities in the atmosphere over the Amazon rainforest. Biogeosciences 2015, 12, 6337–6349. [Google Scholar] [CrossRef]
- Hibbett, D.S.; Binder, M.; Bischoff, J.F.; Blackwell, M.; Cannon, P.F.; Eriksson, O.E.; Huhndorf, S.; James, T.; Kirk, P.M.; Lücking, R. A higher-level phylogenetic classification of the Fungi. Mycol. Res. 2007, 111, 509–547. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis; Academic Press: New York, NY, USA, 2008. [Google Scholar]
- Stajich, J.E.; Berbee, M.L.; Blackwell, M.; Hibbett, D.S.; James, T.Y.; Spatafora, J.W.; Taylor, J.W. The fungi. Curr. Biol. 2009, 19, R840–R845. [Google Scholar] [CrossRef] [PubMed]
- Burford, E.P.; Fomina, M.; Gadd, G.M. Fungal involvement in bioweathering and biotransformation of rocks and minerals. Mineral. Mag. 2003, 67, 1127–1155. [Google Scholar] [CrossRef]
- Gadd, G.M.; Fomina, M.; Burford, E.P. Fungal roles and function in rock, mineral and soil transformations. In Microorganisms and Earth Systems—Advances in Geomicrobiology; Gadd, G.M., Semple, K.T., Lappin-Scott, H.M., Eds.; Cambridge University Press: New York, NY, USA, 2005; pp. 201–231. [Google Scholar]
- Parchert, K.J.; Spilde, M.N.; Porras-Alfaro, A.; Nyberg, A.M.; Northup, D.E. Fungal communities associated with rock varnish in black canyon, New Mexico: Casual inhabitants or essential partners? Geomicrobiol. J. 2012, 29, 752–766. [Google Scholar] [CrossRef]
- Gadd, G.M. Metals, minerals and microbes: Geomicrobiology and bioremediation. Microbiology 2010, 156, 609–643. [Google Scholar] [CrossRef] [PubMed]
- Fomina, M.A.; Alexander, I.J.; Colpaert, J.V.; Gadd, G.M. Solubilization of toxic metal minerals and metal tolerance of mycorrhizal fungi. Soil Biol. Biochem. 2005, 37, 851–866. [Google Scholar] [CrossRef]
- Kolo, K.; Keppens, E.; Preat, A.; Claeys, P. Experimental observations on fungal diagenesis of carbonate substrates. J. Geophys. Res. Biogeosci. 2007, 112. [Google Scholar] [CrossRef]
- Fomina, M.; Burford, E.P.; Hillier, S.; Kierans, M.; Gadd, G.M. Rock-building fungi. Geomicrobiol. J. 2010, 27, 624–629. [Google Scholar] [CrossRef]
- Gadd, G.M. Interactions of fungi with toxic metals. New Phytol. 1993, 124, 25–60. [Google Scholar] [CrossRef]
- Fomina, M.; Hillier, S.; Charnock, J.M.; Melville, K.; Alexander, I.J.; Gadd, G.M. Role of oxalic acid overexcretion in transformations of toxic metal minerals by Beauveria caledonica. Appl. Environ. Microbiol. 2005, 71, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Verrecchia, E.P. Fungi and sediments. In Microbial Sediments; Riding, R.E., Awramik, S.M., Eds.; Springer-Verlag: Berlin, Heidelberg, Germany, 2000. [Google Scholar]
- Hoffland, E.; Kuyper, T.W.; Wallander, H.; Plassard, C.; Gorbushina, A.A.; Haselwandter, K.; Holmstrom, S.; Landeweert, R.; Lundstrom, U.S.; Rosling, A.; et al. The role of fungi in weathering. Front. Ecol. Environ. 2004, 2, 258–264. [Google Scholar] [CrossRef]
- Cromack, K., Jr.; Sollins, P.; Todd, R.L.; Fogel, R.; Todd, A.W.; Fender, W.M.; Crossley, M.E.; Crossley, D.A., Jr. The role of oxalic acid and bicarbonate in calcium cycling by fungi and bacteria: Some possible implications for soil animals. In Proceedings of the VI International Soil Zoology Colloquium, Uppsala, Sweden, 21–25 June 1976.
- Gadd, G.M. Fungal production of citric and oxalic acid: Importance in metal speciation, physiology and biogeochemical processes. In Advances in Microbial Physiology; Academic Press: Cambridge, MA, USA, 1999; Volume 41, pp. 47–92. [Google Scholar]
- Adeyemi, A.O.; Gadd, G.M. Fungal degradation of calcium-, lead-and silicon-bearing minerals. Biometals 2005, 18, 269–281. [Google Scholar] [CrossRef] [PubMed]
- Schilling, J.S.; Jellison, J. Oxalate regulation by two brown rot fungi decaying oxalate-amended and non-amended wood. Holzforschung 2005, 59, 681–688. [Google Scholar] [CrossRef]
- Tuason, M.M.S.; Arocena, J.M. Calcium oxalate biomineralization by Piloderma fallax in response to various levels of calcium and phosphorus. Appl. Environ. Microbiol. 2009, 75, 7079–7085. [Google Scholar] [CrossRef] [PubMed]
- Gadd, G.M. Fungi in Biogeochemical Cycles; Cambridge University Press: Cambridge, UK, 2006; Volume 24. [Google Scholar]
- Gadd, G.M. Bacterial and fungal geomicrobiology: A problem with communities? Geobiology 2008, 6, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Gadd, G.M.; Raven, J.A. Geomicrobiology of eukaryotic microorganisms. Geomicrobiol. J. 2010, 27, 491–519. [Google Scholar] [CrossRef]
- Stumm, W.; Morgan, J.J. Aquatic Chemistry: Chemical Equilibria and Rates in Natural Waters; John Wiley & Sons Inc.: New York, NY, USA, 1996; p. 1022. [Google Scholar]
- Dutton, M.V.; Evans, C.S. Oxalate production by fungi: Its role in pathogenicity and ecology in the soil environment. Can. J. Microbiol. 1996, 42, 881–895. [Google Scholar] [CrossRef]
- Oggerin, M.; Rodriguez, N.; del Moral, C.; Amils, R. Fungal jarosite biomineralization in Rio Tinto. Res. Microbiol. 2014, 165, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Fomina, M.; Podgorsky, V.S.; Olishevska, S.V.; Kadoshnikov, V.M.; Pisanska, I.R.; Hillier, S.; Gadd, G.M. Fungal deterioration of barrier concrete used in nuclear waste disposal. Geomicrobiol. J. 2007, 24, 643–653. [Google Scholar] [CrossRef]
- Fomina, M.; Charnock, J.M.; Hillier, S.; Alvarez, R.; Livens, F.; Gadd, G.M. Role of fungi in the biogeochemical fate of depleted uranium. Curr. Biol. 2008, 18, R375–R377. [Google Scholar] [CrossRef] [PubMed]
- Rhee, Y.J.; Hillier, S.; Gadd, G.M. Lead transformation to pyromorphite by fungi. Curr. Biol. 2012, 22, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Crusberg, T.C. Biomineralization of copper by a fungus revealed by SEM. Microsc. Anal. 2004, 18, 11–13. [Google Scholar]
- Liang, X.; Hillier, S.; Pendlowski, H.; Gray, N.; Ceci, A.; Gadd, G.M. Uranium phosphate biomineralization by fungi. Environ. Microbiol. 2015, 17, 2064–2075. [Google Scholar] [CrossRef] [PubMed]
- Verrecchia, E.P.; Dumont, J.L. A biogeochemical model for chalk alteration by fungi in semiarid environments. Biogeochemistry 1996, 35, 447–470. [Google Scholar] [CrossRef]
- Burford, E.P.; Hillier, S.; Gadd, G.M. Biomineralization of fungal hyphae with calcite (CaCO3) and calcium oxalate mono- and dihydrate in Carboniferous limestone microcosms. Geomicrobiol. J. 2006, 23, 599–611. [Google Scholar] [CrossRef]
- Hou, W.; Dou, C.; Lian, B.; Dong, H. The interaction of fungus with calcite and the effects on aqueous geochemistry in karst systems. Carbonates Evaporites 2013, 28, 413–418. [Google Scholar] [CrossRef]
- Castanier, S.; le Métayer-Levrel, G.L.; Perthuisot, J.-P. Ca-carbonates precipitation and limestone genesis—The microbiogeologist point of view. Sediment. Geol. 1999, 126, 9–23. [Google Scholar] [CrossRef]
- Bowen, A.D.; Davidson, F.A.; Keatch, R.; Gadd, G.M. Induction of contour sensing in Aspergillus niger by stress and its relevance to fungal growth mechanics and hyphal tip structure. Fungal Genet. Biol. 2007, 44, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Verrecchia, E.P.; Braissant, O.; Cailleau, G. The oxalate-carbonate pathway in soil carbon storage: The role of fungi and oxalotrophic bacteria. In Fungi in Biogeochemical Cycles; Cambridge University Press: New York, NY, USA, 2006; pp. 289–310. [Google Scholar]
- Guggiari, M.; Bloque, R.; Aragno, M.; Verrecchia, E.; Job, D.; Junier, P. Experimental calcium-oxalate crystal production and dissolution by selected wood-rot fungi. Int. Biodeterior. Biodegrad. 2011, 65, 803–809. [Google Scholar] [CrossRef]
- Burbank, M.B.; Weaver, T.J.; Green, T.L.; Williams, B.C.; Crawford, R.L. Precipitation of calcite by indigenous microorganisms to strengthen liquefiable soils. Geomicrobiol. J. 2011, 28, 301–312. [Google Scholar] [CrossRef]
- Li, Q.; Csetenyi, L.; Gadd, G.M. Biomineralization of metal carbonates by Neurospora crassa. Environ. Sci. Technol. 2014, 48, 14409–14416. [Google Scholar] [CrossRef] [PubMed]
- Hou, W.; Lian, B.; Zhang, X. CO2 mineralization induced by fungal nitrate assimilation. Bioresour. Technol. 2011, 102, 1562–1566. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.L.; Heath, I.B. Roles of calcium-ions in hyphal tip growth. Microbiol. Rev. 1993, 57, 367–382. [Google Scholar] [PubMed]
- Pitt, D.; Ugalde, U.O. Calcium in fungi. Plant Cell Environ. 1984, 7, 467–475. [Google Scholar] [CrossRef]
- Manoli, F.; Koutsopoulos, S.; Dalas, E. Crystallization of calcite on chitin. J. Cryst. Growth 1997, 182, 116–124. [Google Scholar] [CrossRef]
- Whitney, K. Systems of biomineralization in the fungi. In Origin, Evolution and Modern Aspects of Biomineralization in Plants and Animals; Crick, R.E., Ed.; Springer-Verlag: Berlin, Heidelberg, Germany, 1989. [Google Scholar]
- Simkiss, K. The processes of biomineralization in lower plants and animals—An overview. Biominer. Lower Plants Anim. 1986, 30, 19–37. [Google Scholar]
- Schultze-Lam, S.; Fortin, D.; Davis, B.S.; Beveridge, T.J. Mineralization of bacterial surfaces. Chem. Geol. 1996, 132, 171–181. [Google Scholar] [CrossRef]
- Barton, H.A.; Northup, D.E. Geomicrobiology in cave environments: Past, current and future perspectives. J. Cave Karst Stud. 2007, 69, 163–178. [Google Scholar]
- Klappa, C. Calcified filaments in Quaternary calcretes: Organo-mineral interactions in the subaerial vadose environment. J. Sediment. Petrol. 1979, 49, 955–968. [Google Scholar] [CrossRef]
- Wright, V.P. The role of fungal biomineralization in the formation of Early Carboniferous soil fabrics. Sedimentology 1986, 33, 831–838. [Google Scholar] [CrossRef]
- Monger, H.C.; Daugherty, L.A.; Lindemann, W.C.; Liddell, C.M. Microbial precipitation of pedogenic calcite. Geology 1991, 19, 997–1000. [Google Scholar] [CrossRef]
- Cailleau, G.; Verrecchia, E.P.; Braissant, O.; Emmanuel, L. The biogenic origin of needle fibre calcite. Sedimentology 2009, 56, 1858–1875. [Google Scholar] [CrossRef]
- Bindschedler, S.; Millière, L.; Cailleau, G.; Job, D.; Verrecchia, E.P. Calcitic Nanofibres in Soils and Caves: A Putative Fungal Contribution to Carbonatogenesis; Geological Society, London, Special Publications: London, UK, 2010; Volume 336, pp. 225–238. [Google Scholar]
- Bindschedler, S.; Millière, L.; Cailleau, G.; Job, D.; Verrecchia, E.P. An ultrastructural approach to analogies between fungal structures and needle fiber calcite. Geomicrobiol. J. 2012, 29, 301–313. [Google Scholar] [CrossRef]
- Li, Q.; Csetenyi, L.; Paton, G.I.; Gadd, G.M. CaCO3 and SrCO3 bioprecipitation by fungi isolated from calcareous soil. Environ. Microbiol. 2015, 17, 3082–3097. [Google Scholar] [CrossRef] [PubMed]
- Addadi, L.; Raz, S.; Weiner, S. Taking advantage of disorder: Amorphous calcium carbonate and its roles in biomineralization. Adv. Mater. 2003, 15, 959–970. [Google Scholar] [CrossRef]
- Benzerara, K.; Menguy, N.; Guyot, F.; Dominici, C.; Gillet, P. Nanobacteria-like calcite single crystals at the surface of the tataouine meteorite. Proc. Natl. Acad. Sci. USA 2003, 100, 7438–7442. [Google Scholar] [CrossRef]
- Faatz, M.; Gröhn, F.; Wegner, G. Amorphous calcium carbonate: Synthesis and potential intermediate in biomineralization. Adv. Mater. 2004, 16, 996–1000. [Google Scholar] [CrossRef]
- Rautaray, D.; Ahmad, A.; Sastry, M. Biosynthesis of CaCO3 crystals of complex morphology using a fungus and an actinomycete. J. Am. Chem. Soc. 2003, 125, 14656–14657. [Google Scholar] [CrossRef] [PubMed]
- Rautaray, D.; Ahmad, A.; Sastry, M. Biological synthesis of metal carbonate minerals using fungi and actinomycetes. J. Mater. Chem. 2004, 14, 2333–2340. [Google Scholar] [CrossRef]
- Ahmad, A.; Rautaray, D.; Sastry, M. Biogenic calcium carbonate: Calcite crystals of variable morphology by the reaction of aqueous Ca2+ ions with fungi. Adv. Funct. Mater. 2004, 14, 1075–1080. [Google Scholar] [CrossRef]
- Sanyal, A.; Rautaray, D.; Bansal, V.; Ahmad, A.; Sastry, M. Heavy-metal remediation by a fungus as a means of production of lead and cadmium carbonate crystals. Langmuir 2005, 21, 7220–7224. [Google Scholar] [CrossRef] [PubMed]
- Masaphy, S.; Zabari, L.; Pastrana, J.; Dultz, S. Role of fungal mycelium in the formation of carbonate concretions in growing media—An investigation by SEM and synchrotron-based X-ray tomographic microscopy. Geomicrobiol. J. 2009, 26, 442–450. [Google Scholar] [CrossRef]
- Verrecchia, E.P.; Dumont, J.L.; Rolko, K.E. Do fungi building limestones exist in semi-arid regions? Naturwissenschaften 1990, 77, 584–586. [Google Scholar] [CrossRef]
- Verrecchia, E.P.; Verrecchia, K.E. Needle-fiber calcite—A critical-review and a proposed classification. J. Sediment. Res. Sect. Sediment. Petrol. Processes 1994, 64, 650–664. [Google Scholar]
- Cailleau, G.; Dadras, M.; Abolhassani-Dadras, S.; Braissant, O.; Verrecchia, E.P. Evidence for an organic origin of pedogenic calcitic nanofibres. J. Cryst. Growth 2009, 311, 2490–2495. [Google Scholar] [CrossRef]
- Millière, L.; Hasinger, O.; Bindschedler, S.; Cailleau, G.; Spangenberg, J.E.; Verrecchia, E.P. Stable carbon and oxygen isotope signatures of pedogenic needle fibre calcite. Geoderma 2011, 161, 74–87. [Google Scholar] [CrossRef]
- Kubiëna, W.L. Micropedology. Soil Sci. 1939, 47, 163. [Google Scholar] [CrossRef]
- Verges, V.; Madon, M.; Bruand, A.; Bocquier, G. Morphologie et cristallogenèse de microcristaux supergènes de calcite en aiguilles. Bull. Minéral. 1982, 105, 351–356. (In French) [Google Scholar]
- Jones, B.; Ng, K.C. The structure and diagenesis of rhizoliths from Cayman Brac, British West Indies. J. Sediment. Petrol. 1988, 58, 457–467. [Google Scholar]
- Callot, G.; Guyon, A.; Mousain, D. Relation between calcite needles and fungal hyphae in the soil. Agronomie 1985, 5, 209–216. [Google Scholar] [CrossRef]
- Phillips, S.E.; Self, P.G. Morphology, crystallography and origin of needle-fiber calcite in quaternary pedogenic calcretes of South Australia. Aust. J. Soil Res. 1987, 25, 429–444. [Google Scholar] [CrossRef]
- Ould Mohamed, S.; Bruand, A. Morphology and origin of secondary calcite in soils from Beauce, France. Dev. Soil Sci. 1993, 22, 27–36. [Google Scholar]
- Borsato, A.; Frisia, S.; Jones, B.; Van der Borg, K. Calcite moonmilk: Crystal morphology and environment of formation in caves in the Italian Alps. J. Sediment. Res. 2000, 70, 1171–1182. [Google Scholar] [CrossRef]
- Javaux, E.J. Biomarkers. In Encyclopedia of Astrobiology; Springer-Verlag: Berlin, Germany; Heidelberg, Germany, 2011; pp. 182–183. [Google Scholar]
- Simoneit, B.R.T. A review of current applications of mass spectrometry for biomarker/molecular tracer elucidations. Mass Spectrom. Rev. 2005, 24, 719–765. [Google Scholar] [CrossRef] [PubMed]
- Simkiss, K.; Wilbur, K.M. Biomineralization: Cell Biology and Mineralization; Academic Press: New York, NY, USA, 1989. [Google Scholar]
- Stumm, W. Chemistry of the Solid-Water Interface: Processes at the Mineral-Water and Particle-Water Interface in Natural Systems; John Wiley & Son Inc.: New York, NY, USA, 1992. [Google Scholar]
- Barton, H.A.; Spear, J.R.; Pace, N.R. Microbial life in the underworld: Biogenicity in secondary mineral formations. Geomicrobiol. J. 2001, 18, 359–368. [Google Scholar]
- Blyth, A.J.; Frisia, S. Molecular evidence for bacterial mediation of calcite formation in cold high-altitude caves. Geomicrobiol. J. 2008, 25, 101–111. [Google Scholar] [CrossRef]
- Benzerara, K.; Menguy, N. Looking for traces of life in minerals. Comptes Rendus Palevol 2009, 8, 617–628. [Google Scholar] [CrossRef]
- Yoon, T.H. Applications of soft X-ray spectromicroscopy in material and environmental sciences. Appl. Spectrosc. Rev. 2009, 44, 91–122. [Google Scholar] [CrossRef]
- Gilbert, P.; Abrecht, M.; Frazer, B.H. The organic-mineral interface in biominerals. Rev. Mineral. Geochem. 2005, 59, 157–185. [Google Scholar]
- Cavalazzi, B. Global to nano-scale relevance of Ca-carbonate biosignatures. Boll. Soc. Paleontol. Ital. 2013, 52, 139–140. [Google Scholar]
- Smits, M.M.; Herrmann, A.M.; Duane, M.; Duckworth, O.W.; Bonneville, S.; Benning, L.G.; Lundström, U. The fungal-mineral interface: Challenges and considerations of micro-analytical developments. Fungal Biol. Rev. 2009, 23, 122–131. [Google Scholar]
- Dykstra, M.J.; Reuss, L.E. Biological Electron Microscopy: Theory, Techniques, and Troubleshooting; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2011. [Google Scholar]
- Canaveras, J.C.; Hoyos, M.; Sanchez-Moral, S.; Sanz-Rubio, E.; Bedoya, J.; Soler, V.; Groth, I.; Schumann, P.; Laiz, L.; Gonzalez, I.; et al. Microbial communities associated with hydromagnesite and needle-fiber aragonite deposits in a karstic cave (Altamira, Northern Spain). Geomicrobiol. J. 1999, 16, 9–25. [Google Scholar]
- Benzerara, K.; Menguy, N.; Lopez-Garcia, P.; Yoon, T.H.; Kazmierczak, J.; Tyliszczak, T.; Guyot, F.; Brown, G.E. Nanoscale detection of organic signatures in carbonate microbialites. Proc. Natl. Acad. Sci. USA 2006, 103, 9440–9445. [Google Scholar]
- Benzerara, K.; Miller, V.M.; Barell, G.; Kumar, V.; Miot, J.; Brown, G.E.; Lieske, J.C. Search for microbial signatures within human and microbial calcifications using soft X-ray spectromicroscopy. J. Investig. Med. 2006, 54, 367–379. [Google Scholar] [CrossRef]
- Obst, M.; Wang, J.; Hitchcock, A.P. Soft X-ray spectro-tomography study of cyanobacterial biomineral nucleation. Geobiology 2009, 7, 577–591. [Google Scholar] [PubMed]
- Benzerara, K.; Yoon, T.H.; Tyliszczak, T.; Constantz, B.; Spormann, A.M.; Brown, G.E. Scanning transmission X-ray microscopy study of microbial calcification. Geobiology 2004, 2, 249–259. [Google Scholar] [CrossRef]
- Wagner, M. Single-cell ecophysiology of microbes as revealed by Raman microspectroscopy or secondary ion mass spectrometry imaging. Annu. Rev. Microbiol. 2009, 63, 411–429. [Google Scholar] [CrossRef] [PubMed]
- Kilburn, M.R.; Wacey, D. Nanoscale secondary ion mass spectrometry (NanoSIMS) as an analytical tool in the geosciences. In Principles and practive of analytical techniques in geosciences; Grice, K., Ed.; The Royal Society of Chemistry: Cambridge, UK, 2015; pp. 1–34. [Google Scholar]
- Anderson, I.C.; Cairney, J.W. Diversity and ecology of soil fungal communities: Increased understanding through the application of molecular techniques. Environ. Microbiol. 2004, 6, 769–779. [Google Scholar] [CrossRef] [PubMed]
- Peay, K.G.; Kennedy, P.G.; Bruns, T.D. Fungal community ecology: A hybrid beast with a molecular master. Bioscience 2008, 58, 799–810. [Google Scholar] [CrossRef]
- Lindahl, B.D.; Nilsson, R.H.; Tedersoo, L.; Abarenkov, K.; Carlsen, T.; Kjoller, R.; Koljalg, U.; Pennanen, T.; Rosendahl, S.; Stenlid, J.; et al. Fungal community analysis by high-throughput sequencing of amplified markers : A user’s guide. New Phytol. 2013, 199, 288–299. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, M.J.; Nelson, W.; Soderlund, C.; Maier, R.M.; Pryor, B.M. Assessing fungal community structure from mineral surfaces in Kartchner caverns using multiplexed 454 pyrosequencing. Microb. Ecol. 2015, 70, 175–187. [Google Scholar] [CrossRef] [PubMed]
- Jurado, V.; Fernandez-Cortes, A.; Cuezva, S.; Laiz, L.; Canaveras, J.C.; Sanchez-Moral, S.; Saiz-Jimenez, C. The fungal colonisation of rock-art caves: Experimental evidence. Naturwissenschaften 2009, 96, 1027–1034. [Google Scholar] [CrossRef] [PubMed]
- Seifert, K.A. Progress towards DNA barcoding of fungi. Mol. Ecol. Resour. 2009, 9, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Porter, T.M.; Golding, G.B. Factors that affect large subunit ribosomal DNA amplicon sequencing studies of fungal communities: Classification method, primer choice, and error. PLoS ONE 2012, 7, e35749. [Google Scholar] [CrossRef]
Figure 1.
(A) Hyphal filamentous structure showing cells arranged one after the other, with the formation of ramification by cell branching; (B) Typical two-dimensional morphology of a mycelium as a result of the hyphal branched pattern.
Figure 1.
(A) Hyphal filamentous structure showing cells arranged one after the other, with the formation of ramification by cell branching; (B) Typical two-dimensional morphology of a mycelium as a result of the hyphal branched pattern.

Figure 2.
Simplified phylogenetic tree of Fungi showing the five main phyla currently defined. For each phylum, one typical morphological example is shown.
Figure 2.
Simplified phylogenetic tree of Fungi showing the five main phyla currently defined. For each phylum, one typical morphological example is shown.

Figure 3.
Sketch showing the involvement of fungi in global some biogeochemical cycles; (a) Fungi contribute substantially to mineral weathering, leading to the release of bioavailable metals or nutrients, which eventually may be uptaken by living organisms or precipitated as secondary minerals; (b) Fungi as heterotrophs, recycle organic matter (OM). While doing so, they produce metabolites such as organic acids that can also precipitate as secondary minerals (salts). OM recycling eventually releases constitutive elements such as C, N, P and S; (c) CO2 produced by heterotrophic fungal respiration can dissolve into H2O and depending on the physicochemical conditions precipitate as CaCO3 leading to the formation of a secondary mineral.
Figure 3.
Sketch showing the involvement of fungi in global some biogeochemical cycles; (a) Fungi contribute substantially to mineral weathering, leading to the release of bioavailable metals or nutrients, which eventually may be uptaken by living organisms or precipitated as secondary minerals; (b) Fungi as heterotrophs, recycle organic matter (OM). While doing so, they produce metabolites such as organic acids that can also precipitate as secondary minerals (salts). OM recycling eventually releases constitutive elements such as C, N, P and S; (c) CO2 produced by heterotrophic fungal respiration can dissolve into H2O and depending on the physicochemical conditions precipitate as CaCO3 leading to the formation of a secondary mineral.

Figure 4.
Scanning Electron Microscope (SEM) images of fungal hyphae (white arrow) coated with calcium oxalate crystals (probably wheddelite). Both images (A) and (B) are from a sample of plant root associated to fungal hyphae (possibly mycorrhizal fungi) in a secondary CaCO3 deposit of a calcic Cambisol humic calcaric skeletic soil (WRB 2006) in Villiers, Switzerland. Detailed procedure of sample treatment prior to electron microscopy and operation of SEM can be found in [14].
Figure 4.
Scanning Electron Microscope (SEM) images of fungal hyphae (white arrow) coated with calcium oxalate crystals (probably wheddelite). Both images (A) and (B) are from a sample of plant root associated to fungal hyphae (possibly mycorrhizal fungi) in a secondary CaCO3 deposit of a calcic Cambisol humic calcaric skeletic soil (WRB 2006) in Villiers, Switzerland. Detailed procedure of sample treatment prior to electron microscopy and operation of SEM can be found in [14].

Figure 5.
Processes potentially leading to fungal CaCO3 biomineralization at the fungal microenvironment scale. These processes are mainly linked to both Ca2+ sequestration and metabolic control of carbonate alkalinity within cell compartments (cell wall (cw), cytoplasm (cy), and organelles) of fluids translocated inside the hypha. (i) Carbonate alkalinity levels in the fungal microenvironment influence Ca2+ and CO32− bioavailability; (ii) Ca2+ sequestration rate by fungi in the different cellular compartments influences Ca2+ bioavailability. Ca2+ can be present within the cell-wall (Ca2+-cw) as free cytoplasmic Ca2+ bounded to proteins (Ca2+-B) or stored in organelles. Finally, fungi may exert a metabolic control on intra-hyphal alkalinity levels (represented as CO32−), through pH regulation for instance (e.g., H+ excretion).
Figure 5.
Processes potentially leading to fungal CaCO3 biomineralization at the fungal microenvironment scale. These processes are mainly linked to both Ca2+ sequestration and metabolic control of carbonate alkalinity within cell compartments (cell wall (cw), cytoplasm (cy), and organelles) of fluids translocated inside the hypha. (i) Carbonate alkalinity levels in the fungal microenvironment influence Ca2+ and CO32− bioavailability; (ii) Ca2+ sequestration rate by fungi in the different cellular compartments influences Ca2+ bioavailability. Ca2+ can be present within the cell-wall (Ca2+-cw) as free cytoplasmic Ca2+ bounded to proteins (Ca2+-B) or stored in organelles. Finally, fungi may exert a metabolic control on intra-hyphal alkalinity levels (represented as CO32−), through pH regulation for instance (e.g., H+ excretion).

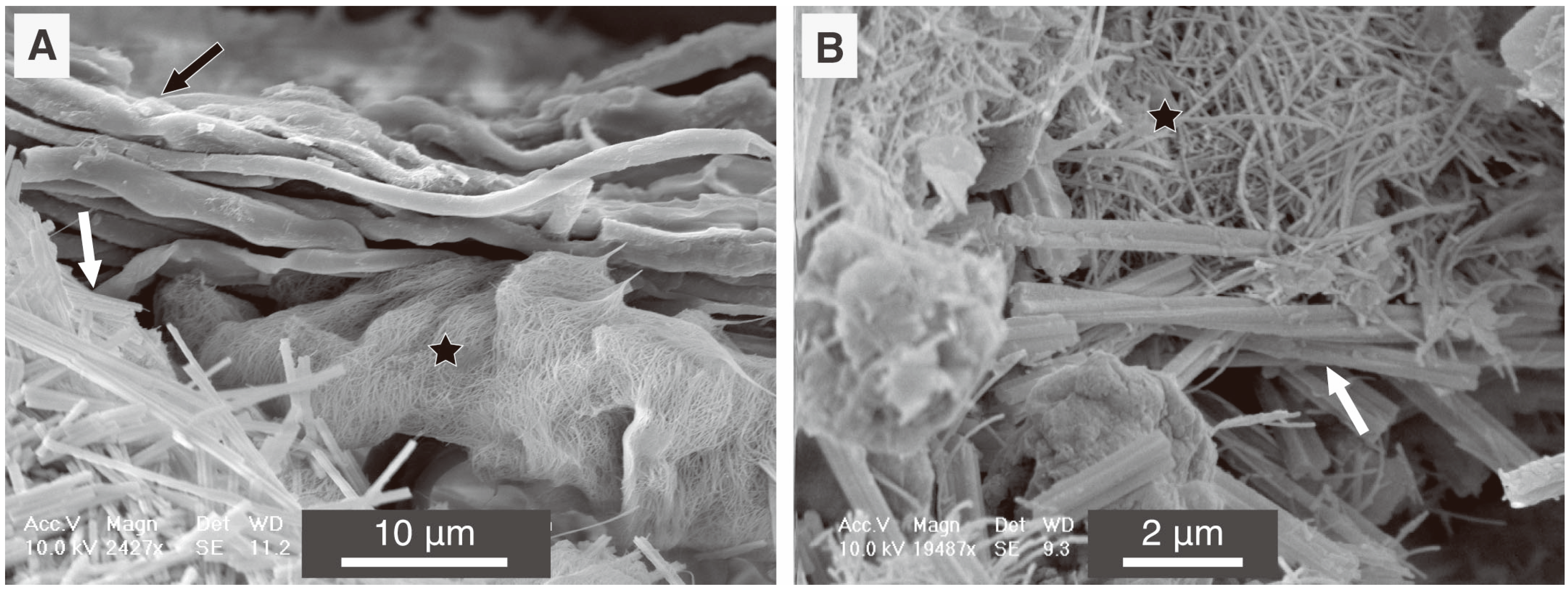
Figure 6.
Scanning Electron Microscope (SEM) images of natural samples of secondary CaCO3 deposit composed of needle fibre calcite (NFC; white arrows) and nanofibres (black stars) from a calcic Cambisol humic calcaric skeletic soil (WRB 2006) in Villiers, Switzerland. Both images (A) and (B) are from a fungal rhizomorph (fungal hyphae are shown in A by a black arrow) associated to a limestone fragment from deep mineral layers of a soil covered with secondary CaCO3 deposits.
Figure 6.
Scanning Electron Microscope (SEM) images of natural samples of secondary CaCO3 deposit composed of needle fibre calcite (NFC; white arrows) and nanofibres (black stars) from a calcic Cambisol humic calcaric skeletic soil (WRB 2006) in Villiers, Switzerland. Both images (A) and (B) are from a fungal rhizomorph (fungal hyphae are shown in A by a black arrow) associated to a limestone fragment from deep mineral layers of a soil covered with secondary CaCO3 deposits.

Figure 7.
Transmission electron microscopy (TEM) images of fungal rhizomorphs and associated EDS spectra. Analysed zones are indicated with a star. Cu is from the TEM grid. Fungal rhizomorphs sampled in a calcic Cambisol humic calcaric skeletic soil (WRB 2006) in Villiers, Switzerland at the interface between the B and C horizons, showing the presence of calcium in both their cell wall (left-hand side) and in intrahyphal inclusions (right-hand side). Os peak indicates that inclusions also contain organic material (see [91] for further details).
Figure 7.
Transmission electron microscopy (TEM) images of fungal rhizomorphs and associated EDS spectra. Analysed zones are indicated with a star. Cu is from the TEM grid. Fungal rhizomorphs sampled in a calcic Cambisol humic calcaric skeletic soil (WRB 2006) in Villiers, Switzerland at the interface between the B and C horizons, showing the presence of calcium in both their cell wall (left-hand side) and in intrahyphal inclusions (right-hand side). Os peak indicates that inclusions also contain organic material (see [91] for further details).

© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bindschedler, S.; Cailleau, G.; Verrecchia, E. Role of Fungi in the Biomineralization of Calcite. Minerals 2016, 6, 41. https://doi.org/10.3390/min6020041
AMA Style
Bindschedler S, Cailleau G, Verrecchia E. Role of Fungi in the Biomineralization of Calcite. Minerals. 2016; 6(2):41. https://doi.org/10.3390/min6020041
Chicago/Turabian StyleBindschedler, Saskia, Guillaume Cailleau, and Eric Verrecchia. 2016. "Role of Fungi in the Biomineralization of Calcite" Minerals 6, no. 2: 41. https://doi.org/10.3390/min6020041
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.