Mineralogical and Elemental Composition of Pectinatella magnifica and Its Statoblasts

Abstract
:1. Introduction
2. Materials and Methods
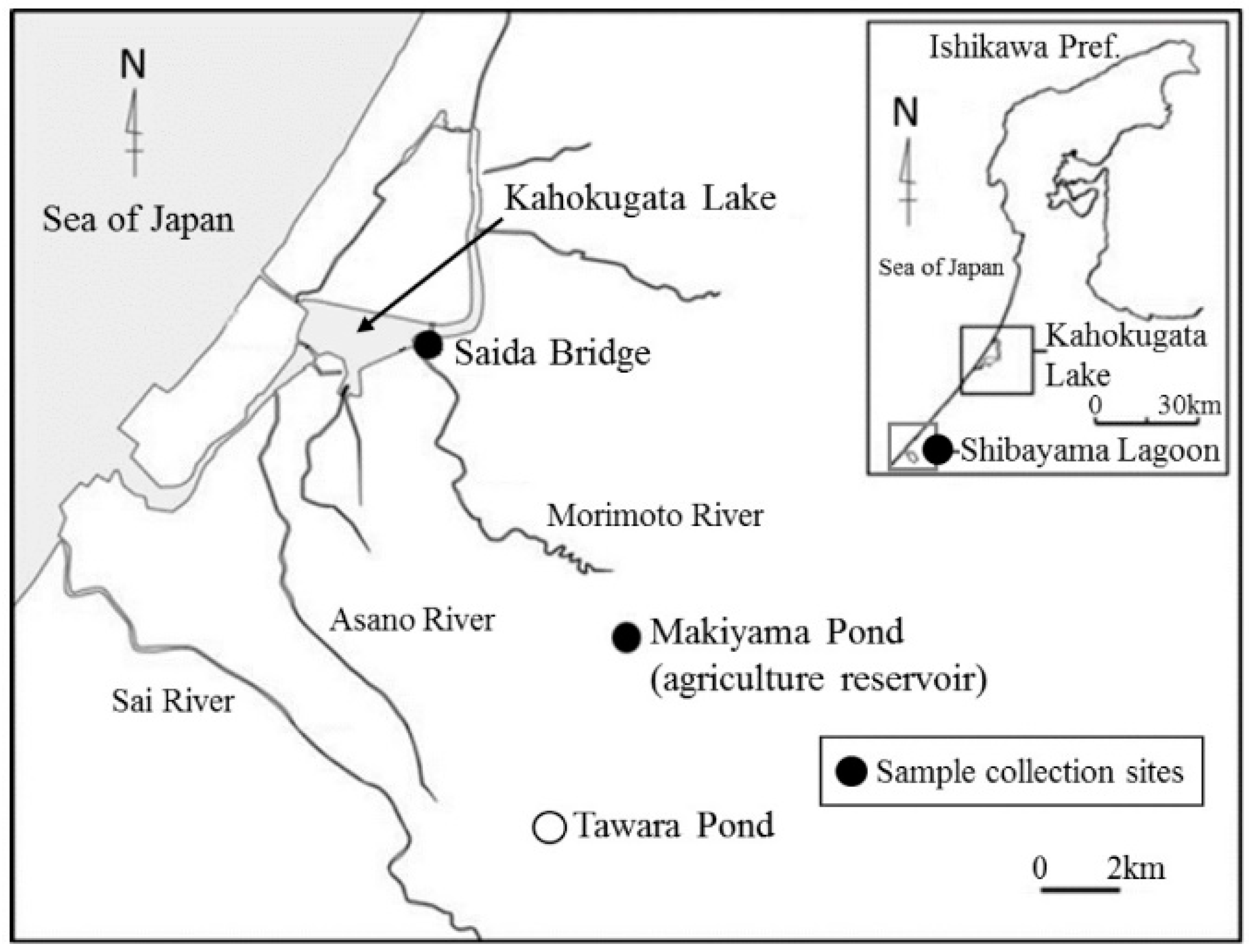
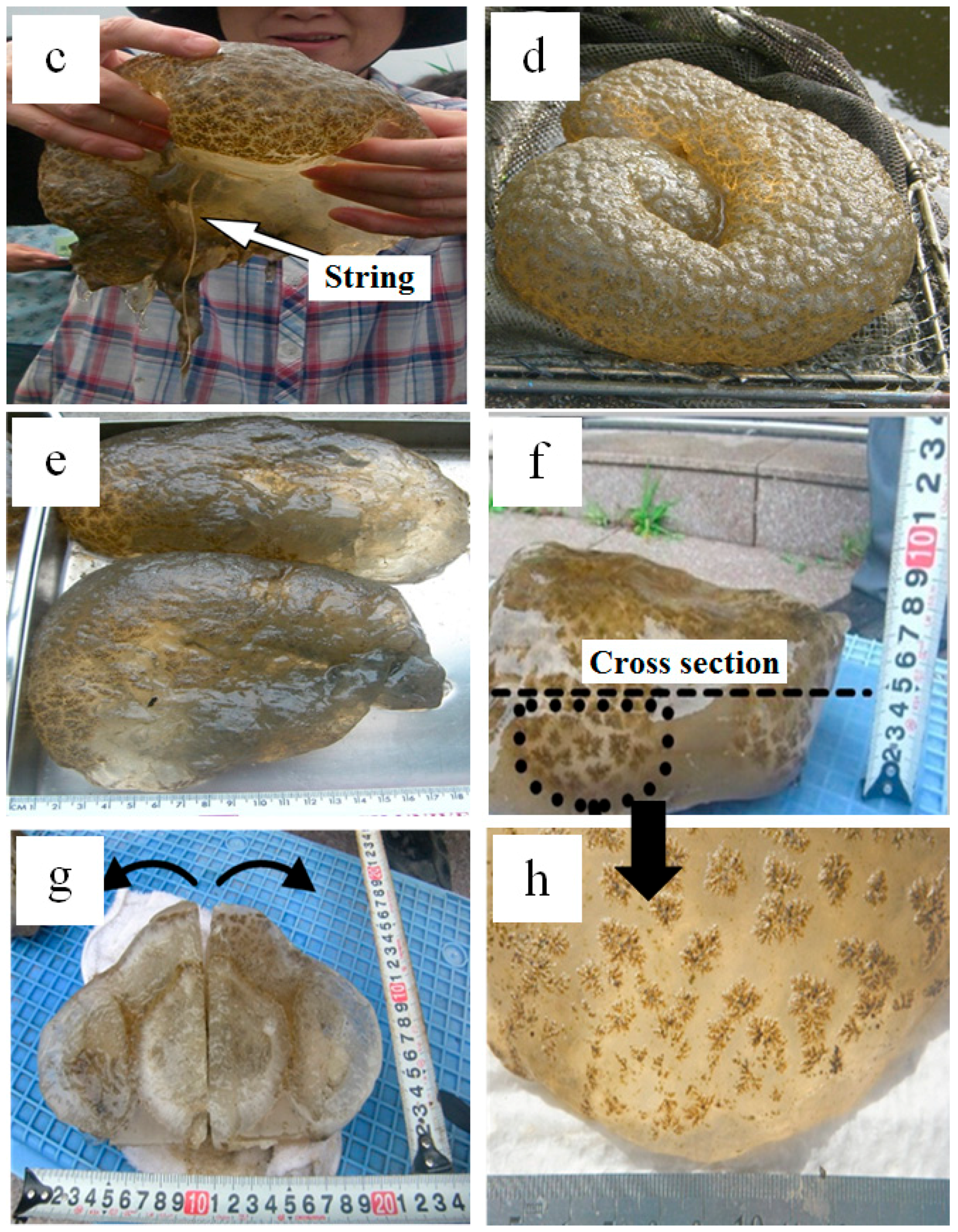
2.1. Exploring Pectinatella magnifica Field and Investigated Specimens
2.2. ICP-MS Analyses
2.3. Scanning Electron Microscopy (SEM) with Energy-Dispersive Spectroscopy (EDS)
2.4. X-Ray Powder Diffraction Measurements
2.5. X-Ray Fluorescence Analyses (XRF) for Deposited Minerals on Makiyama Pond
3. Results
3.1. Optical Microscopy Observation and Identification of Species
3.2. ICP-MS Analyses of Pectinatella magnifica
3.3. SEM Micrograph
3.4. SEM Observation and Energy-Dispersive Spectroscopic (EDS) Analyses
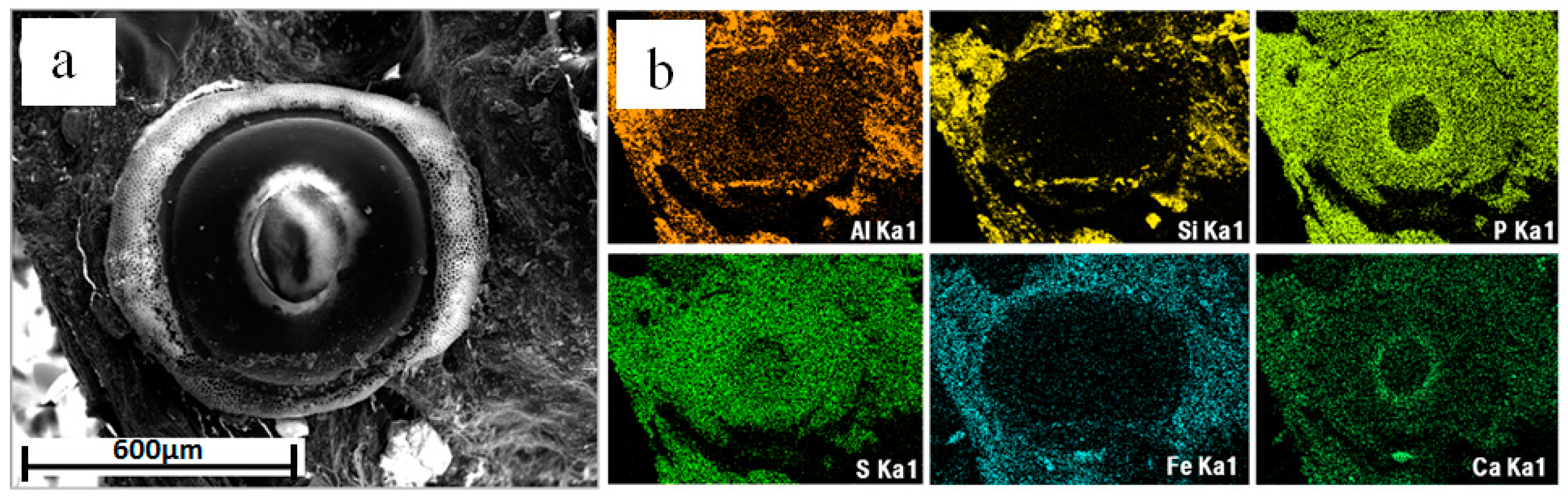
3.5. SEM Elemental Distribution Maps
3.6. X-Ray Powder Diffraction (XRD) for Mineralogy of Pectinatella magnifica in Makiyama Agriculture Reservoir Pond
3.7. X-Ray Fluorescence Analyses (XRF) for Deposited Minerals in Makiyama Agricultural Reservoir Pond
4. Discussion
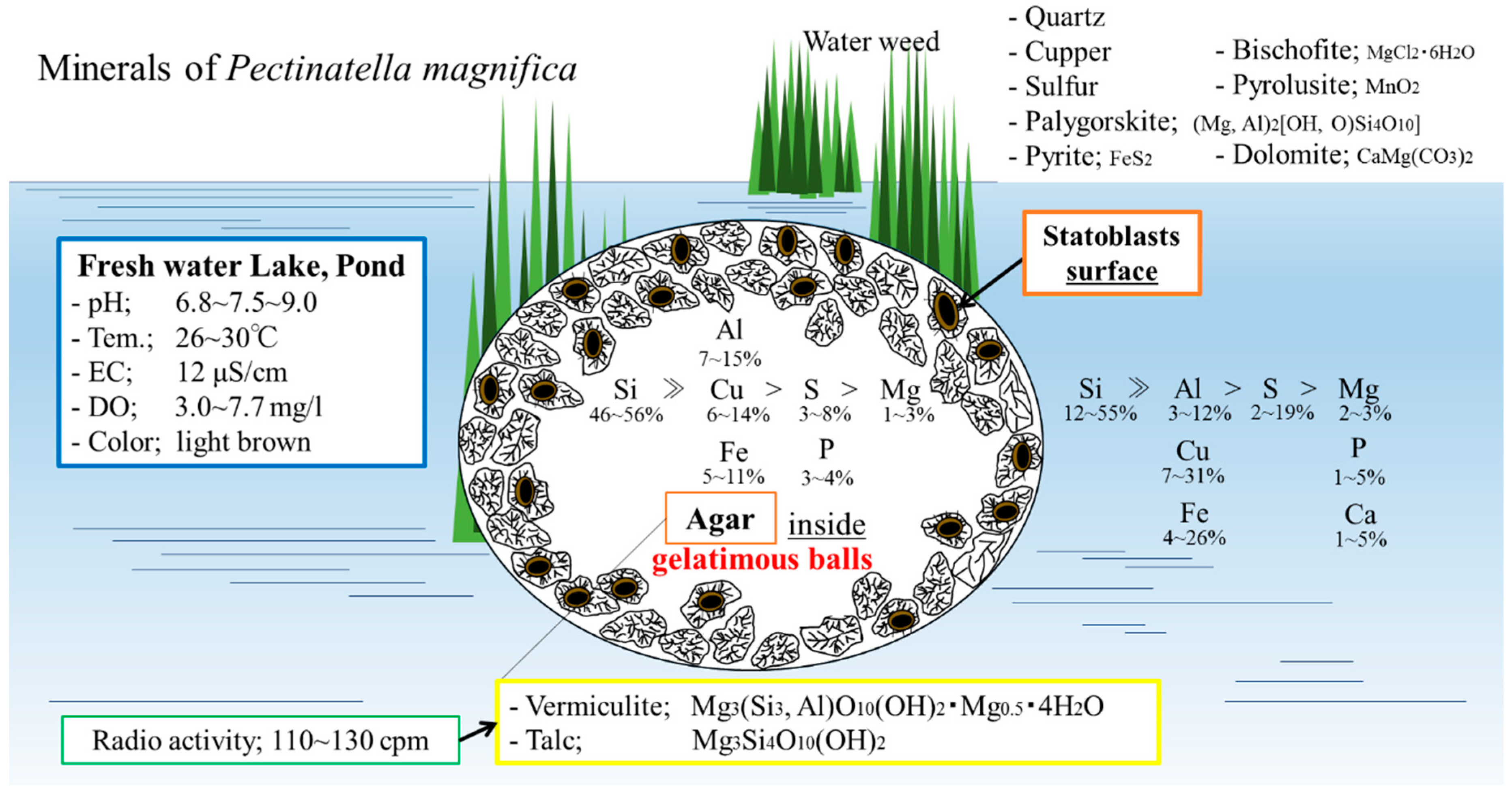
4.1. Characteristics of Statoblasts
4.2. Mineralogical and Elemental Composition of Pectinatella magnifica and Its Statoblasts
4.3. Distribution of Elements and Minerals inside the Bryozoan Colony
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Mawatari, S. New occurrence of Pectinatella magnifica (Leidy) in Japanese Lake. Proc. Jpn. Soc. Syst. Zool. 1973, 9, 41–43. [Google Scholar]
- Oda, S. Pectinatella magnifica occurring in Lake Shoji, Japan. Proc. Jpn. Soc. Syst. Zool. 1974, 10, 31–39. [Google Scholar]
- Oda, S. Notes on Pectinatella magnifica occurring in Japan. Collect. Breed. Tokyo 1975, 37, 40–53. (In Japanese) [Google Scholar]
- Oda, S. A life of Pectinatella magnifica a freshwater bryozoan. Ibid 1976, 38, 434–443. (In Japanese) [Google Scholar]
- Oda, S. A note on bryozoan in Lake Shoji, Japan. Proc. Jpn. Soc. Syst. Zool. 1978, 15, 19–23. [Google Scholar]
- Hirose, M. Taxonomy as a basis and a goal of zoology, with some examples of bryozoology in museums. TAXA Proc. Jpn. Soc. Syst. Zool. 2013, 34, 25–31. [Google Scholar]
- Hirose, M. Taxonomy of Lophophorata; Description of diversity of morphology and biology of bryozoans. TAXA Proc. Jpn. Soc. Syst. Zool. 2014, 37, 1–13. [Google Scholar]
- Smith, A.M.; Key, M.M., Jr.; Gordon, D.P. Skeletal mineralogy of bryozoans: Taxonomic and temporal patterns. Earth Sci. Rev. 2010, 78, 271–277. [Google Scholar] [CrossRef]
- Taylor, P.D.; Lombardi, C.; Cocito, S. Biomineralization in bryozoans: Present, past, future. Biol. Rev. 2015, 90, 1118–1150. [Google Scholar] [CrossRef] [PubMed]
- Withrow, M. The chemical constitution of Pectinatella. Science 1930, 71, 265. [Google Scholar]
- Tazaki, K.; Brandon, K.; Yamamoto, S.; Sato, M.; Hayashi, T.; Takehara, T. Strontium sorption in Phytoplankton biomass and Pectinatell magnifica; Implications for bioremediation. Radioisotopes 2016, 65, 383–392. [Google Scholar] [CrossRef]
- Hirose, M. Taxonomy and identification of freshwater bryozoans in Japan. TAXA Proc. Jpn. Soc. Syst. Zool. 2012, 33, 17–32. [Google Scholar]
- Miyashita, M.; Harada, M.; Noguch, M. First Report of Magnificent Bryozoan Pectinatella magnifica (LEIDY) in Wakayama Prefecture and Its Distribution at Kinokawa and Iwada Cities; Memoirs Institute of Advanced Technology, Kinki University: Osaka, Japan, 2014; Volume 19, pp. 1–12. [Google Scholar]
- Hashida, Y.; Hashida, S.; Tazaki, K.; Nakamura, I.; Takehara, H.; Nakamura, K. Environmental activity and the ecotourism at Makiyama, Kanazawa, Ishikawa in Japan. Kahokugata Lake Sci. 2017, 20, 19–31. [Google Scholar]
- Mukai, H. Germination of the statoblasts of freshwater bryozoan, Pectinatella gelatinosa. J. Exp. Zool. 1974, 183, 27–40. [Google Scholar] [CrossRef]
- Oda, S. Freshwater bryozoan in Japan. Biol. Jpn. 1990, 4, 50–57. [Google Scholar]
- Mawatari, S. USA originated Pectinatella magnifica (LEIOY) found in Japan. Anim. Taxon. 1973, 9, 41–44. [Google Scholar]
- Kaneko, Y.; Iwatsuki, K.; Noda, Y. Pectinatella magnifica in Kagawa Prefecture. J. Biol. Kagawa 1994, 21, 63–66. [Google Scholar]
- Hirose, M.; Ikezawa, H.; Kanek, N.; Mawatari, S. Distribution of freshwater bryozoans in Ibaragi Prefecture. Ibaragi Nat. Mus. Rep. 2006, 9, 31–46. [Google Scholar]
- Iwasaki, K. Introduced sessile animals and invasive alien species act in Japan. Sess. Org. 2006, 23, 13–24. [Google Scholar]
- Oda, S. Germination of the statoblasts in freshwater Gryozoa. Sci. Rep. Tokyo Kyoiku Daigaku B 1959, 9, 90–132. [Google Scholar]
- Robin, N.; Bermard, S.; Miot, I.; Blanc-Valleron, M.M.; Charbonnier, S.; Petit, G. Calcification and diagenesis of bacterial colonies. Minerals 2015, 5, 488–506. [Google Scholar] [CrossRef]
- Bindschedler, S.; Cailleau, G.; Verrecchia, E. Role of Fungi in the biomineralization of calcite. Minerals 2016, 6, 41. [Google Scholar] [CrossRef]
- Simkiss, K.; Wilbur, K.M. Biomineralization: Cell Biology and Mineral Deposition; Academic Press Inc.: Waltham, MA, USA, 1989; p. 337. [Google Scholar]
- Krumbein, W.E. Microbial Geochemistry; Blackwell Scientific Publications: Oxford, UK, 1983; p. 330. [Google Scholar]
- Tazaki, K.; Okuno, M.; Furumoto, M.; Watanabe, H. Biomineralization of pisoliths in hot springs. Mater. Sci. Eng. C 2006, 26, 617–623. [Google Scholar] [CrossRef]
- Tazaki, K.; Morii, I. Microbial immobilization of Si, Mn, Fe and Sr ions in the nacreous layer of Sinohyliopsis schlegeli and environmental factors. Earth Sci. Front. 2008, 15, 55–65. [Google Scholar] [CrossRef]
- Tazaki, K. Microbial Mats; Their Biomineralization and Bioremediation; Takakuwa Art Printing Office: Ishikawa, Japan, 2009; 228p. [Google Scholar]
- Lowenstam, H.A.; Weiner, S. On Biomineralization; Oxford University Press: Oxford, UK, 1989; pp. 45–46. [Google Scholar]
- Mann, H.; Tazaki, K.; Fyfe, W.S.; Kerrich, R. Microbial Accumulation of Iron and Manganese in Different Aquatic Environments: An Electron Optical Study; CATENA SUPPLEMENT 21; CATENA: Cremlingen, Germany, 1992; pp. 115–131. [Google Scholar]
- Tazaki, K.; Takehara, T.; Ishigaki, Y.; Nakagawa, H. The distribution of Sr, Cs, Np and Pu in the radioactive paddy rice field in Fukushima. J. Foss. Res. 2013, 46, 35–57. [Google Scholar]
- Tazaki, K.; Simojima, Y.; Takehara, T.; Nakano, M. Formation of microbial mats and salt in radioactive paddy soils in Fukushima, Japan. Minerals 2015, 5, 849–862. [Google Scholar] [CrossRef]
- Seckbach, J.; Oren, A. Microbial Mats: Modern and Ancient Microorganisms in Stratified Systems; Springer: New York, NY, USA, 2010; p. 606. [Google Scholar]
- Morikawa, H.; Takahashi, M.; Kawamura, Y. New progress in environmental biotechnology using phytoremediation. J. Environ. Biotechnol. 2001, 1, 1–14. [Google Scholar]
- Ehrlich, H.L.; Newman, D.K.; Kappler, A. Ehrlich’s Geomicrobiology, 6th ed.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2016; 635p. [Google Scholar]
- Sekine, M.; Shimizu, F. Chemical Structure of Gel Material of Pectinatella magnifica. Abstract of Japanese Agricultural Chemical Society. 2013. Available online: http://www.jsbba.or.jp/MeetingofJSBBA/2013/MeetingofJSBBA2013.pdf (accessed on 24 September 2017).
- Oono, K.; Shibasaki, T. The lagoon and bay sediments of the Hokuriku district. Earth Sci. 1967, 21, 25–34. [Google Scholar]
- Tsukawaki, S.; Asano, I. Sedimentation in Kahoku-gata Lagoon in Ishikawa Pref. Central Japan; Bulletin of the Japan Sea Research Institute, Kanazawa University: Kanazawa, Japan, 1999; Volume 30, pp. 69–97. [Google Scholar]
- Tazaki, K. Architecture of biomats reveals history of geo-, aqua-, and bio-systems. Episodes 1999, 22, 21–25. [Google Scholar]
- Tazaki, K.; Tazaki, F.; Okuno, M.; Takehara, T.; Ishigaki, Y.; Nakagawa, H. Formation of pisolite at hot springs in Saturnia, Toscana, Italy. J. Geol. Soc. Jpn. 2016, 122, 45–60. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Kahokugata Lake | Makiyama Agriculture Reservoirs Pond | |
|---|---|---|
| Date | 27 June, 14 July in 2016 | 18 June, 1 July in 2016 |
| pH | 6.8–7.7–9.0 | 6.8–7.5 |
| Water Temp. | 26–27 °C | 33–36 °C |
| EC | 12 μS/cm | 12 μS/cm |
| DO | 6.8–7.7 mg/L | 3.0–4.0 mg/L |
| Water Color | light brown | light brown |
| Radio Activity | 110–130 cpm (BG 80–100cpm) | 110–120 cpm (BG 80–100 cpm) |
| Size of | 30–40–60 cm | 20–50 cm |
| Pectinatella magnifica | 200 g, 800 g, 14 kg | 500 g, 1 kg, 5 kg |
| Atomic Number | Elements | mg/L |
|---|---|---|
| 5 | B | 0.046 |
| 11 | Na | 13.100 |
| 12 | Mg | 46.500 |
| 13 | Al | 204.000 |
| 14 | Si | - |
| 15 | P | 26.200 |
| 16 | Si | 53.700 |
| 17 | Cl | 1.400 |
| 19 | K | 22.400 |
| 20 | Ca | 45.600 |
| 22 | Ti | 5.870 |
| 23 | V | 0.630 |
| 24 | Cr | 0.320 |
| 25 | Mn | 3.980 |
| 26 | Fe | 290.000 |
| 27 | Co | 0.100 |
| 28 | Ni | 0.170 |
| 29 | Cu | 0.150 |
| 30 | Zn | 1.580 |
| 33 | As | 0.100 |
| 37 | Rb | 0.048 |
| 38 | Sr | 0.110 |
| 39 | Y | 0.084 |
| 40 | Zr | ND |
| 48 | Cd | 0.004 |
| 55 | Ce | 0.540 |
| 56 | Ba | 0.470 |
| 80 | Hg | ND |
| 82 | Pb | 0.150 |
| Elements | 1 | 2 | 3 | 4 |
|---|---|---|---|---|
| Mg | 1.60 | 1.22 | 2.09 | 3.45 |
| Al | 10.49 | 7.47 | 12.67 | 15.34 |
| Si | 46.05 | 56.48 | 48.67 | 47.03 |
| P | 3.52 | 3.18 | 3.54 | 3.39 |
| S | 7.98 | 7.92 | 7.06 | 3.16 |
| Cl | 0.43 | 0.31 | 0.68 | 0.18 |
| K | 1.14 | 0.26 | 1.24 | 1.31 |
| Ca | 1.70 | 1.51 | 2.04 | 1.30 |
| Ti | 0.24 | ND | ND | ND |
| Mn | 0.04 | ND | ND | ND |
| Fe | 6.50 | 4.86 | 4.73 | 10.90 |
| Cu | 13.69 | 11.09 | 13.71 | 6.55 |
| Total | 93.38 | 94.30 | 96.43 | 92.61 |
| Makiyama Pond: Carbon Coated Statobolasts (mass %, ±3δ) | |||
|---|---|---|---|
| Elements | 1 | 2 | 3 |
| Na | 4.70 | 6.84 | 6.64 |
| Mg | 5.68 | 5.34 | 4.32 |
| Al | 2.26 | 1.97 | 1.20 |
| Si | 9.43 | 5.72 | 10.09 |
| P | 3.05 | 0.53 | 0.47 |
| S | 45.83 | 37.01 | 53.75 |
| Cl | 3.81 | 2.33 | 3.51 |
| K | 3.55 | ND | 3.02 |
| Ca | 14.78 | 12.78 | 12.43 |
| Ti | 0.42 | ND | 0.59 |
| Mn | ND | ND | ND |
| Fe | 4.01 | ND | 1.89 |
| Ni | 0.63 | 0.59 | ND |
| Cu | ND | 15.95 | ND |
| Zn | ND | 8.22 | ND |
| Sr | ND | 0.59 | 0.64 |
| Total | 98.15 | 97.87 | 98.55 |
| Statoblasts | Statoblasts | Anchor shape thorn | |
| (a) Statoblasts | ||
| 2θ (°) | d-Value (Å) | Relative Intensity |
| 20.050 | 4.428 | 10 |
| 21.400 | 4.149 | 25 |
| 21.560 | 4.118 | 25 |
| 22.571 | 3.936 | 20 |
| 22.737 | 3.908 | 20 |
| 24.386 | 3.650 | 10 |
| 27.200 | 3.276 | 100 |
| 27.340 | 3.259 | 85 |
| 27.480 | 3.243 | 50 |
| 28.376 | 3.143 | 20 |
| 30.877 | 2.896 | 10 |
| 35.354 | 2.537 | 15 |
| 36.296 | 2.473 | 15 |
| 39.960 | 2.254 | 20 |
| 40.160 | 2.244 | 20 |
| 43.158 | 2.096 | 15 |
| 50.640 | 1.801 | 20 |
| 50.820 | 1.795 | 20 |
| (b) Agar Part (Gelatinous Part) | ||
| 2θ (°) | d-Value (Å) | Relative Intensity |
| 8.056 | 10.974 | - * |
| 12.237 | 7.233 | - ** |
| 19.730 | 4.488 | - * |
| 35.939 | 2.497 | 40 *** |
| 38.644 | 2.328 | 19 *** |
| 42.881 | 2.107 | 100 *** |
| 54.007 | 1.697 | 25 *** |
| Components | 1 | 2 | 3 | 4 | 5 |
|---|---|---|---|---|---|
| SiO2 | 60.3000 | 58.1000 | 55.8000 | 56.8000 | 57.9000 |
| Al2O3 | 20.1000 | 19.5000 | 23.1000 | 14.9000 | 15.8000 |
| CO2 | 6.5500 | 9.1400 | 9.3100 | 15.6000 | 13.0000 |
| Fe2O3 | 6.3000 | 4.5600 | 5.2000 | 4.6500 | 4.9900 |
| K2O | 2.4100 | 2.2500 | 2.1700 | 2.3800 | 2.4100 |
| MgO | 1.9800 | 1.2200 | 1.1400 | 1.7500 | 1.8100 |
| TiO2 | 0.8050 | 0.4590 | 0.4440 | 0.5730 | 0.6170 |
| Na2O | 0.7310 | 1.6600 | 1.0500 | 1.2500 | 1.2500 |
| CaO | 0.4980 | 2.3200 | 1.1100 | 1.2500 | 1.1900 |
| P2O5 | 0.0947 | 0.4030 | 0.3540 | 0.4040 | 0.4620 |
| SO3 | 0.0603 | 0.1540 | 0.1360 | 0.1850 | 0.1900 |
| MnO | 0.5290 | 0.0434 | 0.0588 | 0.1070 | 0.1060 |
| Total | 100.3590 | 99.8100 | 99.9280 | 99.8590 | 99.7250 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tazaki, K.; Fukuyama, A.; Tazaki, F.; Okuno, M.; Hashida, Y.; Hashida, S.; Takehara, T.; Nakamura, K.; Kato, T. Mineralogical and Elemental Composition of Pectinatella magnifica and Its Statoblasts. Minerals 2018, 8, 242. https://doi.org/10.3390/min8060242
Tazaki K, Fukuyama A, Tazaki F, Okuno M, Hashida Y, Hashida S, Takehara T, Nakamura K, Kato T. Mineralogical and Elemental Composition of Pectinatella magnifica and Its Statoblasts. Minerals. 2018; 8(6):242. https://doi.org/10.3390/min8060242
Chicago/Turabian StyleTazaki, Kazue, Atsuko Fukuyama, Fumie Tazaki, Masayuki Okuno, Yumiko Hashida, Shozo Hashida, Teruaki Takehara, Keiichi Nakamura, and Tomohiro Kato. 2018. "Mineralogical and Elemental Composition of Pectinatella magnifica and Its Statoblasts" Minerals 8, no. 6: 242. https://doi.org/10.3390/min8060242