Río Tinto: A Geochemical and Mineralogical Terrestrial Analogue of Mars

{kind=link}
{kind=link}
{kind=link}
{kind=link}
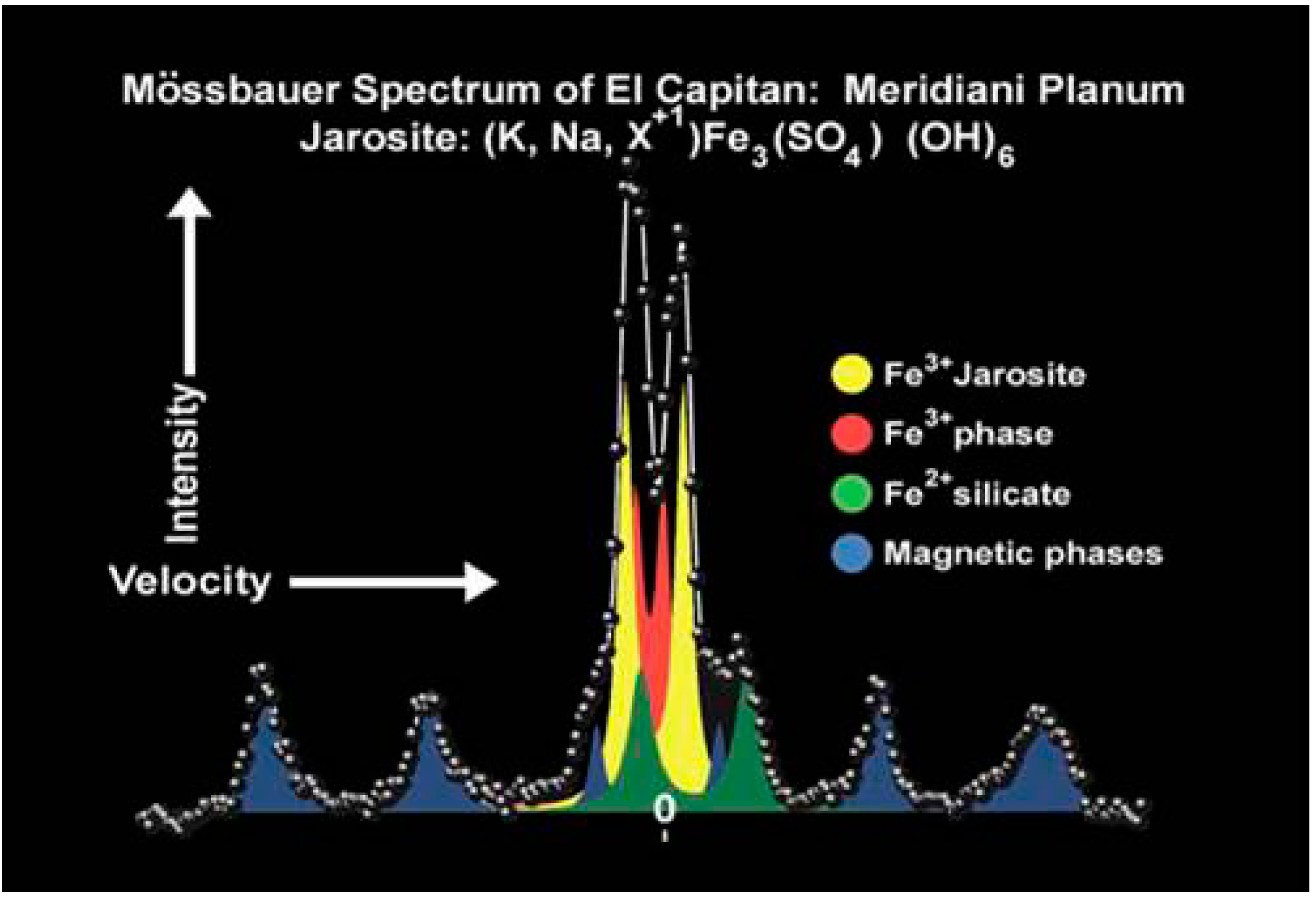{kind=link}
{kind=link}
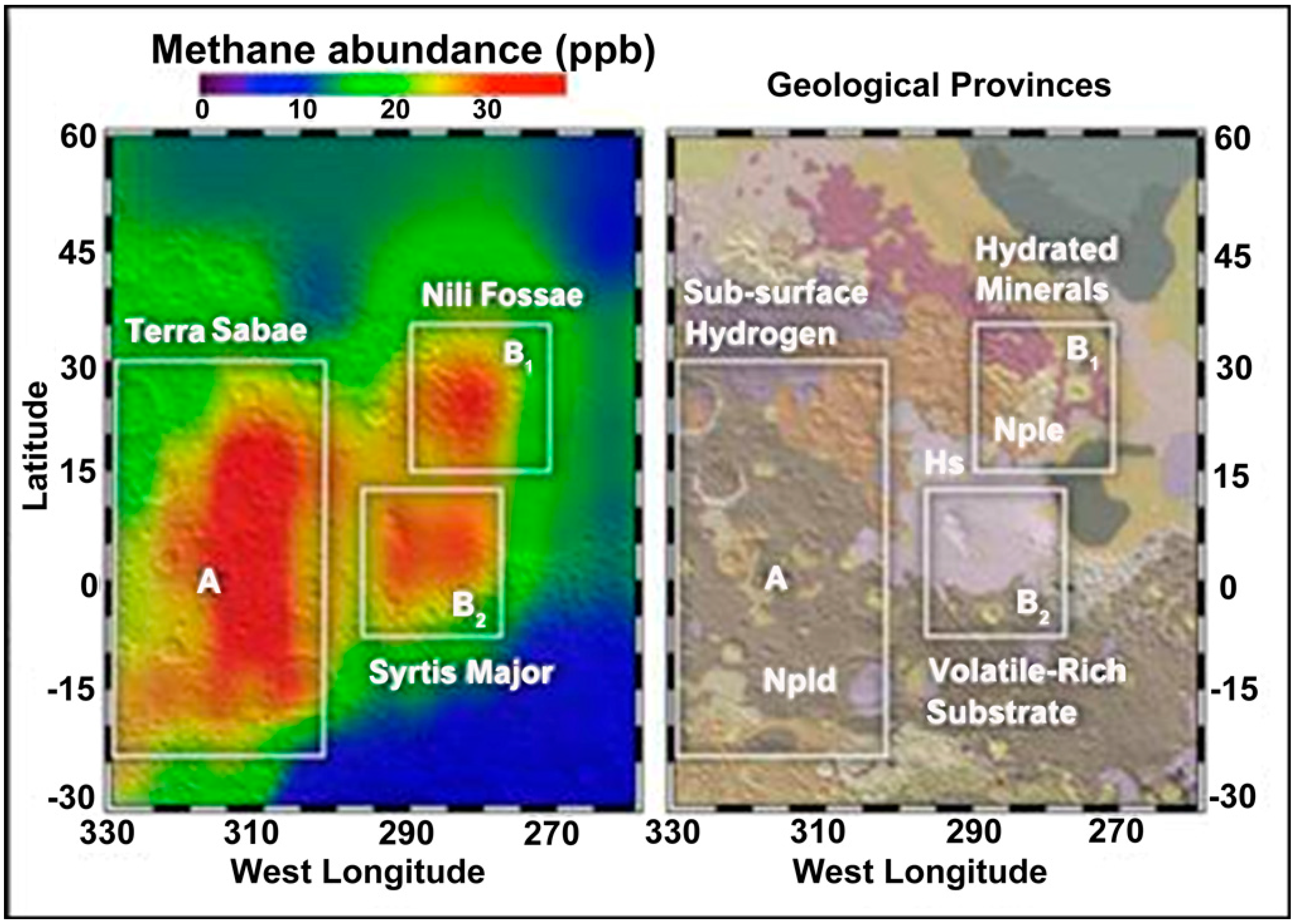{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Rio Tinto

3. Iron Bioformations in the Tinto Basin

4. Iron World
5. Subsurface Geomicrobiology of the Iberian Pyrite Belt


6. Rio Tinto as a Mars Terrestrial Analog




7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Des Marais, D.J.; Nuth, J.A.; Allamandola, L.J.; Boss, A.P.; Farmer, J.D.; Hoehler, T.M.; Jakosky, B.M.; Meadows, V.S.; Pohorille, A.; Runnegar, B.; et al. The NASA Astrobiology Roadmap. Astrobiology 2008, 8, 715–730. [Google Scholar]
- Margulis, L.; Mazur, P.; Barghoorn, E.S.; Halvorson, H.O.; Jukes, T.H.; Kaplan, I.R. The Viking Mission: Implications for life in the Vallis Marineris area. J. Mol. Evol. 1979, 14, 223–232. [Google Scholar] [CrossRef]
- Johnson, D.B.; Hallberg, K.B. The microbiology of acidic mine waters. Res. Microbiol. 2003, 154, 466–473. [Google Scholar] [CrossRef]
- Ehrlich, H.L.; Newman, D.K. Geomicrobiology, 5th ed.; CRC Press: Boca Raton, FL, USA, 2008. [Google Scholar]
- Sand, W.; Gehrke, T.; Hallma, R.; Schippers, A. Sulfur chemistry, biofilm and the (in)direct attack mechanisms—A critical evaluation of bacterial leaching. Appl. Microbiol. Biotechnol 1995, 43, 961–966. [Google Scholar] [CrossRef]
- Sand, W.; Gehrke, T.; Jozsa, P.G.; Schippers, A. Biochemistry of bacterial leaching—Direct vs. indirect bioleaching. Hydrometall 2001, 59, 159–175. [Google Scholar] [CrossRef]
- Colmer, A.R.; Temple, K.L.; Hinkle, H.E. An iron-oxidizing bacterium from the acid drainage of some bituminous coal mines. J. Bacteriol. 1950, 59, 317–328. [Google Scholar]
- Pronk, J.T.; Bruyn, J.C.; Bos, P.; Kuenen, J.G. Anaerobic growth of Thiobacillus ferrooxidans. Appl. Environ. Microbiol. 1992, 58, 2227–2230. [Google Scholar]
- Amils, R.; González-Toril, E.; Gómez, F.; Fernández-Remolar, D.; Rodríguez, N.; Malki, M.; Zuluaga, J.; Aguilera, A.; Amaral-Zettler, L.A. Importance of chemolithotrophy for early life on Earth: The Tinto River (Iberian Pyritic Belt) case. In Origins; Seckbach, J., Ed.; Springer: Amsterdam, The Netherlands, 2004; pp. 463–480. [Google Scholar]
- Rawlings, D.E. Heavy metal mining using microbes. Annu. Rev. Microbiol. 2002, 56, 65–91. [Google Scholar] [CrossRef]
- Benz, M.; Brune, A.; Schink, B. Anaerobic and aerobic oxidation of ferrous iron at neutral pH by chemoheterotrophic nitrate-reducing bacteria. Arch. Microbiol. 1998, 169, 159–165. [Google Scholar] [CrossRef]
- Widdel, F.; Schnell, S.; Heising, S.; Ehrenreich, A.; Assmus, B.; Schink, B. Ferrous iron oxidation by anoxygenic phototrophic bacteria. Nature 1993, 162, 834–836. [Google Scholar] [CrossRef]
- Godd, T. The deep hot biosphere. Proc. Natl. Acad. Sci. USA 1992, 89, 6045–6049. [Google Scholar] [CrossRef]
- Bachofen, R.; Ferloni, P.; Flynn, L. Microorganisms in the subsurface. Microbiol. Res. 1998, 153, 1–22. [Google Scholar] [CrossRef]
- Pedersen, K. Exploration of deep intraterrestrial microbial life: Current perspectives. FEMS Microbiol. Lett. 2000, 185, 9–16. [Google Scholar] [CrossRef]
- Chapelle, F.H.; O’Nelly, K.; Bradley, P.M.; Methé, B.A.; Ciufo, S.A.; Knobel, L.L.; Lovley, D.R. A hydrogen-based subsurface microbial community dominated by methanogens. Nature 2002, 415, 312–315. [Google Scholar] [CrossRef]
- Leblanc, M.; Morales, J.A.; Borrego, J.; Elbaz-Poulichet, F. 4500-year-old mining pollution in Southwestern Spain: Long-Term implications for modern mining pollution. Econ. Geol. 2000, 95, 655–662. [Google Scholar]
- López-Archilla, A.I.; Marín, I.; Amils, R. Microbial community composition and ecology of an acidic aquatic environment: The Tinto River, Spain. Microbiol. Ecol. 2001, 41, 20–35. [Google Scholar]
- Amaral-Zettler, L.A.; Gómez, F.; Zettler, E.; Keenan, B.G.; Amils, R.; Sogin, M.L. Microbiology: Eukaryotic diversity in Spain’s River of Fire. Nature 2002, 417, 137. [Google Scholar] [CrossRef]
- González-Toril, E.; LLobet-Brosa, E.; Casamayor, E.O.; Amann, R.; Amils, R. Microbial ecology of an extreme acidic environment, the Tinto River. Appl. Environ. Microbiol. 2003, 69, 4853–4865. [Google Scholar]
- Boulter, C.A. Did both extensional tectonics and magmas act as major drivers of convection cells during the formation of the Iberian Pyrite Belt massive sulphide deposits? J. Geol. Soc. London 1996, 153, 181–184. [Google Scholar] [CrossRef]
- Leistel, J.M.; Marcoux, E.; Theiblemont, D.; Quesada, C.; Sánchez, A.; Almodóvar, G.R.; Pascual, E.; Saez, R. The volcanic-hosted massive sulphide deposits of the Iberian Pyrite Belt. Miner. Depos. 1997, 33, 2–30. [Google Scholar] [CrossRef]
- Lescuyer, J.L.; Leistel, J.M.; Mrcoux, E.; Milési, J.P.; Thiéblemont, D. Late Devonian-Early Carboniferous peak sulphide mineralization in the Western Hercynides. Miner. Depos. 1997, 33, 208–220. [Google Scholar] [CrossRef]
- González-Toril, E.; Aguilera, A.; Rodríguez, N.; Fernández-Remolar, D.; Gómez, F.; Díaz, E.; García-Moyano, A.; Sanz, J.L.; Amils, R. Microbial ecology of Río Tinto, a natural extreme acidic environment of biohydrometallurgical interest. Hydrometall 2010, 104, 329–333. [Google Scholar]
- García-Moyano, A.; González-Toril, E.; Aguilera, A.; Amils, R. Prokaryotic community composition and ecology of macroscopic floating filaments from an extreme acidic environment, Río Tinto, (SW, Spain). Syst. Appl. Microbiol. 2007, 30, 601–614. [Google Scholar]
- García-Moyano, A.; González-Toril, E.; Aguilera, A.; Amils, R. Comparative microbial ecology study of the sediments and the water column of the Río Tinto, an extreme acidic environment. FEMS Microbiol. Ecol. 2012, 81, 303–314. [Google Scholar]
- Ohmura, N.; Sasaki, K.; Matsumoto, N.; Saiki, H. Anaerobic respiration using Fe3+, S0 and H2 in the chemolithoautotrophic bacterium Acidithiobacillus ferrooxidans. J. Bacteriol. 2002, 18, 2081–2087. [Google Scholar] [CrossRef]
- Malki, M.; González-Toril, E.; Sanz, J.L.; Gómez, F.; Rodríguez, N.; Amils, R. Importance of the iron cycle in biohydrometallurgy. Hydrometall 2006, 83, 223–228. [Google Scholar] [CrossRef]
- Coupland, K.; Johnson, D.B. Evidence that the potential for dissimilatory ferric iron reduction is widespread among acidophilic heterotrophic bacteria. FEMS Microbiol. Lett. 2008, 279, 30–35. [Google Scholar] [CrossRef]
- Malki, M.; de Lacey, A.L.; Rodríguez, N.; Amils, R.; Fernández, V.M. Preferential use of an anode as an electron acceptor by an acidophilic bacterium in the presence of oxygen. Appl. Environ. Microbiol. 2008, 74, 4472–4476. [Google Scholar] [CrossRef]
- Sánchez-Andrea, I.; Rodríguez, N.; Amils, R.; Sanz, J.L. Microbial diversity in anaerobic sediments at Río Tinto, a naturally acidic environment with high heavy metal content. Appl. Environ. Microbiol. 2011, 77, 6085–6093. [Google Scholar] [CrossRef]
- Sánchez-Andrea, I.; Rojas-Ojeda, P.; Amils, R.; Sanz, J.L. Screening of anaerobic activities in sediments of an acidic environment: Tinto River. Extremophiles 2012, 16, 829–839. [Google Scholar]
- Sánchez-Andrea, I.; Stams, A.J.M.; Amils, R.; Sanz, J.L. Enrichment and isolation of acidophilic sulfate-reducing bacteria from Tinto River sediments. Environ. Microbiol. Rep. 2013, 5, 672–678. [Google Scholar] [CrossRef]
- Lu, S.; Gischkat, S.; Reiche, M.; Akob, D.M.; Hallberg, K.B.; Küsel, K. Ecophysiology of Fe-cycling bacteria in acidic sediments. Appl. Environ. Microbiol. 2010, 76, 8174–8183. [Google Scholar] [CrossRef]
- Hallberg, K.B.; Johnson, D.B. Biodiversity of acidophilic prokaryotes. Adv. Appl. Microbiol. 2001, 49, 37–84. [Google Scholar]
- González-Toril, E.; Gómez, F.; Malki, M.; Amils, R. The Isolation and Study of Acidophilic Microorganisms. In Extremophiles, Methods in Microbiology; Rainey, F.A., Oren, A., Eds.; Elsevier Academic Press: San Diego, CA, USA, 2006; Volume 35, pp. 471–510. [Google Scholar]
- Rawlings, D.E. Characteristics and adaptability of iron- and sulfur-oxidizing microorganisms used for the recovery of metals from minerals and their concentrates. Microb. Cell Factor. 2005, 4, 13–28. [Google Scholar] [CrossRef] [Green Version]
- Aguilera, A.; Manrubia, S.C.; Gómez, F.; Rodríguez, N.; Amils, R. Eukaryotic community distribution and its relationship to water physicochemical parameters in an extreme acidic environment, Río Tinto (Southwestern Spain). Appl. Environ. Microbiol. 2006, 72, 5325–5330. [Google Scholar] [CrossRef]
- Aguilera, A.; Zettler, E.; Gómez, F.; Amaral-Zettler, L.; Rodríguez, N.; Amils, R. Distribution and seasonal variability in the benthic eukaryotic community of Río Tinto (SW, Spain), and acidic, high metal extreme environment. Syst. Appl. Microbiol. 2007, 30, 531–546. [Google Scholar] [CrossRef]
- Amaral-Zettler, L.; Zettler, E.R.; Theroux, S.M.; Palacios, C.; Aguilera, A.; Amils, R. Microbial community structure across the tree of life in the extreme Río Tinto. ISME J. 2010, 5, 42–50. [Google Scholar] [CrossRef]
- López-Archilla, A.I.; González, A.E.; Terrón, M.C.; Amils, R. Diversity and ecological relationships of the fungal populations of an acidic river of Southwestern Spain: The Tinto River. Can. J. Microbiol. 2005, 50, 923–934. [Google Scholar]
- Rodríguez, N.; Menéndez, N.; Tornero, J.; Amils, R.; de la Fuente, V. Internal iron biomineralization in Imperata cylindrica, a perennial grass: Chemical composition, speciation and plant localization. New Phytol. 2005, 165, 781–789. [Google Scholar]
- De la Fuente, V.; Rufo, L.; Rodríguez, N.; Amils, R.; Zuluaga, J. Metal accumulation screening of the Río Tinto flora (Huelva, Spain). Biol. Trace Elem. Res. 2010, 134, 318–341. [Google Scholar]
- Franco, A.; Rufo, L.; Rodríguez, N.; Amils, R.; de la Fuente, V. Iron absorption, localization and biomineralization of Cynodon. dactylon, a perennial grass from the Río Tinto basin (SW Iberian Peninsula). J. Plant Nutr. Soil Sci. 2013, 176, 836–842. [Google Scholar]
- Amils, R.; de la Fuente, V.; Rodríguez, N.; Zuluaga, J.; Menéndez, N.; Tornero, J. Composition, speciation and distribution of iron minerals in Imperata cylindrica. Plant Physiol. Biochem. 2007, 45, 335–340. [Google Scholar] [CrossRef]
- De la Fuente, V.; Rodríguez, N.; Amils, R. Immunocytochemical analysis of the subcellular distribution of ferritin in Imperata cylindrica (L.) Raeuschel, an iron hyperaccumulator plant. Acta Histochim. 2012, 114, 232–236. [Google Scholar]
- Fernández-Remolar, D.C.; Rodríguez, N.; Gómez, F.; Amils, R. Geological record of an acidic environment driven by iron hydrochemistry: The Tinto River system. J. Geophys. Res. 2003, 108. [Google Scholar] [CrossRef]
- Fernández-Remolar, D.C.; Morris, R.V.; Gruener, J.E.; Amils, R.; Knoll, A.H. The Rio Tinto Basin, Spain: Mineralogy, sedimentary geobiology and implications for interpretation of ourcrop rocks of meridiani Planum, Mars. Earth Planet Sci. Lett. 2005, 240, 149–167. [Google Scholar]
- Fernández-Remolar, D.C.; Knoll, A.H. Fossilization potential of iron-bearing minerals in acidic environments of Rio Tinto, Spain: Implications for Mars exploration. Icarus 2008, 194, 72–85. [Google Scholar]
- Amils, R.; González-Toril, E.; Fernández-Remolar, D.; Gómez, F.; Aguilera, A.; Rodríguez, N.; Malki, M.; García-Moyano, A.; González-Fairén, A.; de la Fuente, V.; et al. Extreme environments as Mars terrestrial analogs: The Río Tinto case. Planet Space Sci. 2007, 55, 370–381. [Google Scholar] [CrossRef]
- Oggerin, M.; Tornos, F.; Rodríguez, N.; del Moral, C.; Sánchez-Román, M.; Amils, R. Specific jarosite biomineralization by Purpureocillium. lilacinum, an acidophilic fungi isolated from Río Tinto. Environ. Microbiol. 2013, 15, 2228–2237. [Google Scholar]
- Fernández-Remolar, D.C.; Preston, L.J.; Sánchez-Román, M.; Izawa, M.R.M.; Huang, L.; Southam, G.; Banerjee, N.R.; Osinski, G.R.; Flemming, R.; Gómez-Ortíz, D.; et al. Carbonate precipitation under bulk acidic conditions as a potential biosignature for searching life on Mars. Earth Planet Sci. Lett. 2012, 351, 13–26. [Google Scholar]
- Sánchez-Román, M.; Fernández-Remolar, D.; Amils, R.; Sánchez-Navas, A.; Schmid, T.; Martín-Uriz, P.S.; Rodríguez, N.; McKenzie, J.A. Microbial mediated formation of Fe-carbonate minerals under extreme acidic conditions. Sci. Rep. 2014, 4. [Google Scholar] [CrossRef]
- Colín-García, M.; Kanawati, B.; Harir, M.; Schmidt-Kopplin, P.; Amils, R.; Parro, V.; García, M.; Fernández-Remolar, D. Detection of peptidic sequences in the ancient acidic sediments of Río Tinto, Spain. Orig. Life Evol. Biosph. 2011, 41, 523–527. [Google Scholar]
- Preston, L.; Shuster, J.; Fernández-Remolar, D.; Banerjee, N.; Osinski, G.R.; Southam, G. The preservation and degradation of filamentous bacteria and biomolecules within iron oxide deposits at Rio Tinto, Spain. Geobiology 2011, 9, 233–249. [Google Scholar] [CrossRef]
- Geen, A.; van Adkins, J.F.; Boyle, E.A.; Nelson, C.H.; Palanques, A. A 120-yr record of widespread contamination from mining of the Iberian Pyrite Belt. Geology 1997, 25, 291–294. [Google Scholar] [CrossRef]
- Gómez-Ortiz, D.; Fernández-Remolar, D.; Granda, A.; Quesada, C.; Granda, T.; Prieto-Ballesteros, O.; Molina, A.; Amils, R. Identification of the subsurface sulfide bodies responsible for acidity in Río Tinto source water, Spain. Earth Plant. Sci. Lett. 2014, 391, 36–41. [Google Scholar]
- Fernández-Remolar, D.; Gómez, F.; Prieto-Ballesteros, O.; Schelble, R.T.; Rodríguez, N.; Amils, R. Some ecological mechanisms to generate habitability in planetary subsurface areas by chemolithotrophic communities: The Río Tinto subsurface ecosystem as a model system. Astrobiology 2008, 8, 157–173. [Google Scholar]
- Moreno, C.; Capitán, M.A.; Doyle, M.; Nieto, J.M.; Ruiz, F.; Sáez, R. Edad mínima del gossan de Las Cruces: Implicaciones sobre la edad del inicio de los ecosistemas extremos en la Faja Pirítica Ibérica. Geogaceta 2003, 33, 67–70. (In Spainish) [Google Scholar]
- Gómez, F.; Aguilera, A.; Amils, R. Soluble ferric iron as an effective protective agent against UV radiation: Implications for early life. Icarus 2007, 191, 352–359. [Google Scholar]
- Gómez, F.; Mateo-Martí, E.; Prieto-Ballesteros, O.; Martín-Gago, J.; Amils, R. Protection of chemolithotrophic bacteria exposed to Mars environmental conditions. Icarus 2010, 209, 482–487. [Google Scholar] [CrossRef]
- Archibald, F. Lactobacillus plantarum, an organism not requiring iron. FEMS Microbiol. Lett. 1983, 19, 29–32. [Google Scholar] [CrossRef]
- Martin, J.H. Glacial-interglacial CO2 change: The iron hypothesis. Paleooceanography 1990, 5, 1–13. [Google Scholar] [CrossRef]
- Boyd, W.P.; Jickells, T.; Law, C.S.; Blain, S.; Boyle, E.A.; Buesseler, K.O.; Coale, K.H.; Cullen, J.J.; de Baar, H.J.W.; Follows, M.; et al. Mesoscale iron enrichment experiments 1993–2005: Synthesis and future directions. Science 2007, 315, 612–617. [Google Scholar] [CrossRef]
- Reid, R.T.; Live, D.H.; Faulkner, D.J.; Buttler, A. A siderophore from a marine bacterium with an exceptional ferric iron affinity constant. Nature 1993, 366, 455–458. [Google Scholar] [CrossRef]
- Braun, V.; Killmann, H. Bacterial solution to the iron supply problems. Trends Biochem. Sci. 1999, 24, 104–109. [Google Scholar] [CrossRef]
- Gómez, F.; Fernández-Remolar, D.; González-Toril, E.; Amils, R. The Tinto River, an Extreme Gaian Environment. In Scientists Debate Gaia 2000; Margulis, L., Miller, J., Boston, P., Schneider, S., Crist, C., Eds.; MIT Press: Boston, MA, USA, 2003; pp. 321–333. [Google Scholar]
- Anbar, A.D.; Knoll, A.H. Proterozoic ocean chemistry and evolution: A bioinorganic bridge. Science 2002, 297, 1137–1142. [Google Scholar] [CrossRef]
- Fernández-Remolar, D.; Prieto-Ballesteros, O.; Rodríguez, N.; Gómez, F.; Amils, R.; Gomez-Elvira, J.; Stoker, C. Underground habitats found in the Río Tinto Basin: A model for subsurface life habitats on Mars. Astrobiology 2008, 8, 1023–1046. [Google Scholar]
- Puente-Sánchez, F.; Moreno-Paz, M.; Rivas, L.A.; Cruz-Gil, P.; García-Villadangos, M.; Gómez, M.J.; Postigo, M.; Garrido, P.; González-Toril, E.; Briones, C.; et al. Deep subsurface sulfate reduction and methanogenesis in the Iberian Pyrite Belt revealed through geochemistry and molecular biomarkers. Geobiology 2014, 12, 34–47. [Google Scholar]
- Squyres, S.W.; Crotzinger, J.P.; Arvidson, R.E.; Bell, J.F., III; Calvin, W.; Christensen, P.R.; Clark, B.C.; Crisp, J.A.; Farrand, W.H.; Herkenhoff, K.E.; et al. In situ evidence for an ancient aqueous environment in Meridiani Planum, Mars. Science 2004, 306, 1709–1714. [Google Scholar]
- Klingelhöfer, G.; Morris, R.V.; Bernhardt, B.; Schröder, C.S.; de Souza, P.A., Jr.; Yen, A.; Gellert, R.; Evlanov, E.N.; Zubkov, B.; Foh, J.; et al. Jarosite and hematite at Meridiani Planum from the Mössbauer spectrometer on the Opportunity rover. Science 2005, 306, 1740–1745. [Google Scholar]
- Milliken, R.E.; Swayze, G.A.; Arvidson, R.E.; Bishop, J.L.; Clark, R.N.; Ehlmann, B.L.; Green, R.O.; Grotzinger, J.P.; Morris, R.V.; Murchie, S.L.; et al. Opaline silica in young deposits on Mars. Geology 2008, 36, 847–850. [Google Scholar] [CrossRef]
- Farrand, W.H.; Glotch, T.D.; Rice, J.W.; Hurowitz, J.A.; Swayze, G. Discovery of jarosite within Mawrth Vallis region of Mars: Implications for the geological history of the region. Icarus 2009, 204, 478–488. [Google Scholar] [CrossRef]
- Ehdmann, B.L.; Mustard, J.F. An in-situ record of major environmental transitions on early Mars at Northeast Syrtis Major. Geophys. Res. Lett. 2012, 39. [Google Scholar] [CrossRef]
- Fairen, A.G.; Fernández-Remolar, D.; Dohm, J.M.; Baker, V.R.; Amils, R. Inhibition of carbonate synthesis in acidic oceans from Mars. Nature 2004, 431, 423–426. [Google Scholar] [CrossRef]
- Zolotov, M.; Shock, E. Formation of jarosite-bearing deposits through aqueous oxidation of pyrite at the Meridiani Planum, Mars. Geophys. Res. Lett. 2005, 32. [Google Scholar] [CrossRef]
- Christensen, P.R.; Bandfield, J.L.; Clark, R.N.; Edgett, K.S.; Hamilton, V.E.; Hoefen, T.; Kieffer, H.H.; Kuzmin, R.O.; Lane, M.D.; Malin, M.C.; et al. Detection of crystalline hematite mineralization on Mars by the thermal emission spectrometer evidence for near-surface water. J. Geophys. Res. 2000, 104, 9623–9642. [Google Scholar] [CrossRef]
- Christensen, P.R.; Morris, R.V.; Lane, M.D.; Banfield, J.L.; Malin, M.C. Global mapping of martian hematite mineral deposits: remnants of water-driven processes on early Mars. J. Geophys. Res. 2001, 106, 23873–23885. [Google Scholar] [CrossRef]
- Christensen, P.R.; Ruff, S.W. Formation of the hematite-bearing unit in Meridiani Planum: Evidence for deposition in standing water. J. Geophys. Res. Planet. 2004, 109, E08003. [Google Scholar] [CrossRef]
- Rieder, R.; Gellert, R.; Anderson, R.C.; Brückner, J.; Clark, B.C.; Dreibus, G.; Economou, T.; Klingelhöfer, G.; Lugmair, G.W.; Ming, D.W.; et al. Chemistry of rocks and soils at Meridiani Planum from the alpha particle X-ray spectrometer. Science 2004, 306, 1746–1749. [Google Scholar] [CrossRef]
- McLennan, S.M.; Bell, J.F., III; Calvin, W.M.; Christensen, P.R.; Clark, B.C.; de Souza, P.A.; Farmer, J.; Farrand, W.H.; Fike, D.A.; Gellert, R.; et al. Provenance and diagenesis of the Burns formation, Meridiani Planum, Mars. Earth Planet. Sci. Lett. 2005, 240, 95–121. [Google Scholar]
- Bigham, J.M.; Schwertmann, U.; Traina, S.J.; Winland, R.L.; Wolf, M. Schwertmannite and the chemical modelling of iron in acid sulphate waters. Geochim. Cosmochim. Acta 1996, 60, 2111–2121. [Google Scholar] [CrossRef]
- Knoll, A.H.; Carr, M.; Clark, B.; Des Marais, D.J.; Farmer, J.D.; Fische, W.W.; Grotzinger, J.P.; McLennan, S.M.; Malin, M.; Schröder, C.; et al. An astrobiological perspective on Meridiani Planum. Earth Planet. Sci. Lett. 2005, 240, 179–189. [Google Scholar] [CrossRef]
- Grotzinger, J.P.; Arvidson, R.E.; Bell, J.F., III; Calvin, W.; Clark, B.C.; Fike, D.A.; Golombek, M.; Greeley, R.; Haldemann, A.; Herkenhoff, K.E.; et al. Stratigraphy, sedimentology and depositional environment of the Burns formation, Meridiani Planum, Mars. Earth Planet. Sci. Lett. 2005, 240, 11–72. [Google Scholar]
- Formisano, V.; Atreya, S.; Encrenaz, T.; Ignatiev, N.; Giuranna, M. Detection of methane in the atmosphere of Mars. Science 2004, 306, 1758–1761. [Google Scholar] [CrossRef]
- Mumma, M.J.; Villanueva, G.L.; Novak, R.E.; Hewagama, T.; Bonev, B.P.; DiSanti, M.A.; Mandell, A.; Smith, M.D. Strong release of methane on Mars in Northen Summer 2003. Science 2009, 323, 1041–1045. [Google Scholar] [CrossRef]
- Kotsyurbenko, O.R.; Friedrich, M.W.; Simankova, M.V.; Nozhenvnikova, A.N.; Golyshin, P.N.; Timmis, K.N.; Conrad, R. Shift from acetoclastic to H2 dependent methanogenesis in a West Siberian peat bog at low pH values and isolation of an acidophilic Methanobacterium strain. Appl. Environ. Microbiol. 2007, 73, 2344–2348. [Google Scholar] [CrossRef]
- Taconi, K.A.; Zappi, M.E.; French, W.T.; Brown, L.R. Methanogenesis under acidic pH conditions in a semi-continuous reactor system. Bioresour. Technol. 2008, 99, 8075–8081. [Google Scholar] [CrossRef]
- Sanz, J.L.; Rodríguez, N.; Díaz, E.; Amils, R. Methanogenesis in the sediments of Río Tinto, an extreme acidic environment. Environ. Microbiol. 2011, 13, 2336–2341. [Google Scholar] [CrossRef]
- Jakosky, B.M.; Haberle, R.M. The Seasonal Behavior of Water on Mars. In Mars; Kieffer, H.H., Ed.; University of Arizona Press: Tucson, AZ, USA, 1992; pp. 969–1016. [Google Scholar]
- Carr, M.H. Retention of an atmosphere on early Mars. J. Geophys. Res. 1999, 104, 21897–21909. [Google Scholar] [CrossRef]
- Boynton, W.V.; Feldman, W.C.; Squyres, S.W.; Prettyman, T.H.; Brückner, J.; Evans, L.G.; Reedy, R.C.; Starr, R.; Arnold, J.R.; Drake, D.M.; et al. Distribution of hydrogen in the near surface of Mars: Evidence for subsurface ice deposits. Science 2002, 297, 81–85. [Google Scholar]
- Bibring, J.P.; Langevin, Y.; Gendrin, A.; Gondet, B.; Poulet, F.; Berthé, M.; Soufflot, A.; Arvidson, R.; Mangold, N.; Mustard, J.; et al. Mars surface diversity as revealed by the OMEGA/Mars Express observations. Science 2005, 307, 1576–1581. [Google Scholar] [CrossRef]
- Smith, P.H.; Tamppari, L.K.; Arvidson, R.E.; Bass, D.; Blaney, D.; Boynton, W.V.; Carswell, A.; Catling, D.C.; Clark, B.C.; Duck, T.; et al. H2O at the Phoenix landing site. Science 2009, 325, 58–61. [Google Scholar]
- Parker, T.J.; Gorsline, D.S.; Saunders, R.S.; Pieri, D.C.; Schneeberger, D.M. Coastal geomorphology of the martian northern plains. J. Geophys. Res. 1993, 98, 11061–11078. [Google Scholar] [CrossRef]
- Malin, M.C.; Edgett, K.S. Sedimentary rocks of early Mars. Science 2000, 290, 1927–1937. [Google Scholar] [CrossRef]
- Malin, M.C.; Edgett, K.S. Evidence for persistent flow and aqueous sedimentation on early Mars. Science 2003, 302, 1931–1934. [Google Scholar] [CrossRef]
- Fairén, A.G.; Dohm, J.M.; Baker, V.R.; de Pablo, M.A.; Ruiz, J.; Ferris, J.; Anderson, R. Episodic flood inundations of the northern plains of Mars. Icarus 2003, 165, 53–67. [Google Scholar]
- Fairén, A.G. A cold and wet Mars. Icarus 2010, 208, 165–175. [Google Scholar]
- Bhattacharya, J.P.; Payenberg, T.H.D.; Lang, S.C.; Bourke, M. Dynamic river channels suggest a long-lived Noachian crater lake on Mars. Geophys. Res. Lett. 2005, 32. [Google Scholar] [CrossRef]
- Poulet, F.; Bibring, J.P.; Mustard, J.F.; Gendrin, A.; Mangold, N.; Langevin, Y.; Arvidson, R.E.; Gondet, B.; Gómez, C.; the Omega Team. Phyllosilicates on Mars and implications for early martian climate. Nature 2005, 438, 623–627. [Google Scholar]
- Heldmann, J.L.; Toon, O.B.; Pollard, W.H.; Mellon, M.T.; Pitlick, J.; McKay, C.P.; Andersen, D.T. Formation of Martian gullies by the action of liquid water flowing under current Martian environmental conditions. J. Geophys. Res. 2005, 110. [Google Scholar] [CrossRef]
- Baldridge, A.M.; Hook, S.J.; Crowley, J.K.; Marion, G.M.; Kargel, J.S.; Michalski, J.L.; Thomson, B.J.; de Souza Filho, C.R.; Bridges, N.T.; Brown, A.J.; et al. Contemporaneous deposition of phyllosilicates and sulfates: Using Australin acidic lake deposits to describe geochemical variability on Mars. Geophys. Res. Lett. 2009, 36. [Google Scholar] [CrossRef]
- Benison, K.O.; LaClair, D.A. Modern an ancient extremely acid saline deposits: Terrestrial analogs for martian environments. Astrobiology 2003, 3, 609–618. [Google Scholar] [CrossRef]
- Michel, F.A.; Everdingen, R.O. Formation of jarosite deposits on Cretaceous shales in the Fort Norman area, Northwest Territories. Can. Mineral. 1987, 25, 221–226. [Google Scholar]
- Lacelle, D.; Levillé, R. Acid drainage generation and associated Ca-Fe-SO4 minerals in a periglacial environment, Eagle Plains, Northen Yukon, Canada: A potential analogue for low-temperature sulfate formation on Mars. Planet. Space Sci. 2010, 5, 509–521. [Google Scholar] [CrossRef]
- Batller, M.W.; Osinki, G.R.; Lim, D.S.S.; Dávila, A.F.; Michel, F.A.; Craig, M.A.; Izawa, M.R.M.; Leoni, L.; Slater, G.F.; Fairén, A.G.; et al. Characterization of the acidic cold seep emplaced jarosite: Golden Deposit, NWT, Canada, as an analogue for jarosite deposition on Mars. Icarus 2012, 242, 382–398. [Google Scholar]
- West, L.; McGovern, D.J.; Onston, T.C.; Morris, R.V.; Suchecki, P.; Pratt, L.M. High Lake gossan deposit: An Artic analogue for ancient Martian surficial processes? Plant. Sapce. Sci. 2009, 57, 1302–1311. [Google Scholar] [CrossRef]
- Dold, B.; González-Toril, E.; Aguilera, A.; López-Pamo, E.; Cisternas, M.E.; Bucchi, F.; Amils, R. Acid Rock Drainage and Rock Weathering in Antarctica: important sources for iron cycling in the Southern Ocean. Environ. Sci. Technol. 2013, 47, 6129–6136. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Amils, R.; Fernández-Remolar, D.; The IPBSL Team. Río Tinto: A Geochemical and Mineralogical Terrestrial Analogue of Mars. Life 2014, 4, 511-534. https://doi.org/10.3390/life4030511
Amils R, Fernández-Remolar D, The IPBSL Team. Río Tinto: A Geochemical and Mineralogical Terrestrial Analogue of Mars. Life. 2014; 4(3):511-534. https://doi.org/10.3390/life4030511
Chicago/Turabian StyleAmils, Ricardo, David Fernández-Remolar, and The IPBSL Team. 2014. "Río Tinto: A Geochemical and Mineralogical Terrestrial Analogue of Mars" Life 4, no. 3: 511-534. https://doi.org/10.3390/life4030511