
Bipyrimidine Signatures as a Photoprotective Genome Strategy in G + C-rich Halophilic Archaea


Abstract
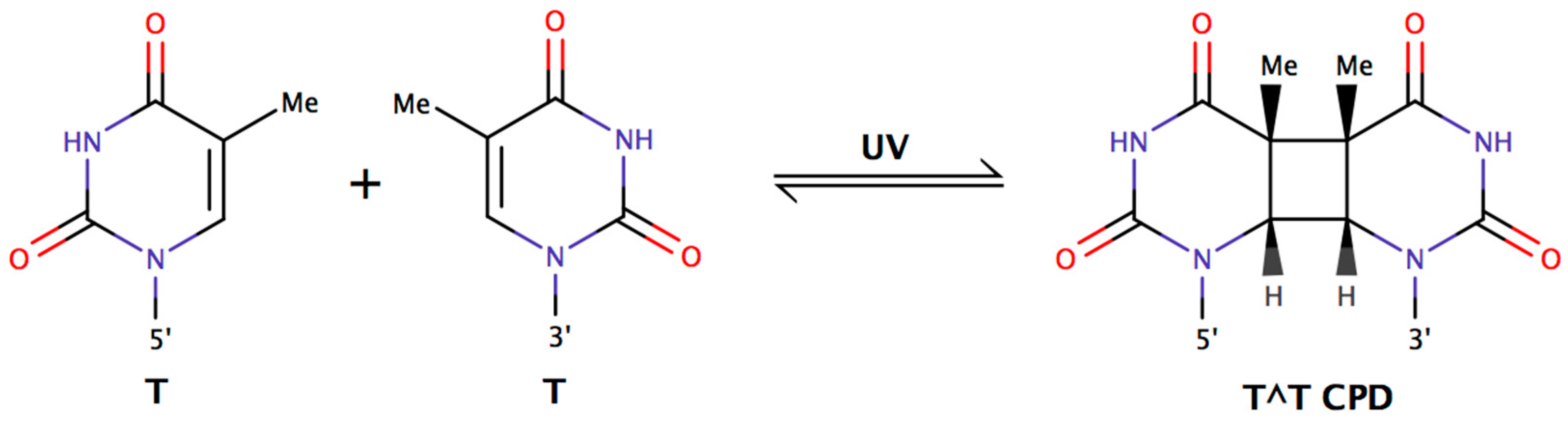
:1. Introduction
2. Materials and Methods
2.1. Comparing G + C Content of Halophilic Archaea Versus Other Prokaryotes
2.2. Genome Sampling
2.3. Determining Bipyrimidine Incidences
2.4. Determining Theoretical Genomic Photoreactivity (Pg)
2.5. Statistical Methods
3. Results
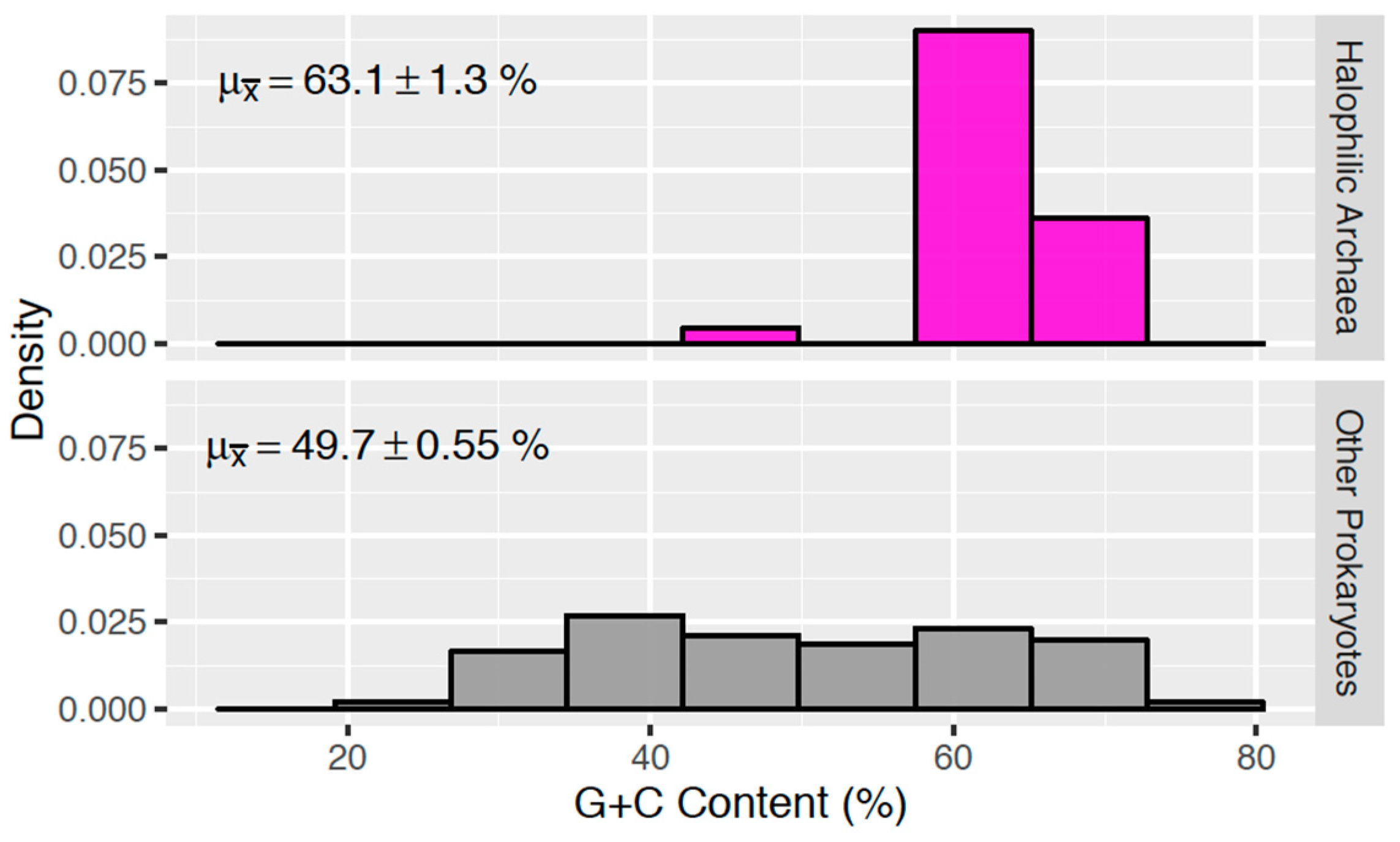
3.1. G + C Content of Halophilic Archaea
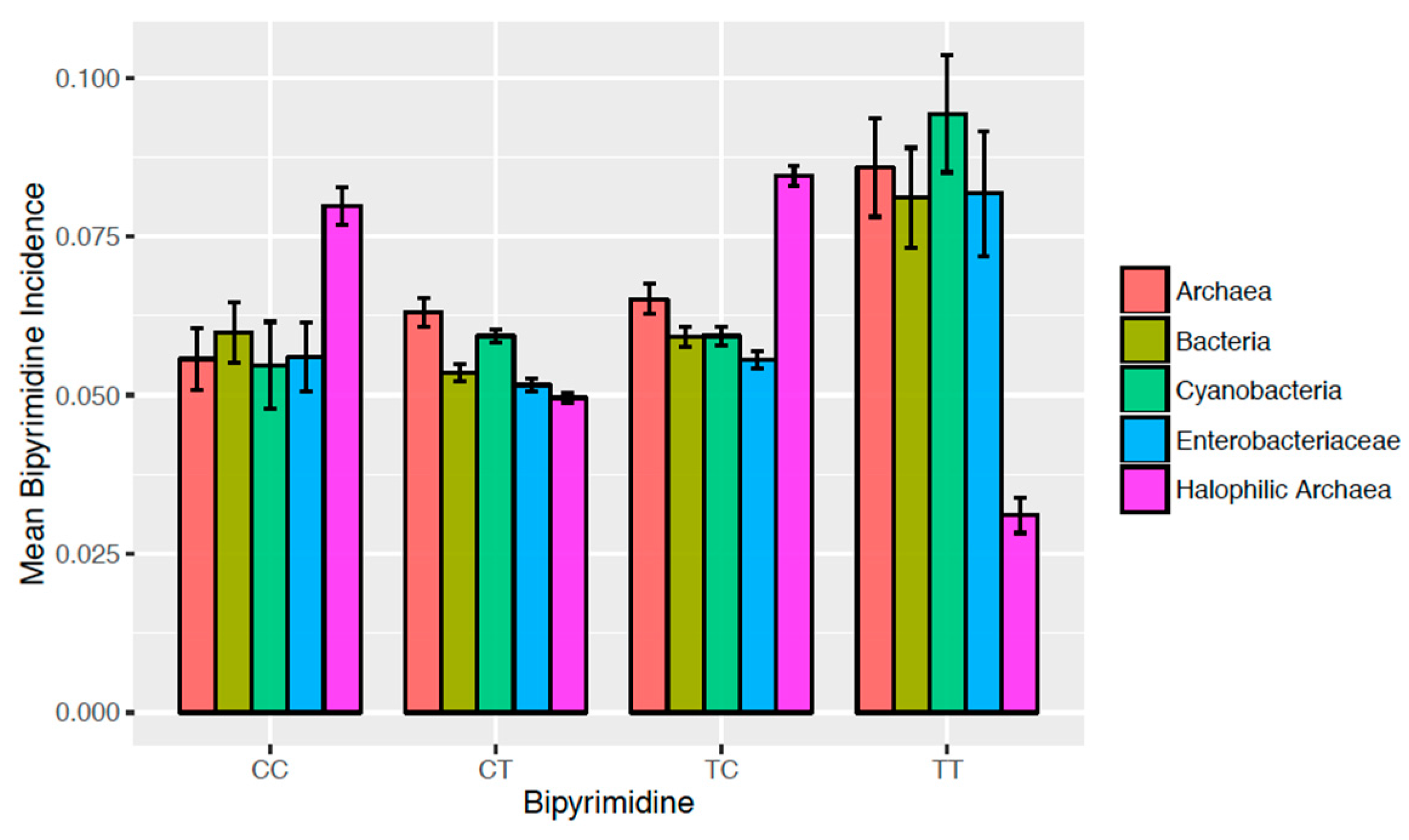
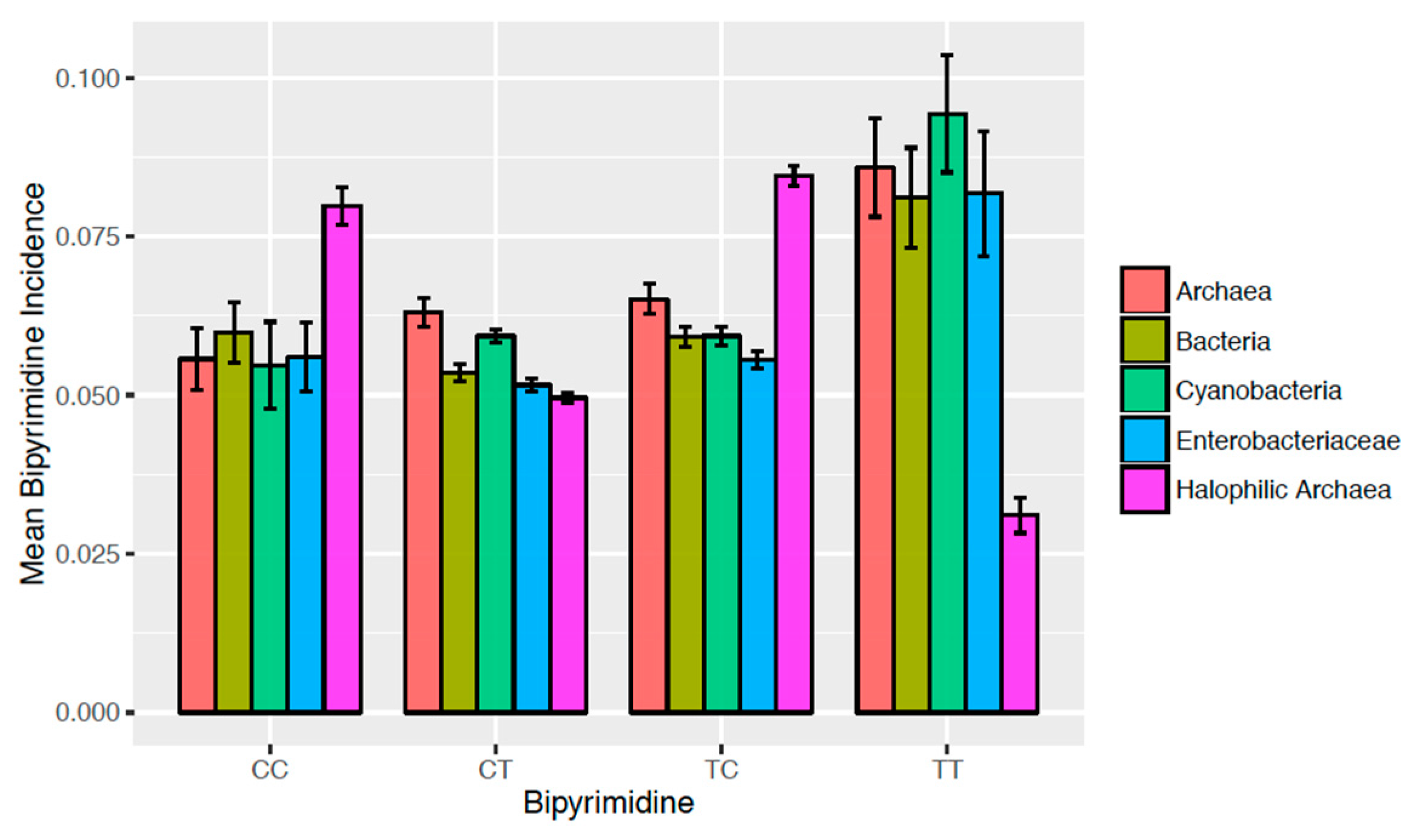
3.2. Bipyrimidine Signature of Halophilic Archaea
- Halophilic archaea have larger CCi than any other group (p < 10−4 each). No other significant intergroup differences in CCi were detected.
- Halophilic archaea have smaller CTi than archaea and cyanobacteria (p < 10−4 each). Other significant differences in CTi were found between enterobacteriaceae vs. archaea (p < 10−4), cyanobacteria vs. bacteria (p = 2.35 × 10−4), and enterobacteriaceae vs. cyanobacteria (p < 10−4).
- Halophilic archaea have larger TCi than any other group (p < 10−4 each). Archaea have the next highest level of TCi, being larger than bacteria (p < 10−4), cyanobacteria (p = 3.27 × 10−3), and enterobacteriaceae (p < 10−4).
- Halophilic archaea have smaller TTi than any other group (p < 10−4 each). No other significant intergroup differences in TTi were detected.
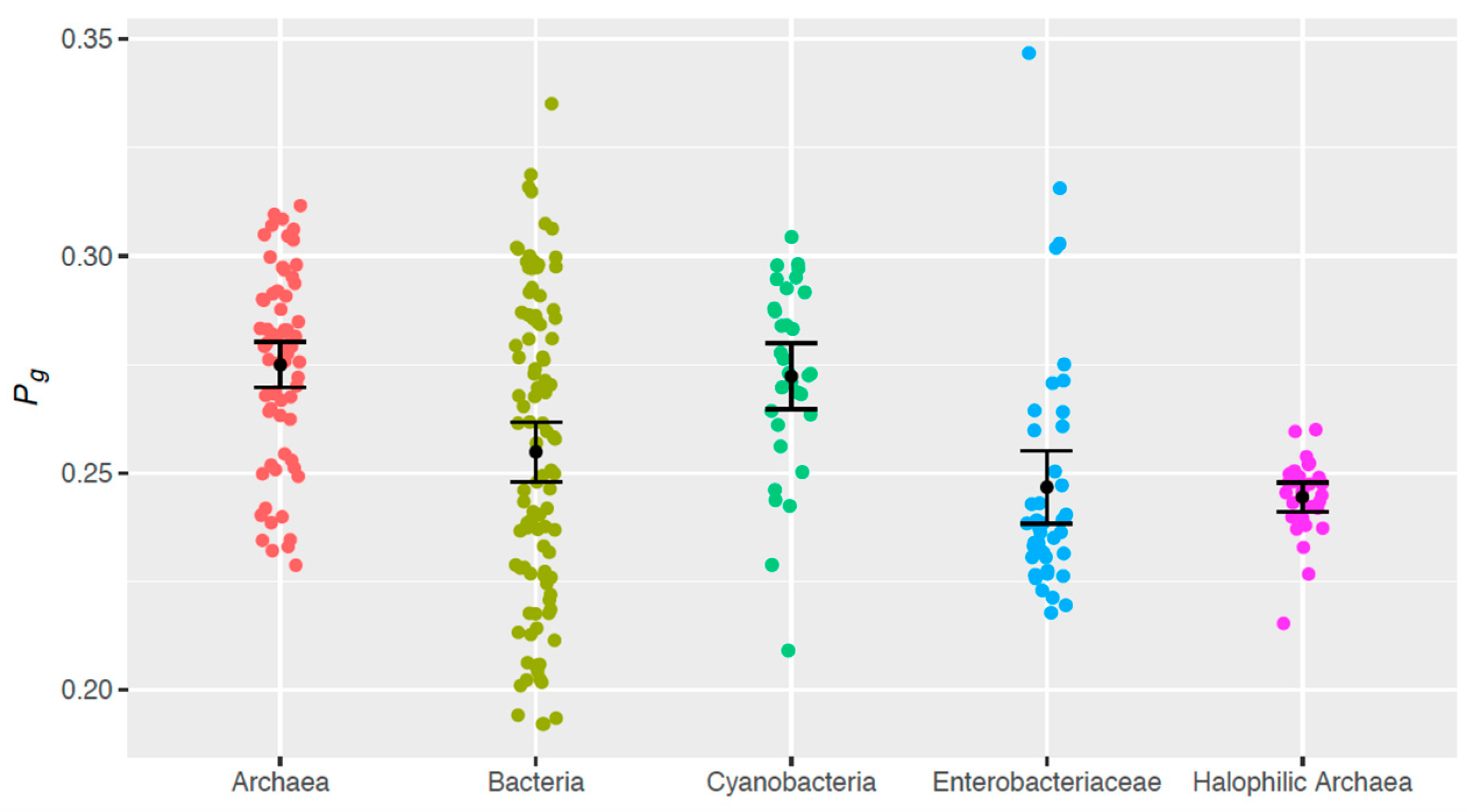
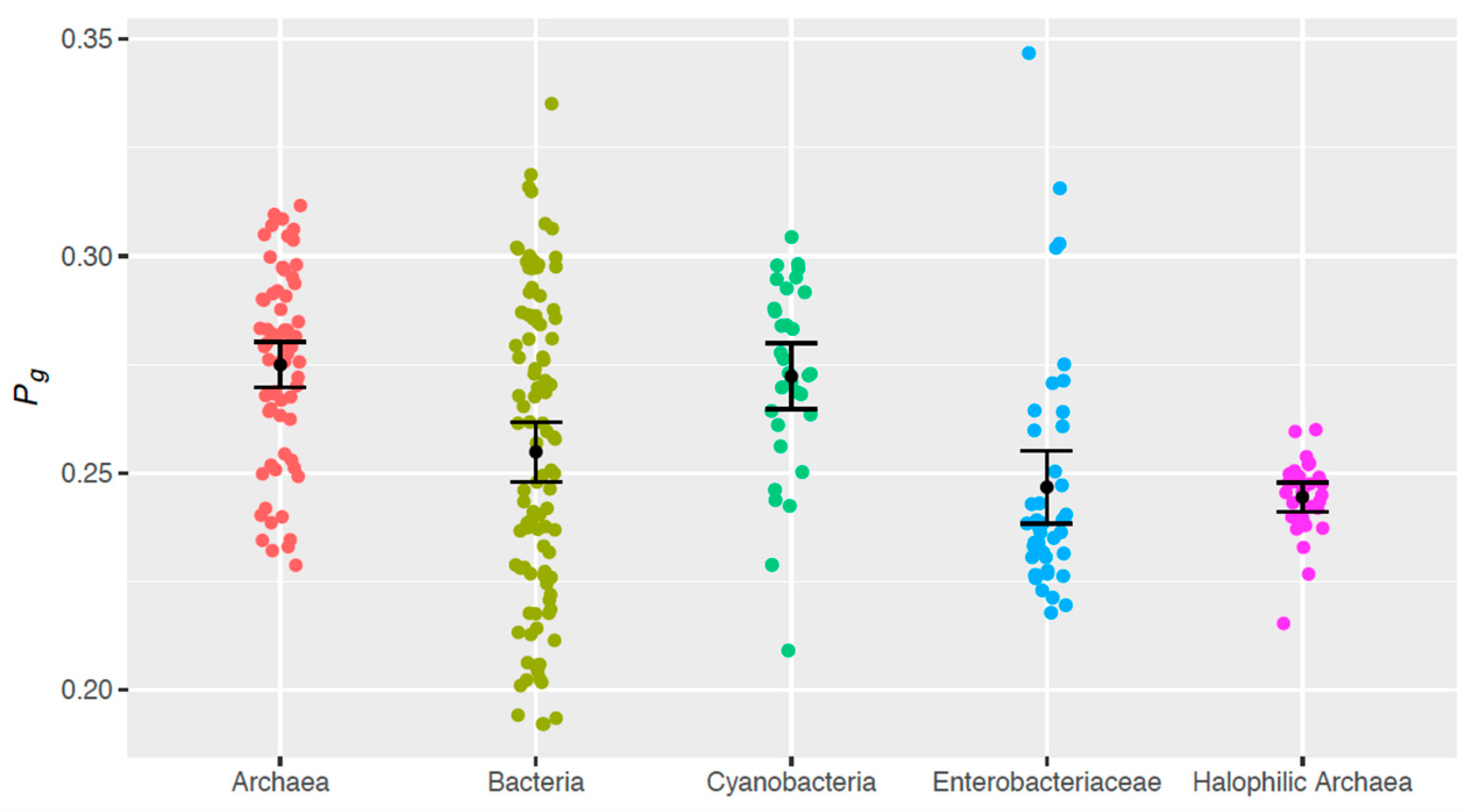
3.3. Intergroup Differences in Theoretical Genomic Photoreactivity (Pg)
- Halophilic archaeal genomes have a smaller Pg than do genomes from (non-halophilic) archaea (p < 10−3) and cyanobacteria (p = 1.01 × 10−3).
- Similarly, enterobacteriaceae genomes are less photoreactive than (non-halophilic) archaea (p < 10−3) and cyanobacteria (p = 1.02 × 10−3).
- (Non-halophilic) archaeal genomes have a larger Pg than do genomes from the other sample groups, with the exception of cyanobacteria (p < 10−3).
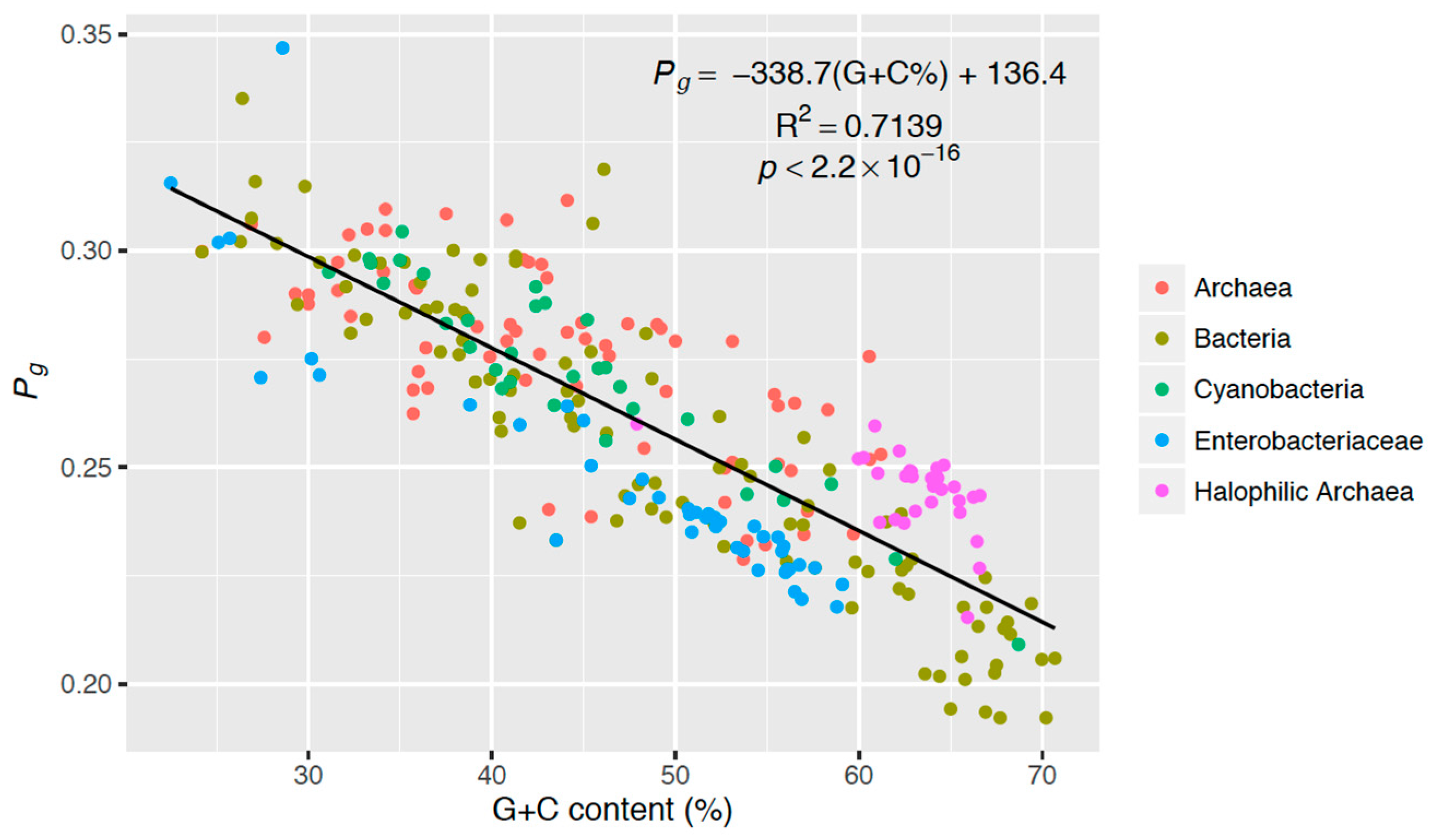
3.4. Genomic Strategy of Photoprotection
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Baxter, B.K.; Litchfield, C.D.; Sowers, K.; Griffith, J.D.; DasSarma, P.A.; DasSarma, S. Great salt lake microbial diversity. In Adaptation to Life in High Salt Concentrations in Archaea Bacteria, and Eukarya; Gunde-Cimerron, N., Oren, A., Plemenita, A., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 9–21. [Google Scholar]
- Oren, A. Microbial life at high salt concentrations: phylogenetic and metabolic diversity. Saline Syst. 2008, 4, 2. [Google Scholar] [CrossRef] [PubMed]
- Grant, W.D.; Gemmell, R.T.; McGenity, T.J. Halophiles. In Extremophiles: Microbial Life in Extreme Environments; Horikoshi, K., Grant, W.D., Eds.; Wiley-Liss: New York, NY, USA, 1998; pp. 93–132. [Google Scholar]
- Almeida-Dalmet, S.; Sikaroodi, M.; Gillevet, P.M.; Litchfield, C.D.; Baxter, B.K. Temporal study of the microbial diversity of the north arm of Great Salt Lake, Utah. Microorganisms 2015, 3, 310–326. [Google Scholar] [CrossRef]
- Dundas, I.D.; Larsen, H. A study on the killing by light of photosensitized cells of Halobacterium Salinarium. Arch. Microbiol. 1963, 46, 19–28. [Google Scholar] [CrossRef]
- Baxter, B.K.; Eddington, B.; Riddle, M.R.; Webster, T.N.; Avery, B.J. Great Salt Lake halophilic microorganisms as models for astrobiology: Evidence for desiccation tolerance and ultraviolet radiation resistance. In Proceeding of the SPIE 6694, Instruments, Methods, and Missions for Astrobiology X, San Diego, CA, USA, 26 August 2007; p. 669415. [CrossRef]
- Hescox, M.A.; Carlberg, D.M. Photoreactivation in Halobacterium cutirubrum. Can. J. Microbiol. 1972, 18, 981–985. [Google Scholar] [CrossRef] [PubMed]
- McCready, S. The repair of ultraviolet light-induced DNA damage in the halophilic archaebacteria, Halobacterium cutirubrum, Halobacterium halobium and Haloferax volcanii. Mutat. Res. 1996, 364, 25–32. [Google Scholar] [CrossRef]
- McCready, S.; Marcello, L. Repair of UV damage in Halobacterium salinarum. Biochem. Soc. Trans. 2003, 31, 694–698. [Google Scholar] [CrossRef] [PubMed]
- Crowley, D.J.; Boubriak, I.; Berquist, B.R.; Clark, M.; Richard, E.; Sullivan, L.; DasSarma, S.; McCready, S. The uvrA, uvrB and uvrC genes are required for repair of ultraviolet light induced DNA photoproducts in Halobacterium sp. NRC-1. Saline Syst. 2006, 2, 11. [Google Scholar] [CrossRef] [PubMed]
- Stantial, N.; Dumpe, J.; Pietrosimone, K.; Baltazar, F.; Crowley, D.J. Transcription-coupled repair of UV damage in the halophilic archaea. DNA Repair (Amst) 2016, 11, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Hepburn, D.; Fitt, P.S. Photoreactivation in pigmented and non-pigmented extreme halophiles. Biochim. Biophys. Acta 1984, 799, 135–142. [Google Scholar] [CrossRef]
- Shahmohammadi, H.R.; Asgarani, E.; Terato, H.; Saito, T.; Ohyama, Y.; Gekko, K.; Yamamoto, O.; Ide, H. Protective roles of bacterioruberin and intracellular KCl in the resistance of Halobacterium salinarium against DNA-Damaging agents. J. Radiat. Res. 1998, 39, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Kelly, M.; Norgard, S.; Liaaen-Jensen, S. C50-carotenoids. 5. carotenoids of Halobacterium Salinarium, especially bacterioruberin. Acta Chem. Scand. 1970, 24, 2169–2182. [Google Scholar] [CrossRef] [PubMed]
- Kushwaha, S.C.; Gochnauer, M.B.; Kushner, D.J.; Kates, M. Pigments and isoprenoid compounds in extremely and moderately halophilic bacteria. Can. J. Microbiol. 1974, 20, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Kushwaha, S.C.; Kramer, J.K.G.; Kates, M. Isolation and characterization of C50-carotenoid pigments and other polar isoprenoids from Halobacterium cutirubrum. Biochim. Biophys. Acta 1975, 398, 303–314. [Google Scholar] [CrossRef]
- Marshall, C.P.; Leuko, S.; Coyle, C.M.; Walter, M.R.; Burns, B.P.; Neilan, B.A. Carotenoid analysis of halophilic archaea by resonance raman spectroscopy. Astrobiology 2007, 7, 631–643. [Google Scholar] [CrossRef] [PubMed]
- DasSarma, S.; Kennedy, S.P.; Berquist, B.; Ng, W.V.; Baliga, N.S.; Spudich, J.L.; Krebs, M.P.; Eisen, J.A.; Johnson, E.A.; Johnson, C.H.; Hood, L. Genomic perspective on the photobiology of Halobacterium species NRC-1, a phototrophic, phototactic, and UV-tolerant haloarchaeon. Photosynth. Res. 2001, 70, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Oesterhelt, D.; Stoekenius, W. Functions of a new photoreceptor membrane. Proc. Natl. Acad. Sci. USA 1973, 70, 2853–2857. [Google Scholar] [CrossRef] [PubMed]
- Papke, R.T.; Douady, C.J.; Doolittle, W.F.; Rodriguez-Valera, F. Diversity of bacteriorhodopsins in different hypersaline waters from a single Spanish saltern. Environ. Microbiol. 2003, 5, 1039–1045. [Google Scholar] [CrossRef] [PubMed]
- NCBI Genome Database. Available online: http://www.ncbi.nlm.nih.gov/genome/browse/# (accessed on 23 August 2016).
- Haynes, R.H. Molecular localization of radiation damage relevant to bacterial inactivation. In Physical Processes in Radiation Biology; Augenstein, L., Mason, R., Rosenberg, B., Eds.; Academic Press: New York, NY, USA, 1964; p. 51. [Google Scholar]
- Setlow, R.B. Cyclobutane-type pyrimidine dimers in polynucleotides. Science 1966, 153, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Setlow, R.B.; Carrier, W.L. Pyrimidine dimers in ultraviolet-Irradiated DNA’s. J. Mol. Biol. 1966, 17, 237–254. [Google Scholar] [CrossRef]
- Joux, F.; Jeffrey, W.H.; Lebaron, P.; Mitchell, D.L. Marine bacterial isolates display diverse responses to UV-B radiation. Appl. Environ. Microbiol. 1999, 65, 93820–93827. [Google Scholar]
- Kennedy, S.P.; Ng, W.V.; Salzberg, S.L.; Hood, L.; DasSarma, S. Understanding the adaptation of Halobacterium species NRC-1 to its extreme environment through computational analysis of its genome sequence. Genome. Res. 2001, 11, 1641–1650. [Google Scholar] [CrossRef] [PubMed]
- Douki, T.; Cadet, J. Individual determination of the yield of the main UV-Induced dimeric pyrimidine photoproducts in DNA suggests a high mutagenicity of CC photolesions. Biochemistry 2001, 40, 2495–2501. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.P.; Häder, D.P. UV-induced DNA damage and repair: A review. Photochem. Photobiol. Sci. 2002, 1, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Besaratinia, A.; Yoon, J.; Schroeder, C.; Bradforth, S.E.; Cockburn, M.; Pfeifer, G.P. Wavelength dependence of ultraviolet radiation-induced DNA damage as determined by laser irradiation suggests that cyclobutane pyrimidine dimers are the principal DNA lesions produced by terrestrial sunlight. FASEB J. 2011, 25, 3079–3091. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D.L.; Jen, J.; Cleaver, J.E. Sequence specificity of cyclobutane pyrimidine dimers in DNA treated with solar (ultraviolet B) radiation. Nucleic Acids Res. 1991, 20, 225–229. [Google Scholar] [CrossRef]
- Yoon, J.H.; Lee, C.S.; O’Connor, T.R.; Yasui, A.; Pfeifer, G.P. The DNA damage spectrum produced by simulated sunlight. J. Mol. Biol. 2000, 299, 681–693. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, R.P.; Richa; Kumar, A.; Tyagi, M.B.; Sinha, R.P. Molecular Mechanisms of Ultraviolet Radiation-Induced DNA Damage and Repair. J. Nucleic Acids 2010, 2010, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Matallana-Surget, S.; Meador, J.A.; Joux, F.; Douki, T. Effect of the GC content of DNA on the distribution of UVB-induced bipyrimidine photoproducts. Photochem. Photobiol. Sci. 2008, 7, 794–801. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.L. Dinucleotide Counts. Available online: https://github.com/jones1040/DinucleotideCounts (accessed on 23 August 2016).
- R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 23 August 2016).
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biometrical. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed]
- Bolhuis, H.; Palm, P.; Wende, A.; Falb, M.; Rampp, M.; Rodriguez-Valera, F.; Pfeiffer, F.; Oesterhelt, D. The genome of the square archaeon Haloquadratum walsbyi: Life at the limits of water activity. BMC Genom. 2006, 7, 169. [Google Scholar] [CrossRef] [PubMed]
- Burns, D.G.; Janssen, P.H.; Itoh, T.; Kamekura, M.; Li, Z.; Jensen, G.; Rodrıguez-Valera, F.; Bolhuis, H.; Dyall-Smith, M. Haloquadratum walsbyi gen. nov., sp. nov., the square haloarchaeon of Walsby, isolated from saltern crystallizers in Australia and Spain. Int. J. Syst. Evolut. Microbiol. 2007, 57, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Porter, K.; Russ, B.; Burns, D.; Dyall-Smith, M. Diversity of Haloquadratum and other haloarchaea in three, geographically distant, Australian saltern crystallizer ponds. Extremophiles 2010, 14, 161–169. [Google Scholar]
- Sancar, G.B. Enzymatic photoreactivation: 50 years and counting. Mutat. Res. 2000, 451, 25–37. [Google Scholar] [CrossRef]
- Weber, S. Light-driven enzymatic catalysis of DNA repair: A review of recent biophysical studies on photolyase. Biochim. Biophys. Acta 2005, 1707, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Sancar, A. DNA excision repair. Annu Rev. Biochem. 1996, 65, 43–81. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, R.L.; Björn, L.O.; Bais, A.; Ilyasd, M. Changes in biologically active ultraviolet radiation reaching the Earth's surface. Photochem. Photobiol. Sci. 2003, 2, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Chou, M.D.; Lee, K.T. Parameterizations for the absorption of solar radiation by water vapor and ozone. J. Atmos. Sci. 1996, 53, 1203–1208. [Google Scholar] [CrossRef]
- Litchfield, C.D. Survival strategies for microorganisms in hypersaline environments and their relevance to life on early Mars. Meteorit. Planet Sci. 1998, 33, 813–819. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Bag, S.K.; Das, S.; Harvill, E.T.; Dutta, C. Molecular signature of hypersaline adaptation: Insights from genome and proteome composition of halophilic prokaryotes. Genome Biol. 2008, 9, R70. [Google Scholar] [CrossRef] [PubMed]
- Knight, R.D.; Freeland, S.J.; Landweber, L.F. A simple model based on mutation and selection explains trends in codon and amino-acid usage and GC composition within and across genomes. Genome Biol. 2001, 2, research0010.1–0010.13. [Google Scholar] [CrossRef]
- Musto, H.; Naya, H.; Zavala, A.; Romero, H.; Alvarez-Valín, F.; Bernardi, G. Genomic GC level, optimal growth temperature, and genome size in prokaryotes. Biochem. Biophys. Res. Commun. 2006, 347, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Hildebrand, F.; Meyer, A.; Eyre-Walker, A. Evidence of selection upon genomic GC-content in bacteria. PLoS Genet. 2010, 6, e1001107. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain | G + C (%) | Size (Mb) | Genes |
|---|---|---|---|---|
| Halalkalicoccus jeotgali | B3 | 62.56 | 3.699 | 3717 |
| Halanaeroarchaeum sulfurireducens | HSR2 | 62.86 | 2.210 | 2213 |
| Haloarcula hispanica | N601 | 62.47 | 3.902 | 3825 |
| Haloarcula marismortui | ATCC 43049 | 61.14 | 4.275 | 4226 |
| Haloarcula sp. | CBA1115 | 61.98 | 4.225 | 4108 |
| Halobacterium hubeiense | JI20-1 | 66.58 | 3.130 | 3189 |
| Halobacterium salinarum | NRC-1 | 65.92 | 2.571 | 2629 |
| Halobacterium sp. | DL1 | 66.44 | 3.163 | 3237 |
| Haloferax gibbonsii | ARA6 | 66.24 | 3.918 | 3783 |
| Haloferax mediterranei | ATCC 33500 | 60.26 | 3.905 | 3800 |
| Haloferax volcanii | DS2 | 65.46 | 4.013 | 3925 |
| Halogeometricum borinquense | DSM 11551 | 59.97 | 3.944 | 3838 |
| Halomicrobium mukohataei | DSM 12286 | 65.51 | 3.332 | 3293 |
| Halopiger xanaduensis | SH-6 | 65.20 | 4.355 | 4174 |
| Haloquadratum walsbyi | DSM 16790 | 47.90 | 3.179 | 2827 |
| Halorhabdus tiamatea | SARL4B | 62.77 | 3.146 | 3069 |
| Halorhabdus utahensis | DSM 12940 | 62.90 | 3.117 | 2969 |
| Halorubrum lacusprofundi | ATCC 49239 | 63.95 | 3.693 | 3523 |
| Halostagnicola larsenii | XH-48 | 60.87 | 4.131 | 3954 |
| Haloterrigena turkmenica | DSM 5511 | 64.25 | 5.441 | 5074 |
| Halovivax ruber | XH-70 | 64.30 | 3.224 | 3187 |
| Natrialba magadii | ATCC 43099 | 61.03 | 4.444 | 4128 |
| Natrinema pellirubrum | DSM 15624 | 63.96 | 4.354 | 4178 |
| Natrinema sp. | J7-2 | 64.06 | 3.794 | 3681 |
| Natronobacterium gregoryi | SP2 | 62.20 | 3.788 | 3720 |
| Natronococcus occultus | SP4 | 64.63 | 4.314 | 4162 |
| Natronomonas moolapensis | 8.8.11 | 64.50 | 2.913 | 2793 |
| Natronomonas pharaonis | DSM 2160 | 63.08 | 2.750 | 2799 |
| Salinarchaeum sp. | Harcht-Bsk1 | 66.60 | 3.255 | 3036 |
| Sample Group | Mean G + C Content (%) | Median (%) |
|---|---|---|
| (Non-Halophilic) Archaea | 43.87 +/− 2.30 | 43.05 |
| Bacteria | 49.10 +/− 2.55 | 47.25 |
| Cyanobacteria | 44.21 +/− 3.08 | 42.65 |
| Enterobacteriaceae | 47.83 +/− 3.17 | 51.73 |
| Halophilic Archaea | 63.09 +/− 1.28 | 63.95 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jones, D.L.; Baxter, B.K. Bipyrimidine Signatures as a Photoprotective Genome Strategy in G + C-rich Halophilic Archaea. Life 2016, 6, 37. https://doi.org/10.3390/life6030037
Jones DL, Baxter BK. Bipyrimidine Signatures as a Photoprotective Genome Strategy in G + C-rich Halophilic Archaea. Life. 2016; 6(3):37. https://doi.org/10.3390/life6030037
Chicago/Turabian StyleJones, Daniel L., and Bonnie K. Baxter. 2016. "Bipyrimidine Signatures as a Photoprotective Genome Strategy in G + C-rich Halophilic Archaea" Life 6, no. 3: 37. https://doi.org/10.3390/life6030037