Spread of a Gammabaculovirus within Larval Populations of Its Natural Balsam Fir Sawfly (Neodiprion abietis) Host Following Its Aerial Application

Abstract
:1. Introduction
2. Results and Discussion
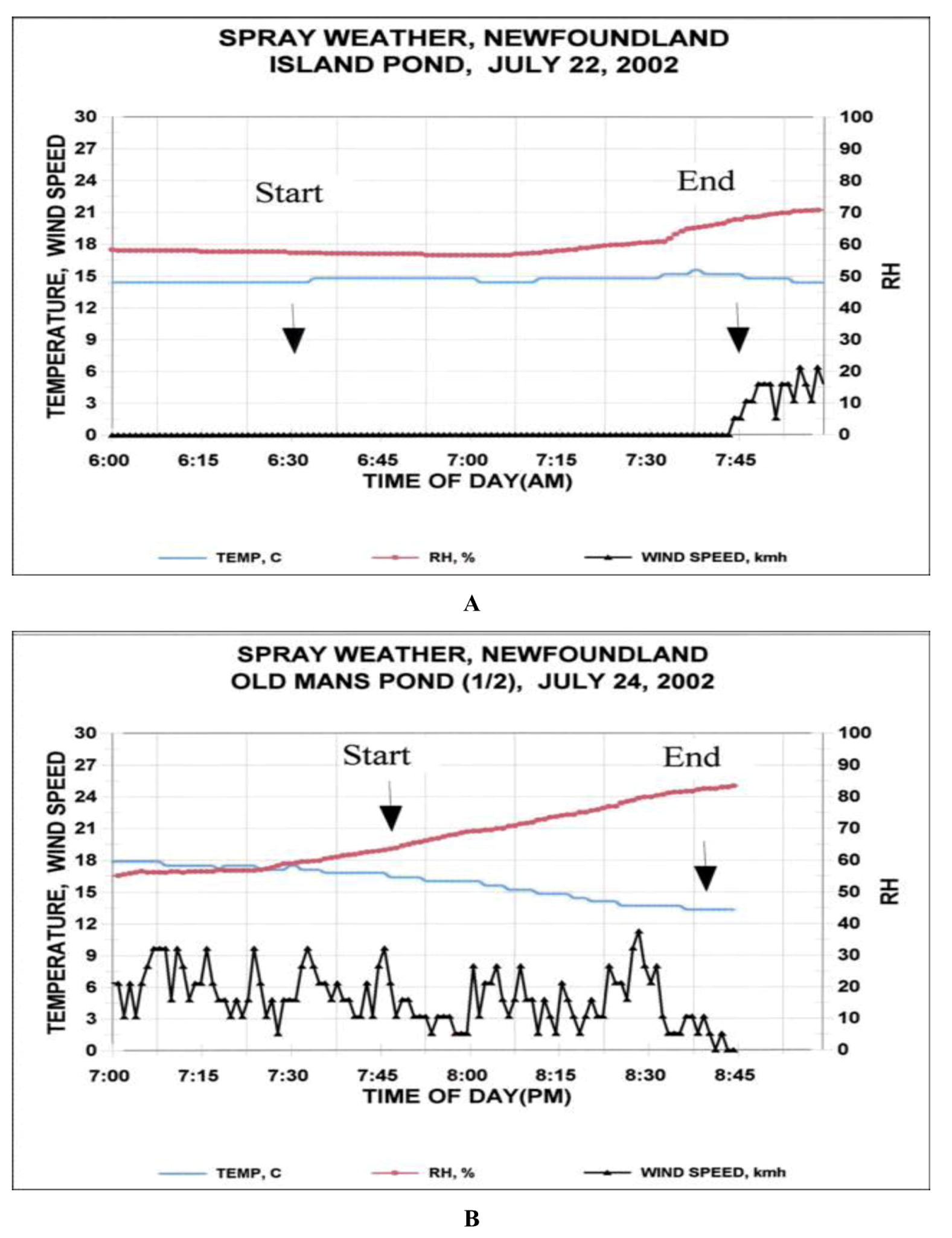
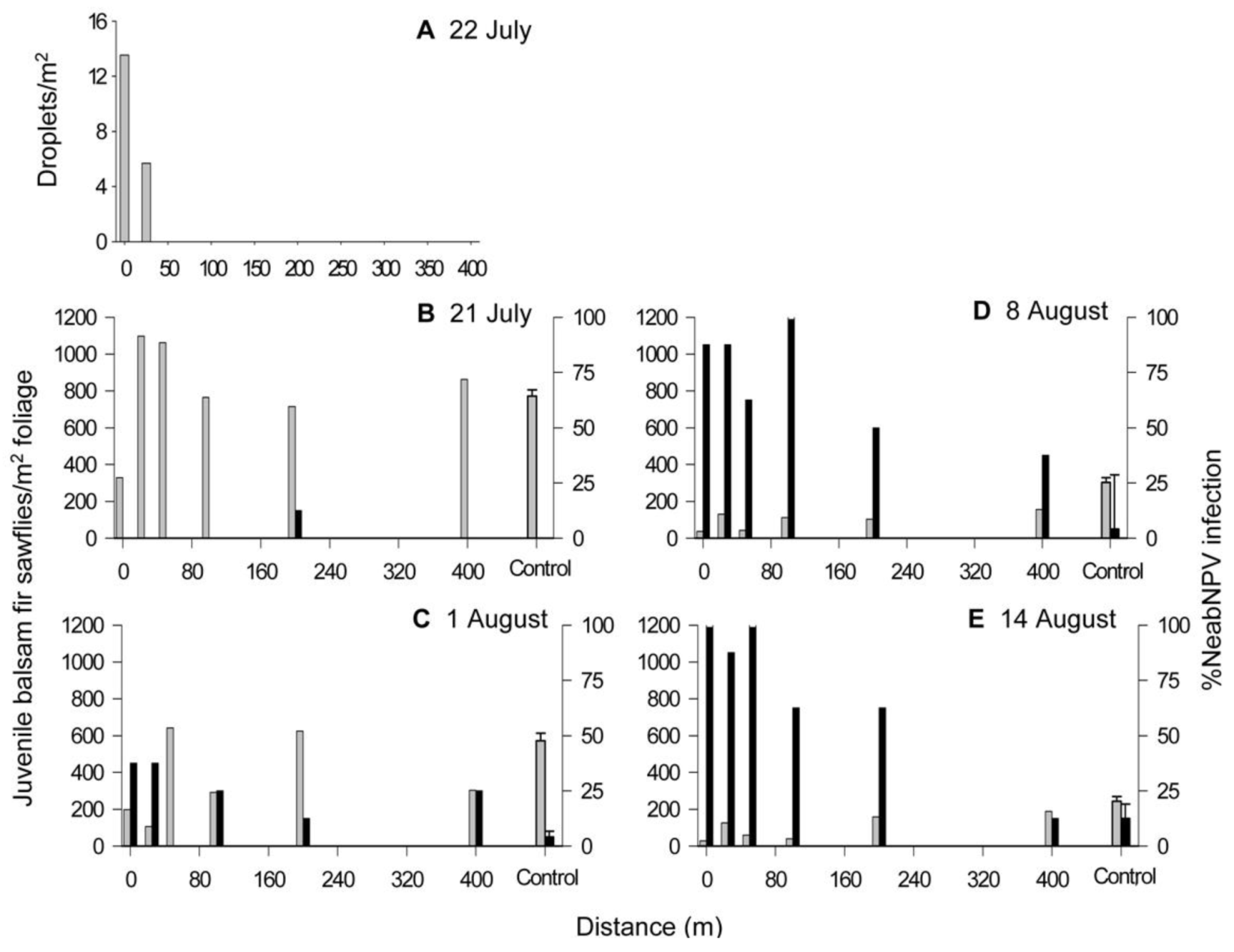
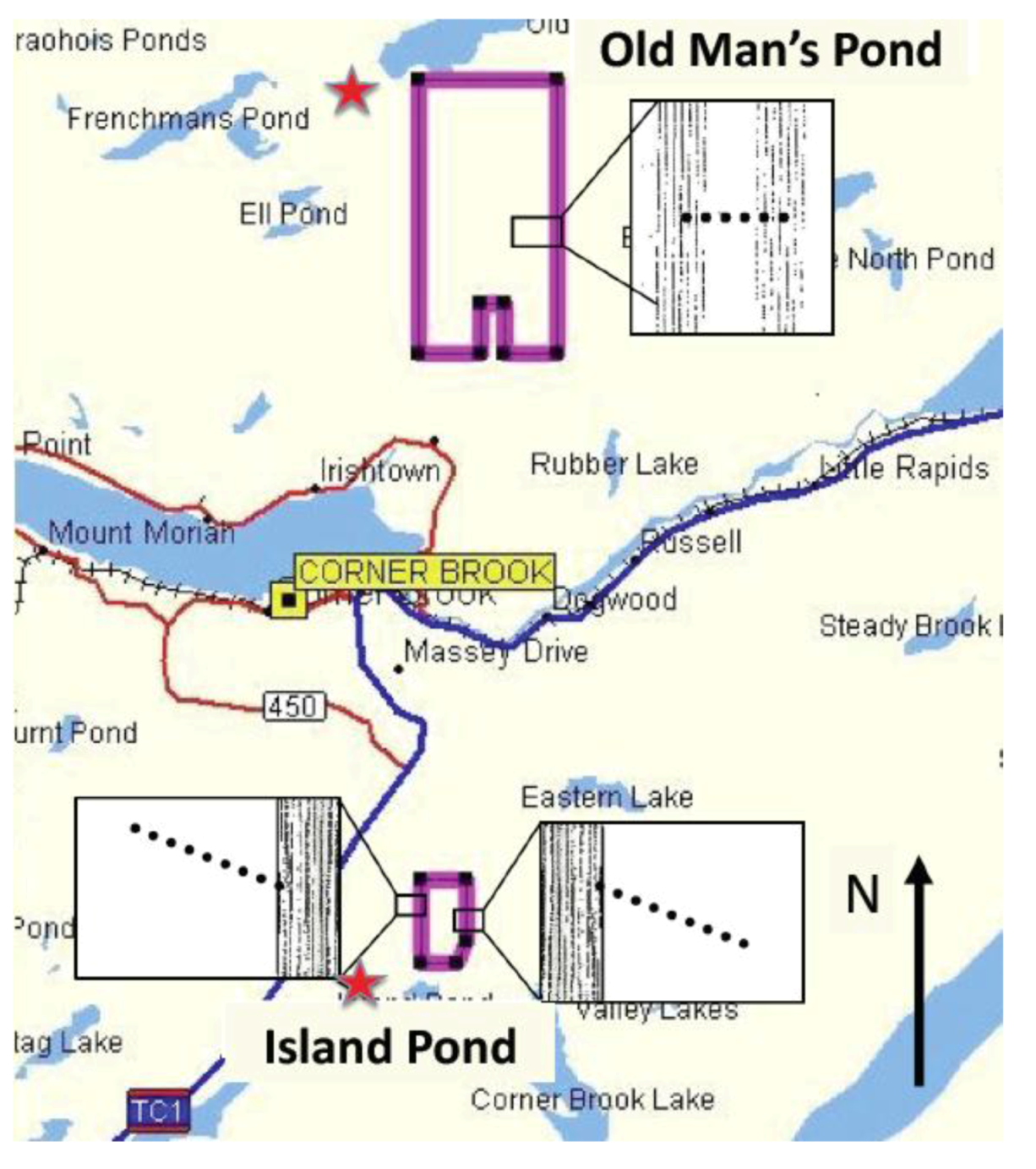
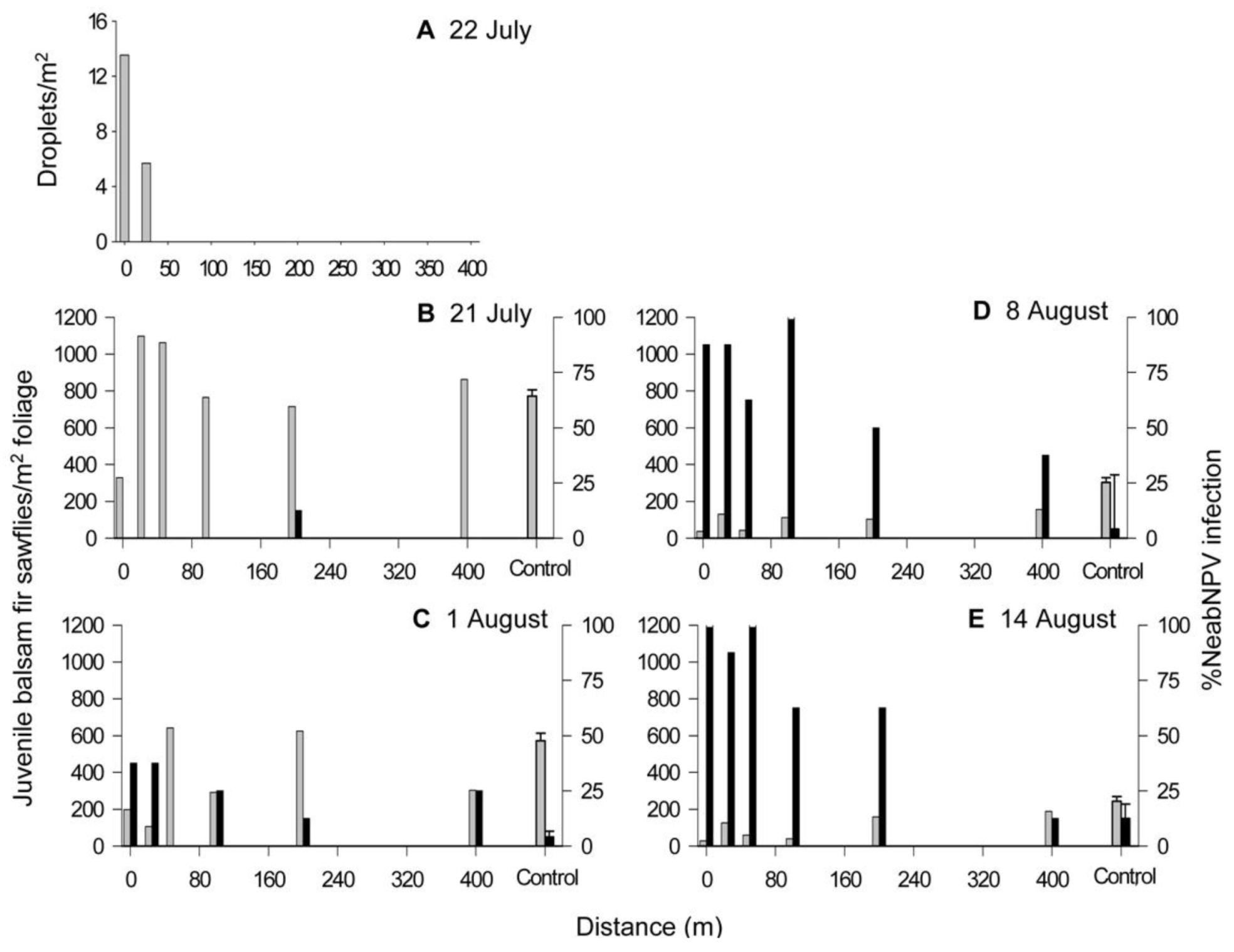
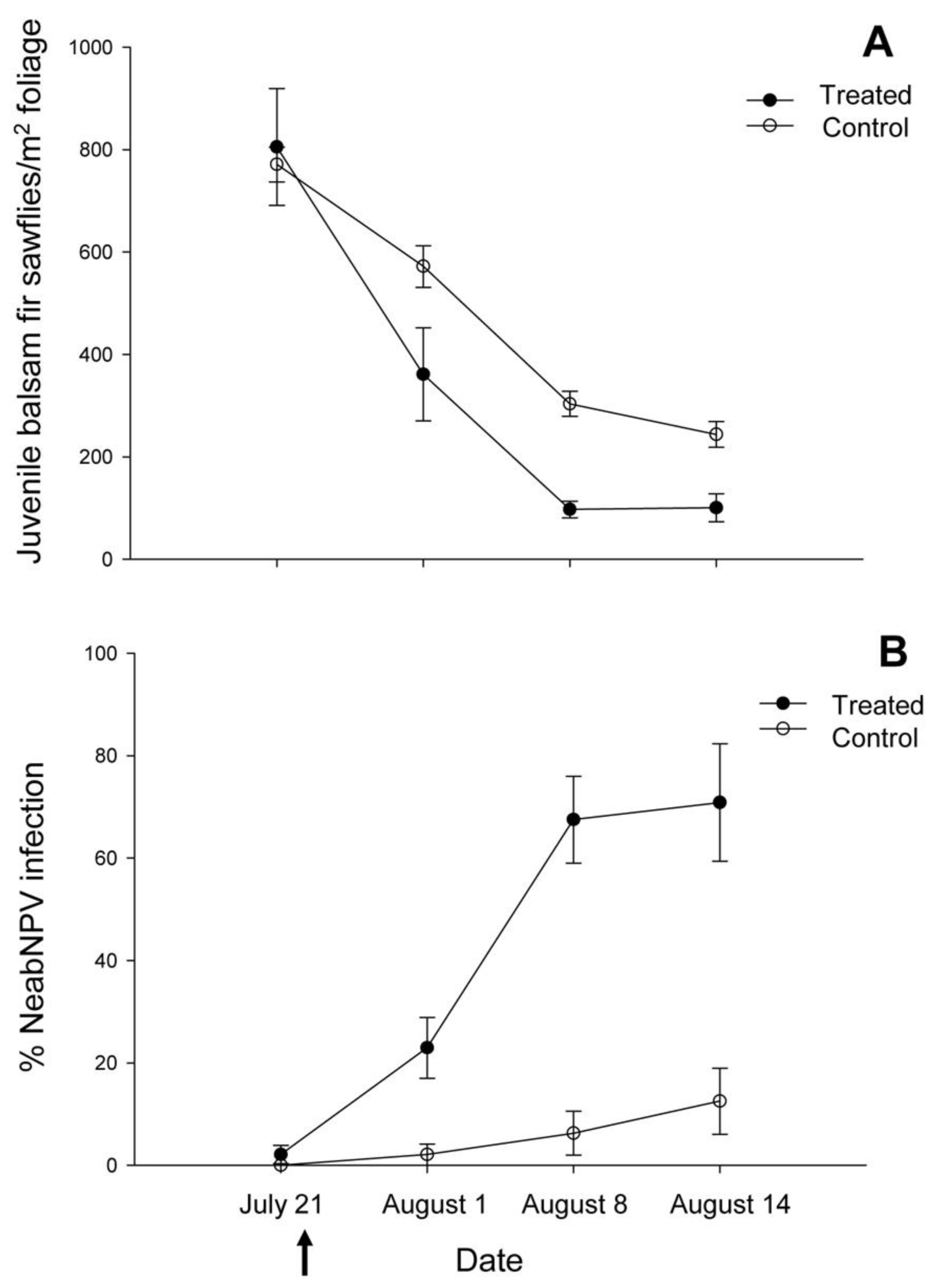
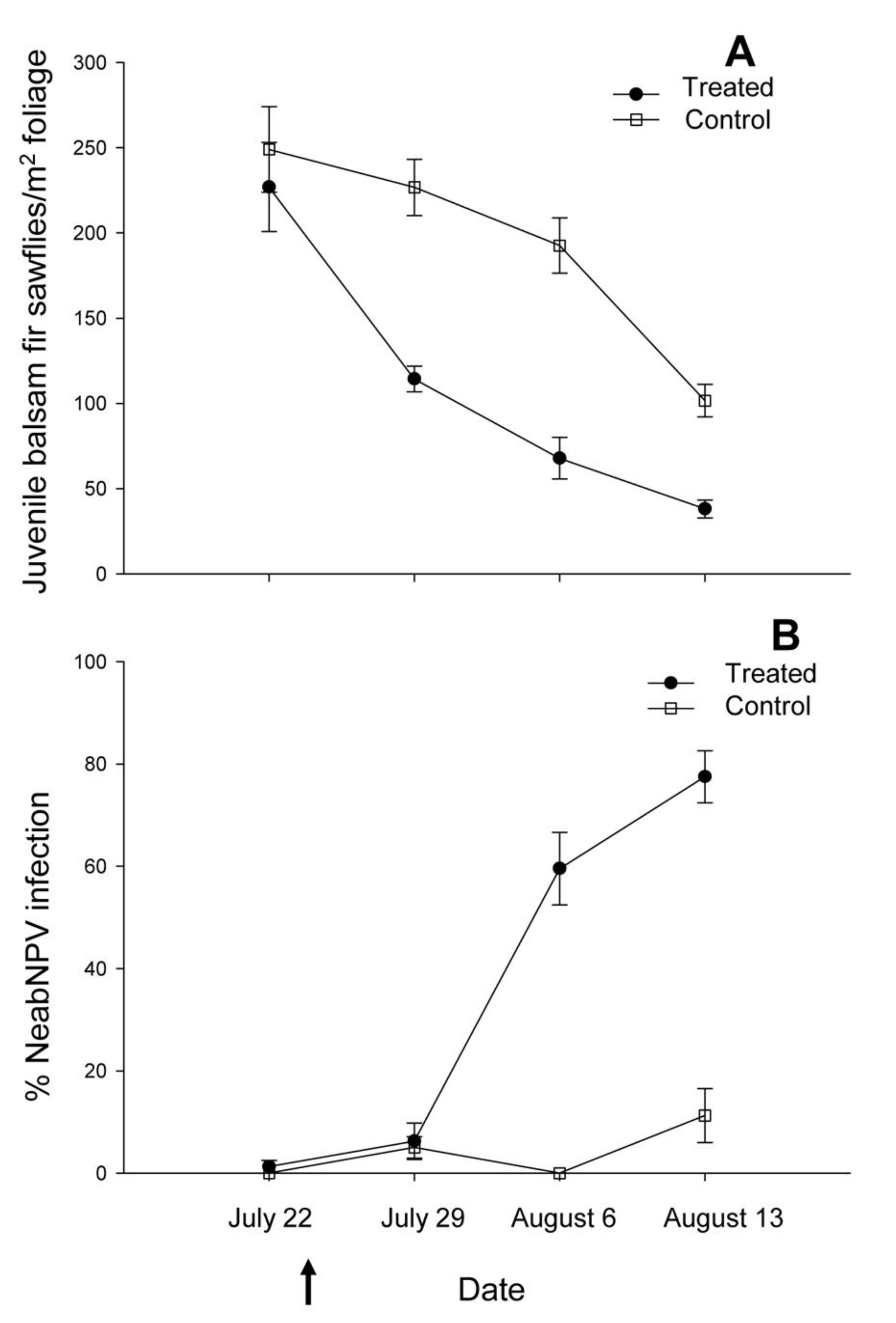
2.1. Island Pond
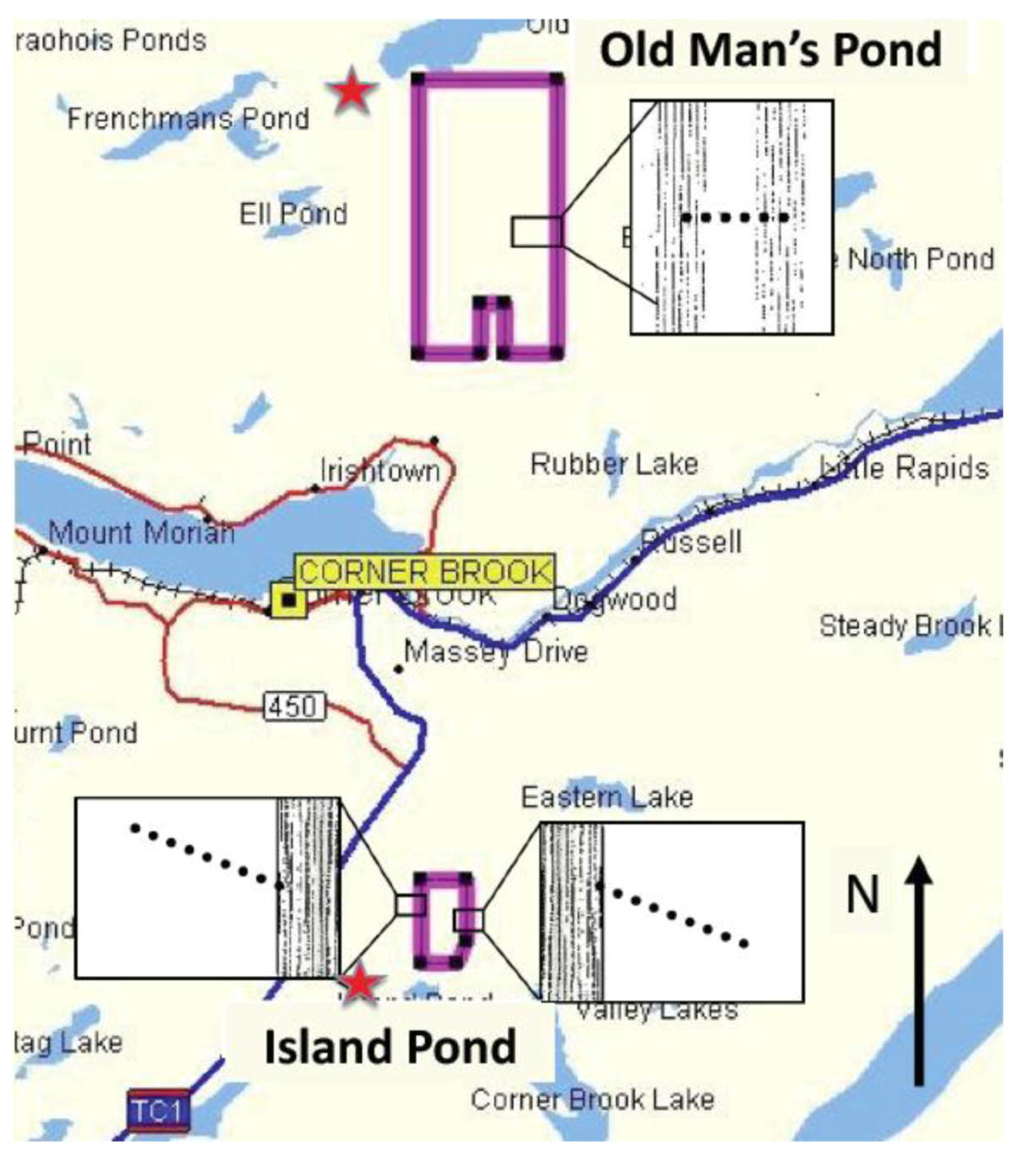
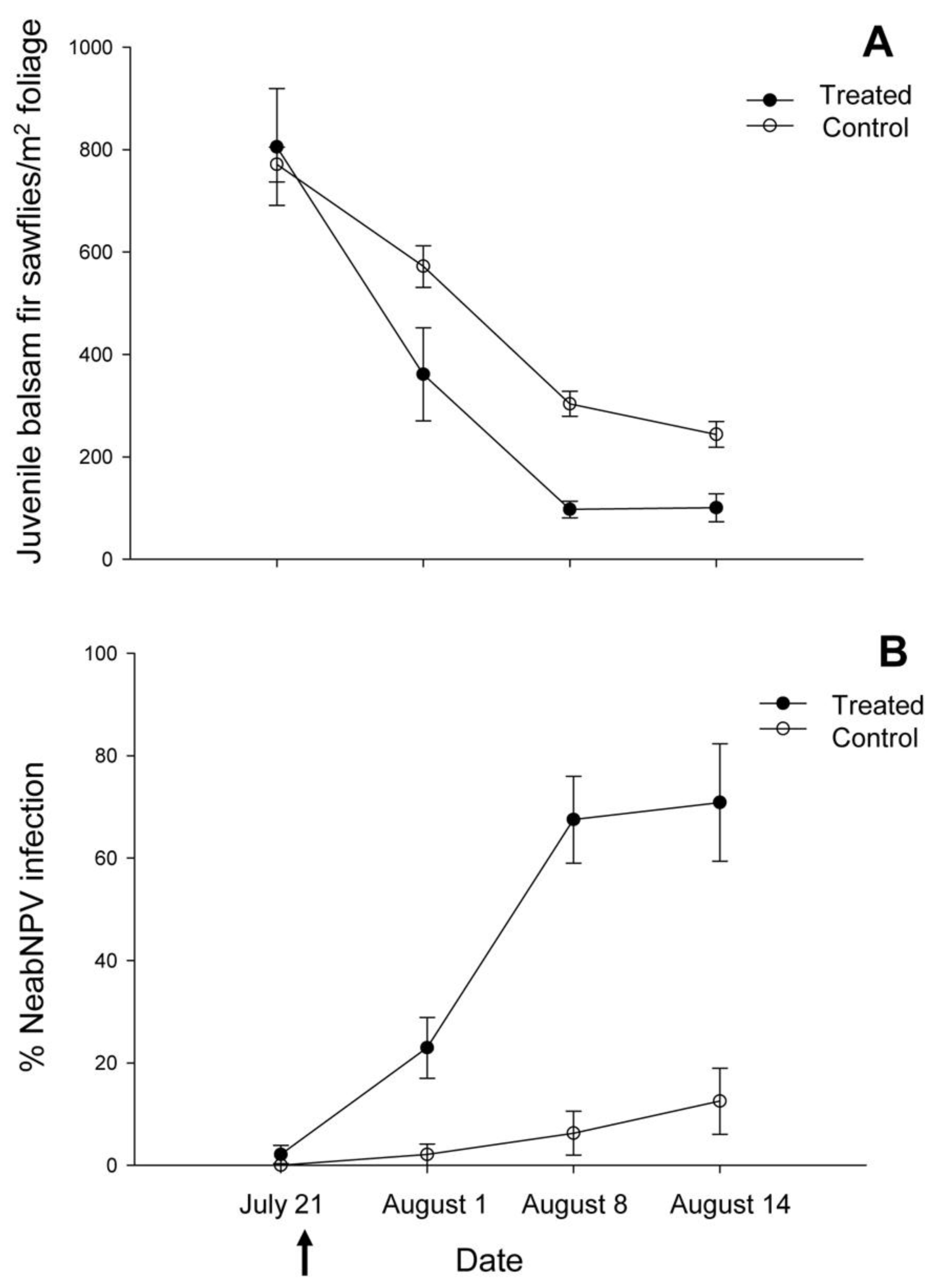

{kind=link}
{kind=link}
{kind=link}
{kind=link}
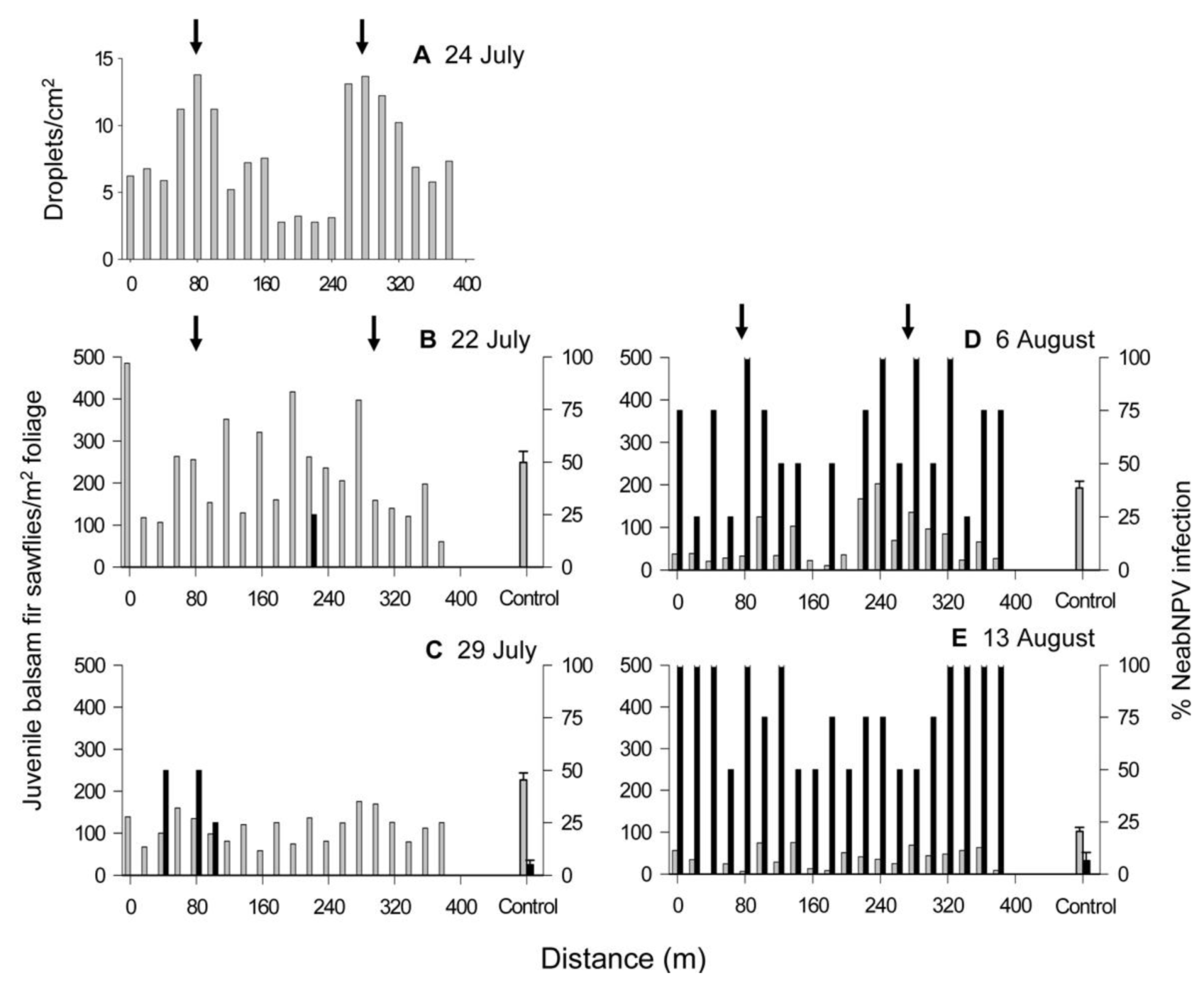{kind=link}
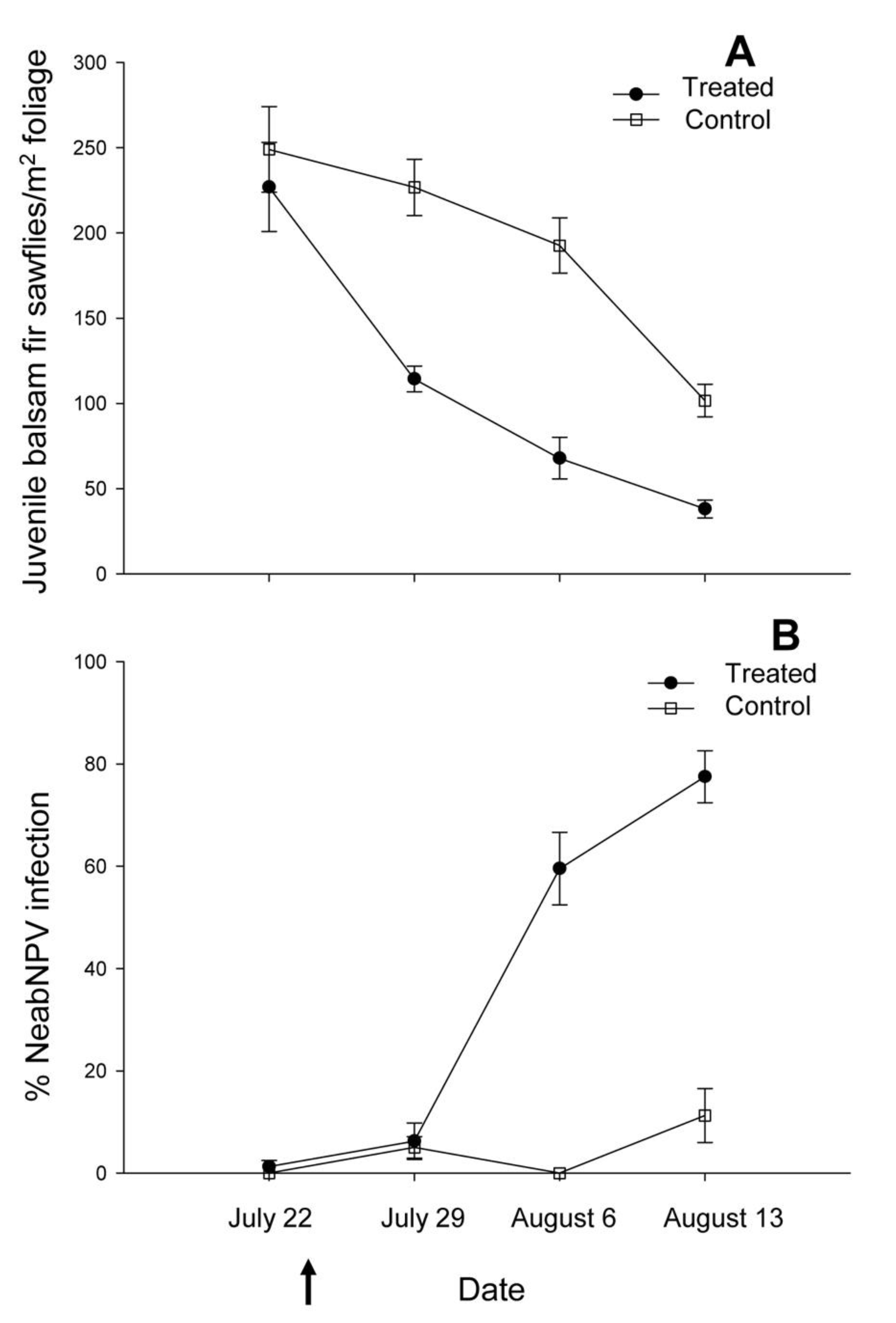{kind=link}
{kind=link}
| Treatment | Weeks post spray | N | Density Pearson’s r | p value | Infection Pearson’s r | p value |
|---|---|---|---|---|---|---|
| Control | 1 | 12 | −0.469 | 0.12 | −0.343 | 0.27 |
| Control | 2 | 12 | −0.254 | 0.43 | −0.229 | 0.47 |
| Control | 3 | 12 | −0.434 | 0.16 | −0.421 | 0.17 |
| NeabNPV | 1 | 12 | 0.156 | 0.63 | −0.103 | 0.75 |
| NeabNPV | 2 | 12 | 0.466 | 0.13 | −0.718 | 0.01 |
| NeabNPV | 3 | 12 | 0.722 | 0.01 | −0.909 | 0.00 |
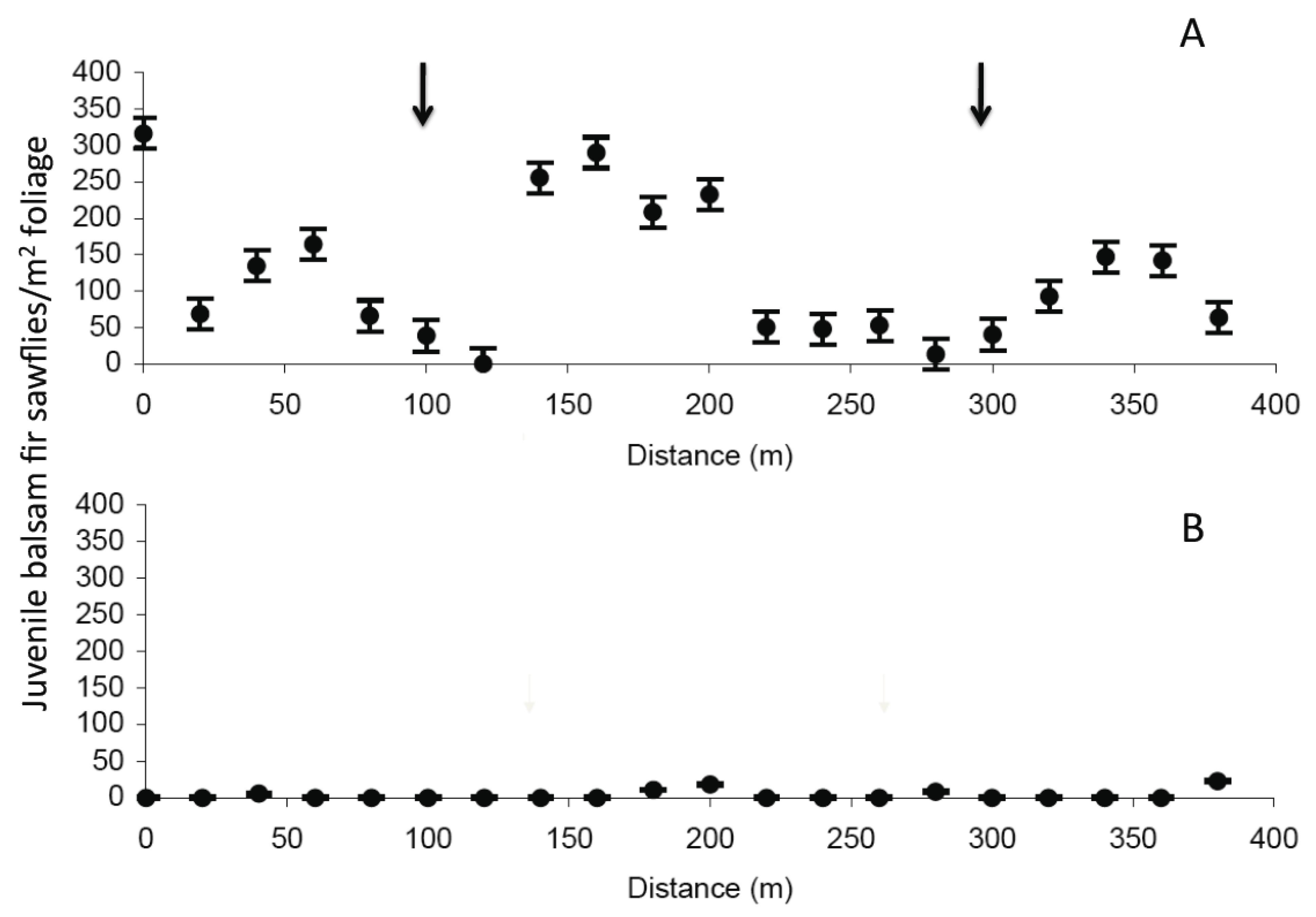
2.2. Old Man’s Pond

| Treatment | Weeks post spray | N | Density Pearson’s r | p value | Infection Pearson’s r | p value |
|---|---|---|---|---|---|---|
| Control | 1 | 20 | −0.237 | 0.31 | 0.150 | 0.02 |
| Control | 2 | 20 | 0.505 | 0.62 | -* | -* |
| Control | 3 | 20 | 0.058 | 0.81 | −0.408 | 0.07 |
| NeabNPV | 1 | 20 | −0.418 | 0.07 | −0.402 | 0.01 |
| NeabNPV | 2 | 20 | −0.402 | 0.50 | −0.364 | 0.12 |
| NeabNPV | 3 | 20 | −0.103 | 0.01 | −0.098 | 0.68 |
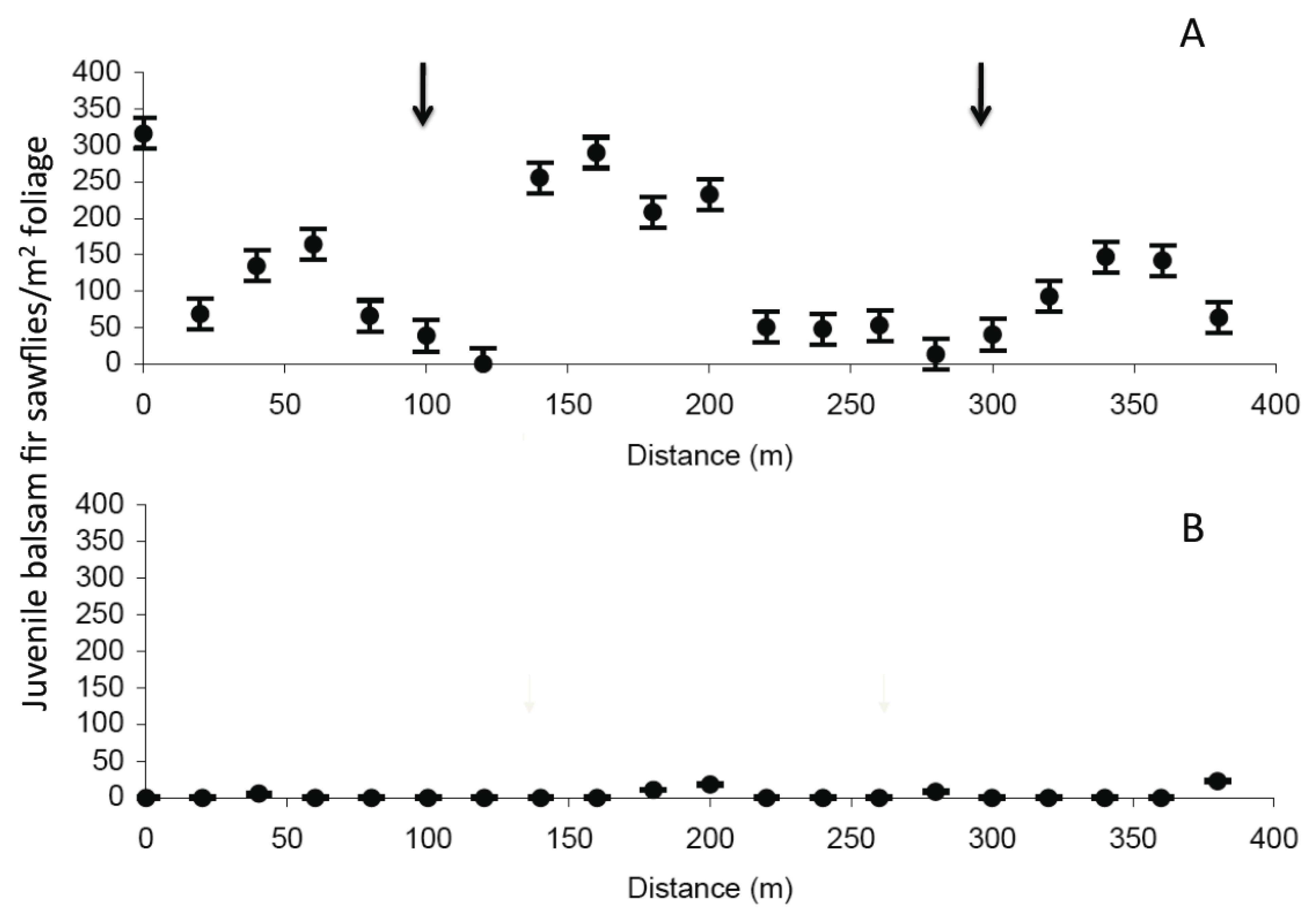
2.3. Significance of NeabNPV Field Trials
3. Materials and Methods
3.1. Aerial Application of NeabNPV
3.2. Amplification and Purification of NeabNPV for Use in Field Trials
3.3. Field Sites and Study Design
3.4. Balsam Fir Sawfly Sampling
3.5. Molecular Probing for NeabNPV
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
References and Notes
- Fuxa, J.R. Ecological considerations for the use of entomopathogens in IPM. Ann. Rev. Entomol. 1987, 32, 225–251. [Google Scholar] [CrossRef]
- Fuxa, J.R. Ecology of insect nucleopolyhedroviruses (Review). Agric. Ecosyst. Environ. 2004, 103, 27–43. [Google Scholar] [CrossRef]
- Payne, N.J. Factors influencing aerial insecticide application to forests. Integr. Pest Manag. Rev. 2000, 5, 1–10. [Google Scholar] [CrossRef]
- Cory, J.S.; Evans, H.F. Viruses. In Field Manual of Techniques in Invertebrate Pathology, 2nd ed.; Lacey, L.A., Kaya, H.K., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 149–174. [Google Scholar]
- Lapointe, R.; Thumbi, D.K.; Lucarotti, C.J. Recent advances in our knowledge of baculovirus molecular biology and its relevance for the registration of baculovirus-based products for insect pest population control. In Integrated Pest Management and Pest Control; Soloneski, S., Larramendy, M.L., Eds.; InTech Open Access Publisher: Rijeka, Croatia, 2012; pp. 481–522. [Google Scholar]
- Lauzon, H.A.M.; Lucarotti, C.J.; Krell, P.J.; Feng, Q.; Retnakaran, A.; Arif, B.M. Sequence and organization of the Neodiprion lecontei nucleopolyhedrovirus genome. J. Virol. 2004, 78, 7023–7035. [Google Scholar]
- Jehle, J.A.; Blissard, G.W.; Bonning, B.C.; Cory, J.S.; Herniou, E.A.; Rohrmann, G.F.; Theilmann, D.A.; Thiem, S.M.; Vlak, J.M. On the classification and nomenclature of baculoviruses: A proposal for revision. Arch. Virol. 2006, 151, 1257–1266. [Google Scholar] [CrossRef]
- Bird, F.T.; Elgee, D.E. A virus disease and introduced parasites as factors controlling the European spruce sawfly, Diprion hercyniae (Htg.), in central New Brunswick. Can. Entomol. 1957, 89, 371–378. [Google Scholar] [CrossRef]
- Bird, F.T.; Birk, J.M. Artificially disseminated virus as a factor controlling the European spruce sawfly, Diprion hercyniae (Htg.) in the absence of introduced parasites. Can. Entomol. 1961, 93, 228–238. [Google Scholar] [CrossRef]
- Stairs, G.R. Artificial initiation of virus epizootics in forest tent caterpillar populations. Can. Entomol. 1965, 97, 1059–1062. [Google Scholar] [CrossRef]
- Young, E.C. The epizootiology of two pathogens of the coconut palm rhinoceros beetle. J. Invertebr. Pathol. 1974, 24, 82–92. [Google Scholar] [CrossRef]
- Entwistle, P.F.; Adams, P.H.W.; Evans, H.F.; Rivers, C.F. Epizootology of a nuclear polyhedrosis virus (Baculoviridae) in European spruce sawfly (Gilpinia hercyniae): Spread of disease from small epicentres in comparison with spread of baculovirus diseases in other hosts. J. Appl. Ecol. 1983, 20, 473–487. [Google Scholar] [CrossRef]
- Shepherd, R.F.; Otvos, I.S.; Chorney, R.J.; Cunningham, J.C. Pest management of the Douglas-fir tussock moth (Lepidoptera: Lymantriidae): Prevention of an outbreak through early treatment with a nuclear polyhedrosis virus by ground and aerial applications. Can. Entomol. 1984, 116, 1533–1542. [Google Scholar] [CrossRef]
- Otvos, I.S.; Cunningham, J.C.; Friskie, L.M. Aerial application of nuclear polyhedrosis virus against Douglas-fir tussock moth, Orgyia pseudotsugata (McDunnough) (Lepidoptera: Lymantriidae). I. Impacts in the year of application. Can. Entomol. 1987, 119, 697–706. [Google Scholar] [CrossRef]
- Otvos, I.S.; Cunningham, J.C.; Alfaro, R.I. Aerial applications of nuclear polyhedrosis virus against Douglas-fir tussock moth, Orgyia pseudotsugata (McDunnough) (Lepidoptera: Lymantriidae). II. Impacts 1 and 2 years after application. Can. Entomol. 1987, 119, 707–715. [Google Scholar] [CrossRef]
- Fuxa, J.R.; Richter, A.R. Classical biological control in an ephemeral crop habitat with Anticarsia gemmatalis nucleopolyhedrovirus. BioControl 1999, 44, 403–419. [Google Scholar]
- Moreau, G.; Lucarotti, C.J.; Kettela, E.G.; Thurston, G.S.; Holmes, S.; Weaver, C.; Levin, D.B.; Morin, B. Aerial application of nucleopolyhedrovirus induces decline in increasing and peaking populations of Neodiprion abietis. Biol. Control 2005, 33, 65–73. [Google Scholar] [CrossRef]
- Moreau, G. Past and present outbreaks of the balsam fir sawfly: An analytical review. Forest Ecol. Manag. 2006, 221, 215–219. [Google Scholar] [CrossRef]
- Anderson, R.M.; May, R.M. The population dynamics of microparasites and their invertebrate hosts. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1981, 291, 451–524. [Google Scholar] [CrossRef]
- Wallace, D.R.; Cunningham, J.C. Diprionid sawflies. In Forest Insect Pests in Canada; Armstrong, J.A., Ives, W.G.H., Eds.; Natural Resources Canada: Ottawa, Canada, 1995; pp. 193–232. [Google Scholar]
- Linnen, C.R.; Farrell, B.D. Phylogenetic analysis of nuclear and mitochondrial genes reveals evolutionary relationships and mitochondrial introgression in the sertifer species group of the genus Neodiprion (Hymenoptera: Diprionidae). Mol. Phylogenet. Evol. 2008, 48, 240–257. [Google Scholar] [CrossRef]
- Carroll, W.J. Some aspects of the Neodiprion abietis complex in Newfoundland.
- Anstey, L.J.; Quiring, D.T.; Ostaff, D.P. Seasonal changes in intra-tree distribution of immature balsam fir sawfly (Hymenoptera: Diprionidae). Can. Entomol. 2002, 134, 529–538. [Google Scholar] [CrossRef]
- Parsons, K.H.; Piene, H.; Farrell, J.; Quiring, D.T. Temporal patterns of balsam fir sawfly defoliation and growth loss in young balsam fir. Forest Ecol. Manag. 2003, 184, 33–46. [Google Scholar] [CrossRef]
- Parsons, K.; Quiring, D.; Piene, H.; Moreau, G. Relationship between balsam fir sawfly density and defoliation in balsam fir. Forest Ecol. Manag. 2005, 205, 325–331. [Google Scholar] [CrossRef]
- Lucarotti, C.J.; Moreau, G.; Kettela, E.G. Abietiv™—A viral biopesticide for control of the balsam fir sawfly. In Biological Control: A Global Pespective; Vincent, C., Goettel, M., Lazarovits, G., Eds.; CABI International: Wallingford, United Kingdom, 2007; pp. 353–361. [Google Scholar]
- Lucarotti, C.J.; Morin, B.; Graham, R.I.; Lapointe, R. Production, application, and field performance of Abietiv™, the balsam fir sawfly nucleopolyhedrovirus. Virol. Sin. 2007, 22, 163–172. [Google Scholar] [CrossRef]
- Canadian Council of Forest Ministers. Forest Insect National Tables for Areas of Moderate to Severe Defoliation. Available online: http://nfdp.ccfm.org/insects/national_e.php/ (accessed on 27 July 2012).
- Lucarotti, C.J. Abietiv™. In Proceedings of the SERG International 2010 Workshop, St. John’s, NL, USA, 9–11 February 2010; pp. 81–87.
- Graves, R.; Quiring, D.T.; Lucarotti, C.J. Transmission of a Gammabaculovirus within cohorts of balsam fir sawfly (Neodiprion abietis) larvae. Insects 2012, in press.. [Google Scholar]
- Weisner, C.J. Review of the role of droplet size effects on spray efficacy. In Forest Insect Pests in Canada; Armstrong, J.A., Ives, W.G.H., Eds.; Natural Resources Canada: Ottawa, Canada, 1995; pp. 493–496. [Google Scholar]
- Payne, N.J. Spray dispersal, deposition, and assessment. In Forest Insect Pests in Canada; Armstrong, J.A., Ives, W.G.H., Eds.; Natural Resources Canada: Ottawa, Canada, 1995; pp. 465–478. [Google Scholar]
- Mickle, R.E.; Rousseau, G. Optimization Trials for Insecticide Spraying into Small Blocks—Murdochville Results; Spray Efficacy Research Group (SERG) Report: Quebec, Canada, 1998; p. 37. April 1998.
- Mierzejewski, K.; Reardon, R.C.; Thistle, H.; Dubois, N.R. Conventional application equipment: Aerial application. In Field Manual of Techniques in Invertebrate Pathology; Lacey, L.A., Kaya, H.K., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 99–126. [Google Scholar]
- Teshke, K.; Chow, Y.; Bartlett, K.; Ross, R.; van Netten, C. Spatial and temporal distribution of airborne Bacillus thuringiensis var. kurstaki during an aerial spray program for gypsy moth eradication. Environ. Health Perspect. 2001, 109, 47–54. [Google Scholar] [CrossRef]
- Yates, W.E.; Akesson, N.B.; Bayer, D.E. Drift of glyphosate sprays applied with aerial and ground equipment. Weed Sci. 1978, 26, 597–604. [Google Scholar]
- Renne, D.S.; Wolf, M.A. Experimental studies of 2,4-D herbicide drift characteristics. Agric. Meteorol. 1979, 20, 7–24. [Google Scholar] [CrossRef]
- Chester, G.; Ward, R.J. Occupational exposure and drift hazard during aerial application of paraquat to cotton. Arch. Environ. Contam. Toxicol. 1984, 13, 551–563. [Google Scholar] [CrossRef]
- Oeseburg, F.; van Leeuwen, D. Dispersion of aerial agricultural sprays; model and validation. Agric. Forest Meteorol. 1990, 53, 223–255. [Google Scholar] [CrossRef]
- Payne, N.J. Spray dispersal from aerial glyphosate applications. Crop Prot. 1993, 12, 463–469. [Google Scholar] [CrossRef]
- Woods, N.; Craig, I.P.; Dorr, G.; Young, B. Spray drift of pesticides arising from aerial application in cotton. J. Environ. Qual. 2001, 30, 697–701. [Google Scholar] [CrossRef] [Green Version]
- Moreau, G.; Eveleigh, E.S.; Lucarotti, C.J.; Quiring, D.T. Stage-specific responses to ecosystem alteration in an eruptive herbivorous insect. J. Appl. Ecol. 2006, 43, 28–34. [Google Scholar]
- Moreau, G.; Lucarotti, C.J. A brief review of the past use of baculoviruses for the management of eruptive forest defoliators and recent developments on a sawfly virus in Canada. For. Chron. 2007, 83, 105–112. [Google Scholar]
- Health Canada. Registration Decision Neodiprion abietis Newfoundland Strain. RD-2009-05; Health Canada Pest Management Regulatory Agency: Ottawa, Canada, 2009 March 25; p. 9. [Google Scholar]
- McLeod, I.M.; Lucarotti, C.J.; Hennigar, C.R.; MacLean, D.A.; Holloway, A.G.L.; Cormier, G.A.; Davies, D.C. Advances in aerial application technologies and decision support for integrated pest management. In Integrated Pest Management and Pest Control; Soloneski, S., Larramendy, M.L., Eds.; InTech Open Access Publisher: Rijeka, Croatia, 2012; pp. 651–668. [Google Scholar]
- Moreau, G.; Eveleigh, E.S.; Lucarotti, C.J.; Quiring, D.T. Ecosystem alteration modifies the relative strengths of bottom-up and top-down forces in a herbivore population. J. Anim. Ecol. 2006, 75, 853–861. [Google Scholar] [CrossRef]
- Campbell, C.S.; Quiring, D.T.; Kettela, E.G.; Lucarotti, C.J. Application of Balsam Fir Sawfly Nucleopolyhedrovirus against Its Natural Host Neodiprion abietis (Hymenoptera: Diprionidae). In Proceedings of the IUFRO Workshop on Forest Insect Population Dynamics and Host Influences, Kanazawa, Japan, 14–19 September 2003; pp. 86–89.
- Lucarotti, C.J.; Whittome-Waygood, B.H.; Lapointe, R.; Levin, D.B. Pathology of a Gammabaculovirus in its natural balsam fir sawfly (Neodiprion abietis) host. Psyche 2012, in press.. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Graves, R.; Lucarotti, C.J.; Quiring, D.T. Spread of a Gammabaculovirus within Larval Populations of Its Natural Balsam Fir Sawfly (Neodiprion abietis) Host Following Its Aerial Application. Insects 2012, 3, 912-929. https://doi.org/10.3390/insects3040912
Graves R, Lucarotti CJ, Quiring DT. Spread of a Gammabaculovirus within Larval Populations of Its Natural Balsam Fir Sawfly (Neodiprion abietis) Host Following Its Aerial Application. Insects. 2012; 3(4):912-929. https://doi.org/10.3390/insects3040912
Chicago/Turabian StyleGraves, Roger, Christopher J. Lucarotti, and Dan T. Quiring. 2012. "Spread of a Gammabaculovirus within Larval Populations of Its Natural Balsam Fir Sawfly (Neodiprion abietis) Host Following Its Aerial Application" Insects 3, no. 4: 912-929. https://doi.org/10.3390/insects3040912