Update on the Status of Bemisia tabaci in the UK and the Use of Entomopathogenic Fungi within Eradication Programmes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

2. Pest Status of Bemisia tabaci
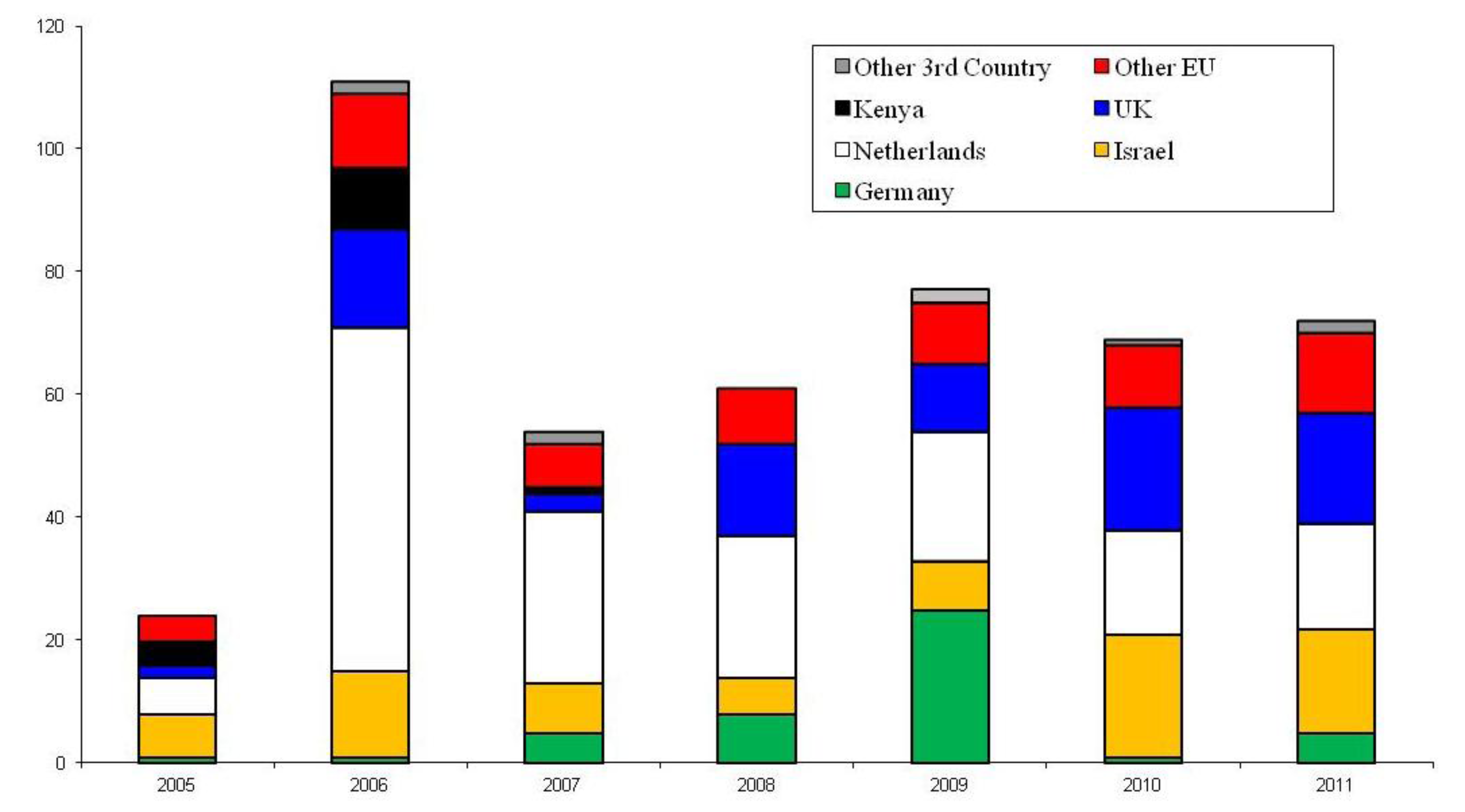
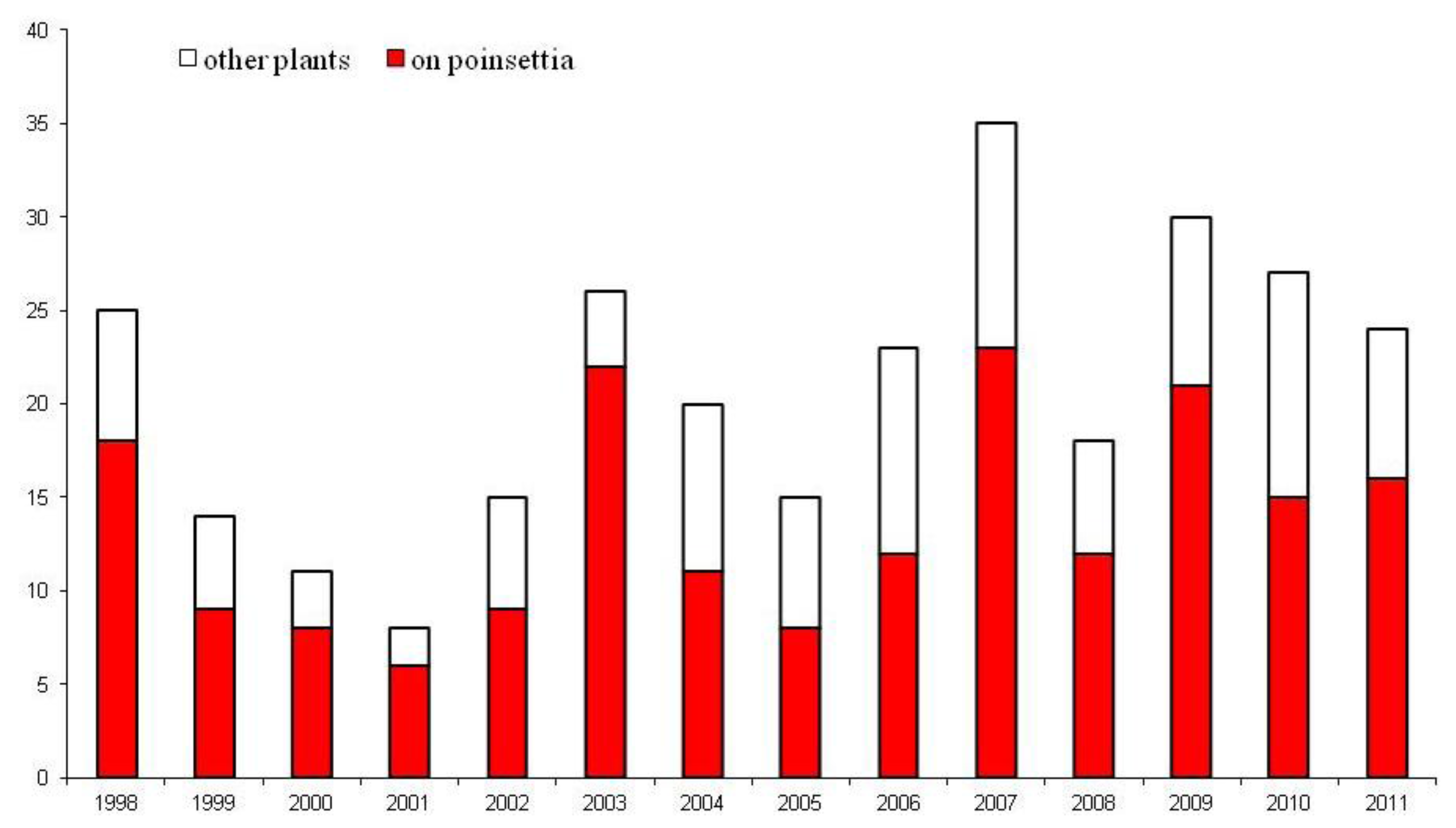
3. UK Interceptions of Bemisia tabaci
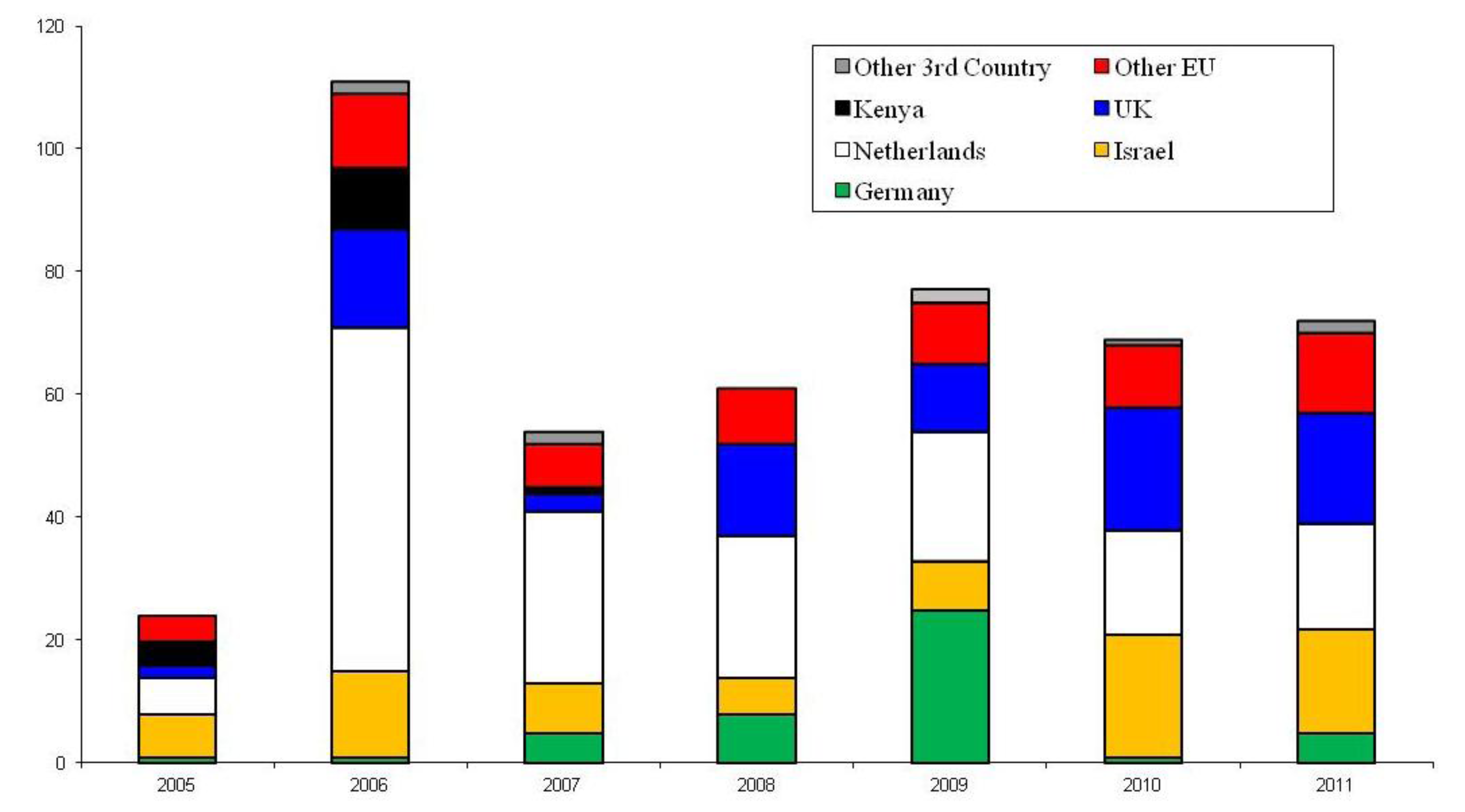
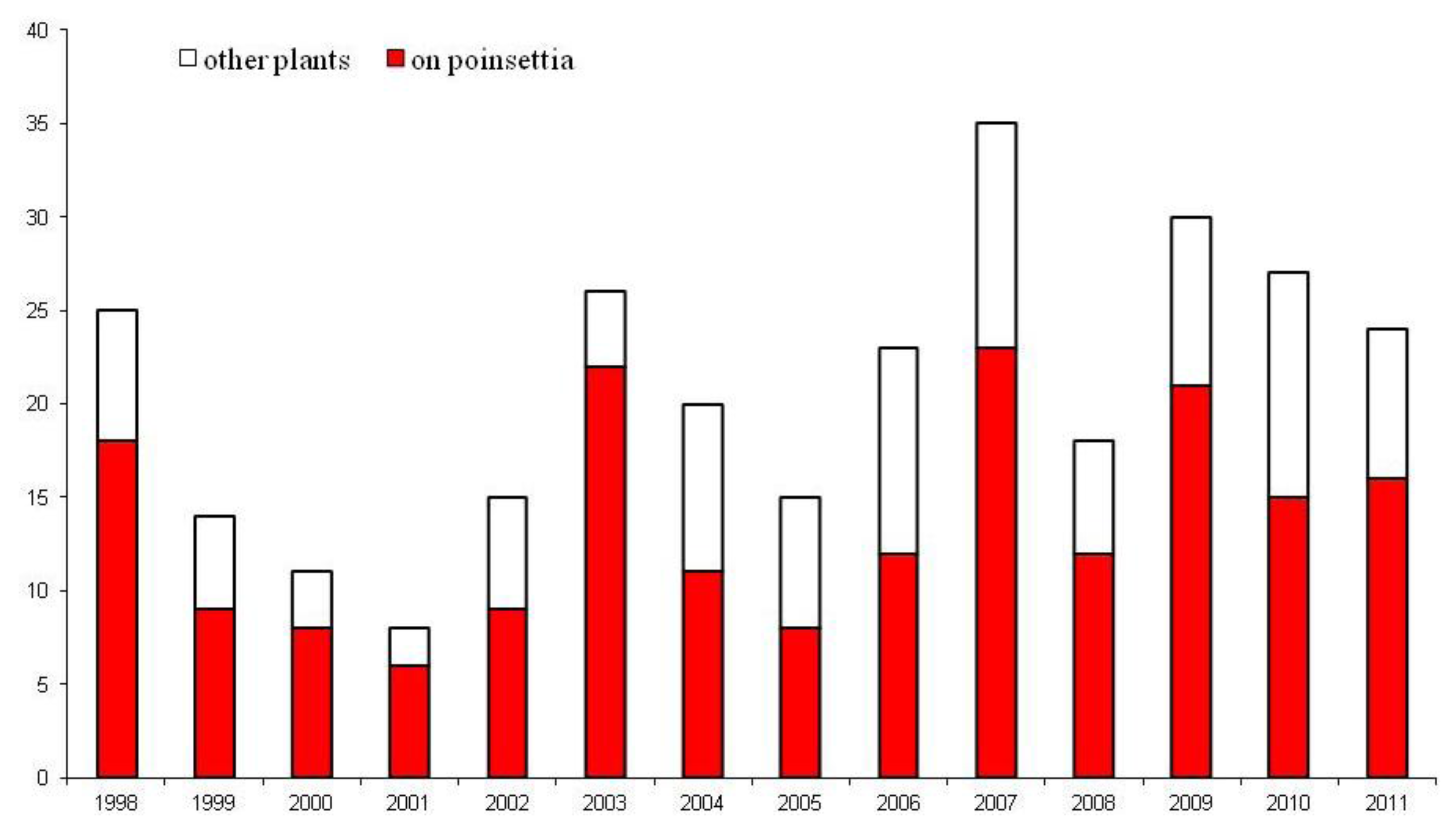
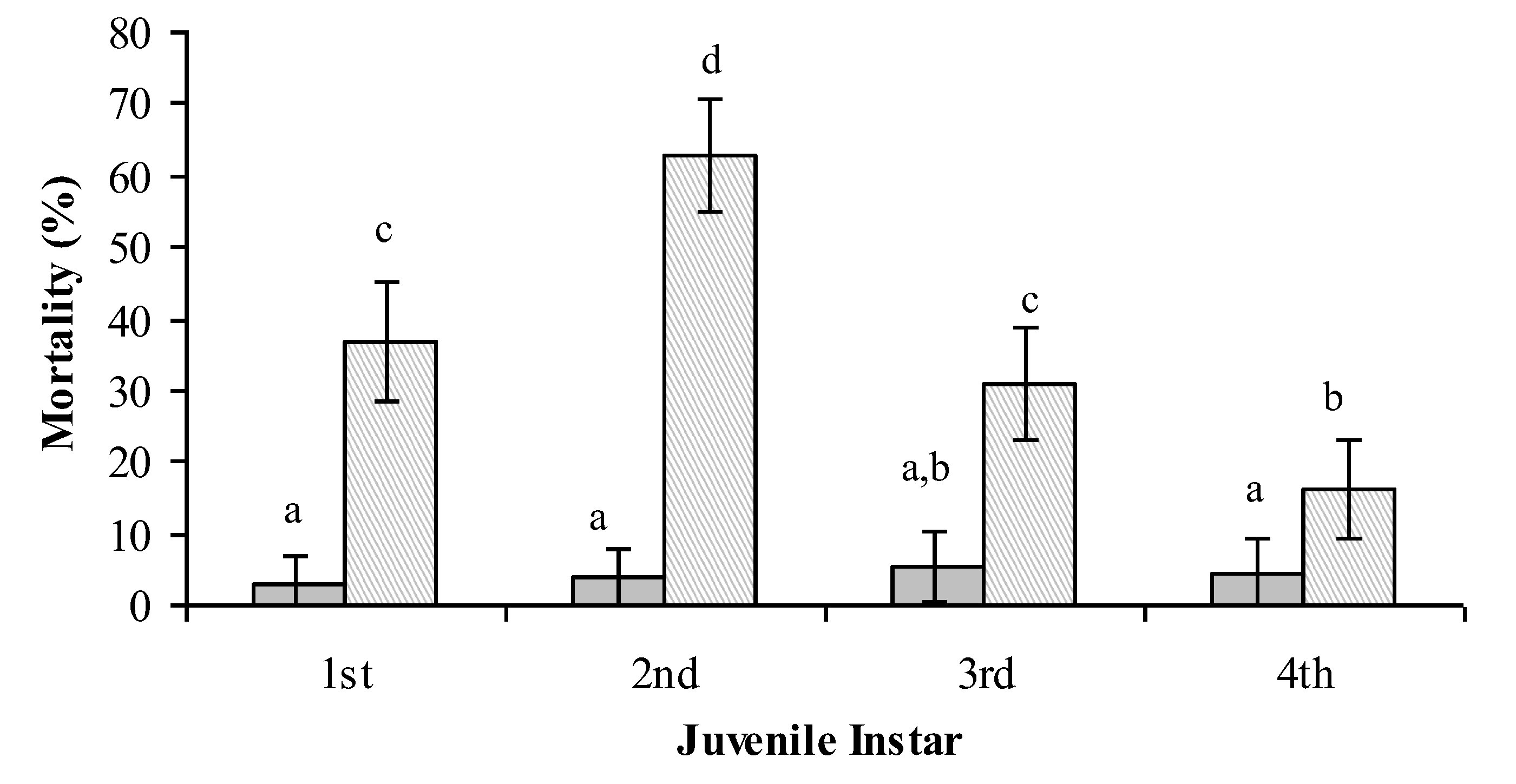
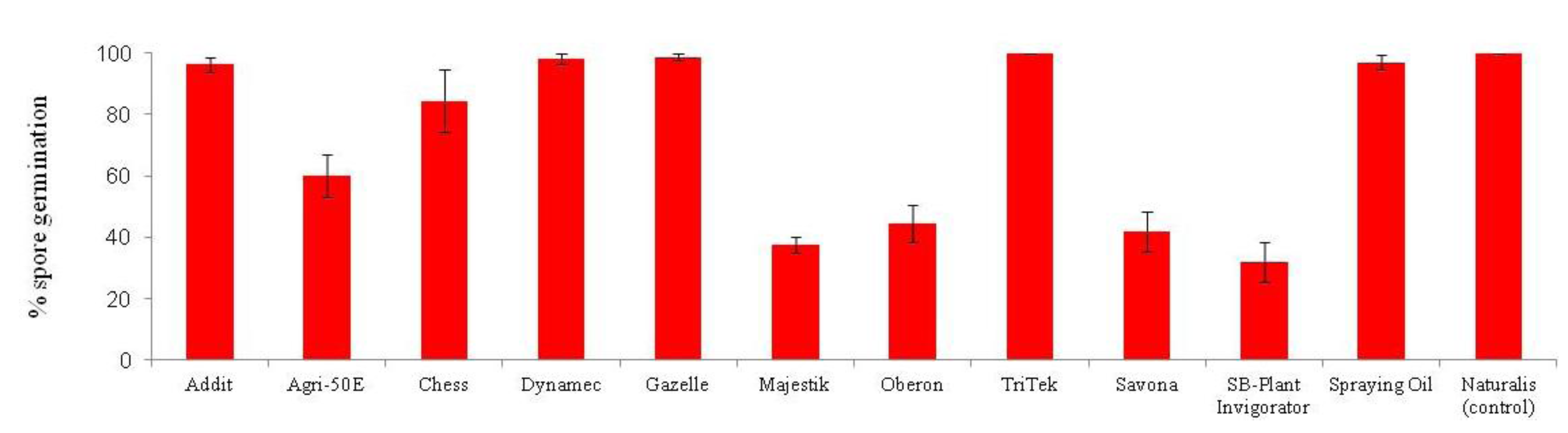
4. Entomopathogenic Fungi for the Control of Bemisia tabaci

 Lecanicillium muscarium + 0.02% Agral
Lecanicillium muscarium + 0.02% Agral  [26].
Lecanicillium muscarium + 0.02% Agral [26].
[26].
Lecanicillium muscarium + 0.02% Agral [26].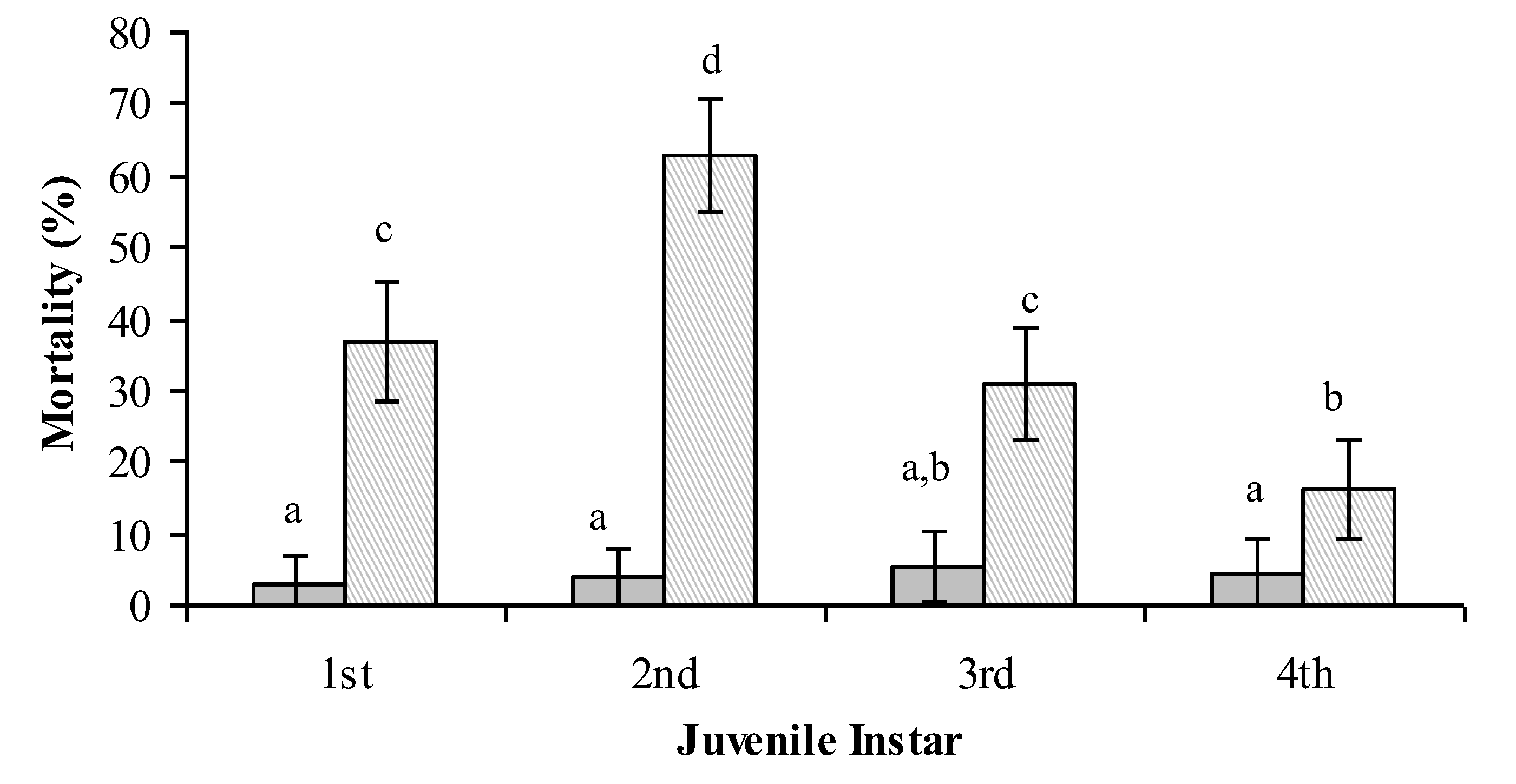
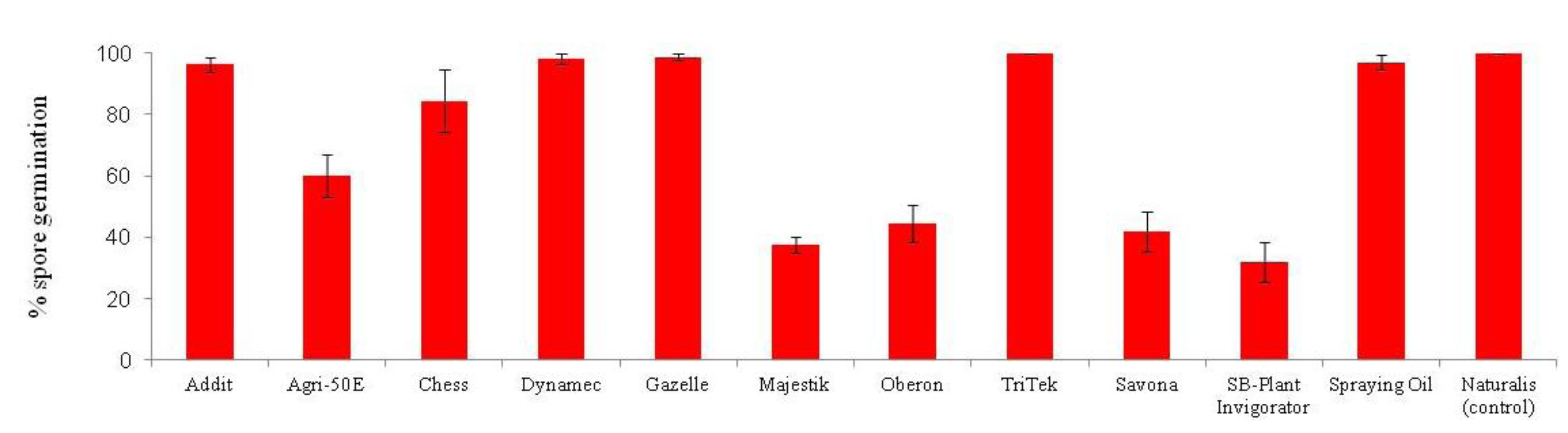
5. Conclusions
Acknowledgements
References
- Xu, C.; Qiu, B.-L.; Cuthbertson, A.G.S.; Zhang, Y.; Ren, S.-X. Adaptability of sweetpotato whitefly Bemisia tabaci (Hemipetera: Aleyrodidae) on seven marginal host plants. Int. J. Pest Manage. 2012, 58, 297–301. [Google Scholar] [CrossRef]
- Powell, M.E.; Cuthbertson, A.G.S.; Boonham, N.; Morris, J.; Bell, H.A.; Northing, P. First record of the Q Biotype of the sweetpotato whitefly, Bemisia tabaci, intercepted in the U. Eur. J. Plant Pathol. 2012, 133, 797–801. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S. Protecting the UK from alien pests. Bio. News 2005, 32, 9. [Google Scholar]
- Cuthbertson, A.G.S.; Blackburn, L.F.; Eyre, D.P.; Cannon, R.J.C.; Millar, J.; Northing, P. Bemisia tabaci: The current situation in the UK and the prospect of developing strategies for eradication using entomopathogens. Insect Sci. 2011, 18, 1–10. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; Powell, M.E. Bemisia tabaci—A cryptic species complex. Bio. News 2012, 57, 17–18. [Google Scholar]
- Powell, M.E.; Cuthbertson, A.G.S. Pest control: Distinguishing between different biotypes of Bemisia tabaci in the UK. The Biologist 2013, 60, 18–21. [Google Scholar]
- De Barro, P.J.; Liu, S.S.; Boykin, L.M.; Dinsdale, A.B. Bemisia tabaci: A statement of species status. Ann. Rev. Entomol. 2011, 56, 1–19. [Google Scholar] [CrossRef]
- Dinsdale, A.; Cook, L.; Riginos, C.; Buckley, Y.M.; De Barro, P. Refined global analysis of Bemisia tabaci (Hemiptera: Sternorrhyncha: Aleyrodoidea: Aleyrodidae) mitochondrial cytochrome oxidase 1 to identify species level genetic boundaries. Ann. Entomol. Soc. Am. 2010, 103, 196–208. [Google Scholar] [CrossRef]
- Shatters, R.G.; Powell, C.A.; Boykin, L.; Liansheng, H.; McKenzie, C.L. Improved DNA barcoding method for Bemisia tabaci and related Aleyrodidae: Development of universal and Bemisia tabaci biotype-specific mitochondrial cytochrome coxidase I polymerase chain reaction primers. J. Econ. Entomol. 2009, 102, 750–758. [Google Scholar] [CrossRef]
- Bethke, J.A.; Byrne, F.J.; Hodges, G.S.; McKenzie, C.L.; Shatters, R.G. First record of the Q biotype of the sweetpotato whitefly, Bemisia tabaci, in Guatemal. Phytoparasitica 2009, 37, 61–64. [Google Scholar] [CrossRef]
- Jones, C.M.; Gorman, K.; Denholm, I.; Williamson, M.S. High-throughput allelic discrimination of B and Q biotypes of the whitefly, Bemisia tabaci, using TaqMan allele-selective PCR. Pest Manag. Sci. 2008, 64, 12–15. [Google Scholar] [CrossRef]
- McKenzie, C.I.; Hodges, G.; Osborne, L.; Byrne, F.J.; Shatters, R.G. Distribution of Bemisia tabaci (Hemiptera: Aleyrodidae) biotypes in Florida—investigating the Q Invasion. J. Econ. Entomol. 2009, 102, 670–676. [Google Scholar] [CrossRef]
- Cannon, R.J.C.; Eyre, D.; MacLeod, A.; Matthews, L.; Malumphy, C.; Cheek, S.; Bartlett, P.W. Interceptions and outbreaks of Bemisia tabaci in the UK. In The BCPC International Congress—Crop Science and Technology; The British Crop Protection Council: Alton, UK, 2005; Volume 2, pp. 1007–1012. [Google Scholar]
- Ahmad, M.; Arif, M.I.; Ahmad, Z.; Denholm, I. Cotton whitefly (Bemisia tabaci) resistance to organophosphate and pyrethroid insecticides in Pakistan. Pest Manag. Sci. 2002, 58, 203–208. [Google Scholar] [CrossRef]
- Byrne, F.J.; Oletting, R.D.; Bethke, J.A.; Green, C.; Chamberlin, J. Understanding the dynamics of neonicotinoid activity in the management of Bemisia tabaci whiteflies on poinsettias. Crop Prot. 2010, 29, 260–266. [Google Scholar] [CrossRef]
- Luo, C.; Jones, C.M.; Devine, G.; Zhang, F.; Denholm, I.; Gorman, K. Insecticide resistance in Bemisia tabaci biotype Q (Hemiptera: Aleyrodidae) from China. Crop Prot. 2010, 29, 429–434. [Google Scholar] [CrossRef]
- Schuster, D.J.; Mann, R.S.; Toapanta, M.; Cordero, R.; Thompson, S.; Cyman, S.; Shurtleff, A.; Morris, R.F. Monitoring neonicotinoid resistance in biotype B of Bemisia tabaci in Florida. Pest Manag. Sci. 2010, 66, 186–195. [Google Scholar]
- Wang, Z.; Yan, H.; Yang, Y.; Wu, Y. Biotype and insecticide resistance status of the whitefly Bemisia tabaci in China. Pest Manag. Sci. 2010, 66, 1360–1366. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; Buxton, J.H.; Blackburn, L.F.; Mathers, J.J.; Robinson, K.; Powell, M.E.; Fleming, D.A.; Bell, H.A. Eradicating Bemisia tabaci Q on poinsettia plants in the UK. Crop Prot. 2012, 42, 42–48. [Google Scholar] [CrossRef]
- Lacey, L.A.; Fransen, J.J.; Carruthers, R.I. Global distribution of naturally occurring fungi of Bemisia, their biologies and use as biological control agents. In Bemisia: 1995 Taxonomy, Biology, Damage, Control and Management; Gerling, D., Mayer, R.T., Eds.; Intercept Ltd: Andover, UK, 1996; pp. 401–433. [Google Scholar]
- Wraight, S.P.; Carruthers, R.I.; Bradley, C.A.; Jaronski, S.T.; Lacey, L.A.; Wood, P.; Galaini-Wraight, S. Pathogenicity of the entomopathogenic fungi Paecilomyces spp. and Beauveria bassiana against the silverleaf whitefly, Bemisia argentifolii. J. Invertebr. Pathol. 1988, 71, 217–226. [Google Scholar]
- Wraight, S.P.; Carruthers, R.I.; Jaronski, S.T.; Bradley, C.A.; Garza, C.J.; Galaini-Wraight, S. Evaluation of the entomopathogenic fungi Beauveria bassiana and Paecilomyces fumosoroseus for microbial control of the silverleaf whitefly, Bemisia argentifolii. Biol. Control 2000, 17, 203–217. [Google Scholar] [CrossRef]
- Fransen, J.J. Natural enemies of whiteflies: Fungi. In Whiteflies: Their Bionomics, Pest Status and Management; Gerling, D., Ed.; Intercept Ltd: Andover, UK, 1990; pp. 187–210. [Google Scholar]
- Steenberg, T.; Humber, R. Entomopathogenic potential of Verticillium lecanii and Acremonium species (Deuteromycotina: Hyphomycetes). J. Invertebr. Pathol. 1999, 73, 309–314. [Google Scholar] [CrossRef]
- Poprawski, T.J.; Greenberg, S.M.; Ciomperlik, M.A. Effect of host plant on Beauveria bassiana and Paecilomyces fumosoroseus induced mortality of Trialeurodes vaporariorum (Homoptera: Aleyrodidae). Environ. Entomol. 2000, 29, 1048–1053. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; Walters, K.F.A.; Northing, P. Susceptibility of Bemisia tabaci immature stages to the entomopathogenic fungus Lecanicillium muscarium on tomato and verbena foliage. Mycopathologia 2005, 159, 23–29. [Google Scholar] [CrossRef]
- Meade, D.L.; Byrne, D.N. The use of Verticillium lecanii against subimaginal instars of Bemisia tabaci. J. Invertebr. Pathol. 1991, 57, 296–298. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; Walters, K.F.A.; Deppe, C. Compatibility of the entomopathogenic fungus Lecanicillium muscarium and insecticides for eradication of sweetpotato whitefly, Bemisia tabaci. Mycopathologia 2005, 160, 35–41. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; Blackburn, L.F.; Northing, P.; Luo, W.; Cannon, R.J.C.; Walters, K.F.A. Further chemical compatibility testing of the entomopathogenic fungus Lecaniillium muscarium to control Bemisia tabaci in glasshouses. Int. J. Environ. Sci. Tech. 2010, 7, 405–409. [Google Scholar]
- Cuthbertson, A.G.S.; Walters, K.F.A. Pathogenicity of the entomopathogenic fungus Lecanicillium muscarium against the sweetpotato whitefly Bemisia tabaci under laboratory and glasshouse conditions. Mycopathologia 2005, 160, 315–319. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cuthbertson, A.G.S. Update on the Status of Bemisia tabaci in the UK and the Use of Entomopathogenic Fungi within Eradication Programmes. Insects 2013, 4, 198-205. https://doi.org/10.3390/insects4020198
Cuthbertson AGS. Update on the Status of Bemisia tabaci in the UK and the Use of Entomopathogenic Fungi within Eradication Programmes. Insects. 2013; 4(2):198-205. https://doi.org/10.3390/insects4020198
Chicago/Turabian StyleCuthbertson, Andrew G. S. 2013. "Update on the Status of Bemisia tabaci in the UK and the Use of Entomopathogenic Fungi within Eradication Programmes" Insects 4, no. 2: 198-205. https://doi.org/10.3390/insects4020198