Ecology, Behaviour and Control of Apis cerana with a Focus on Relevance to the Australian Incursion

Abstract
:1. Introduction
- What is known about the ecology and behaviour of Indo-Malayan Apis cerana? How are tropical and temperate strains different?
- What is known about honey production and pollination services in A. cerana? How does this differ between tropical and temperate strains?
- How does A. cerana ecology and behaviour compare to Apis mellifera, and what is the likely ecological overlap between these two species where they co-occur?
- What has been the impact of A. cerana where it has been introduced? How is A. cerana controlled and managed where it has been introduced?
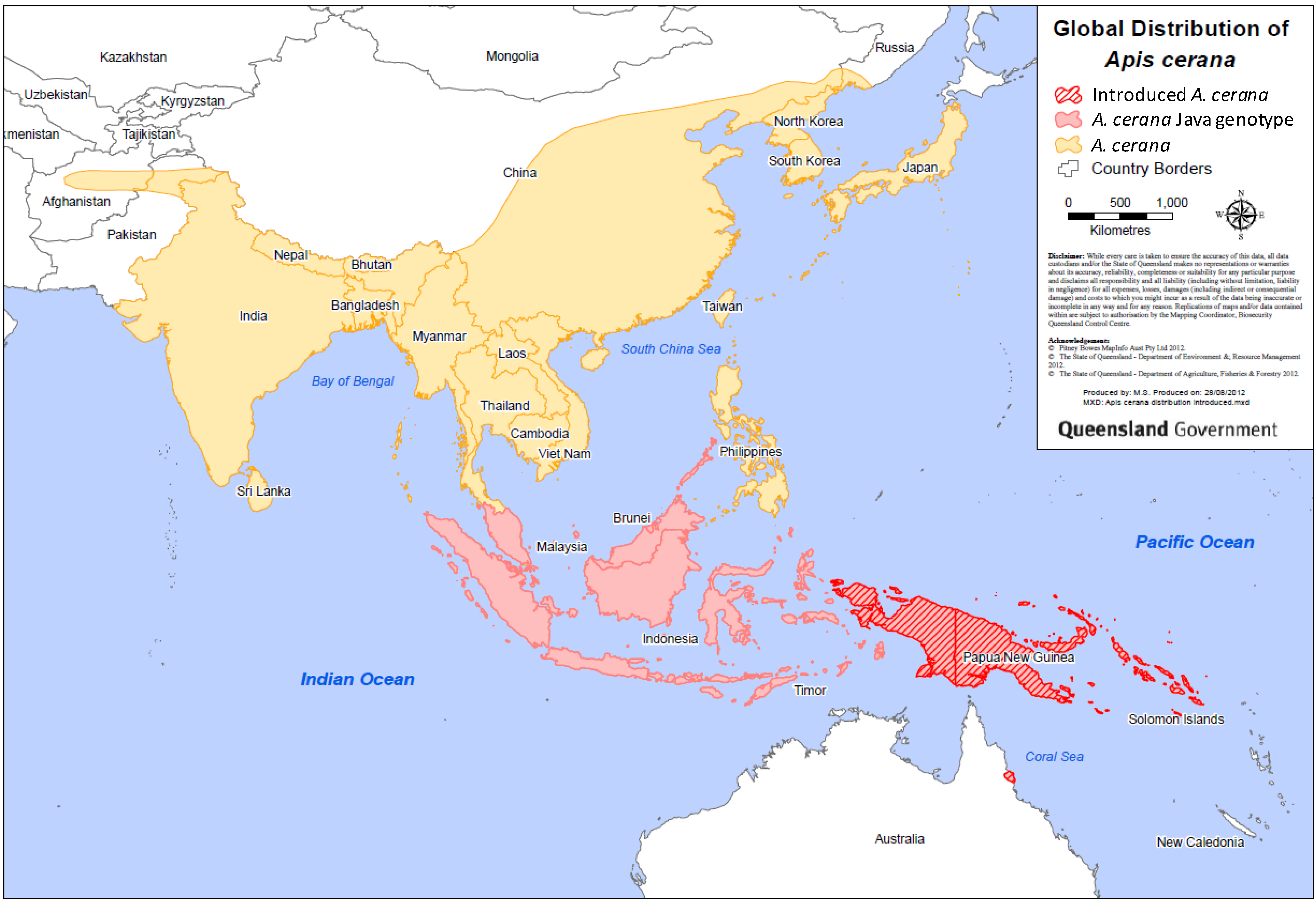
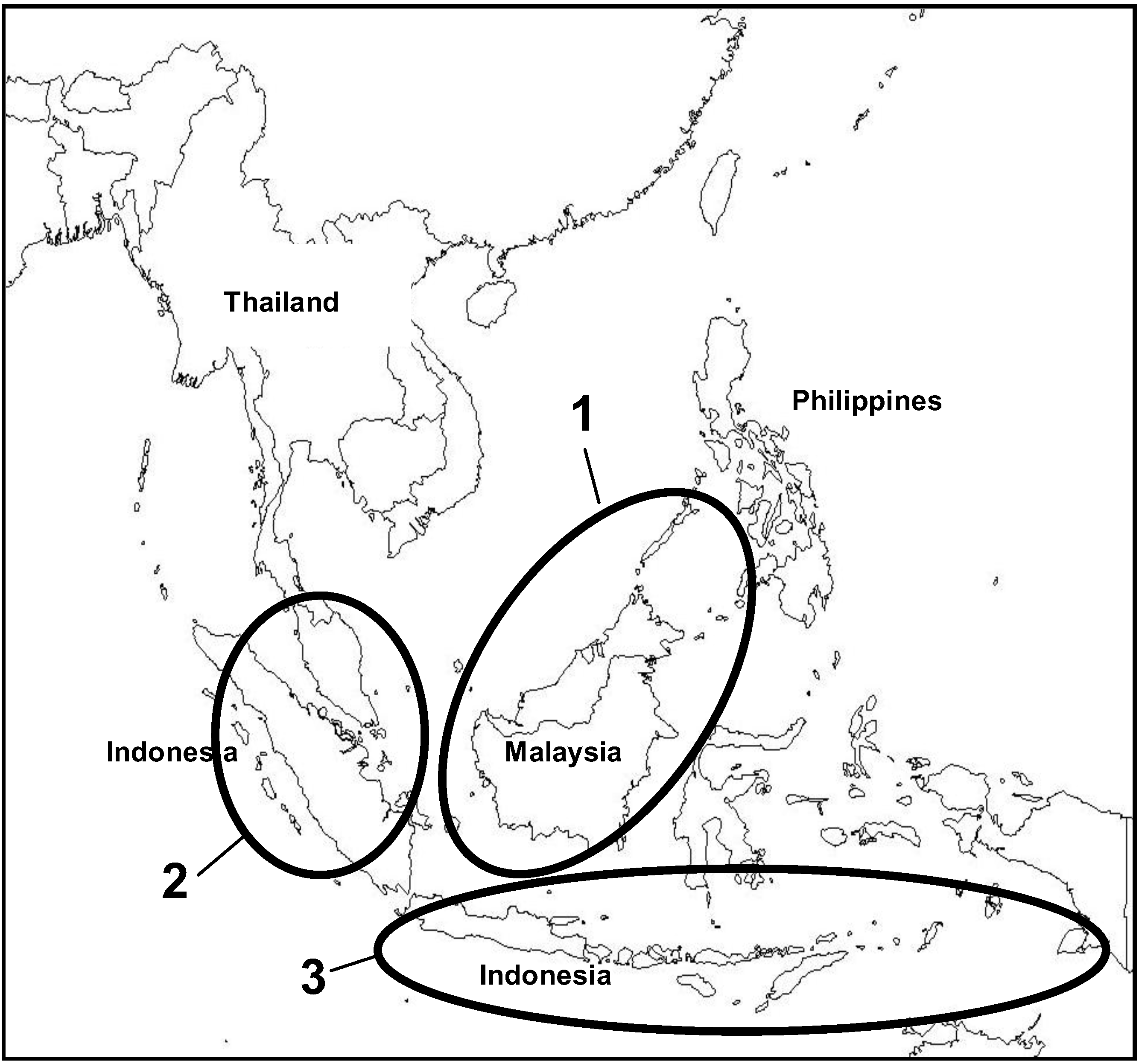
2. Distribution
Natural and Introduced Range

3. Morphology and Genetic Diversity
3.1. Appearance
3.2. Morphological Divergence

3.3. Genetic Divergence

4. Ecology and Behaviour
Foraging
5. Nesting
5.1. Nesting Habitats and Sites
5.2. Nest Characteristics
5.3. Colony Size and Nest Density
6. Mating and Reproduction
6.1. Timing of Mating Flights, Sex Attractants, and Drone Congregation Areas
6.2. Brood Development

{kind=link}
{kind=link}
{kind=link}
| Stage | Worker | Drone | Queen | |||
|---|---|---|---|---|---|---|
| A. cerana | A. mellifera | A. cerana | A. mellifera | A. cerana | A. mellifera | |
| Egg to larva | 3 | 3 | 3 | 3 | 3 | 3 |
| Larva to pupa | 5 | 6 | 6 | 7 | 4–5.5 | 5 |
| Pupa to adult | 11 | 12 | 14 | 14 | 6–7.5 | 5 |
| Total | 19 | 21 | 23 | 24 | 13–16 | 13 |
7. Swarming and Absconding
7.1. Seasonal Absconding
7.2. Predation (Disturbance Absconding)
7.3. Reproductive Swarming
8. Other Behaviour
8.1. Temperament
8.2. Diseases and Hygiene
8.3. Fanning Behaviour
9. Pollination
9.1. Pollination Services—Crops
9.2. Pollination Services—Wild Flora
10. A. cerana vs. A. mellifera
10.1. Competition between A. cerana and A. mellifera
10.2. Competition for Floral Resources
10.3. Robbing and Direct Fighting
10.4. Mating Interference
11. Examples where A. cerana and A. mellifera Co-Exist (through Human Introduction)
11.1. Solomon Islands
11.2. Asia
11.3. Far North Queensland, Australia
12. Control Strategies
12.1. Australia
12.2. Solomon Islands
13. Detection and Capturing Techniques
13.1. Detection
13.2. Swarm Capture
13.3. Trap Attractants—Scents
13.4. Trap Attractants—Pheromones
13.4.1. Pheromones in A. cerana
13.4.2. Attractiveness of Pheromones
14. Conclusions
Acknowledgments
Conflicts of Interest
References
- Ruttner, F. Biogeography and Taxonomy of Honeybees; Springer-Verlag Berlin: Heidelberg, Germany, 1988. [Google Scholar]
- Anderson, D. Control of Asian honeybees in the Solomon Islands; Australian Centre for International Agricultural Research (ACIAR): Canberra, Australia, 2010.
- Anderson, D.L.; Annand, N.; Lacey, M.; Ete, S. Control of Asian Honey Bees in Solomon Islands; Australian Centre for International Agricultural Research (ACIAR): Canberra, Australia, 2012.
- Anderson, D.L. Assessment of the Varroa Mite and Asian Bee Incursion in the Solomon Islands; Australian Centre for International Agricultural Research (ACIAR): Canberra, Australia, 2004.
- Anderson, D.; CSIRO, Canberra, Australia. Unpublished work. 2013.
- Hepburn, C.; Radloff, S.E. Honeybees of Asia; Springer-Verlag Berlin: Heidelberg, Germany, 2011. [Google Scholar]
- Crane, E. The World History of Beekeeping and Honey Hunting; Duckworth: London, UK, 1999. [Google Scholar]
- Bradbear, N. Bees and Their Role in Forest Livelihoods a Guide to the Services Provided by Bees and the Sustainable Harvesting, Processing and Marketing of Their Products; Food and Agriculture Organization of the United Nations: Rome, Italy, 2009.
- Anderson, D.L. Non-Reproduction of Varroa jacobsoni in Apis mellifera colonies in Papua New Guinea and Indonesia. Apidologie 1994, 25, 412–421. [Google Scholar] [CrossRef]
- Dunn, K.J. Exotic Asian bee detected in Torres Strait. Bee Briefs 1992, 9, 18–19. [Google Scholar]
- Barry, S.; Cook, D.; Duthie, R.; Clifford, D.; Anderson, D. Future surveillance needs for honeybee biosecurity; Rural Industries Research and Development Corporation: Canberra, Australia, 2010. [Google Scholar]
- Shield, J. The Asian Honey Bee: Report of an Incursion in Cairns 2007—Technical Aspects of the Response; Department of Primary Industries and Fisheries: Brisbane, Australia, 2007; pp. 1–106.
- Radloff, S.E.; Hepburn, C.; Hepburn, H.R.; Fuchs, S.; Hadisoesilo, S.; Tan, K.; Engel, M.S.; Kuznetsov, V. Population structure and classification of Apis cerana. Apidologie 2010, 41, 589–601. [Google Scholar] [CrossRef]
- Smith, D.R.; Villafuerte, L.; Otis, G.; Palmer, M.R. Biogeography of Apis cerana F. and A-nigrocincta Smith: Insights from mtDNA studies. Apidologie 2000, 31, 265–279. [Google Scholar] [CrossRef]
- Oldroyd, B.P.; Wongsiri, S. Asian Honey Bees: Biology, Conservation and Human Interactions; Harvard University Press: Cambridge, MA, USA, 2006. [Google Scholar]
- Ken, T.; Fuchs, S.; Koeniger, N.; Zan, R.G. Morphological characterization of Apis cerana in the Yunnan Province of China. Apidologie 2003, 34, 553–561. [Google Scholar] [CrossRef]
- Verma, L.R.; Mattu, V.K.; Daly, H.V. Multivariate morphometrics of the Indian honeybee inthe Northwest Himalayan region. Apidologie 1994, 25, 203–223. [Google Scholar] [CrossRef]
- Koetz, A.H.; Biosecurity Queensland, Department of Agriculture, Fisheries and Forestry (DAFF), Cairns, Australia. Unpublished work. 2013.
- Radloff, S.E.; Hepburn, H.R.; Hepburn, C.; Fuchs, S.; Otis, G.W.; Sein, M.M.; Aung, H.L.; Pham, H.T.; Tam, D.Q.; Nuru, A.M.; et al. Multivariate morphometric analysis of Apis cerana of southern mainland Asia. Apidologie 2005, 36, 127–139. [Google Scholar] [CrossRef]
- Smith, D.R. Asian Honeybees and Mitochondrial DNA. In Honeybees of Asia; Hepburn, H.R., Radloff, S.E., Eds.; Springer-Verlag Berlin: Heidelberg, Germany, 2011; pp. 69–93. [Google Scholar]
- Rueppell, O.; Hayes, A.M.; Warrit, N.; Smith, D.R. Population structure of Apis cerana in Thailand reflects biogeography and current gene flow rather than Varroa mite association. Insectes Soc. 2011, 58, 445–452. [Google Scholar] [CrossRef]
- Songram, O.; Sittipraneed, S.; Klinbunga, S. Mitochondrial DNA diversity and genetic differentiation of the honeybee (Apis cerana) in Thailand. Biochem. Genet. 2006, 44, 256–269. [Google Scholar] [CrossRef]
- Oakey, J. May 2012: Short Study of Microsatellite Alleles in Asian Honey Bees Sourced from PNG/Solomons and North Queensland: Summary of Data Interpretation; Biosecurity Queensland, Department of Agriculture, Fisheries and Forestry: Brisbane, Australia, 2012.
- Winston, M.L. The Biology of the Honey Bee; Harvard University Press: Cambridge, MA, USA, 1987. [Google Scholar]
- Bakker, D.R. Foraging and Habitat Selection by Two Species of Honey Bee near Lore Lindu National Park in Sulawesi, Indonesia; The University of Guelph: Guelph, Canada, 1999. [Google Scholar]
- Corlett, R.T. Honeybees in Natural Ecosystems. In Honeybees of Asia; Hepburn, H.R., Radloff, S.E., Eds.; Springer-Verlag Berlin: Heidelberg, Germany, 2011; pp. 215–226. [Google Scholar]
- Partap, U. The Pollination Role of Honeybees. In Honeybees of Asia; Hepburn, H.R., Radloff, S.E., Eds.; Springer-Verlag Berlin: Heidelberg, Germany, 2011; pp. 227–255. [Google Scholar]
- Dyer, F.C.; Seeley, T.D. Dance dialects and foraging range in three Asian honey bee species. Behav. Ecol. Sociobiol. 1991, 28, 227–233. [Google Scholar]
- Hyatt, S. Asian Honey Bee (Apis cerana javana) in Cairns, Far North Queensland: Foraging, Nesting and Swarming Behaviour—Report of Field Observations April 2007–September 2011; Department of Employment, Economic Development and Innovation: Cairns, Queensland, Australia, 2011; pp. 1–24.
- Bhuiyan, M.K.H.; Hossain, M.M.; Bari, M.N. Rearing and management of Apis cerana (F.) and occurence of pests in honeybee colonies. OnLine J. Biol. Sci. 2002, 2, 14–17. [Google Scholar] [CrossRef]
- Dhaliwai, H.S.; Sharma, P.L. Foraging range of the Indian honeybee. J. Apic Res. 1974, 13, 137–141. [Google Scholar]
- Kevan, P.G.; Punchihewa, R.W.K.; Greco, C.F. Foraging Range for Apis cerana and Its Implications for Honey Production and Apiary Management. In The Asiatic Hive Bee: Apiculture, Biology, and Role in Sustainable Development in Tropical and Subtropical Asia; Kevan, P.G., Ed.; Enviroquest Ltd.: Cambridge, ON, Canada, 1995; pp. 223–228. [Google Scholar]
- Hisashi, F. Profitable beekeeping with Apis cerana. Bees Dev. J. 2010, 94, 8–10. [Google Scholar]
- Visscher, P.K.; Seeley, T.D. Foraging strategy of honeybee colonies in a temperate deciduous forest. Ecology 1982, 63, 1790–1801. [Google Scholar] [CrossRef]
- Beekman, M.; Ratnieks, F.L.W. Long-Range foraging by the honey-bee, Apis mellifera L. Funct. Ecol. 2000, 14, 490–496. [Google Scholar] [CrossRef]
- Abrol, D.P. Foraging. In Honeybees of Asia; Hepburn, H.R., Radloff, S.E., Eds.; Springer-Verlag Berlin: Heidelberg, Germany, 2011; pp. 257–292. [Google Scholar]
- Abrol, D.P. Diversity of pollinating insects visiting litchi flowers (Litchi chinensis Sonn.) and path analysis of environmental factors influencing foraging behaviour of four honeybee species. J. Apic Res. 2006, 45, 180–187. [Google Scholar] [CrossRef]
- Verma, L.R.; Dulta, P.C. Foraging behaviour of Apis cerana indica and Apis mellifera in pollinating apple flowers. J. Apic Res. 1986, 25, 197–201. [Google Scholar]
- Singh, M.M. Forgaging behaviour of the Himalayan honeybee (Apis cerana F.) on flowers of Fagopyrum esculentum M. and its impact on grain quality and yield. Ecoprint 2008, 15, 37–46. [Google Scholar]
- Seeley, T.D.; Seeley, R.H.; Akratanakul, P. Colony defense strategies of the Honeybees in Thailand. Ecol. Monogr. 1982, 52, 43–63. [Google Scholar] [CrossRef]
- Corlett, R.T. Flower visitors and pollination in the Oriental (Indomalayan) Region. Biol. Rev. 2004, 79, 497–532. [Google Scholar] [CrossRef]
- Phiancharoen, M.; Duangphakdee, O.; Hepburn, H.R. Biology of Nesting. In Honeybees of Asia; Hepburn, H.R., Radloff, S.E., Eds.; Springer-Verlag Berlin: Heidelberg, Germany, 2011; pp. 109–131. [Google Scholar]
- Otis, G.W. Distributions of recently recognized species of honey bees (Hymenoptera: Apidae; Apis) in Asia. J. Kans Entomol. Soc. 1996, 69, 311–333. [Google Scholar]
- Hadisoesilo, S. A Comparative Study of Two Species of Cavity-Nesting Honey Bees of Sulawesi, Indonesia; University of Guelph: Guelph, Canada, 1997. [Google Scholar]
- Inoue, T.; Adri, S.; Salmah, S. Nest Site Selection and Reproductive Ecology of the Asian Honey Bee, Apis cerana indica, in Central Sumatra. In Natural History of Social Wasps and Bees in Equatorial Sumatra; Sakagami, S.F., Ohgushi, R.I., Roubik, D.W., Eds.; Hokkaido University Press: New York, NY, USA, 1990; pp. 219–232. [Google Scholar]
- Karlsson, T. The natural nest of the Asian hive bee (Apis cerana) in Bangladesh—A minor field study. Swed. Univ. Agric. Sci. 1990, 134, 32. [Google Scholar]
- Ruttner, F.; Woyke, J.; Koeniger, N. Reproduction in Apis cerana 1. Mating behaviour. J. Apic. Res. 1972, 11, 141–146. [Google Scholar]
- Muzaffar, N.; Ahmad, R. Apis spp. (Hymenoptera, Apidae) and their distribution in Pakistan. Pak. J. Agric. Res. 1990, 11, 65–69. [Google Scholar]
- Kuntadi. Nesting habit of Apis cerana F. in nature. Bul. Penelit. Kehutan. 1989, 5, 21–25. [Google Scholar]
- Seeley, T.D. The Ecology of Temperate and Tropical Honeybee Societies: Ecological studies complement physiological ones, offering a new perspective on patterns of honeybee adaptation. Am. Sci. 1983, 71, 264–272. [Google Scholar]
- Seeley, T.D. Measurement of nest cavity volume by the honey bee (Apis mellifera). Behav. Ecol. Sociobiol. 1977, 2, 201–227. [Google Scholar]
- Seeley, T.D.; Morse, R.A. Nest site selection by the honey bee. Insectes Soc. 1978, 25, 323–337. [Google Scholar] [CrossRef]
- Carr, A.J. Asian Honeybee: Possible Environmental Impacts. Report for the Department of Sustainability, Environment, Water, Population and Communities; Sustineo Pty Ltd.: Canberra, Australia, 2011. [Google Scholar]
- New, T.R. Significance of honey bees in the Australian environment: Setting the scene. Vic. Nat. 1997, 114, 4–7. [Google Scholar]
- Manning, R. The honey bee debate: a critique of scientific studies of honey bees Apis mellifera and their alleged impact on Australian wildlife. Vic. Nat. 1997, 114, 13–22. [Google Scholar]
- Oldroyd, B.P.; Lawler, S.H.; Crozier, R.H. Do feral honey-bees (Apis mellifera) and Regent parrots (Polytelis anthopeplus) compete for nest sites? Aust. J. Ecol. 1994, 19, 444–450. [Google Scholar] [CrossRef]
- Paini, D. The Impact of the European Honey Bee (Apis mellifera) on Australian Native Bees; University of Western Australia: Perth, Australia, 2004. [Google Scholar]
- Paini, D.R.; Williams, M.R.; Roberts, J.D. No short-term impact of honey bees on the reproductive success of an Australian native bee. Apidologie 2005, 36, 613–621. [Google Scholar] [CrossRef]
- Paini, D.R. Impact of the introduced honey bee (Apis mellifera) (Hymenoptera: Apidae) on native bees: A review. Austral. Ecol. 2004, 29, 399–407. [Google Scholar] [CrossRef]
- Pyke, G.H. The introduced Honeybee Apis mellifera and the Precautionary Principle: Reducing the conflict. Aust. Zool. 1999, 31, 181–186. [Google Scholar]
- Sugden, E.; Thorp, R.W.; Buchmann, S. Honey bee—Native bee competition: Focal point for environmental change and apicultural response in Australia. Bee World 1996, 77, 26–44. [Google Scholar]
- Paton, D.C. Honeybees in the Australian Environment. Bioscience 1993, 43, 95–103. [Google Scholar] [CrossRef]
- Punchihewa, R.W.K. Beekeeping for Honey Production in Sri Lanka: Management of Asiatic Hive Honeybee Apis Cerana in Its Natural Tropical Monsoonal Environment; Sri Lanka Department of Agriculture and Canadian International Development Agency: Peradeniya, Sri Lanka, 1994.
- Berry, J.A.; Delaplane, K.S. Effects of comb age on honey bee colony growth and brood survivorship. J. Apic Res. 2001, 40, 3–8. [Google Scholar]
- Hadisoesilo, S.; Otis, G.W. Differences in drone cappings of Apis cerana and Apis nigrocincta. J. Apic. Res. 1998, 37, 11–15. [Google Scholar]
- Dyer, F.C.; Seeley, T.D. Nesting behavior and the evolution of worker tempo in four honey bee species. Ecology 1991, 72, 156–170. [Google Scholar] [CrossRef]
- Tong, X.C.; Boot, W.J. Production of reproductives in the honey bee species Apis cerana in northern Vietnam. J. Apic. Res. 2005, 44, 41–48. [Google Scholar]
- Suwannapong, G.; Benbow, M.E.; Nieh, J.C. Biology of Thai Honeybees: Natural History and Threats. In Bees: Biology, Threats and Colonies; Florio, R.M., Ed.; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2011; pp. 1–98. [Google Scholar]
- Kajobe, R.; Roubik, D.W. Honey-Making bee colony abundance and predation by apes and humans in a Uganda forest reserve. Biotropica 2006, 38, 210–218. [Google Scholar] [CrossRef]
- Jaffe, R.; Dietemann, V.; Allsopp, M.H.; Costa, C.; Crewe, R.M.; Dall’Olio, R.; de la Rua, P.; El-Niweiri, M.A.A.; Fries, I.; Kezic, N.; et al. Estimating the density of honeybee colonies across their natural range to fill the gap in pollinator decline censuses. Conserv. Biol. 2009, 24, 583–593. [Google Scholar]
- Moritz, R.F.A.; Kraus, F.B.; Kryger, P.; Crewe, R.M. The size of wild honeybee populations (Apis mellifera) and its implications for the conservation of honeybees. J. Insect Conserv. 2007, 11, 391–397. [Google Scholar] [CrossRef]
- Paton, D.C. Overview of Feral and Managed Honeybees in Australia; Department of Zoology, The University of Adelaide: Canberra, Australia, 1996. [Google Scholar]
- Oldroyd, B.P.; Thexton, E.G.; Lawler, S.H.; Crozier, R.H. Population demography of Australian feral bees (Apis mellifera). Oecologia 1997, 111, 381–387. [Google Scholar] [CrossRef]
- Baum, K.A.; Rubink, W.L.; Pinto, M.A.; Coulson, R.N. Spatial and temporal distribution and nest site characteristics of feral honey bee (Hymenoptera : Apidae) colonies in a coastal prairie landscape. Environ. Entomol. 2005, 34, 610–618. [Google Scholar] [CrossRef]
- Hinson, E. Estimating Feral Honey Bee (Apis mellifera) Colony Density in South Eastern Australia Using Linked Microsatellite Loci; University of Sydney: Sydney, NSW, Australia, 2011. [Google Scholar]
- Koeniger, N.; Koeniger, G.; Gries, M.; Tingek, S. Drone competition at drone congregation areas in four Apis species. Apidologie 2005, 36, 211–221. [Google Scholar] [CrossRef]
- Koeniger, N.; Koeniger, G. Reproductive isolation among species of the genus Apis. Apidologie 2000, 31, 313–339. [Google Scholar] [CrossRef]
- Otis, G.W.; Koeniger, N.; Rinderer, T.E.; Hadisoesilo, S.; Yoshida, T.; Tingek, S.; Wongsiri, S.; Mardan, M. Comparative Mating Flight Times of Asian Honey Bees. In Proceedings of Seventh international conference on tropical bees: Management and diversity and fifth Asian Apicultural Association conference, Chiang Mai, Thailand, 19–25 March 2000; pp. 138–141.
- Commerford, M.M.; Koetz, A.H. Ecology and behaviour of Asian honey bees (Apis cerana Java) in Cairns, Australia. In Asian Honey Bee Transition to Management Program; Department of Agriculture, Fisheries and Forestry (DAFF): Cairns, Queensland, 2013. [Google Scholar]
- Punchihewa, R.W.K.; Koeniger, N.; Koeniger, G. Congregation of Apis cerana indica Fabricius 1798 drones in the canopy of trees in Sri Lanka. Apidologie 1990, 21, 201–208. [Google Scholar] [CrossRef]
- Fujiwara, S.; Miura, H.; Kumagai, T.; Sawaguchi, T.; Naya, S.; Goto, T.; Asanuma, H.; Suzuki, K. Drone congregation of Apis cerana japonica (Radoszkowski, 1877) above large trees. Apidologie 1994, 25, 331–337. [Google Scholar] [CrossRef]
- Koeniger, G.; Koeniger, N.; Phiancharoen, M. Comparative Reproductive Biology and Honeybees. In Honeybees of Asia; Hepburn, C., Radloff, S.E., Eds.; Springer-Verlag berlin: Heidelberg, Germany, 2011. [Google Scholar]
- Hepburn, H.R.; Reece, S.L.; Neumann, P.; Moritz, R.F.A.; Radloff, S.E. Absconding in honeybees (Apis mellifera) in relation to queen status and mode of worker reproduction. Insectes Soc. 1999, 46, 323–326. [Google Scholar] [CrossRef]
- Verma, S.; Attri, P.K. Indigenous beekeeping for sustainable development in Himachal Himalaya. Indian J. Tradit. Knowl. 2008, 7, 221–225. [Google Scholar]
- Hepburn, H.R. Absconding, Migration and Swarming. In Honeybees of Asia; Hepburn, H.R., Radloff, S.E., Eds.; Springer-Verlag Berlin: Heidelberg, Germany, 2011; pp. 133–158. [Google Scholar]
- Dulta, P.C.; Rana, B.S.; Verma, L.R.; Mattu, V.K. Absconding behaviour if the Indian honey bee. Indian Bee J. 1988, 50, 67. [Google Scholar]
- Pokhrel, S.; Thapa, R.; Neupane, F.; Shrestha, S. Absconding behavior and management of Apis cerana F. Honeybee in Chitwan, Nepal. J. Inst. Agric. Anim. Sci. 2006, 27, 77–86. [Google Scholar]
- Fuchs, S.; Tautz, J. Colony Defence and Natural Enemies. In Honeybees of Asia; Hepburn, H.R., Radloff, S.E., Eds.; Springer-Verlag Berlin: Heidelberg, Germany, 2011; pp. 369–395. [Google Scholar]
- Koetz, A.H.; Hyatt, S. Asian Honey Bee (Apis cerana) Remote Nest Treatment. In Asian honey bee Transition to Management Program; Department of Agriculture, Fisheries and Forestry (DAFF): Cairns, Queensland, Australia, 2013. [Google Scholar]
- Tan, K.; Wang, Z.W.; Li, H.; Yang, S.; Hu, Z.W.; Kastberger, G.; Oldroyd, B.P. An “I see you” prey-predator signal between the Asian honeybee, Apis cerana, and the hornet, Vespa velutina. Anim. Behav. 2012, 83, 879–882. [Google Scholar] [CrossRef]
- Chinh, T.X.; Boot, W.J.; Sommeijer, M.J. Production of reproductives in the honey bee species Apis cerana in northern Vietnam. J. Apic Res. 2005, 44, 41–48. [Google Scholar]
- Warhurst, P.; Goebel, R. The Bee Book: Beekeeping in Australia, 2nd ed.Department of Primary Industries and Fisheries: Brisbane, Australia, 2005.
- Sharma, P.L. Observations on the swarming and mating habits of the Indian honeybee. Bee World 1960, 41, 230–232. [Google Scholar]
- Kojima, Y.; Toki, T.; Morimoto, T.; Yoshiyama, M.; Kimura, K.; Kadowaki, T. Infestation of Japanese native honey bees by tracheal mite and virus from non-native European honey bees in Japan. Microb. Ecol. 2011, 62, 895–906. [Google Scholar] [CrossRef]
- Mack, R.N.; Simberloff, D.; Lonsdale, W.M.; Evans, H.; Clout, M.; Bazzaz, F.A. Biotic invasions: Causes, epidemiology, global consequences, and control. Ecol. Appl. 2000, 10, 689–710. [Google Scholar] [CrossRef]
- Oldroyd, B.P. Coevolution while you wait: Varroa jacobsoni, a new parasite of western honeybees. Trends Ecol. Evol. 1999, 14, 312–315. [Google Scholar] [CrossRef]
- Fries, I. Diseases of Asian Honeybees. In Honeybees of Asia; Hepburn, H.R., Radloff, S.E., Eds.; Springer-Verlag Berlin: Heidelberg, Germany, 2011; pp. 333–345. [Google Scholar]
- Anderson, D.L. Viruses of Apis cerana and Apis mellifera. In The Asiatic Hive Bee: Apiculture, Biology, and Role in Sustainable Development in Tropical and Subtropical Asia; Kevan, P.G., Ed.; Enviroquest Ltd.: Cambridge, ON, Canada, 1995; pp. 161–170. [Google Scholar]
- Yang, B.; Peng, G.D.; Li, T.B.; Kadowaki, T. Molecular and phylogenetic characterization of honey bee viruses, Nosema microsporidia, protozoan parasites, and parasitic mites in China. Ecol. Evol. 2013, 3, 298–311. [Google Scholar] [CrossRef]
- Ai, H.X.; Yan, X.; Han, R.C. Occurrence and prevalence of seven bee viruses in Apis mellifera and Apis cerana apiaries in China. J. Invertebr. Pathol. 2012, 109, 160–164. [Google Scholar] [CrossRef]
- Choe, S.E.; Lien, T.K.N.; Noh, J.H.; Koh, H.B.; Jean, Y.H.; Kweon, C.H.; Kang, S.W. Prevalence and distribution of six bee viruses in Korean Apis cerana populations. J. Invertebr. Pathol. 2012, 109, 330–333. [Google Scholar] [CrossRef]
- Li, J.L.; Qin, H.R.; Wu, J.; Sadd, B.M.; Wang, X.H.; Evans, J.D.; Peng, W.J.; Chen, Y.P. The Prevalence of parasites and pathogens in Asian Honeybees Apis cerana in China. PLoS One 2012, 7, 1–12. [Google Scholar]
- Warrit, N.; Lekprayoon, C. Asian Honeybee Mites. In Honeybees of Asia; Hepburn, H.R., Radloff, S.E., Eds.; Springer-Verlag Berlin: Heidelberg, Germany, 2011; pp. 347–368. [Google Scholar]
- Roberts, J.D.; Anderson, D.L. Establishing the Disease Status of the Asian Honeybee in the Cairns Region. Publication No. 13/082; Rural Industries Research and Development Corporation (RIRDC): Canberra, Australia, 2013. [Google Scholar]
- Rath, W. Co-adaptation of Apis cerana Fabr. and Varroa jacobsoni Oud. Apidologie 1999, 30, 97–110. [Google Scholar] [CrossRef]
- Boecking, O. Sealing up and non-removal of diseased and Varroa jacobsoni infested drone brood cells is part of the hygienic behaviour in Apis cerana. J. Apic Res. 1999, 38, 159–168. [Google Scholar]
- Boecking, O.; Spivak, M. Behavioral defenses of honey bees against Varroa jacobsoni Oud. Apidologie 1999, 30, 141–158. [Google Scholar] [CrossRef]
- Rath, W.; Drescher, W. Response of Apis cerana Fabr towards brood infested with Varroa jacobsoni Oud and infestation rate of colonies in Thailand. Apidologie 1990, 21, 311–321. [Google Scholar] [CrossRef]
- Rosenkranz, P.; Tewarson, N.C.; Singh, A.; Engels, W. Differential hygienic behavior towards Varroa jacobsoni in capped worker brood of Apis cerana depends on alien scent adhering to the mites. J. Apic Res. 1993, 32, 89–93. [Google Scholar]
- Sasagawa, H.; Matsuyama, S.; Leal, W.S.; Peng, C.Y.S. Hygienic Grooming Behaviour Induced by Parasitic Varroa Mites in Apis cerana japonica. Asian Bees and Beekeeping: Progress of Research and Development. In Proceedings of fourth Asian Apicultural Association International Conference, Kathmandu, Nepal, 23–28 March 1998.
- Verma, L.R. Beekeeping: In Integrated Mountain Development: Economic And Scientific Perspectives; International Centre for Integrated Mountain Development (ICIMOD): Kathmandu, Nepal, 1990.
- Klein, A.-M.; Vaissiere, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef]
- Genersch, E. Honey bee pathology: Current threats to honey bees and beekeeping. Appl. Microbiol. Biotechnol. 2010, 87, 87–97. [Google Scholar] [CrossRef]
- Momose, K.; Yumoto, T.; Nagamitsu, T.; Kato, M.; Nagamasu, H.; Sakai, S.; Harrison, R.D.; Itioka, T.; Hamid, A.A.; Inoue, T. Pollination biology in a lowland dipterocarp forest in Sarawak, Malaysia. I. Characteristics of the plant-pollinator community in a lowland dipterocarp forest. Am. J. Bot. 1998, 85, 1477–1501. [Google Scholar] [CrossRef]
- Pritchard, K. The Unseen Costs of Agricultural Expansion across a Rainforest Landscape: Depauperate pollinator Communities and Reduced Yield in Isolated Crops. Ph.D. Thesis, James Cook University, Cairns, Australia, 2005. [Google Scholar]
- De Barro, P. The Future of the Australian honey Bee Industry; CSIRO: Canberra, Australia, 2007. [Google Scholar]
- Cunningham, S.A.; FitzGibbon, F.; Heard, T.A. The future of pollinators for Australian agriculture. Aust. J. Agric. Res. 2002, 53, 893–900. [Google Scholar] [CrossRef]
- Verma, L.R.; Partap, U. Foraging behaviour of Apis cerana on cauliflower and cabbage and its impact on seed production. J. Apic. Res. 1994, 33, 231–236. [Google Scholar]
- Verma, L.R. A Framework for R & D on Beekeeping with Asian Hive Bee Apis Cerana. In Proceedings of BEENET Asia: Workshop on Priorities in R&D on beekeeping in tropical Asia, Serdang, Malaysia, 1992.
- Verma, L.R.; Rana, R.S. Further studies on the behaviour of Apis cerana and Apis mellifera foraging on apple flowers. J. Apic Res. 1994, 33, 175–179. [Google Scholar]
- Partap, U.; Verma, L.R. Pollination of radish by Apis cerana. J. Apic Res. 1994, 33, 237–241. [Google Scholar]
- Matsuka, M.; Verma, L.R.; Wongsiri, S.; Shrestha, K.K.; Partap, U. Asian Bees and Beekeeping: Progress of Research and Development. In Proceedings of the Fourth Asian Apicultural Association International Conference, Kathmandu, Nepal, 23–28 March 1998.
- Sihag, R.C.; Mishra, R.C. Crop Pollination and Apis cerana. In The Asiatic Hive Bee: Apiculture, Biology, and Role in Sustainable Development in Tropical and Subtropical Asia; Kevan, P.G., Ed.; Enviroquest Ltd.: Cambridge, ON, Canada, 1995; pp. 135–141. [Google Scholar]
- Khan, B.M. Comparative Study on Pollination Effect of Honeybee Species Apis cerana and Apis mellifera on the Fruit Yield of Toria (Brassica napus) in Peshawar, Pakistan. In The Asiatic Hive Bee: Apiculture, Biology, and Role in Sustainable Development in Tropical and Subtropical Asia; Kevan, P.G., Ed.; Enviroquest Ltd.: Cambridge, Canada, 1995; pp. 151–152. [Google Scholar]
- Klein, A.M.; Steffan-Dewenter, I.; Tscharntke, T. Fruit set of highland coffee increases with the diversity of pollinating bees. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2003, 270, 955–961. [Google Scholar] [CrossRef]
- Atmowidi, T.; Riyanti, F.; Sutrisna, A. Pollination effectiveness of Apis cerana Fabricus and Apis mellifera Linnaeus (Hymenoptera: Apidae) in Jatropha curcas L. (Euphorbiaceae). Biotropia 2008, 15, 129–134. [Google Scholar]
- Gross, C.L. The effect of introduced honeybees on native bee visitation and fruit-set in Dillwynia juniperina (Fabaceae) in a fragmented ecosystem. Biol. Conserv. 2001, 102, 89–95. [Google Scholar] [CrossRef]
- Paton, D.C. Honey bees Apis mellifera and the disruption of plant-pollinator systems in Australia. Vic. Nat. 1997, 114, 23–29. [Google Scholar]
- Gross, C.L.; Mackay, D. Honeybees reduce fitness in the pioneer shrub Melastoma affine (Melastomataceae). Biol. Conserv. 1998, 86, 169–178. [Google Scholar] [CrossRef]
- Gross, C.L.; Gorrell, L.; Macdonald, M.J.; Fatemi, M. Honeybees facilitate the invasion of Phyla canescens (Verbenaceae) in Australia—No bees, no seed! Weed Res. 2010, 50, 364–372. [Google Scholar]
- Simpson, S.R.; Gross, C.L.; Silberbauer, L.X. Broom and honeybees in Australia: An alien liaison. Plant. Biol. 2005, 7, 541–548. [Google Scholar] [CrossRef]
- Gordon, C.E. The coexistence of species. Rev. Chil. Hist. Nat. 2000, 73, 175–198. [Google Scholar] [CrossRef]
- Krebs, C.J. Ecology: The Experimental Analysis of Distribution and Abundance; Harper and Row: New York, NY, USA, 1972. [Google Scholar]
- Sharma, H.K.; Gupta, J.K.; Rana, B.S. Diurnal activity of Apis cerana F. and A. mellifera L. on different flora during spring and honey flow period. Pest Manag. Econ. Zool. 2000, 8, 151–154. [Google Scholar]
- Yang, M.-X.; Tan, K.; Radloff, S.E.; Hepburn, H.R. Interspecific Interactions among Asian Honeybees. In Honeybees of Asia; Hepburn, H.R., Radloff, S.E., Eds.; Springer-Verlag Berlin: Heidelberg, Germany, 2011; pp. 445–472. [Google Scholar]
- Partap, U. Foraging Competition between Apis Cerana and Apis Mellifera and Its Impact on Crop Pollination. Asian bees and beekeeping: Progress of research and development. In Proceedings of fourth Asian Apicultural Association International Conference, Kathmandu, Nepal, 23–28 March, 1998; pp. 206–208.
- Sakagami, S.F. Some interspecific relations between Japanese and European honeybees. J. Anim. Ecol. 1959, 28, 51–68. [Google Scholar] [CrossRef]
- Sharma, H.K.; Gupta, J.K.; Rana, B.S. Resource Partitioning among Apis mellifera and Apis cerana under Mid-Hill Conditions of Himachal Pradesh. Asian Bees and Beekeeping: Progress of Research and Development. In Proceedings of Fourth Asian Apicultural Association International Conference, Kathmandu, Nepal, March 23–28 1998; pp. 213–215.
- Rinderer, T.E.; Marx, B.D.; Gries, M.; Tingek, S. A scientific note on stratified foraging by Sabahan bees on the yellow flame tree (Peltophorum pterocarpum). Apidologie 1996, 27, 423–425. [Google Scholar] [CrossRef]
- Partap, U.; Shukla, A.N.; Verma, L.R. Comparative Foraging Behaviour of Apis cerana and Apis mellifera in Pollinating Peach and Plum Flowers in Kathmandu Valley, Nepal. Asian Bees and Beekeeping: Progress of Research and Development. In proceedings of Fourth Asian Apicultural Association International Conference, Kathmandu, Nepal, 23–28 March 1998; pp. 193–198.
- Wongsiri, S.; Lai, Y.-S.; Liu, Z.-S. Beekeeping in the Guandong province of China and some observations on the Chinese Honeybee Apis cerana cerana and the European honey bee Apis mellifera lingustica. Am. Bee J. 1986, 126, 748–752. [Google Scholar]
- Breed, M.D.; Deng, X.-B.; Buchwald, R. Comparative nestmate recognition in Asian honey bees, Apis florea, Apis andreniformis, Apis dorsata, and Apis cerana. Apidologie 2007, 38, 411–418. [Google Scholar] [CrossRef]
- Annand, N. The Solomon experience with Asian honey bees. Available online: http://www.theabk.com/article/solomon-experience-asian-honey-bees/ (accessed on 1 July 2012).
- Manila-Fajardo, A.C.; Cervancia, C.R. Performance of honey bees (Apis mellifera L.) in three ecosystems in Laguna, Philippines. Philipp. Agric. Sci. 2003, 86, 146–157. [Google Scholar]
- Yoshikawa, K.; Ohgushi, R. Tropical beekeeping in Cambodia. J. Biol. 1965, 16, 81–88. [Google Scholar]
- Juntawong, N.; Pechhacker, H. Apis Mellifera Versus Apis cerana in the North of Thailand. Bees Dev. J. 1994, 30, 6–7. [Google Scholar]
- Yang, G. Harm of introducing the western honeybee Apis mellifera L. to the Chinese honeybee Apis cerana F. and its ecological impact. Acta Entomol. Sin. 2005, 48, 401–406. [Google Scholar]
- Yu, L.; Han, S. Effect of habitat and interspecific competition on Apis cerana cerana colony distribution. J. Appl. Ecol. 2003, 14, 553–556. [Google Scholar]
- Moritz, R.F.A.; Haertel, S.; Neumann, P. Global invasions of the western honeybee (Apis mellifera) and the consequences for biodiversity. Ecoscience 2005, 12, 289–301. [Google Scholar] [CrossRef]
- Tan, N.Q.; Binh, P.T. Harmony or conflict? Apis mellifera and Apis cerana in Southern Vietnam. Beekeep. Dev. 1994, 32, 4–7. [Google Scholar]
- Bellis, G.A.; Profke, A.M. Rainbow bee-eaters (Merops ornatus) as a monitoring tool for honeybees (Apis mellifera L.; Hymenoptera : Apidae). Aust. J. Entomol. 2013, 42, 266–270. [Google Scholar]
- Wilson, D. Use of Bee-Eater Birds in Monitoring for the Asian Honey Bee; Animal Health Australia: Deakin, Australia, 2009. [Google Scholar]
- Harvey, C.T.; Qureshi, S.A.; MacIsaac, H.J. Detection of a colonizing, aquatic, non-indigenous species. Divers. Distrib. 2009, 15, 429–437. [Google Scholar] [CrossRef]
- Ashcroft, M.B.; Gollan, J.R.; Batley, M. Combining citizen science, bioclimatic envelope models and observed habitat preferences to determine the distribution of an inconspicuous, recently detected introduced bee (Halictus smaragdulus Vachal Hymenoptera: Halictidae) in Australia. Biol. Invasions 2012, 14, 515–527. [Google Scholar] [CrossRef]
- Frey, J.K. Distinguishing range expansions from previously undocumented populations using background data from museum records. Divers. Distrib. 2009, 15, 183–187. [Google Scholar] [CrossRef]
- Koetz, A.H. Detection efficacy of Asian honey bees (Apis cerana) in Cairns, Australia. In Asian Honey Bee Transition to Management Program; Department of Agriculture, Fisheries and Forestry (DAFF): Cairns, Queensland, Austraila, 2013. [Google Scholar]
- Crane, E. A short history of knowledge about honey bees (Apis) up to 1800. Bee World 2004, 85, 6–11. [Google Scholar]
- Sasaki, M.; Ono, M.; Asada, S.; Yoshida, T. Oriental orchid (Cymbidium pumilum) attracts drones of the Japanese honeybee (Apis cerana japonica) as pollinators. Experientia 1991, 47, 1229–1231. [Google Scholar] [CrossRef]
- Sasaki, M. Deception of the Japanese honey bee, Apis cerana japonica, by the oriental orchid Cymbidium pumilum. Honeybee Sci. 1992, 13, 167–172. [Google Scholar]
- Sugahara, M.; Minamoto, T.; Fuchikawa, T.; Michinomae, M.; Shimizu, I. Apis cerana japonica Discriminates between floral color phases of the oriental orchid, Cymbidium floribundum. Zool. Sci. 2010, 27, 901–906. [Google Scholar] [CrossRef]
- Koltermann, R. Race-Specific and species-specific scent evaluation in honeybees and their ecological adaptation. Rassen- bzw. artspezifische Duftbewertung bei der Honigbiene und ökologische Adaptation. (German). J. Comp. Physiol. 1973, 85, 327–360. [Google Scholar] [CrossRef]
- Williams, J.L. Wind-Directed pheromone trap for drone honey bees (Hymenoptera: Apidae). J. Econ. Entomol. 1987, 80, 532–536. [Google Scholar]
- Plettner, E.; Otis, G.W.; Wimalaratne, P.D.C.; Winston, M.L.; Slessor, K.N.; Pankiw, T.; Punchihewa, P.W.K. Species- and caste-determined mandibular gland signals in honeybees (Apis). J. Chem. Ecol. 1997, 23, 363–377. [Google Scholar]
- Lacey, M. Identification and Application of the Aggregation Pheromone of Apis Cerana. In RIRDC Project No CSE-74A; Rural Industries Research And Development Corporation (RIRDC): Barton, Australia, 1999. [Google Scholar]
- Lacey, M. Development of a Specific Aggregation Lure for Apis Cerana Javana—A Report for the Rural Industries Research and Development Corporation; Rural Industries Research And Development Corporation (RIRDC): Barton, Australia, 2000. [Google Scholar]
- Keeling, C.I.; Otis, G.W.; Hadisoesilo, S.; Slessor, K.N. Mandibular gland component analysis in the head extracts of Apis cerana and Apis nigrocincta. Apidologie 2001, 32, 243–252. [Google Scholar] [CrossRef]
- Schmidt, J.O.; Morgan, E.D.; Oldham, N.J.; DoNascimento, R.R.; Dani, F.R. (Z)-11-eicosen-1-ol, a major component of Apis cerana venom. J. Chem. Ecol. 1997, 23, 1929–1939. [Google Scholar] [CrossRef]
- Anderson, D.L.; Trueman, J.W.H. Varroa jacobsoni (Acari : Varroidae) is more than one species. Exp. Appl. Acarol. 2000, 24, 165–189. [Google Scholar] [CrossRef]
- Pirk, C.W.W.; Sole, C.L.; Crewe, R.M. Pheromones. In Honeybees of Asia; Hepburn, H.R., Radloff, S.E., Eds.; Springer-Verlag Berlin: Heidelberg, Germany, 2011; pp. 207–214. [Google Scholar]
- Naik, D.G.; Gadre, R.V.; Kapadi, A.H.; Singh, M.K.; Suryanarayana, M.C.; Kshirsagar, K.K. Nasonov gland pheromone of the Indian honeybee, Apis cerana indica. J. Apic Res. 1988, 27, 205–206. [Google Scholar]
- Matsuyama, S.; Suzuki, H.; Sasagawa, H. Chemical Analysis of Worker Pheromone Component of Asian Honeybees. In Proceedings of the 4th Asian Apiculture Association International Conference, Kathmandu, Nepal, 2000; p. 24.
- Kuang, B.; Kuang, H.; Tan, K. Effect of Synthetic Queen Pheromone on the Behaviour of Apis cerana. Asian Bees and Beekeeping: Progress of Research and Development. In Proceedings of the Fourth Asian Apicultural Association International Conference, Kathmandu, Nepal, 23–28 March 1998.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Koetz, A.H. Ecology, Behaviour and Control of Apis cerana with a Focus on Relevance to the Australian Incursion. Insects 2013, 4, 558-592. https://doi.org/10.3390/insects4040558
Koetz AH. Ecology, Behaviour and Control of Apis cerana with a Focus on Relevance to the Australian Incursion. Insects. 2013; 4(4):558-592. https://doi.org/10.3390/insects4040558
Chicago/Turabian StyleKoetz, Anna H. 2013. "Ecology, Behaviour and Control of Apis cerana with a Focus on Relevance to the Australian Incursion" Insects 4, no. 4: 558-592. https://doi.org/10.3390/insects4040558