Responses to Pheromones in a Complex Odor World: Sensory Processing and Behavior


Abstract
:
1. Introduction

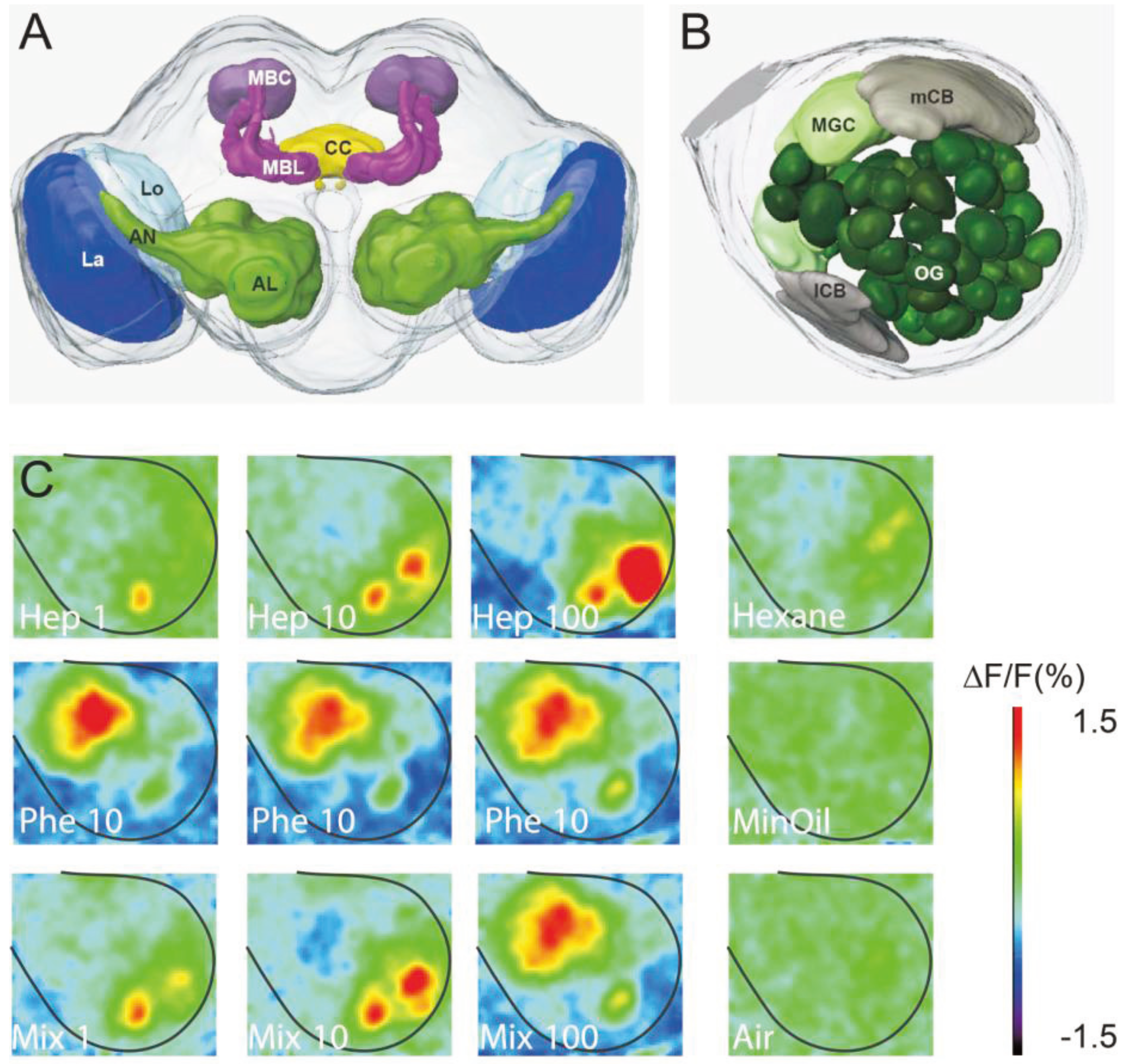
2. The Reception of Sex Pheromones and Plant Volatiles

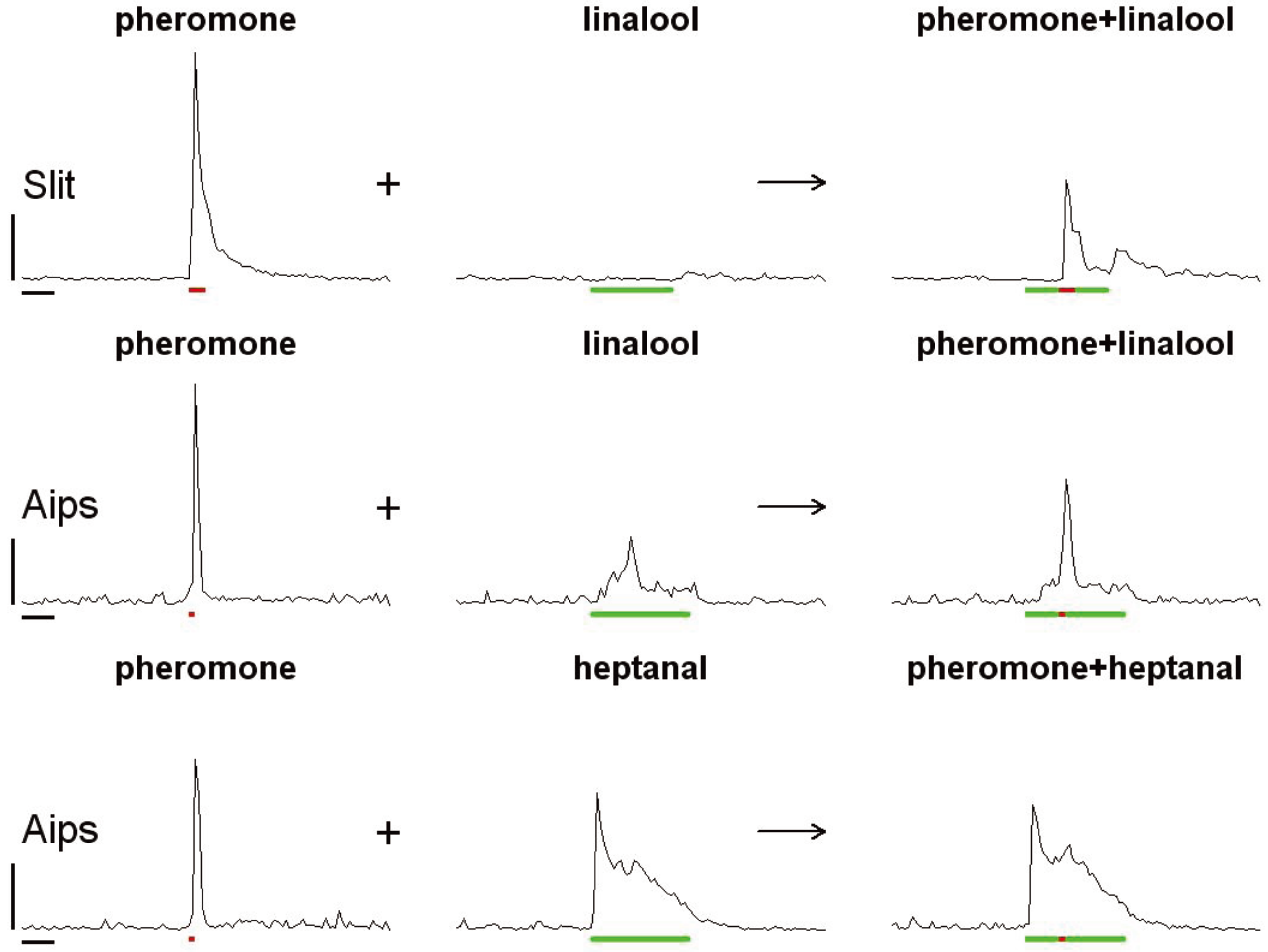
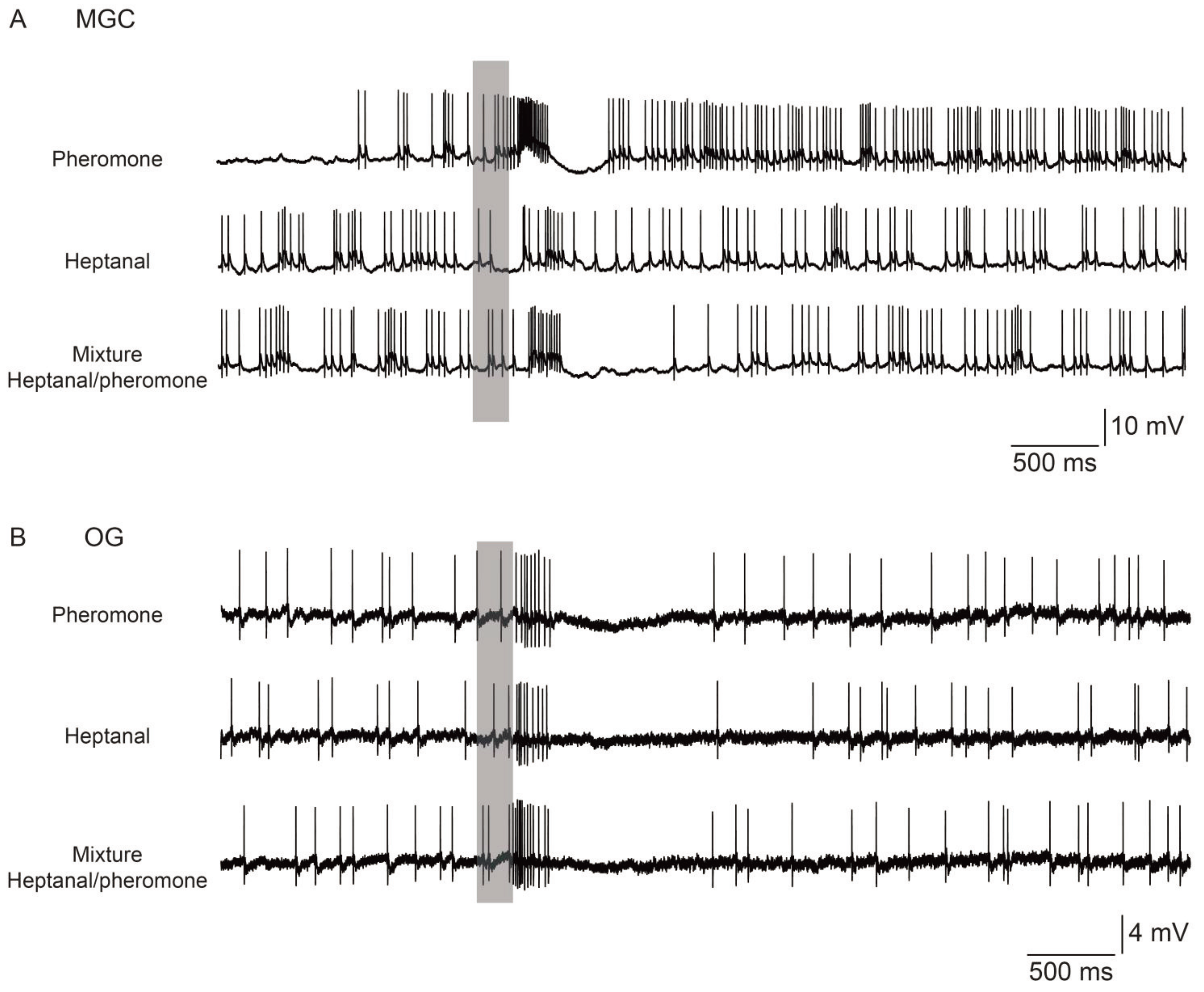
3. Pheromone-Plant Odor Interactions and Signal Coding


4. Plasticity of Pheromone-Plant Odor Interactions
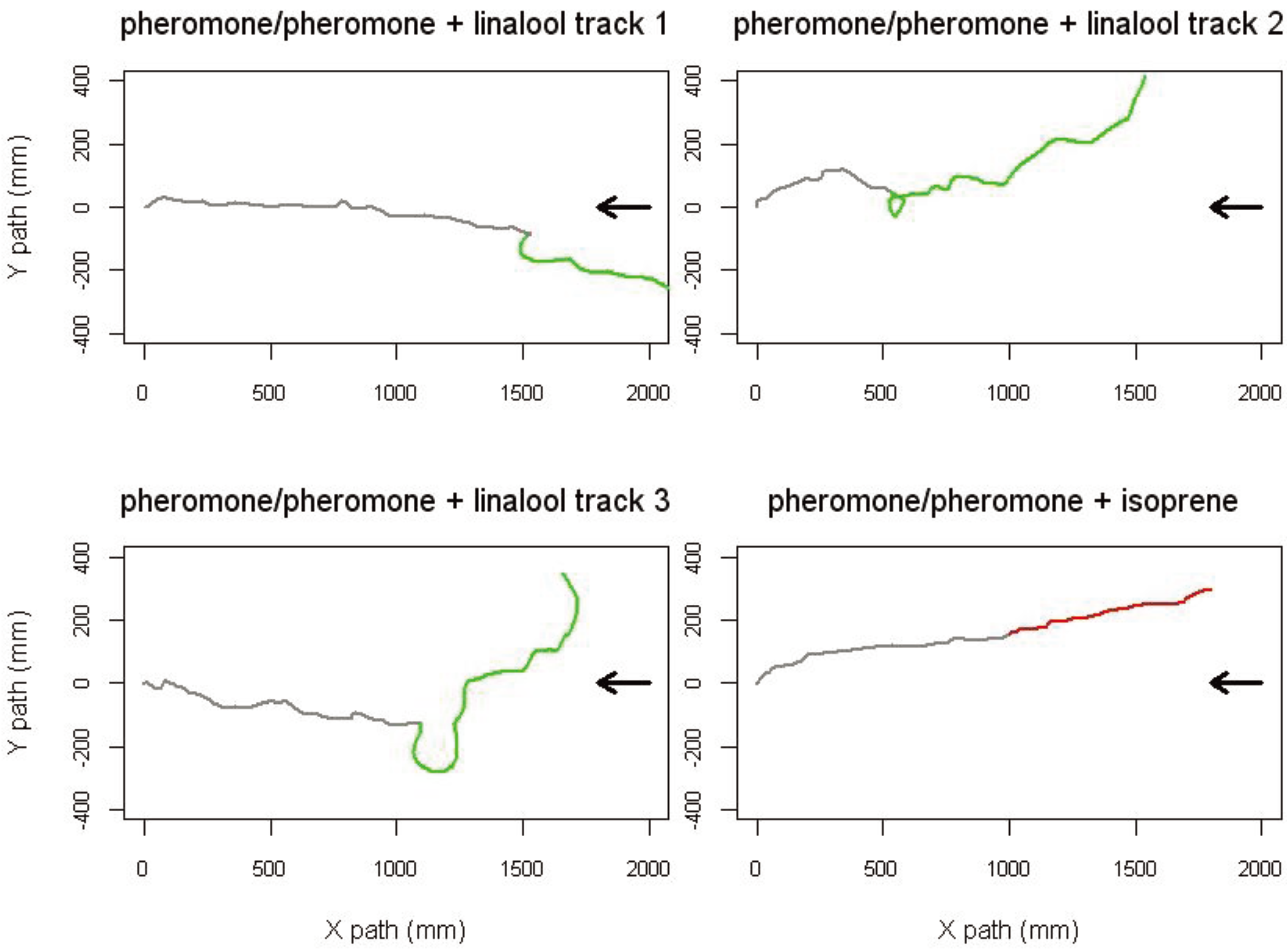
5. Orientation Behavior

6. Adaptive Value of Pheromone-Plant Volatile Interactions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Insect group | species | Pheromone function | Plant odour type | Compounds | Effects | References |
|---|---|---|---|---|---|---|
| Homoptera (various Aphids) | Rhopalosiphon padi, Phorodon humuli | Sex pheromone (female-emitted) | Single host plant volatiles | Benzaldehyde, methyl salicylate | Increased catches to pheromone baited traps in field studies and increased specificity. | [89] |
| Dysaphis plantaginea | Induced host plant volatiles | Adult-feeding induced green leaf volatiles. (Short chain esters) | [90] | |||
| Coleoptera | Leptinotarsa decemlineata | Aggregation pheromone (male produced) | Blend of host plant volatiles | (Z)-3-hexenyl acetate, linalool, methyl salicylate, nonanal, 2-phenylethanol | Increased attraction in a sex-dependent manner | [91] |
| Melolontha hippocastani, Melolontha melolontha | Sex pheromone (female emitted) | Induced host plant volatiles | (Z)3-hexenal, (Z)-2-hexenal, (E)-2-hexenal, (Z)-3-hexen-1-ol, and other green leaf alcohols | Synergistic attraction of males | [92,93] | |
| Pityogenes bidentatus (Scolytidae) | Aggregation pheromone involved in host colonization | Volatiles from non-host trees | Monoterpenes (α- and β-pinene, terpinolene, 3-carene) and green leaf alcohols, (Z)-3-hexenol, (E)-2-hexenol, 1-hexanol | Inhibition of attraction to pheromone | [4,94] | |
| Rhynchophorus spp. | Aggregation pheromone involved in reproduction | Natural material or volatiles from damaged host-plant | Short chain products of fermentation (ethyl acetate, acetoin) | Synergy | [95,96,97] |
6.1. Improving Mate Finding?
6.2. Better Host Exploitation?
6.3. Better Identification?
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cardé, R.T.; Willis, M.A. Navigational strategies used by insects to find distant, wind-borne sources of odor. J. Chem. Ecol. 2008, 34, 854–866. [Google Scholar] [CrossRef]
- Allmann, S.; Späthe, A.; Bisch-Knaden, S.; Kallenbach, M.; Reinecke, A.; Sachse, S.; Baldwin, I.T.; Hansson, B.S. Feeding-induced rearrangement of green leaf volatiles reduces moth oviposition. eLife 2013, 2, e00421. [Google Scholar] [CrossRef]
- Clavijo McCormick, A.; Irmisch, S.; Reinecke, A.; Boeckler, G.A.; Veit, D.; Reichelt, M.; Hansson, B.S.; Gershenzon, J.; Köllner, T.G.; Unsicker, S.B. Herbivore-induced volatile emission in black poplar—Regulation and role in attracting herbivore enemies. Plant Cell Environ. 2014. [Google Scholar] [CrossRef]
- Byers, J.A.; Zhang, Q.-H.; Birgersson, G. Avoidance of nonhost plants by a bark beetle, Pityogenes bidentatus, in a forest of odors. Naturwissenschaften 2004, 91, 215–219. [Google Scholar] [CrossRef]
- Reddy, G.V.P.; Guerrero, A. Interactions of insect pheromones and plant semiochemicals. Trends Plant Sci. 2004, 9, 253–261. [Google Scholar] [CrossRef]
- Vickers, N.J.; Christensen, T.A.; Baker, T.C.; Hildebrand, J.G. Odour-plume dynamics influence the brain’s olfactory code. Nature 2001, 410, 466–470. [Google Scholar]
- Beyaert, I.; Hilker, M. Plant odour plumes as mediators of plant-insect interactions. Biol. Rev. 2014, 89, 68–81. [Google Scholar] [CrossRef]
- Hansson, B.S. Insect Olfaction; Springer: Berlin, Germany, 1999. [Google Scholar]
- Hansson, B.S.; Larsson, M.C.; Leal, W.S. Green leaf volatile-detecting olfactory receptor neurones display very high sensitivity and specifity in a scarab beetle. Physiol. Entomol. 1999, 24, 121–126. [Google Scholar]
- Bengtsson, J.M.; Khbaish, H.; Reinecke, A.; Wolde-Hawariat, Y.; Negash, M.; Seyoum, E.; Hansson, B.S.; Hillbur, Y.; Larsson, M.C. Conserved, highly specialized olfactory receptor neurons for food compounds in two congeneric scarab beetles, Pachnoda interrupta and Pachnoda marginata. Chem. Senses 2011, 36, 499–513. [Google Scholar] [CrossRef]
- Ansebo, L.; Ignell, R.; Löfqvist, J.; Hansson, B.S. Responses to sex pheromone and plant odours by olfactory receptor neurons housed in sensilla auricillica of the codling moth, Cydia pomonella (Lepidoptera: Tortricidae). J. Insect Physiol. 2005, 51, 1066–1074. [Google Scholar] [CrossRef]
- Van der Pers, J.; Thomas, G.; den Otter, C. Interactions between plant odours and pheromone reception in small ermine moths (Lepidoptera: Yponomeutidae). Chem. Senses 1980, 5, 367–371. [Google Scholar] [CrossRef]
- Party, V.; Hanot, C.; Said, I.; Rochat, D.; Renou, M. Plant terpenes affect intensity and temporal parameters of pheromone detection in a moth. Chem. Senses 2009, 34, 763–774. [Google Scholar] [CrossRef]
- Party, V.; Hanot, C.; Busser, D.S.; Rochat, D.; Renou, M. Changes in odor background affect the locomotory response to pheromone in moths. PLoS One 2013, 8, e52897. [Google Scholar]
- Deisig, N.; Kropf, J.; Vitecek, S.; Pevergne, D.; Rouyar, A.; Sandoz, J.-C.; Lucas, P.; Gadenne, C.; Anton, S.; Barrozo, R. Differential interactions of sex pheromone and plant odour in the olfactory pathway of a male moth. PLoS One 2012, 7, e33159. [Google Scholar] [CrossRef] [Green Version]
- Rouyar, A. Rôle des odeurs de plantes sur l’orientation à la phéromone sexuelle chez les noctuelles. PhD Thesis, Université Pierre et Marie Curie, Paris, France, 2013. [Google Scholar]
- Ochieng, S.A.; Park, K.C.; Baker, T.C. Host plant volatiles synergise responses of sex pheromone-specific olfactory receptor neurons in male Helicoverpa zea. J. Comp. Physiol. A 2002, 188, 325–333. [Google Scholar] [CrossRef]
- Hillier, N.K.; Vickers, N.J. Mixture interactions in moth olfactory physiology: Examining the effects of odorant mixture, concentration, distal stimulation, and antennal nerve transection on sensillar responses. Chem. Senses 2011, 36, 93–108. [Google Scholar] [CrossRef]
- Pregitzer, P.; Schubert, M.; Breer, H.; Hansson, B.S.; Sachse, S.; Krieger, J. Plant odorants interfere with detection of sex pheromone signals by male Heliothis virescens. Front. Cell. Neurosci. 2012, 6, 42. [Google Scholar]
- Galizia, C.G.; Sachse, S.; Mustaparta, H. Calcium responses to pheromones and plant odours in the antennal lobe of the male and female moth Heliothis virescens. J. Comp. Physiol. A 2000, 186, 1049–1063. [Google Scholar] [CrossRef]
- Blejec, A. Statistical method for detection of firing rate changes in spontaneously active neurons. Neurocomputing 2005, 65–66, 557–563. [Google Scholar] [CrossRef]
- Vogt, R.G. Molecular basis of pheromone detection in insects. In Comprehensive Insect Physiology Biochemistry Pharmacology and Molecular Biology. Endocrinology; Gilbert, L.I., Iatro, K., Gill, S., Eds.; Elsevier: London, UK, 2005; Volume 3, pp. 753–804. [Google Scholar]
- Pottier, M.A.; Bozzolan, F.; Chertemps, T.; Jacquin-Joly, E.; Lalouette, L.; Siaussat, D.; Maibeche-Coisne, M. Cytochrome p450s and cytochrome p450 reductase in the olfactory organ of the cotton leafworm Spodoptera littoralis. Insect Mol. Biol. 2012, 21, 568–580. [Google Scholar] [CrossRef]
- Durand, N.; Carot-Sans, G.; Bozzolan, F.; Rosell, G.; Siaussat, D.; Debernard, S.; Chertemps, T.; Maibeche-Coisne, M. Degradation of pheromone and plant volatile components by a same odorant-degrading enzyme in the cotton leafworm, Spodoptera littoralis. PLoS One 2011, 6, e29147. [Google Scholar] [CrossRef]
- Anton, S.; Homberg, U. Antennal lobe structure. In Insect Olfaction; Hansson, B.S., Ed.; Springer: Berlin, Gemany, 1999; pp. 98–125. [Google Scholar]
- Gao, Q.; Yuan, B.; Chess, A. Convergent projections of drosophila olfactory neurons to specific glomeruli in the antennal lobe. Nat. Neurosci. 2000, 3, 780–785. [Google Scholar] [CrossRef]
- Vosshall, L.B.; Wong, A.M.; Axel, R. An olfactory sensory map in the fly brain. Cell 2000, 102, 147–159. [Google Scholar] [CrossRef]
- Galizia, C.G.; Sachse, S.; Rappert, A.; Menzel, R. The glomerular code for odor representation is species specific in the honeybee Apis mellifera. Nat. Neurosci. 1999, 2, 473–478. [Google Scholar] [CrossRef]
- Rospars, J.P. Structure and development of the insect antennodeutocerebral system. Int. J. Insect Morphol. Embryol. 1988, 17, 243–294. [Google Scholar] [CrossRef]
- Rospars, J.P.; Hildebrandt, J.G. Sexually dimorphic and isomorphic glomeruli in the antennal lobes of the sphinx moth Manduca sexta. Chem. Senses 2000, 25, 119–129. [Google Scholar] [CrossRef]
- Reisenmann, C.E.; Christensen, T.A.; Francke, W.; Hildebrand, J.G. Enantioselectivity of projection neurons innervating identified olfactory glomeruli. J. Neurosci. 2004, 24, 2602–2611. [Google Scholar] [CrossRef]
- Ha, T.S.; Smith, D.P. A pheromone receptor mediates 11-cis-vaccenyl acetate-induced responses in Drosophila. J. Neurosci. 2006, 26, 8727–8733. [Google Scholar] [CrossRef]
- Krieger, J.; Grosse-Wilde, E.; Gohl, T.; Dewer, Y.M.E.; Raming, K.; Breer, H. Genes encoding candidate pheromone receptors in a moth (Heliothis virescens). Proc. Natl. Acad. Sci. USA 2004, 101, 11845–11850. [Google Scholar]
- Sakurai, T.; Nakagawa, T.; Mitsuno, H.; Mori, H.; Endo, Y. Identification and functional characterization of a sex pheromone receptor in the silkmoth Bombyx mori. Proc. Natl. Acad. Sci. USA 2004, 101, 16653–16658. [Google Scholar]
- Berg, B.G.; Almaas, T.J.; Bjaalie, J.G. The macroglomerular complex of the antennal lobe in the tobacco budworm moth Heliothis virescens: Specified subdivision in four compartments according to information about biologically significant compounds. J. Comp. Physiol. A 1998, 183, 669–682. [Google Scholar] [CrossRef]
- Kárpáti, Z.; Dekker, T.; Hansson, B.S. Reversed functional topology in the antennal lobe of the male european corn borer. J. Exp. Biol. 2008, 211, 2841–2848. [Google Scholar]
- Homberg, U.; Christensen, T.A.; Hildebrand, J.G. Structure and function of the deutocerebrum in insects. Annu. Rev. Entomol. 1989, 34, 477–501. [Google Scholar] [CrossRef]
- Hansson, B.S.; Christensen, T.A. Functional characteristics of the antennal lobe. In Insect Olfaction; Hansson, B.S., Ed.; Springer: Berlin, Gemany, 1999; pp. 126–164. [Google Scholar]
- Martin, J.P.; Beyerlein, A.; Dacks, A.M.; Reisenman, C.E.; Riffell, J.A.; Lei, H.; Hildebrand, J.G. The neurobiology of insect olfaction: Sensory processing in a comparative context. Prog. Neurobiol. 2011, 95, 427–447. [Google Scholar] [CrossRef]
- Husch, A.; Paehler, M.; Fusca, D.; Paeger, L.; Kloppenburg, P. Distinct electrophysiological properties in subtypes of nonspiking olfactory local interneurons correlate with their cell type-specific Ca2+ current profiles. J. Neurophysiol. 2009, 102, 2834–2845. [Google Scholar] [CrossRef]
- Seki, Y.; Rybak, J.; Wicher, D.; Sachse, S.; Hansson, B.S. Physiological and morphological characterization of local interneurons in the Drosophila antennal lobe. J. Neurophysiol. 2010, 104, 1007–1019. [Google Scholar] [CrossRef]
- Reisenman, C.E.; Dacks, A.M.; Hildebrand, J.G. Local interneuron diversity in the primary olfactory center of the moth Manduca sexta. J. Comp. Physiol. A 2011, 197, 653–665. [Google Scholar] [CrossRef]
- Schachtner, J.; Schmidt, M.; Homberg, U. Organization and evolutionary trends of primary olfactory brain centers in Tetraconata (Crustacea plus Hexapoda). Arthropod Struct. Dev. 2005, 34, 257–299. [Google Scholar] [CrossRef]
- Ignell, R.; Root, C.M.; Birse, R.T.; Wang, J.W.; Nässel, D.R.; Winther, A.M.E. Presynaptic peptidergic modulation of olfactory receptor neurons in Drosophila. Proc. Natl. Acad. Sci. USA 2009, 106, 13070–13075. [Google Scholar]
- Rescorla, R.A.; Wagner, A.R. A theory of pavlovian conditioning: Variations in the effectiveness of reinforcement and non-reinforcement. In Classical Conditioning II: Current Research and Theory; Black, A.H., Prokasy, W.F., Eds.; Appleton-Century-Crofts: New York, NY, USA, 1972; pp. 64–99. [Google Scholar]
- Deisig, N.; Giurfa, M.; Sandoz, J.C. Antennal Lobe Processing Increases Separability of Odor Mixture Representations in the Honeybee. J. Neurophysiol. 2010, 103, 2185–2195. [Google Scholar] [CrossRef]
- Lei, H.; Vickers, N. Central processing of natural odor mixtures in insects. J. Chem. Ecol. 2008, 34, 915–927. [Google Scholar] [CrossRef]
- Hallem, E.A.; Carlson, J.R. Coding of odors by a receptor repertoire. Cell 2006, 125, 143–160. [Google Scholar] [CrossRef]
- Andersson, M.N.; Larsson, M.C.; Schlyter, F. Specificity and redundancy in the olfactory system of the bark beetle Ips typographus: Single-cell responses to ecologically relevant odors. J. Insect Physiol. 2009, 55, 556–567. [Google Scholar] [CrossRef]
- Joerges, J.; Küttner, A.; Galizia, C.G.; Menzel, R. Representations of odours and odour mixtures visualized in the honeybee brain. Nature 1997, 387, 285–288. [Google Scholar] [CrossRef]
- Deisig, N.; Giurfa, M.; Lachnit, H.; Sandoz, J.-C. Neural representation of olfactory mixtures in the honeybee antennal lobe. Eur. J. Neurosci. 2006, 24, 1161–1174. [Google Scholar] [CrossRef]
- Carlsson, M.A.; Chong, K.Y.; Daniels, W.; Hansson, B.S.; Pearce, T.C. Component information is preserved in glomerular responses to binary odor mixtures in the moth Spodoptera littoralis. Chem. Senses 2007, 32, 433–443. [Google Scholar] [CrossRef]
- Silbering, A.F.; Galizia, C.G. Processing of odor mixtures in the Drosophila antennal lobe reveals both global inhibition and glomerulus-specific interactions. J. Neurosci. 2007, 27, 11966–11977. [Google Scholar] [CrossRef]
- Sachse, S.; Galizia, C.G. Role of inhibition for temporal and spatial odor representation in olfactory output neurons: A calcium imaging study. J. Neurophysiol. 2002, 87, 1106–1117. [Google Scholar]
- Anton, S.; Hansson, B.S. Sex-pheromone and plant-associated odor processing in antennal lobe interneurons of male Spodoptera littoralis (Lepidoptera, Noctuidae). J. Comp. Physiol. A 1995, 176, 773–789. [Google Scholar]
- Chaffiol, A.; Kropf, J.; Barrozo, R.B.; Gadenne, C.; Rospars, J.-P.; Anton, S. Plant odour stimuli reshape pheromonal representation in neurons of the antennal lobe macroglomerular complex of a male moth. J. Exp. Biol. 2012, 215, 1670–1680. [Google Scholar] [CrossRef] [Green Version]
- Trona, F.; Anfora, G.; Bengtsson, M.; Witzgall, P.; Ignell, R. Coding and interaction of sex pheromone and plant volatile signals in the antennal lobe of the codling moth Cydia pomonella. J. Exp. Biol. 2010, 213, 4291–4303. [Google Scholar] [CrossRef]
- Chaffiol, A.; Dupuy, F.; Barrozo, R.B.; Kropf, J.; Renou, M.; Rospars, J.P.; Anton, S. Pheromone modulates plant odour responses in the antennal lobe a moth. Chem. Senses 2014. [Google Scholar] [CrossRef] [Green Version]
- Namiki, S.; Iwabuchi, S.; Kanzaki, R. Representation of a mixture of pheromone and host plant odor by antennal lobe projection neurons of the silkmoth Bombyx mori. J. Comp. Physiol. A 2008, 194, 501–515. [Google Scholar]
- Barrozo, R.B.; Gadenne, C.; Anton, S. Switching attraction to inhibition: Mating-induced reversed role of sex pheromone in an insect. J. Exp. Biol. 2010, 213, 2933–2939. [Google Scholar] [CrossRef]
- Gronenberg, W.; López-Riquelme, G.O. Multisensory convergence in the mushroom bodies of ants and bees. Acta Biol. Hung. 2005, 55, 31–37. [Google Scholar] [CrossRef]
- Balkenius, A.; Bisch-Knaden, S.; Hansson, B. Interaction of visual and odour cues in the mushroom body of the hawkmoth Manduca sexta. J. Exp. Biol. 2009, 212, 535–541. [Google Scholar] [CrossRef]
- Tanaka, N.K.; Awasaki, T.; Shimada, T.; Ito, K. Integration of chemosensory pathways in the Drosophila second-order olfactory centers. Curr. Biol. 2004, 14, 449–457. [Google Scholar] [CrossRef]
- Jefferis, G.S.X.E.; Potter, C.J.; Chan, A.M.; Marin, E.C.; Rohlfing, T.; Maurer, C.R.; Luo, L. Comprehensive maps of Drosophila higher olfactory centers: Spatially segregated fruit and pheromone representation. Cell 2007, 128, 1187–1203. [Google Scholar] [CrossRef]
- Homberg, U.; Montague, R.A.; Hildebrand, J.G. Anatomy of antenno-cerebral pathways in the brain of the sphinx moth Manduca sexta. Cell Tissue Res. 1988, 254, 255–281. [Google Scholar]
- Kanzaki, R.; Soo, K.; Seki, Y.; Wada, S. Projections to higher olfactory centers from subdivisions of the antennal lobe macroglomerular complex of the male silkmoth. Chem. Senses 2003, 28, 113–130. [Google Scholar] [CrossRef]
- Szyszka, P.; Ditzen, M.; Galkin, A.; Galizia, C.; Menzel, R. Sparsening and temporal sharpening of olfactory representations in the honeybee mushroom bodies. J. Neurophysiol. 2005, 94, 3303–3313. [Google Scholar] [CrossRef]
- Kirschner, S.; Kleineidam, C.; Zube, C.; Rybak, J.; Grünewald, B.; Rössler, W. Dual olfactory pathway in the honeybee, Apis mellifera. J. Comp. Neurol. 2006, 499, 933–952. [Google Scholar] [CrossRef]
- Gadenne, C.; Dufour, M.; Anton, S. Transient post-mating inhibition of behavioural and central nervous responses to sex pheromone in an insect. Proc. R. Soc. B Biol. Sci. 2001, 268, 1631–1635. [Google Scholar] [CrossRef]
- Evenden, M.L.; Gries, R. Plasticity of male response to sex pheromone depends on physiological state in a long-lived moth. Anim. Behav. 2008, 75, 663–672. [Google Scholar] [CrossRef]
- Barrozo, R.B.; Jarriault, D.; Deisig, N.; Gemeno, C.; Monsempes, C.; Lucas, P.; Gadenne, C.; Anton, S. Mating-induced differential coding of plant odour and sex pheromone in a male moth. Eur. J. Neurosci. 2011, 33, 1841–1850. [Google Scholar] [CrossRef]
- Saveer, A. Recognition and modulation of olfactory signals in the noctuid moth Spodoptera littoralis. PhD Thesis, Swedish Agricultural University, Alnarp, Sweden, 2012. [Google Scholar]
- Minoli, S.; Kauer, I.; Colson, V.; Party, V.; Renou, M.; Anderson, P.; Gadenne, C.; Marion-Poll, F.; Anton, S.; Coleman, M.J. Brief exposure to sensory cues elicits stimulus-nonspecific general sensitization in an insect. PLoS One 2012, 7, e34141. [Google Scholar] [CrossRef] [Green Version]
- Anton, S.; Evengaard, K.; Barrozo, R.B.; Anderson, P.; Skals, N. Brief predator sound exposure elicits behavioral and neuronal long-term sensitization in the olfactory system of an insect. Proc. Natl. Acad. Sci. USA 2011, 108, 3401–3405. [Google Scholar] [CrossRef]
- Yang, Z.H.; Casado, D.; Ioriatti, C.; Bengtsson, M.; Witzgall, P. Pheromone pre-exposure and mating modulate codling moth (Lepidoptera: Tortricidae) response to host plant volatiles. Agric. For. Entomol. 2005, 7, 231–236. [Google Scholar] [CrossRef]
- Thöming, G.; Larsson, M.C.; Hansson, B.S.; Anderson, P. Comparison of plant preference hierarchies of male and female moths and the impact of larval rearing hosts. Ecology 2013, 94, 1744–1752. [Google Scholar] [CrossRef]
- Deng, J.-Y.; Wei, H.; Huang, Y.-P.; Du, J.-W. Enhancement of attraction to sex pheromones of Spodoptera exigua by volatile compounds produced by host plants. J. Chem. Ecol. 2004, 30, 2037–2045. [Google Scholar] [CrossRef]
- Meagher, J.R.L.; Mitchell, E.R. Phenylacetaldehyde enhances upwind flight of male fall armyworm (Lepidoptera: Noctuidae) to its sex pheromone. Fla. Entomol. 1998, 81, 554–559. [Google Scholar]
- Light, D.M.; Flath, R.A.; Buttery, R.G.; Zalom, F.G.; Rice, R.E.; Dickens, J.C.; Jang, E.B. Host-plant green-leaf volatiles synergize the synthetic sex pheromones of the corn earworm and codling moth (Lepidoptera). Chemoecology 1993, 4, 145–152. [Google Scholar] [CrossRef]
- Meagher, R.L.; Landolt, P.J. Attractiveness of binary blends of floral odorant compounds to moths in florida, USA. Entomol. Exp. Appl. 2008, 128, 323–329. [Google Scholar] [CrossRef]
- Dickens, J.C.; Smith, J.W.; Light, D.M. Green leaf volatiles enhance sex attractant pheromone of the tobacco budworm, Heliothis virescens (Lepidoptera: Noctuidae). Chemoecology 1993, 4, 175–177. [Google Scholar] [CrossRef]
- Meagher, J.R.L. Trapping fall armyworm (Lepidoptera: Noctuidae) adults in traps baited with pheromone and a synthetic floral compound. Fla. Entomol. 2001, 84, 288–292. [Google Scholar] [CrossRef]
- Kvedaras, O.L.; del Socorro, A.; Gregg, P.C. Effects of phenylacetaldehyde and (z)-3-hexenyl acetate on male response to synthetic sex pheromone in Helicoverpa armigera (Hübner) (Lepidoptera : Noctuidae). Aust. J. Entomol. 2007, 46, 224–230. [Google Scholar] [CrossRef]
- Coracini, M.; Bengtsson, M.; Liblikas, I.; Witzgall, P. Attraction of codling moth males to apple volatiles. Entomol. Exp. Appl. 2004, 110, 1–10. [Google Scholar] [CrossRef]
- Yang, Z.H.; Bengtsson, M.; Witzgall, P. Host plant volatiles synergize response to sex pheromone in codling moth, Cydia pomonella. J. Chem. Ecol. 2004, 30, 619–629. [Google Scholar] [CrossRef]
- Varela, N.; Avilla, J.; Anton, S.; Gemeno, C. Synergism of pheromone and host-plant volatile blends in the attraction of Grapholita molesta males. Entomol. Exp. Appl. 2011, 141, 114–122. [Google Scholar] [CrossRef]
- Von Arx, M.; Schmidt-Büsser, D.; Guerin, P.M. Plant volatiles enhance behavioral responses of grapevine moth males, Lobesia botrana to sex pheromone. J. Chem. Ecol. 2012, 38, 222–225. [Google Scholar] [CrossRef]
- Schmidt-Büsser, D.; von Arx, M.; Guerin, P.M. Host plant volatiles serve to increase the response of male european grape berry moths, Eupoecilia ambiguella, to their sex pheromone. J. Comp. Physiol. A 2009, 195, 853–864. [Google Scholar] [CrossRef]
- Pope, T.W.; Campbell, C.A.; Hardie, J.; Pickett, J.A.; Wadhams, L.J. Interactions between host-plant volatiles and the sex pheromones of the bird cherry-oat aphid, Rhopalosiphum padi and the damson-hop aphid, Phorodon humuli. J. Chem. Ecol. 2007, 33, 157–165. [Google Scholar]
- Van Tol, R.W.H.M.; Helsen, H.H.M.; Griepink, F.C.; de Kogel, W.J. Female-induced increase of host-plant volatiles enhance specific attraction of aphid male Dysaphis plantaginea (Homoptera: Aphididae) to the sex pheromone. Bull. Entomol. Res. 2009, 99, 593–602. [Google Scholar] [CrossRef]
- Dickens, J.C. Plant volatiles moderate response to aggregation pheromone in colorado potato beetle. J. Appl. Entomol. 2006, 130, 26–31. [Google Scholar] [CrossRef]
- Ruther, J.; Reinecke, A.; Tolasch, T.; Hilker, M. Mate finding in the forest cockchafer, Melolontha hippocastani, mediated by volatiles from plants and females. Physiol. Entomol. 2001, 25, 172–179. [Google Scholar]
- Reinecke, A.; Ruther, J.; Hilker, M. The scent of food and defence: Green leaf volatiles and toluquinone as sex attractant mediate mate finding in the European cockchafer Melolontha melolontha. Ecol. Lett. 2002, 5, 257–263. [Google Scholar] [CrossRef]
- Byers, J.A.; Zhang, Q.-H.; Birgersson, G. Strategies of a bark beetle, Pityogenes bidentatus, in an olfactory landscape. Naturwissenschaften 2000, 87, 503–507. [Google Scholar] [CrossRef]
- Rochat, D.; Malosse, C.; Lettere, M.; Ducrot, P.H.; Zagatti, P.; Renou, M.; Descoins, C. Male-produced aggregation pheromone of the american palm weevil, Rhynchophorus palmarum (L.), (Coleoptera, Curculionidae): Collection, identification, electrophysiological activity, and laboratory bioassay. J. Chem. Ecol. 1991, 17, 2127–2141. [Google Scholar] [CrossRef]
- Oehlschlager, A.C.; Chinchilla, C.M.; Gonzales, L.M.; Jiron, R.; Mexzon, L.F.; Morgan, B. Development of a pheromone based trapping system for Rhynchophorus palmarum (Coleoptera : Curculionidae). J. Econ. Entomol. 1993, 86, 1382–1392. [Google Scholar]
- Saïd, I.; Renou, M.; Morin, J.-P.; Ferreira, J.M.S.; Rochat, D. Interactions between acetoin, a plant volatile, and pheromone in Rhynchophorus palmarum: Behavioral and olfactory neuron responses. J. Chem. Ecol. 2005, 31, 1789–1805. [Google Scholar] [CrossRef]
- Le Metayer, M.; Pham-Delegue, M.H.; Thiéry, D.; Masson, C. Influence of host- and non-host plant pollen on the calling behaviour of the European sunflower moth Homeosoma nebulellum (Lepidoptera: Pyralidae). Acta Oecol. 1993, 14, 619–626. [Google Scholar]
- McNeil, J.N.; Delisle, J. Host plant pollen influences calling behaviour and ovarian development of the sunflower moth, Homeosoma Electellum. Oecologia 1989, 80, 201–205. [Google Scholar]
- Raina, A.K.; Kingan, T.G.; Matoo, A.K. Chemical signals from host plant and sexual behavior in a moth. Science 1992, 255, 592–594. [Google Scholar]
- Jactel, H.; Birgersson, G.; Andersson, S.; Schlyter, F. Non-host volatiles mediate associational resistance to the pine processionary moth. Oecologia 2011, 166, 703–711. [Google Scholar] [CrossRef]
- Estrada, C.; Gilbert, L.E. Host plants and immatures as mate-searching cues in Heliconius butterflies. Anim. Behav. 2010, 80, 231–239. [Google Scholar] [CrossRef]
- Byers, J.A. Chemical ecology of bark beetles. Experientia 1989, 45, 271–283. [Google Scholar] [CrossRef]
- Emelianov, I.; Dres, M.; Baltensweiler, W.; Mallet, J. Host-induced assortative mating in host races of the larch budmoth. Evolution 2001, 55, 2002–2010. [Google Scholar] [CrossRef]
- Emelianov, I.; Mallet, J.; Baltensweiler, W. Genetic differentiation in Zeiraphera diniana (Lepidoptera: Tortricidae, the larch budmoth): Polymorphism, host races or sibling species. Heredity 1995, 75, 416–424. [Google Scholar] [CrossRef]
- Emelianov, I.; Marec, F.; Mallet, J. Genomic evidence for divergence with gene flow in host races of the larch budmoth. Proc. R. Soc. B Biol. Sci. 2004, 271, 97–105. [Google Scholar] [CrossRef]
- Emelianov, I.; Simpson, F.; Narang, P.; Mallet, J. Host choice promotes reproductive isolation between host races of the larch budmoth Zeiraphera diniana. J. Evol. Biol. 2003, 16, 208–218. [Google Scholar] [CrossRef]
- Witzgall, P.; Bengtsson, M.; Chambon, J.-P.; Priesner, E.; Wildbolz, T.; Arn, H. Sex pheromones of Spilonota ocellana and Spilonota laricana. Entomol. Exp. Appl. 1991, 60, 219–223. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Deisig, N.; Dupuy, F.; Anton, S.; Renou, M. Responses to Pheromones in a Complex Odor World: Sensory Processing and Behavior. Insects 2014, 5, 399-422. https://doi.org/10.3390/insects5020399
Deisig N, Dupuy F, Anton S, Renou M. Responses to Pheromones in a Complex Odor World: Sensory Processing and Behavior. Insects. 2014; 5(2):399-422. https://doi.org/10.3390/insects5020399
Chicago/Turabian StyleDeisig, Nina, Fabienne Dupuy, Sylvia Anton, and Michel Renou. 2014. "Responses to Pheromones in a Complex Odor World: Sensory Processing and Behavior" Insects 5, no. 2: 399-422. https://doi.org/10.3390/insects5020399