
Cold Ambient Temperature Promotes Nosema spp. Intensity in Honey Bees (Apis mellifera)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Nosema spp. Spore Quantification and Species Identification
2.3. Temperature and Honey Bee Flight Days
2.4. Statistical Analyses
3. Results
3.1. Nosema spp. Intensity and Species Identification
3.2. Association of Flight Days with Nosema spp. Intensity
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Evans, J.D.; Schwarz, R.S. Bees brought to their knees: Microbes affecting honey bee health. Trends Microbiol. 2011, 19, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Le Conte, Y.; Ellis, M.; Ritter, W. Varroa mites and honey bee health: Can Varroa explain part of the colony losses? Apidologie 2010, 41, 353–363. [Google Scholar] [CrossRef]
- Van engelsdorp, D.; Meixner, M.D. A historical review of managed honey bee populations in Europe and the United States and the factors that may affect them. J. Invertebr. Pathol. 2010, 103, 80–95. [Google Scholar] [CrossRef] [PubMed]
- Neumann, P.; Carreck, N.L. Honey bee colony losses. J. Apic. Res. 2010, 49, 1–6. [Google Scholar] [CrossRef]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Wassom, D.L.; Dick, T.A.; Arnason, N.; Strickland, D.; Grundmann, A.W. Host genetics: A key factor in regulating the distribution of parasites in natural host populations. J. Parasitol. 1986, 72, 334–337. [Google Scholar] [CrossRef] [PubMed]
- Roulin, A.; Brinkhof, M.W.G.; Bize, P.; Richner, H.; Jungi, T.W.; Bavoux, C.; Boileau, N.; Burneleau, G. Which chick is tasty to parasites? The importance of host immunology vs. parasite life history. J. Anim. Ecol. 2003, 72, 75–81. [Google Scholar] [CrossRef]
- Dunn, A.M.; Hogg, J.C.; Hatcher, M.J. Transmission and burden and the impact of temperature on two species of vertically transmitted microsporidia. Int. J. Parasitol. 2006, 36, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Polley, L.; Hoberg, E.; Kutz, S. Climate change, parasites and shifting boundaries. Acta Vet. Scand. 2010, 52, 1–5. [Google Scholar] [CrossRef]
- Fenoy, S.; Rueda, C.; Higes, M.; Martín-Hernández, R.; del Aguila, C. High-level resistance of Nosema ceranae, a parasite of the honeybee, to temperature and desiccation. Appl. Environ. Microbiol. 2009, 75, 6886–6889. [Google Scholar] [CrossRef] [PubMed]
- Le Conte, Y.; Navajas, M. Climate change: Impact on honey bee populations and diseases. Rev. Sci. Tech. Oie. 2008, 27, 499–510. [Google Scholar]
- Lafferty, K.D.; Kuris, A.M. How environmental stress affects the impacts of parasites. Limnol. Oceanogr. 1999, 44, 925–931. [Google Scholar] [CrossRef]
- Rozsa, L.; Reiczigel, J.; Majoros, G. Quantifying parasites in samples of hosts. J. Parasitol. 2000, 86, 228–232. [Google Scholar] [CrossRef]
- Rosenkranz, P.; Aumeier, P.; Ziegelmann, B. Biology and control of Varroa destructor. J. Invertebr. Pathol. 2010, 103, 96–119. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.L.; Morgan, M.J. Genetic and morphological variation of bee-parasitic Tropilaelaps mites (Acari: Laelapidae): New and re-defined species. Exp. Appl. Acarol. 2007, 43, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Fries, I. Nosema ceranae in European honey bees (Apis mellifera). J. Invertebr. Pathol. 2010, 103, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.P.; Siede, R. Honey bee viruses. Adv. Virus Res. 2007, 70, 33–80. [Google Scholar] [PubMed]
- Fries, I. Nosema apis—A parasite in the honey bee colony. Bee World 1993, 74, 5–19. [Google Scholar] [CrossRef]
- Winston, M.L. The Biology of the Honey Bee; Harvard University Press: Cambridge, MA, USA, 1987; pp. 46–198. [Google Scholar]
- Southwick, E.E. The honey bee cluster as a homeothermic superorganism. Comp. Biochem. Phys. A 1983, 75, 641–645. [Google Scholar] [CrossRef]
- Zander, E. Tierische Parasiten als Krankenheitserreger bei der Biene. Münchener Bienenztg. 1909, 31, 196–204. [Google Scholar]
- Fries, I.; Feng, F.; da Silva, A.; Slemenda, S.B.; Pieniazek, N.J. Nosema ceranae n. sp. (Microspora, Nosematidae), morphological and molecular characterization of a microsporidian parasite of the Asian honey bee Apis cerana (Hymenoptera, Apidae). Eur. J. Protistol. 1996, 32, 356–365. [Google Scholar] [CrossRef]
- Webster, T.C. Nosema apis spore transmission among honey bees. Am. Bee J. 1993, 133, 869–870. [Google Scholar]
- Keeling, P.J.; Fast, N.M. Microsporidia: Biology and evolution of highly reduced intracellular parasites. Annu. Rev. Microbiol. 2002, 56, 93–116. [Google Scholar] [CrossRef] [PubMed]
- Malone, L.A.; Gatehouse, H.S.; Tregidga, E.L. Effects of time, temperature, and honey on Nosema apis (Microsporidia: Nosematidae), a parasite of the honeybee, Apis mellifera (Hymenoptera: Apidae). J. Invertebr. Pathol. 2001, 77, 258–268. [Google Scholar] [CrossRef] [PubMed]
- Martín-Hernández, R.; Meana, A.; García-Palencia, P.; Marín, P.; Botías, C.; Garrido-Bailón, E.; Barrios, L.; Higes, M. Effect of temperature on the biotic potential of honeybee microsporidia. Appl. Environ. Microbiol. 2009, 75, 2554–2557. [Google Scholar] [CrossRef] [PubMed]
- Dainat, B.; van Engelsdorp, D.; Neumann, P. Colony collapse disorder in Europe. Environ. Microbiol. Rep. 2012, 4, 123–125. [Google Scholar] [CrossRef] [PubMed]
- Dainat, B.; Evans, J.D.; Chen, Y.P.; Gauthier, L.; Neumann, P. Predictive markers of honey bee colony collapse. PLoS ONE 2012, 7, e32151. [Google Scholar] [CrossRef] [PubMed]
- Martín-Hernández, R.; Meana, A.; Prieto, L.; Salvador, A.M.; Garrido-Bailón, E.; Higes, M. Outcome of colonization of Apis mellifera by Nosema ceranae. Appl. Environ. Microbiol. 2007, 73, 6331–6338. [Google Scholar] [CrossRef] [PubMed]
- Borchert, A. Beiträge zur Kenntnis des Bienenparasiten Nosema apis. Arch. Für Bienenkd. 1928, 9, 115–178. [Google Scholar]
- Bailey, L. The epidemiology and control of Nosema disease of the honey bee. Ann. Appl. Biol. 1955, 43, 379–389. [Google Scholar] [CrossRef]
- Traver, B.E.; Williams, M.R.; Fell, R.D. Comparison of within hive sampling and seasonal activity of Nosema ceranae in honey bee colonies. J. Invertebr. Pathol. 2012, 109, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.W.; Chung, W.P.; Wang, C.H.; Softer, L.F.; Huang, W.F. Nosema ceranae infection intensity highly correlates with temperature. J. Invertebr. Pathol. 2012, 111, 264–267. [Google Scholar] [CrossRef] [PubMed]
- Rangel, J.; Baum, K.; Rubink, W.L.; Coulson, R.N.; Johnston, J.S.; Traver, B.E. Prevalence of Nosema species in a feral honey bee population: A 20-year survey. Apidologie 2016, 47, 561–571. [Google Scholar] [CrossRef]
- Burrill, R.M.; Dietz, A. The response of honey bees to variations in solar radiation and temperature. Apidologie 1981, 12, 319–328. [Google Scholar] [CrossRef]
- Hertig, M. The normal and pathological histology of the ventriculus of the honey-bee, with special reference to infection with Nosema apis. J. Parasitol. 1923, 9, 109–140. [Google Scholar] [CrossRef]
- Lotmar, R. Über den Einfluss der Temperatur auf den Parasiten Nosema apis. Beih. Schweiz. Bienenztg. 1944, 67, 17–19. [Google Scholar]
- Bailey, L.; Ball, B.V. Honey Bee Pathology, 2nd ed.; Academic Press Inc.: San Diego, CA, USA, 1991; pp. 64–72. [Google Scholar]
- Fries, I.; Ekbohm, G.; Villumstad, E. Nosema apis, sampling techniques and honey yield. J. Apic. Res. 1984, 23, 102–105. [Google Scholar] [CrossRef]
- Pirk, C.W.W.; de Miranda, J.R.; Kramer, M.; Murray, T.E.; Nazzi, F.; Shutler, D.; van der Steen, J.J.M.; van Dooremalen, C. Statistical guidelines for Apis mellifera research. J. Apicult. Res. 2013. [Google Scholar] [CrossRef]
- Cantwell, G.E. Standard methods for counting Nosema spores. Am. Bee J. 1970, 110, 222–223. [Google Scholar]
- Fries, I.; Chauzat, M.P.; Chen, Y.P.; Doublet, V.; Genersch, E.; Gisder, S.; Higes, M.; McMahon, D.P.; Martín-Hernández, R.; Natsopoulou, M.; et al. Standard methods for Nosema research. J. Apic. Res. 2013. [Google Scholar] [CrossRef]
- Van engelsdorp, D.; Evans, J.D.; Saegerman, C.; Mullin, C.; Haubruge, E.; Nguyen, B.K.; Frazier, M.; Frazier, J.; Cox-Foster, D.; Chen, Y.P.; et al. Colony collapse disorder: A descriptive study. PLoS ONE 2009, 4, e6481. [Google Scholar]
- Gisder, S.; Hedtke, K.; Mockel, N.; Frielitz, M.C.; Linde, A.; Genersch, E. Five-year cohort study of Nosema spp. in Germany: Does climate shape virulence and assertiveness of Nosema ceranae? Appl. Environ. Microbiol. 2010, 76, 3032–3038. [Google Scholar] [CrossRef] [PubMed]
- Higes, M.; García-Palencia, P.; Martín-Hernández, R.; Meana, A. Experimental infection of Apis mellifera honeybees with Nosema ceranae (Microsporidia). J. Invertebr. Pathol. 2007, 94, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Paxton, R.J.; Klee, J.; Korpela, S.; Fries, I. Nosema ceranae has infected Apis mellifera in Europe since at least 1998 and may be more virulent than Nosema apis. Apidologie 2007, 38, 558–565. [Google Scholar] [CrossRef]
- Aufauvre, J.; Biron, D.G.; Vidau, C.; Fontbonne, R.; Roudel, M.; Diogon, M.; Viguès, B.; Belzunces, L.P.; Delbac, F.; Blot, N. Parasite-insecticide interactions: A case study of Nosema ceranae and fipronil synergy on honeybee. Sci. Rep. UK 2012. [Google Scholar] [CrossRef] [PubMed]
- Bourgeois, A.L.; Rinderer, T.E.; Beaman, L.D.; Danka, R.G. Genetic detection and quantification of Nosema apis and N. ceranae in the honey bee. J. Invertebr. Pathol. 2010, 103, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Traver, B.E.; Fell, R.D. Prevalence and infection intensity of Nosema in honey bee (Apis mellifera L.) colonies in Virginia. J. Invertebr. Pathol. 2011, 107, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Giersch, T.; Berg, T.; Galea, F.; Hornitzky, M. Nosema ceranae infects honey bees (Apis mellifera) and contaminates honey in Australia. Apidologie 2009, 40, 117–123. [Google Scholar] [CrossRef]
- Yoshiyama, M.; Kimura, K. Distribution of Nosema ceranae in the European honeybee, Apis mellifera in Japan. J. Invertebr. Pathol. 2011, 106, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Cox-Foster, D.L.; Conlan, S.; Holmes, E.C.; Palacios, G.; Evans, J.D.; Moran, N.A.; Quan, P.L.; Briese, T.; Hornig, M.; Geiser, D.M.; et al. A metagenomic survey of microbes in honey bee colony collapse disorder. Science 2007, 318, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Evans, J.D.; Zhou, L.; Boncristiani, H.; Kimura, K.; Xiao, T.; Litkowski, A.M.; Pettis, J.S. Asymmetrical coexistence of Nosema ceranae and Nosema apis in honey bees. J. Invertebr. Pathol. 2009, 101, 204–209. [Google Scholar] [CrossRef] [PubMed]
- Chaimanee, V.; Warrit, N.; Chantawannakul, P. Infections of Nosema ceranae in four different honeybee species. J. Invertebr. Pathol. 2010, 105, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Medici, S.K.; Sarlo, E.G.; Porrini, M.P.; Braunstein, M.; Eguaras, M.J. Genetic variation and widespread dispersal of Nosema ceranae in Apis mellifera apiaries from Argentina. Parasitol. Res. 2012, 110, 859–864. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.P.; Evans, J.D.; Murphy, C.; Gutell, R.; Zuker, M.; Gundensen-Rindal, D.; Pettis, J.S. Morphological, molecular, and phylogenetic characterization of Nosema ceranae, a microsporidian parasite isolated from the European honey bee, Apis mellifera. J. Eukaryot. Microbiol. 2009, 56, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Forsgren, E.; Fries, I. Comparative virulence of Nosema ceranae and Nosema apis in individual European honey bees. Vet. Parasitol. 2010, 170, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.F.; Solter, L.F. Comparative development and tissue tropism of Nosema apis and Nosema ceranae. J. Invertebr. Pathol. 2013, 113, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Moeller, F. Nosema Disease—Its Control in Honey Bee Colonies; U.S. Department of Agriculture Technical Bulletin: Washington, DC, USA, 1978.
- Retschnig, G.; Williams, G.R.; Odemer, R.; Boltin, J.; Di Poto, C.; Mehmann, M.M.; Retschnig, P.; Winiger, P.; Rosenkranz, P.; Neumann, P. Effects, but no interactions, of ubiquitous pesticide and parasite stressors on honey bee (Apis mellifera) lifespan and behaviour in a colony environment. Environ. Microbiol. 2015, 17, 4322–4331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desai, S.D.; Currie, R.W. Effects of wintering environment and parasite–pathogen interactions on honey bee colony loss in North Temperate regions. PLoS ONE 2016, 11, e0159615. [Google Scholar] [CrossRef] [PubMed]
- Moeller, F.E. Effects of emerging bees and of winter flights on Nosema disease in honeybee colonies. J. Apic. Res. 1972, 11, 117–120. [Google Scholar] [CrossRef]
- Lotmar, R. Bestehen Beziehungen zwischen der Witterung und dem seuchenhaften Auftreten der Frühjahrsschwindsucht (Nosema-Amöben-Seuche)? Schweiz. Bienenztg. 1943, 66, 68–80. [Google Scholar]
- Williams, G.R.; Shutler, D.; Rogers, R.E.L. Effects at Nearctic north-temperate latitudes of indoor versus outdoor overwintering on the microsporidium Nosema ceranae and western honey bees (Apis mellifera). J. Invertebr. Pathol. 2010, 104, 4–7. [Google Scholar] [CrossRef] [PubMed]
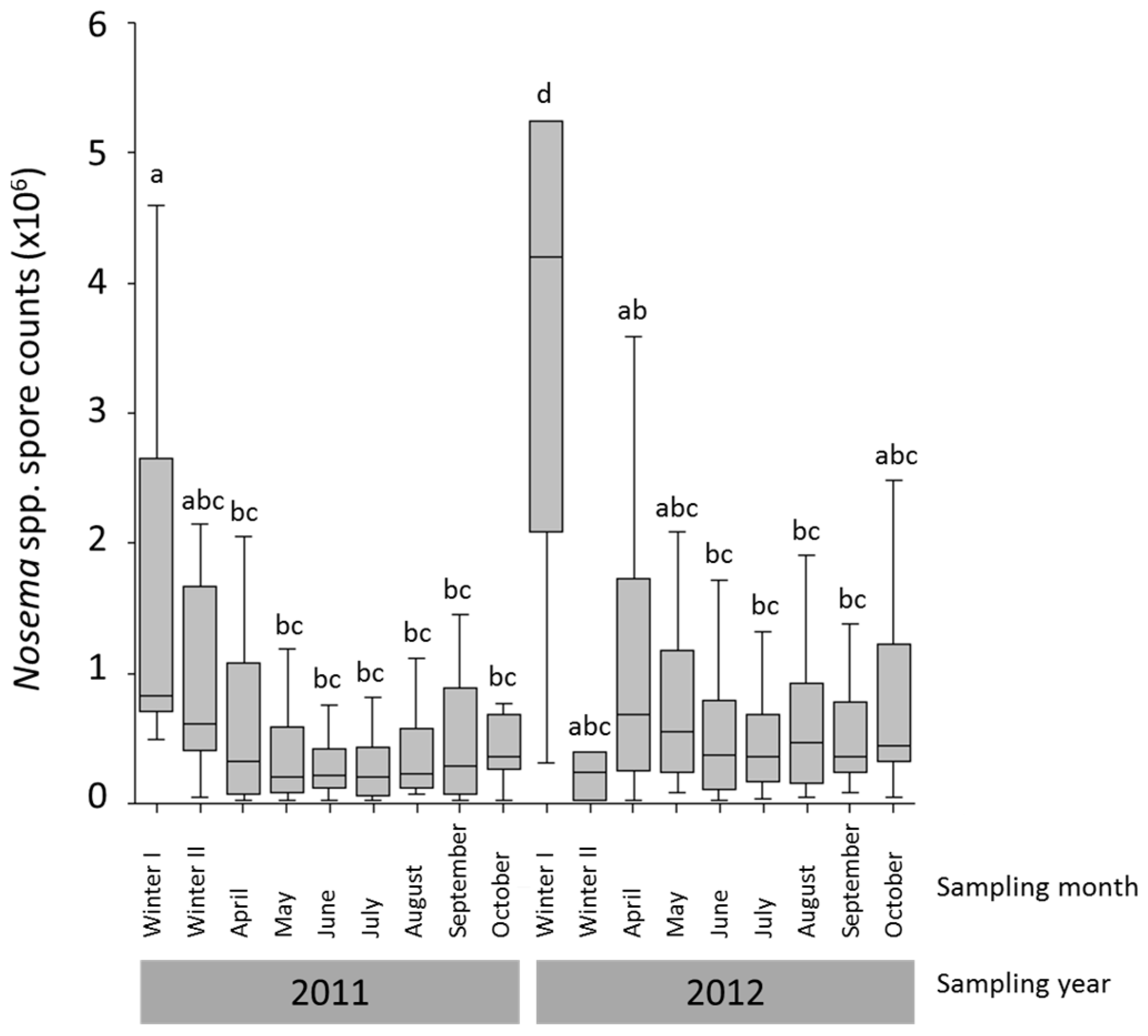



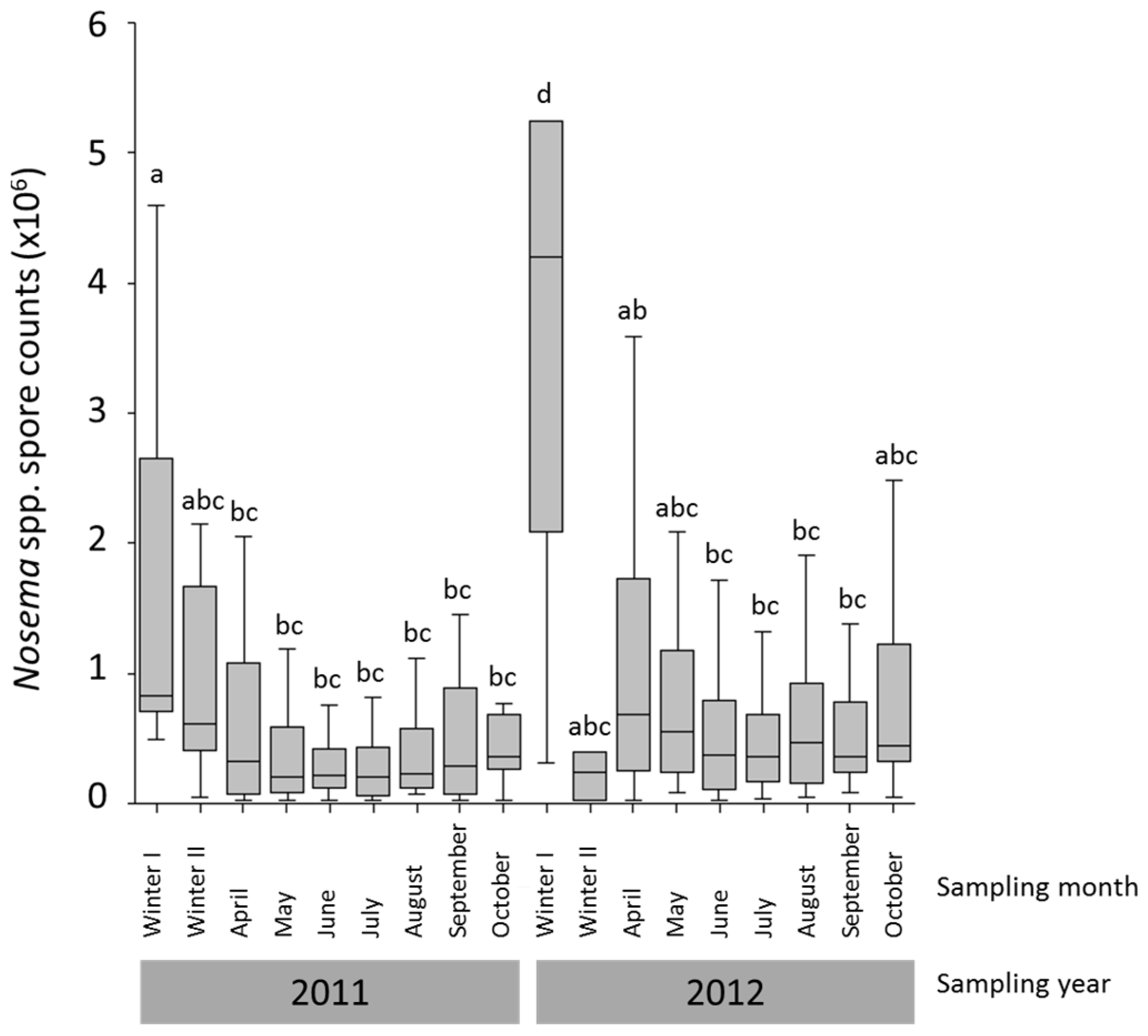{kind=link}
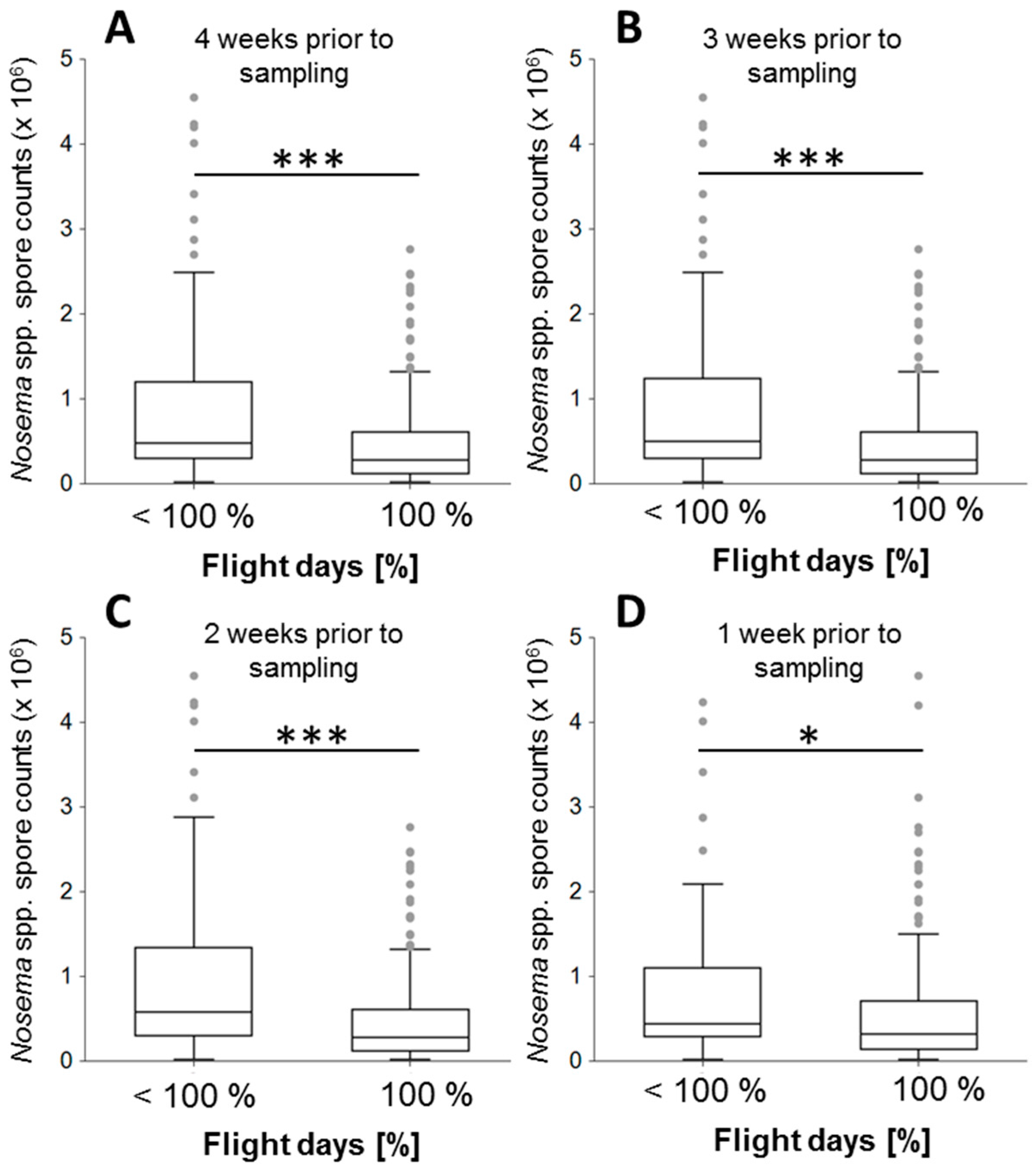{kind=link}
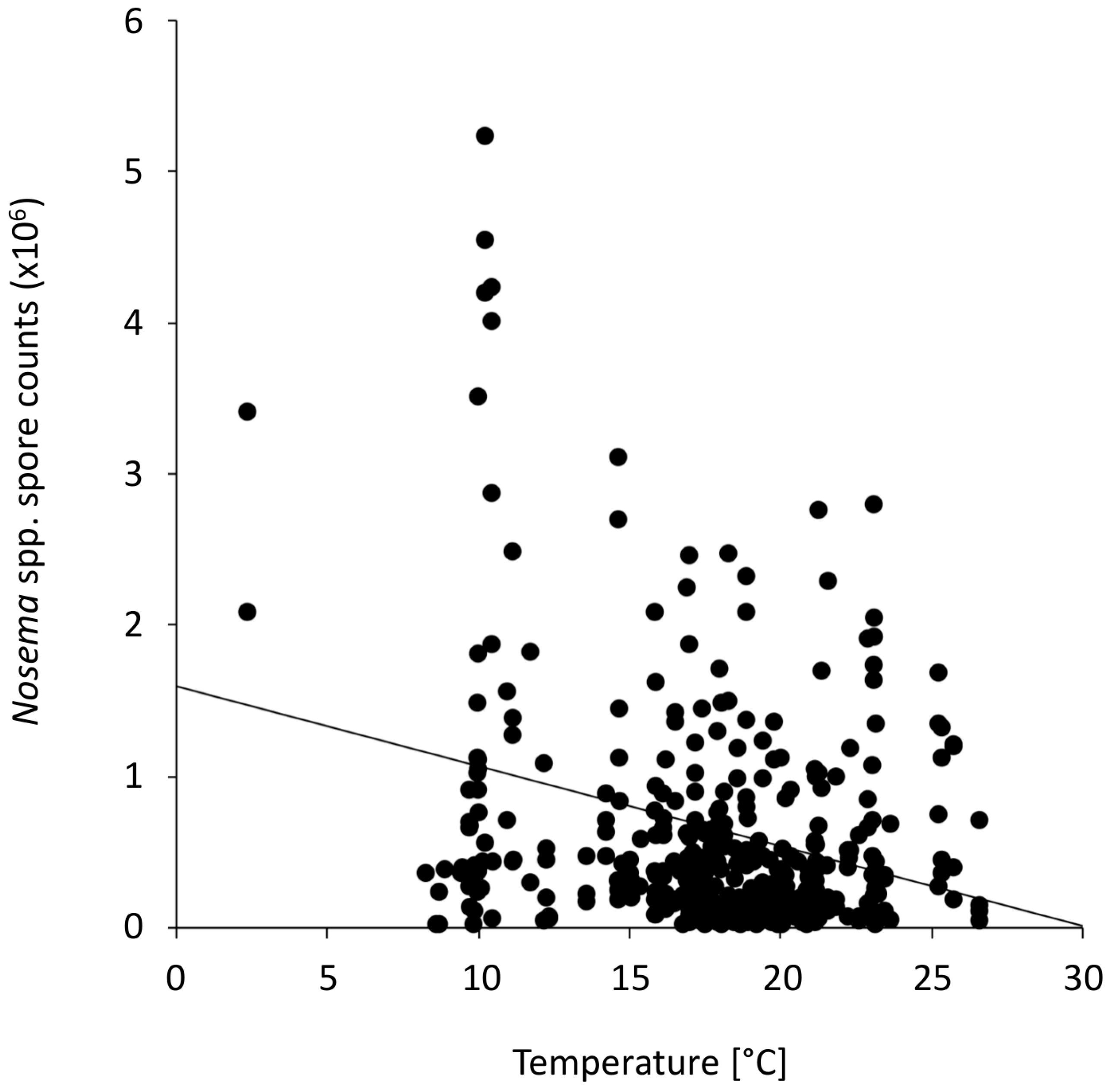{kind=link}
| Group | Time Interval | Median Nosema spp. Intensities (×106) | N | p-Value |
|---|---|---|---|---|
| Colony as the Experimental Unit | ||||
| 100% vs. <100% | 4 weeks b.s. 1 | 0.28, 0.48 | 240, 101 | 0.000002 *** |
| 3 weeks b.s. 1 | 0.28, 0.50 | 241, 100 | 0.000002 *** | |
| 2 weeks b.s. 1 | 0.28, 0.58 | 245, 96 | 0.000000 *** | |
| 1 weeks b.s. 1 | 0.33, 0.44 | 287, 54 | 0.022882 * | |
| ≥90% vs. <90% | 4 weeks b.s. 1 | 0.31, 0.44 | 279, 62 | 0.000564 *** |
| 3 weeks b.s. 1 | 0.31, 0.44 | 275, 66 | 0.000251 *** | |
| 2 weeks b.s. 1 | 0.30, 0.48 | 264, 77 | 0.00005 *** | |
| 1 weeks b.s. 1 | 0.33, 0.44 | 293, 48 | 0.027709 * | |
| ≥75% vs. <75% | 4 weeks b.s. 1 | 0.31, 0.44 | 285, 56 | 0.000629 *** |
| 3 weeks b.s. 1 | 0.31, 0.45 | 284, 57 | 0.000565 *** | |
| 2 weeks b.s. 1 | 0.31, 0.44 | 292, 49 | 0.009982 ** | |
| 1 weeks b.s. 1 | 0.34, 0.44 | 299, 42 | 0.032027 * | |
| ≥50% vs. <50% | 4 weeks b.s. 1 | 0.36, 0.34 | 333, 8 | 0.489272 |
| 3 weeks b.s. 1 | 0.36, 0.36 | 322, 19 | 0.683875 | |
| 2 weeks b.s. 1 | 0.34, 0.45 | 320, 21 | 0.09639 | |
| 1 weeks b.s. 1 | 0.34, 0.44 | 317, 24 | 0.065977 | |
| Apiary as the Experimental Unit | ||||
| 100% vs. <100% | 4 weeks b.s. 1 | 0.29, 0.49 | 97, 47 | 0.00052 *** |
| 3 weeks b.s. 1 | 0.31, 0.53 | 98, 46 | 0.000472 *** | |
| 2 weeks b.s. 1 | 0.31, 0.57 | 100, 44 | 0.000157 *** | |
| 1 weeks b.s. 1 | 0.37, 0.43 | 116, 28 | 0.205144 | |
| Correlation | Time Interval | N | Spearman Rank Correlation Coefficient | p-Value |
|---|---|---|---|---|
| Nosema spp. and temperature | whole study period | 383 | −0.1788 | 0.0004 *** |
| Nosema spp. and flight day prop. 1 | 4 weeks b.s. 2 | 341 | −0.2469 | 3.96 × 10−6 *** |
| Nosema spp. and flight day prop. 1 | 3 weeks b.s. 2 | 341 | −0.2485 | 3.4 × 10−6 *** |
| Nosema spp. and flight day prop. 1 | 2 weeks b.s. 2 | 341 | −0.2626 | 8.76 × 10−7 *** |
| Nosema spp. and flight day prop. 1 | 1 week b.s. 2 | 341 | −0.1282 | 0.0179 * |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Retschnig, G.; Williams, G.R.; Schneeberger, A.; Neumann, P. Cold Ambient Temperature Promotes Nosema spp. Intensity in Honey Bees (Apis mellifera). Insects 2017, 8, 20. https://doi.org/10.3390/insects8010020
Retschnig G, Williams GR, Schneeberger A, Neumann P. Cold Ambient Temperature Promotes Nosema spp. Intensity in Honey Bees (Apis mellifera). Insects. 2017; 8(1):20. https://doi.org/10.3390/insects8010020
Chicago/Turabian StyleRetschnig, Gina, Geoffrey R. Williams, Annette Schneeberger, and Peter Neumann. 2017. "Cold Ambient Temperature Promotes Nosema spp. Intensity in Honey Bees (Apis mellifera)" Insects 8, no. 1: 20. https://doi.org/10.3390/insects8010020