
Impact of Consuming ‘Toxic’ Monarch Caterpillars on Adult Chinese Mantid Mass Gain and Fecundity

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Rearing and Maintenance
2.2. Experimental Design
2.3. Statistical Analyses
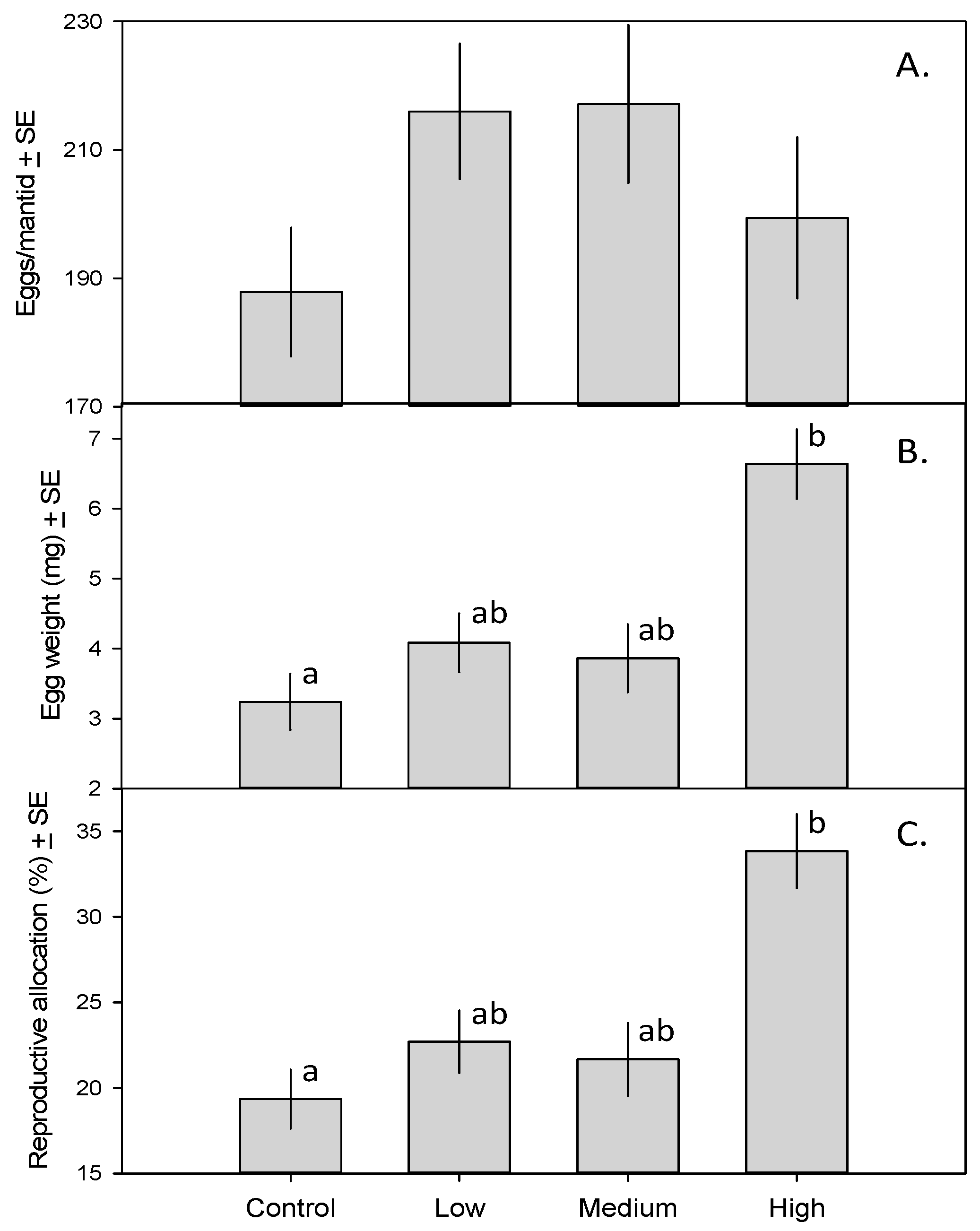
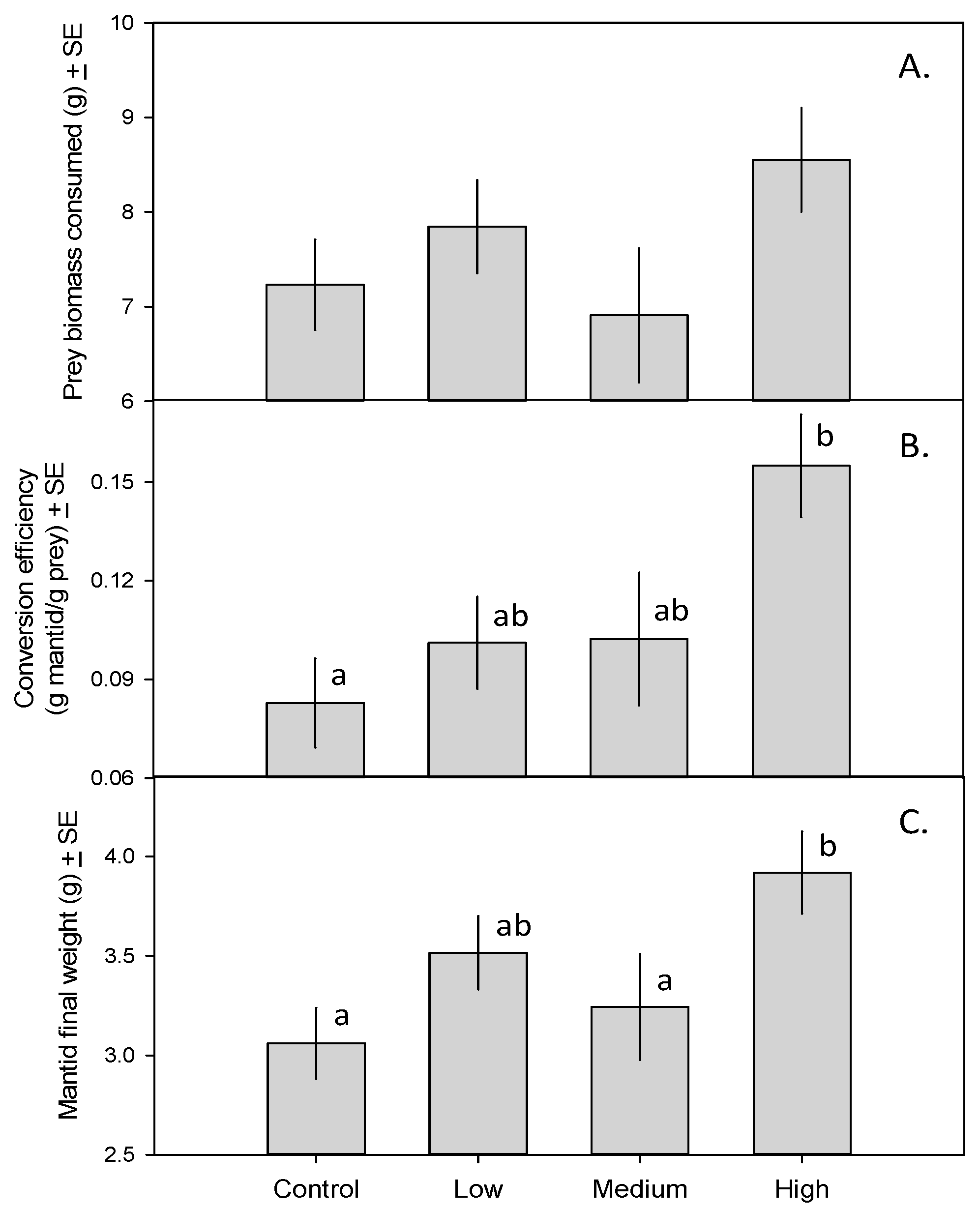
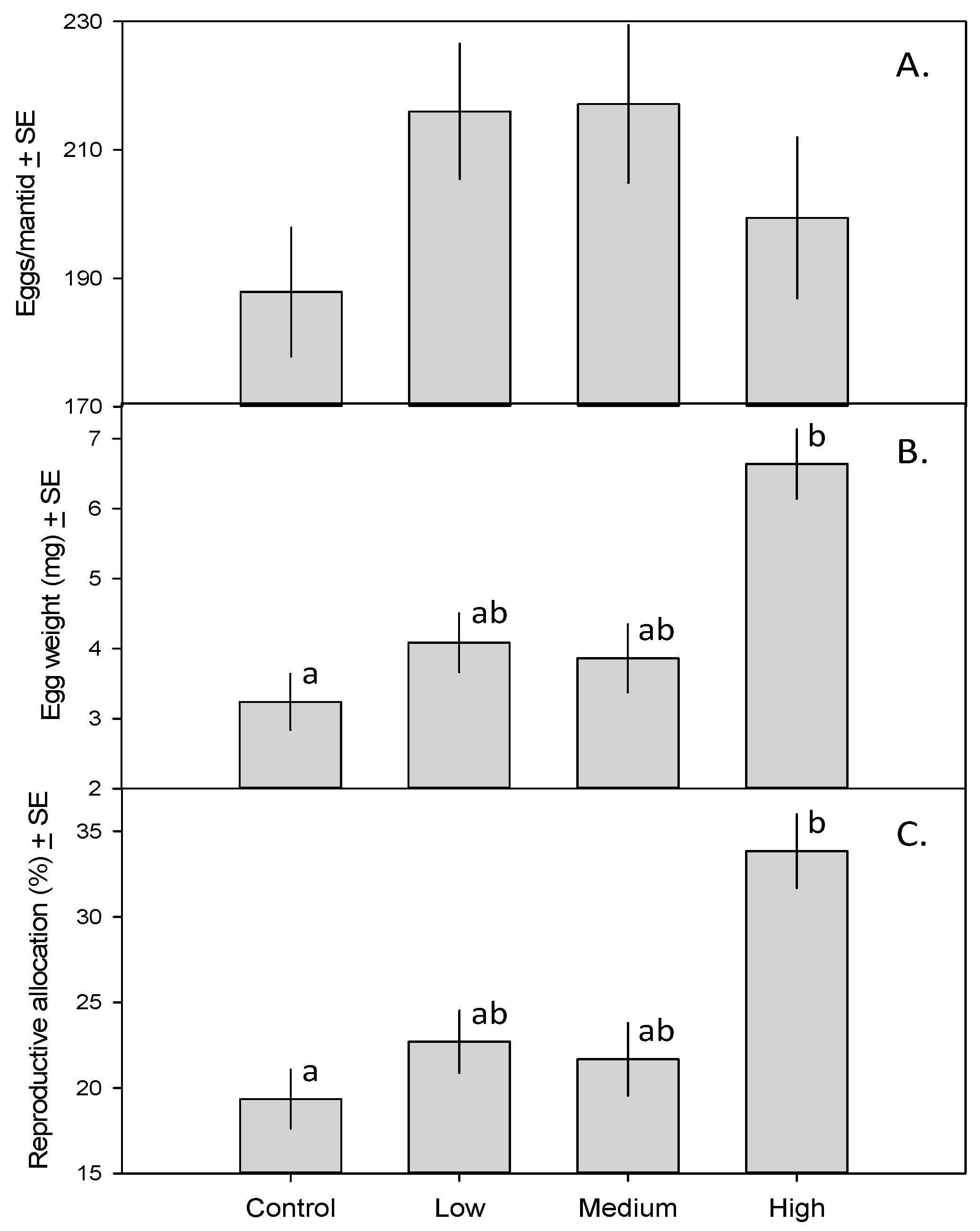
3. Results
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Duffey, S.S. Sequestration of plant natural products by insects. Annu. Rev. Entomol. 1980, 25, 447–477. [Google Scholar] [CrossRef]
- Nishida, R. Sequestration of defensive substances from plants by Lepidoptera. Annu. Rev. Entomol. 2002, 47, 57–92. [Google Scholar] [CrossRef] [PubMed]
- Ruxton, G.D.; Sherratt, T.N.; Speed, M.P. Avoiding Attack: The Evolutionary Ecology of Crypsis, Warning Signals, and Mimicry; Oxford University Press: Oxford, UK, 2004; Volume 249. [Google Scholar]
- Gittleman, J.L.; Harvey, P.H. Why are distasteful prey not cryptic? Nature 1980, 286, 149–150. [Google Scholar] [CrossRef]
- Sloggett, J.J.; Davis, A.J. Eating chemically defended prey: Alkaloid metabolism in an invasive ladybird predator of other ladybirds (Coleoptera: Coccinellidae). J. Exp. Biol. 2010, 213, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, T.; Häggström, H.; Theuring, C.; Lindigkeit, R.; Rahier, M. Detoxification of pyrrolizidine alkaloids by the harvestman Mitopus morio (Phalangidae) a predator of alkaloid defended leaf beetles. Chemoecology 2003, 13, 123–127. [Google Scholar] [CrossRef]
- Brown, K.S.J.; Neto, J.V. Predation on aposematic Ithomiine butterflies by tanagers, Pipraeidea melanonota. Biotropica 1976, 8, 136–141. [Google Scholar] [CrossRef]
- Glendinning, J.I. How do predators cope with chemically defended foods? Biol. Bull. 2007, 213, 252–266. [Google Scholar] [CrossRef] [PubMed]
- Rayor, L.S.; Mooney, L.J.; Renwick, J.A. Predatory behavior of Polistes dominulus wasps in response to cardenolides and glucosinolates in Pieris napi caterpillars. J. Chem. Ecol. 2007, 33, 1177–1185. [Google Scholar] [CrossRef] [PubMed]
- Skelhorn, J.; Rowe, C. Predators’ toxin burdens influence their strategic decisions to eat toxic prey. Curr. Biol. 2007, 17, 1479–1483. [Google Scholar] [CrossRef] [PubMed]
- Malcolm, S. Disruption of web structure and predatory behavior of a spider by plant-derived chemical defenses of an aposematic aphid. J. Chem. Ecol. 1989, 15, 1699–1716. [Google Scholar] [CrossRef] [PubMed]
- Francis, F. Effect of aphid host plant on development and reproduction of the third trophic level, the predator Adalia bipunctata (Coleoptera: Coccinellidae). Environ. Entomol. 2001, 30, 947–952. [Google Scholar] [CrossRef]
- Cox, D.J.; Preisser, E.; University of Rhode Island, Kingston, RI, USA. Personal observations, 2009.
- Agrawal, A.A.; Petschenka, G.; Bingham, R.A.; Weber, M.G.; Rasmann, S. Toxic cardenolides: Chemical ecology and coevolution of specialized plant–herbivore interactions. New Phytol. 2012, 194, 28–45. [Google Scholar] [CrossRef] [PubMed]
- Rafter, J.L.; Agrawal, A.A.; Preisser, E.L. Chinese mantids gut toxic monarch caterpillars: Avoidance of prey defence? Ecol. Entomol. 2013, 38, 76–82. [Google Scholar] [CrossRef]
- Rafter, J.L.; Vendettuoli, J.F.; Gonda-King, L.; Preisser, E.L. Pretty picky for a generalist: Impacts of toxicity and nutritional quality on mantid prey processing. Environ. Entomol. 2017, in press. [Google Scholar]
- Paradise, C.; Stamp, N. Variable quantities of toxic diet cause different degrees of compensatory and inhibitory responses by juvenile praying mantids. Entomol. Exp. Appl. 1990, 55, 213–222. [Google Scholar] [CrossRef]
- Paradise, C.; Stamp, N. Episodes of unpalatable prey reduce consumption and growth of juvenile praying mantids. J. Insect Behav. 1993, 6, 155–166. [Google Scholar] [CrossRef]
- Beukeboom, L.W.; Zwaan, B.J. Genetics. In Insects as Natural Enemies: A Practical Perspective; Jervis, M.A., Ed.; Springer: Dordrecht, The Netherlands, 2005; pp. 167–218. [Google Scholar]
- Rasmann, S.; Agrawal, A.A. Latitudinal patterns in plant defense: Evolution of cardenolides, their toxicity and induction following herbivory. Ecol. Lett. 2011, 14, 476–483. [Google Scholar] [CrossRef] [PubMed]
- Nachappa, P. Ecological Consequences of Genetic Variation in Foraging Behaviors of a Predatory Mite; Kansas State University: Manhattan, KS, USA, 2008. [Google Scholar]
- Rowson, J.M. Studies in the genus Digitalis part I. The colorimetric estimation of digitoxin and of preparations of Digitalis purpurea. J. Pharm. Pharmacol. 1952, 4, 814–830. [Google Scholar] [CrossRef] [PubMed]
- Dobler, S.; Rowell-Rahier, M. Production of cardenolides versus sequestration of pyrrolizidine alkaloids in larvae of Oreina species (Coleoptera, Chrysomelidae). J. Chem. Ecol. 1994, 20, 555–568. [Google Scholar] [CrossRef] [PubMed]
- Brower, L.P.; Seiber, J.N.; Nelson, C.J.; Lynch, S.P.; Hoggard, M.P.; Cohen, J.A. Plant-determined variation in cardenolide content and thin-layer chromatography profiles of monarch butterflies, Danaus plexippus reared on milkweed plants in California. J. Chem. Ecol. 1984, 10, 1823–1857. [Google Scholar] [CrossRef] [PubMed]
- Eisenberg, R.M.; Hurd, L.E.; Bartley, J.A. Ecological consequences of food limitation for adult mantids (Tenodera ardifolia sinensis saussure). Am. Midl. Nat. 1981, 106, 209–218. [Google Scholar] [CrossRef]
- Harshman, L.G.; Zera, A.J. The cost of reproduction: The devil in the details. Trends Ecol. Evol. 2007, 22, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Lefcheck, J.S.; Whalen, M.A.; Davenport, T.M.; Stone, J.P.; Duffy, J.E. Physiological effects of diet mixing on consumer fitness: A meta-analysis. Ecology 2013, 94, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Beckman, N.; Hurd, L.E. Pollen feeding and fitness in praying mantids: The vegetarian side of a tritrophic predator. Environ. Entomol. 2003, 32, 881–885. [Google Scholar] [CrossRef]
- Bilde, T.; Toft, S. Prey preference and egg production of the carabid beetle Agonum dorsale. Entomol. Exp. Appl. 1994, 73, 151–156. [Google Scholar] [CrossRef]
- Berenbaum, M.R.; Miliczky, E. Mantids and milkweed bugs: Efficacy of aposematic coloration against invertebrate predators. Am. Midl. Nat. 1984, 111, 64–68. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Treatment Group | n | Treatment Description |
|---|---|---|
| Control | 9 | Offered two crickets daily from day 4 to day 35 |
| Low Toxicity | 8 | Offered one monarch caterpillar on day 11. Offered two crickets per day all other days until day 35 |
| Medium Toxicity | 7 | Offered one monarch on days 6, 9, 12, 15, and 18. Offered two crickets per day all other days |
| High Toxicity | 7 | Offered one monarch caterpillar each day on days 4–19. Subsequently, offered two crickets per day until day 35 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rafter, J.L.; Gonda-King, L.; Niesen, D.; Seeram, N.P.; Rigsby, C.M.; Preisser, E.L. Impact of Consuming ‘Toxic’ Monarch Caterpillars on Adult Chinese Mantid Mass Gain and Fecundity. Insects 2017, 8, 23. https://doi.org/10.3390/insects8010023
Rafter JL, Gonda-King L, Niesen D, Seeram NP, Rigsby CM, Preisser EL. Impact of Consuming ‘Toxic’ Monarch Caterpillars on Adult Chinese Mantid Mass Gain and Fecundity. Insects. 2017; 8(1):23. https://doi.org/10.3390/insects8010023
Chicago/Turabian StyleRafter, Jamie L., Liahna Gonda-King, Daniel Niesen, Navindra P. Seeram, Chad M. Rigsby, and Evan L. Preisser. 2017. "Impact of Consuming ‘Toxic’ Monarch Caterpillars on Adult Chinese Mantid Mass Gain and Fecundity" Insects 8, no. 1: 23. https://doi.org/10.3390/insects8010023