
Influence of Varroa Mite (Varroa destructor) Management Practices on Insecticide Sensitivity in the Honey Bee (Apis mellifera)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Seasonal Management Experiments
2.2. Pesticides
2.3. Bioassays
2.4. Statistical Analyses
3. Results
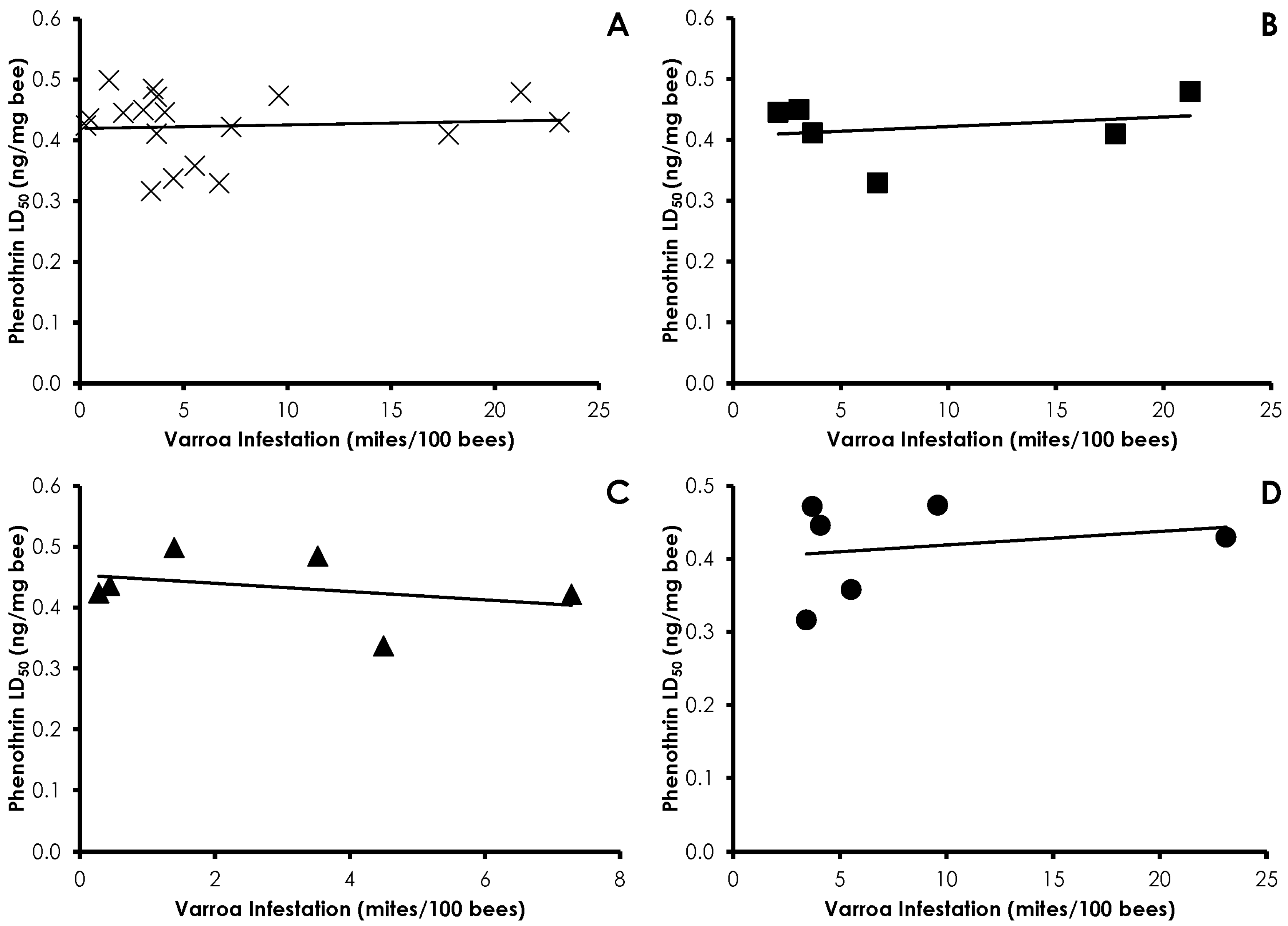
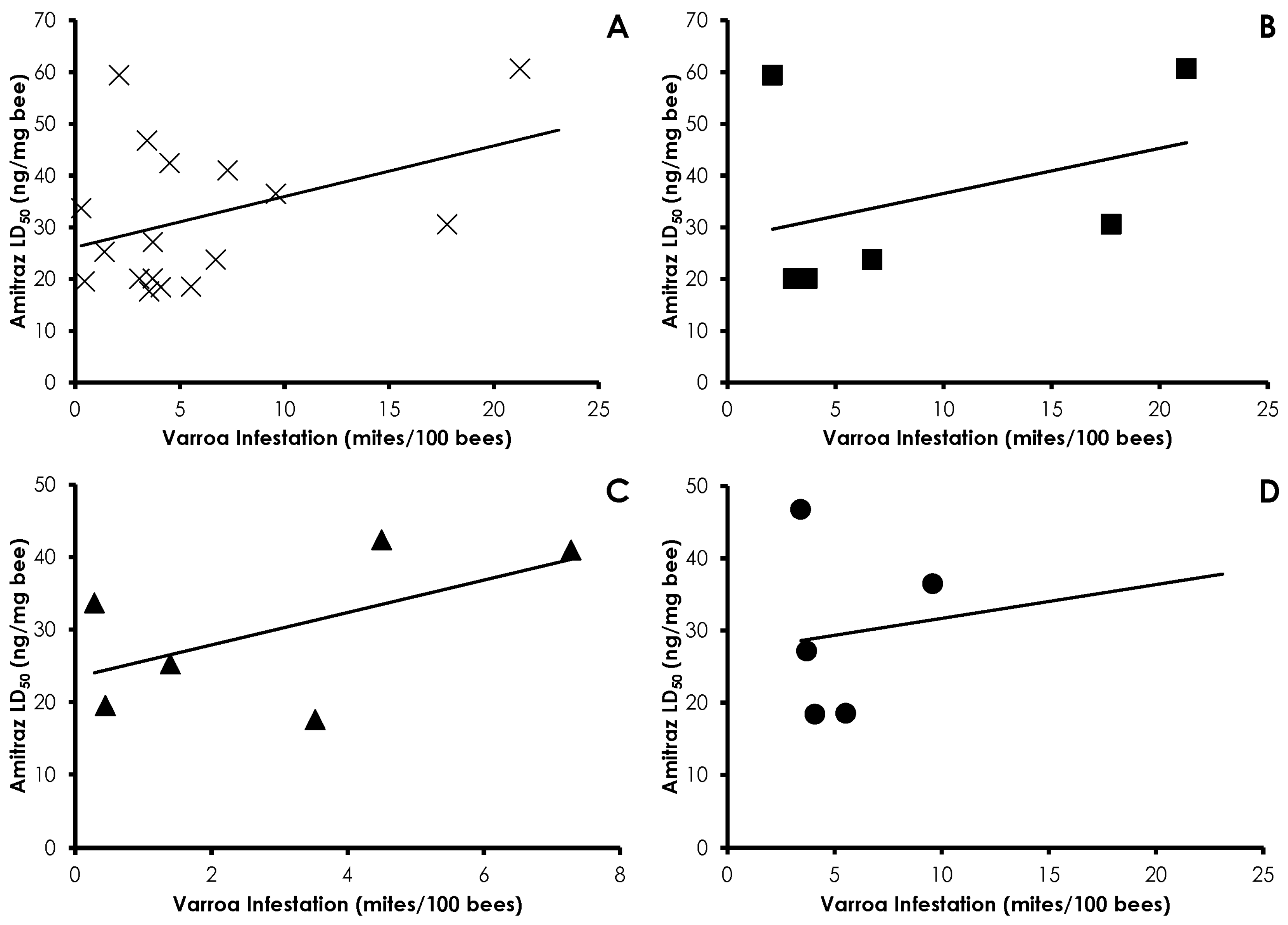
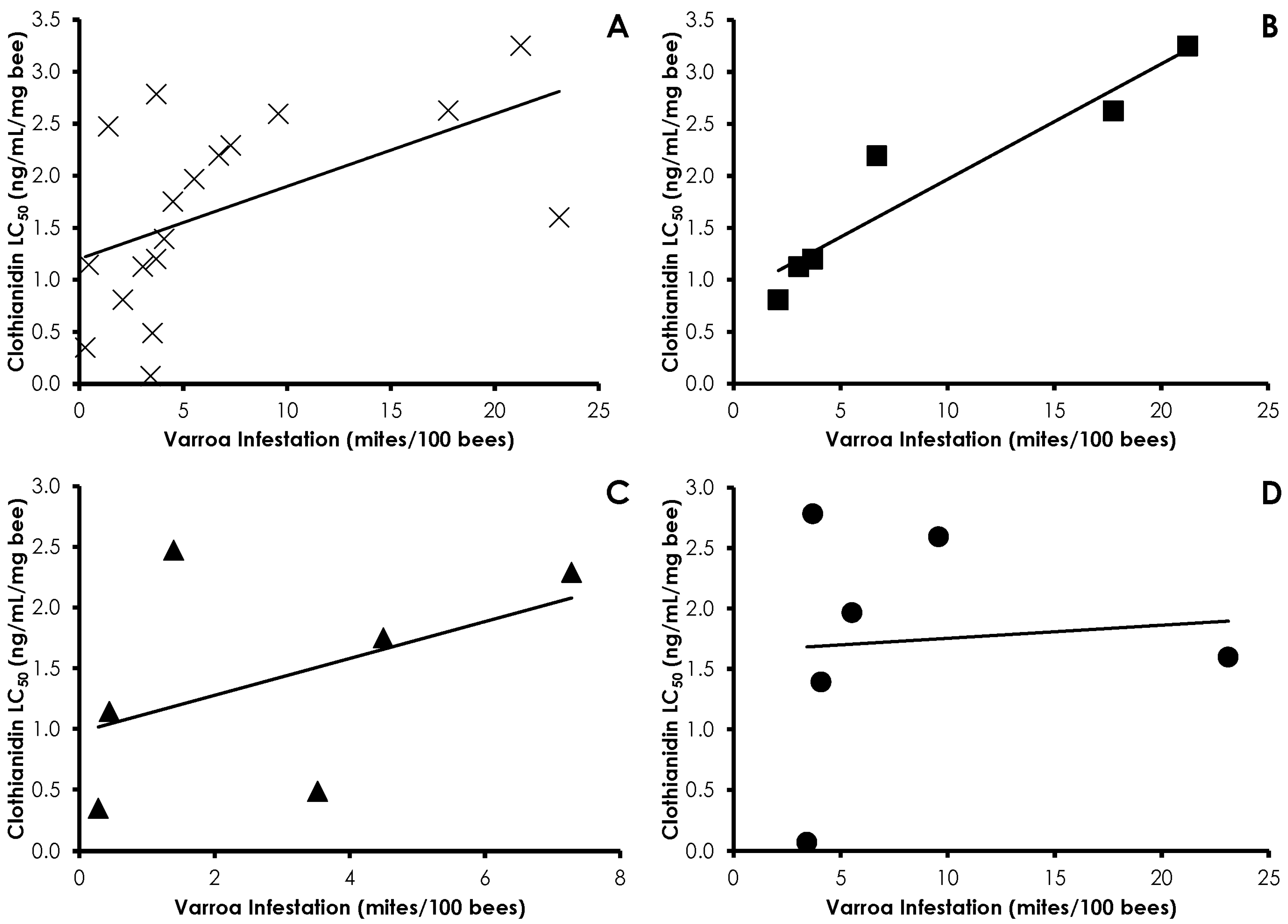
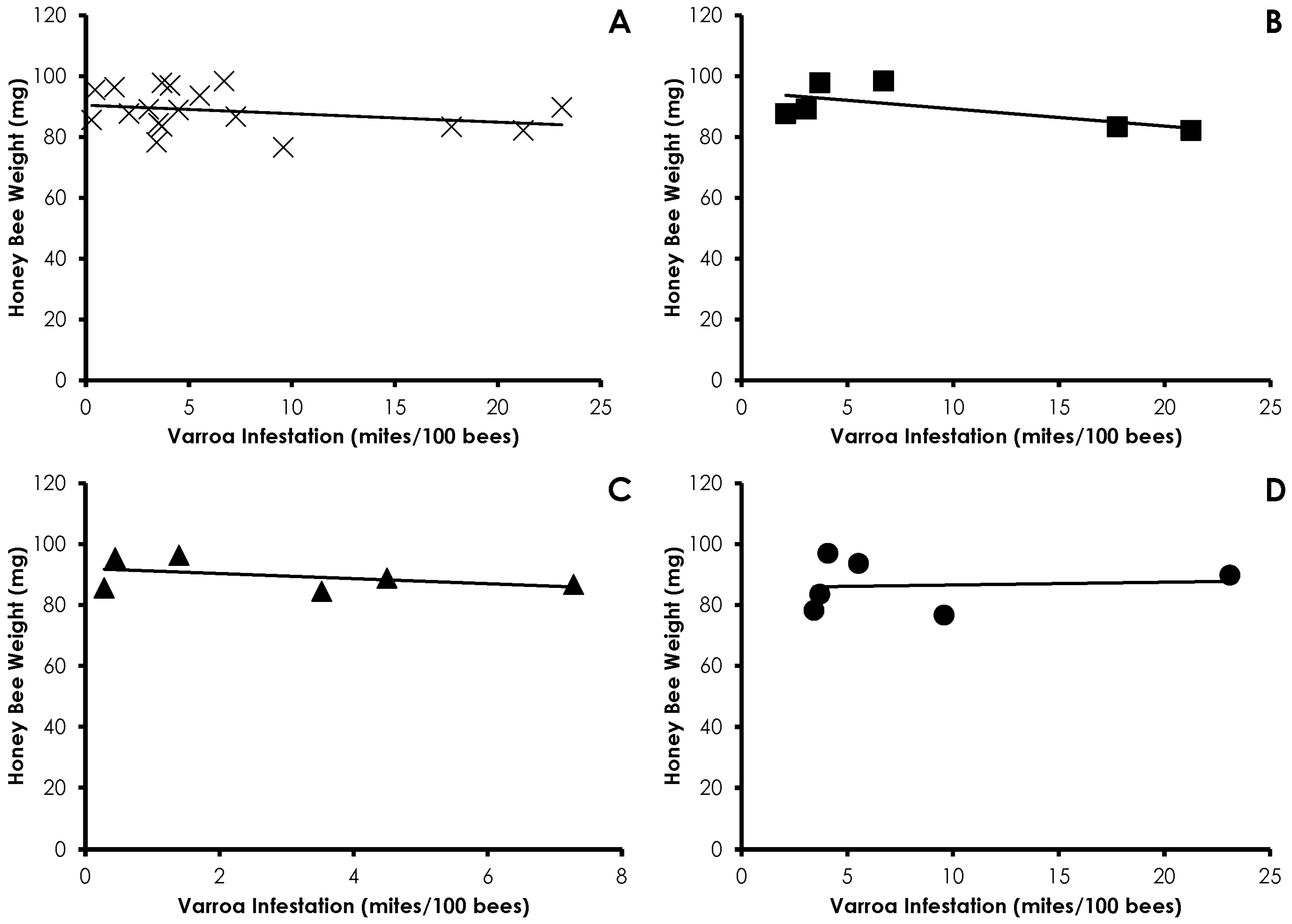
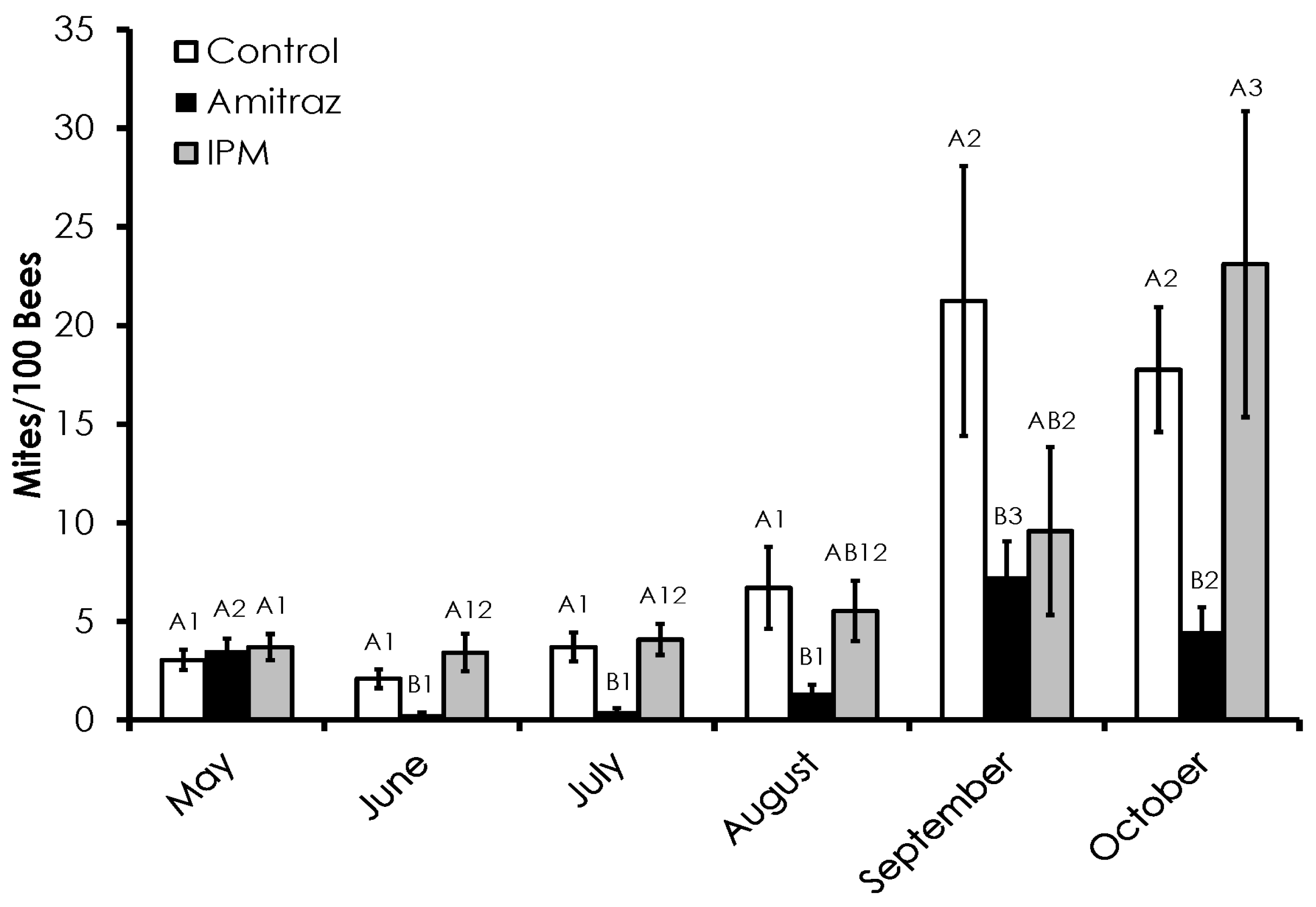
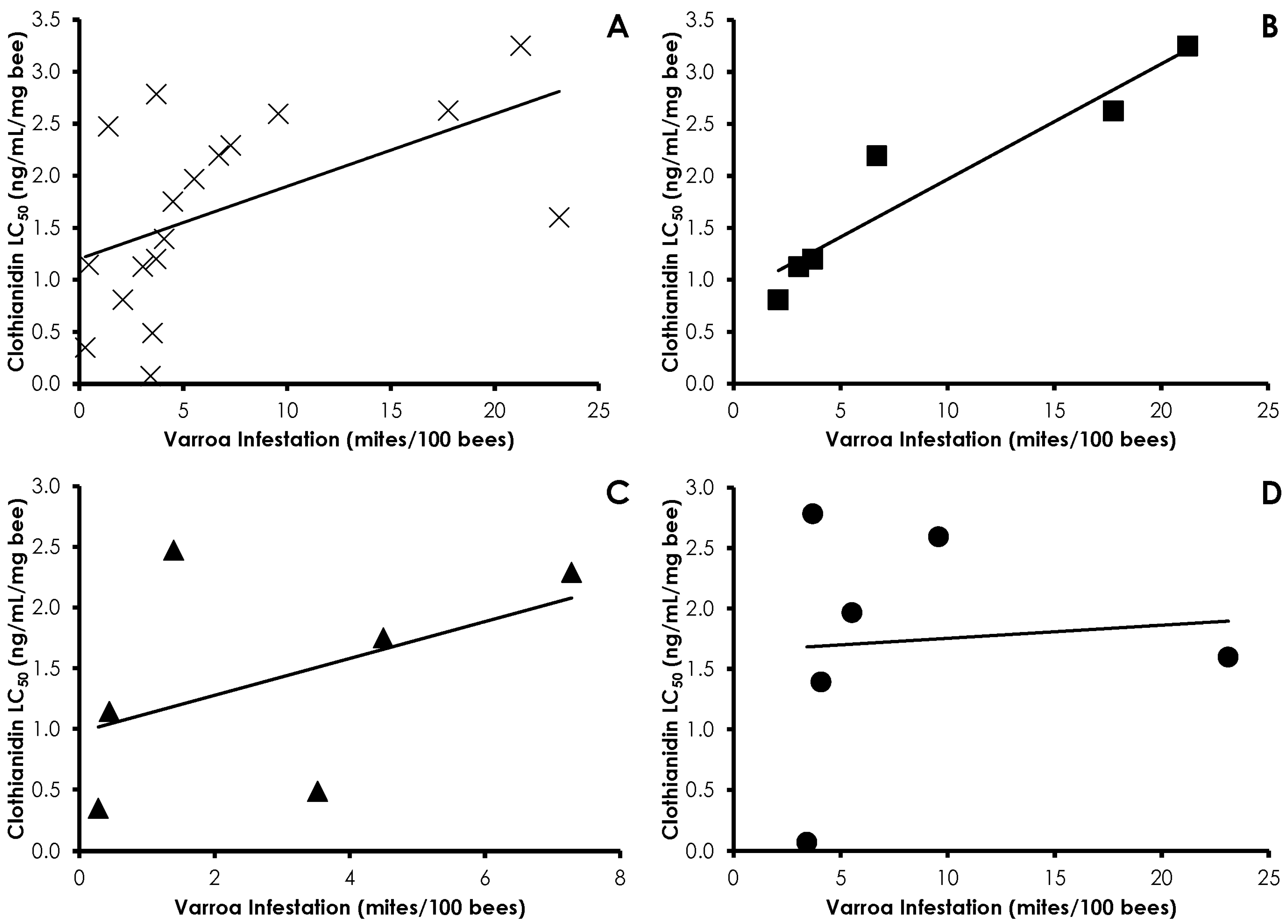
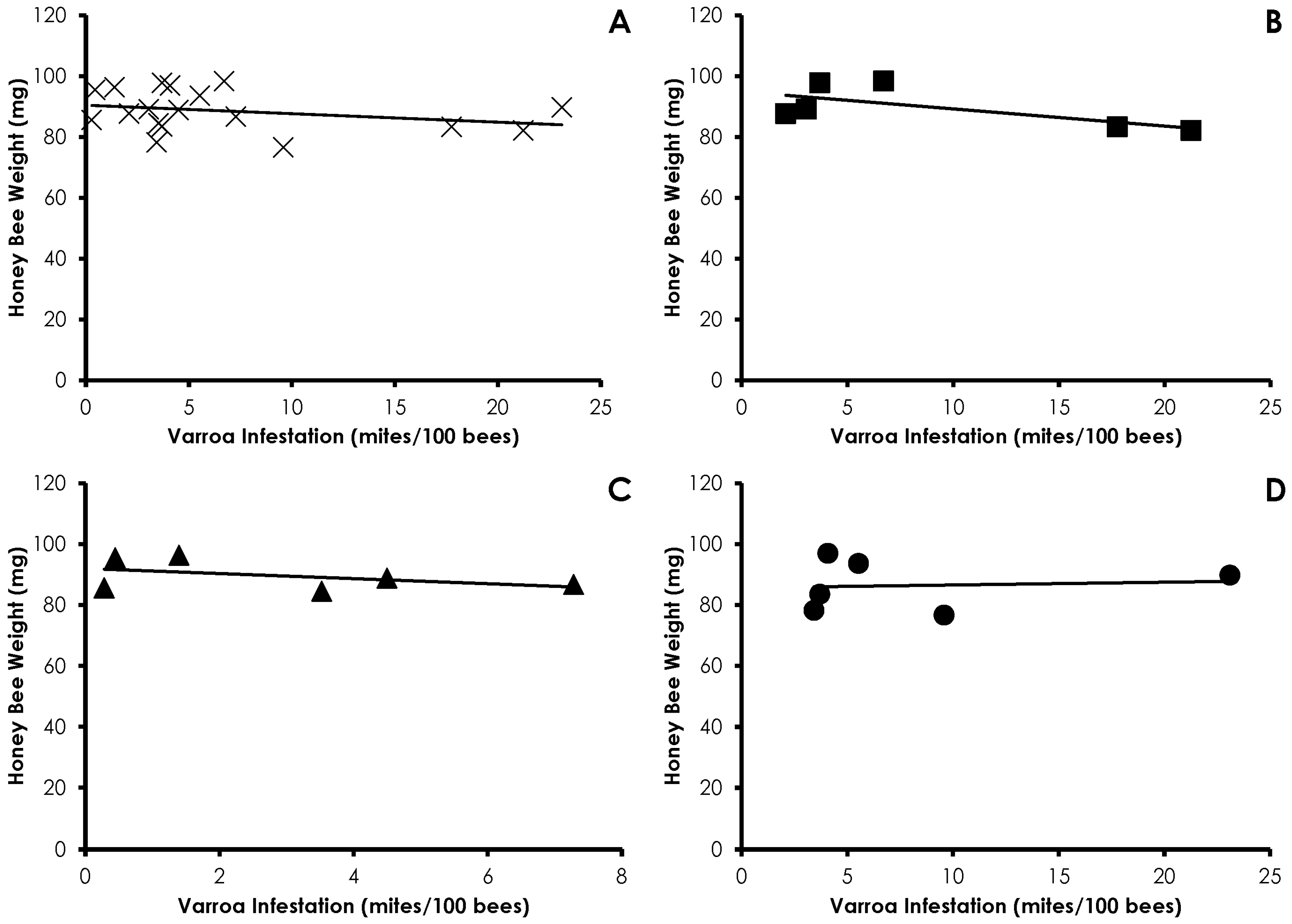
3.1. Varroa Infestation
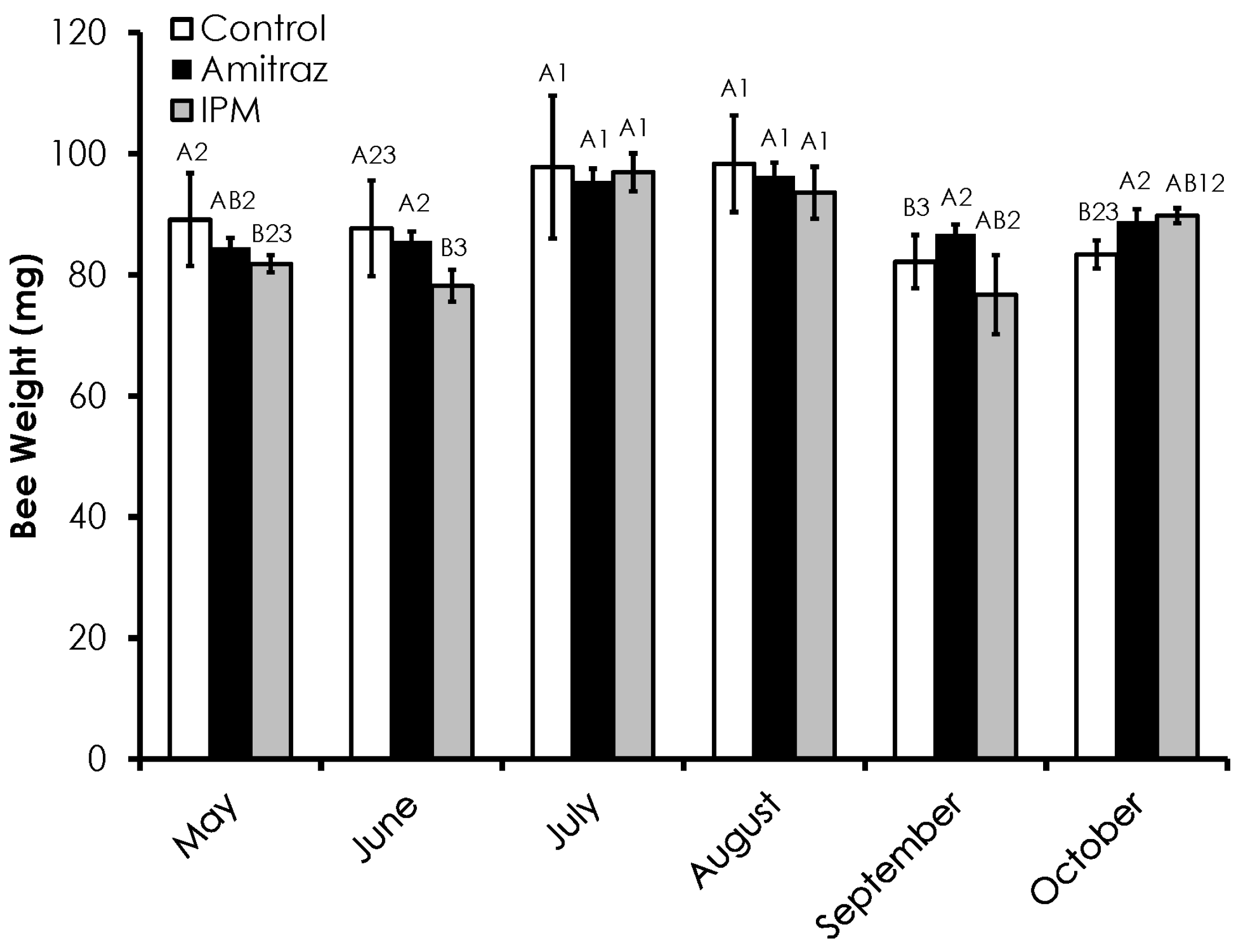
3.2. Honey Bee Weight
3.3. Bioassays
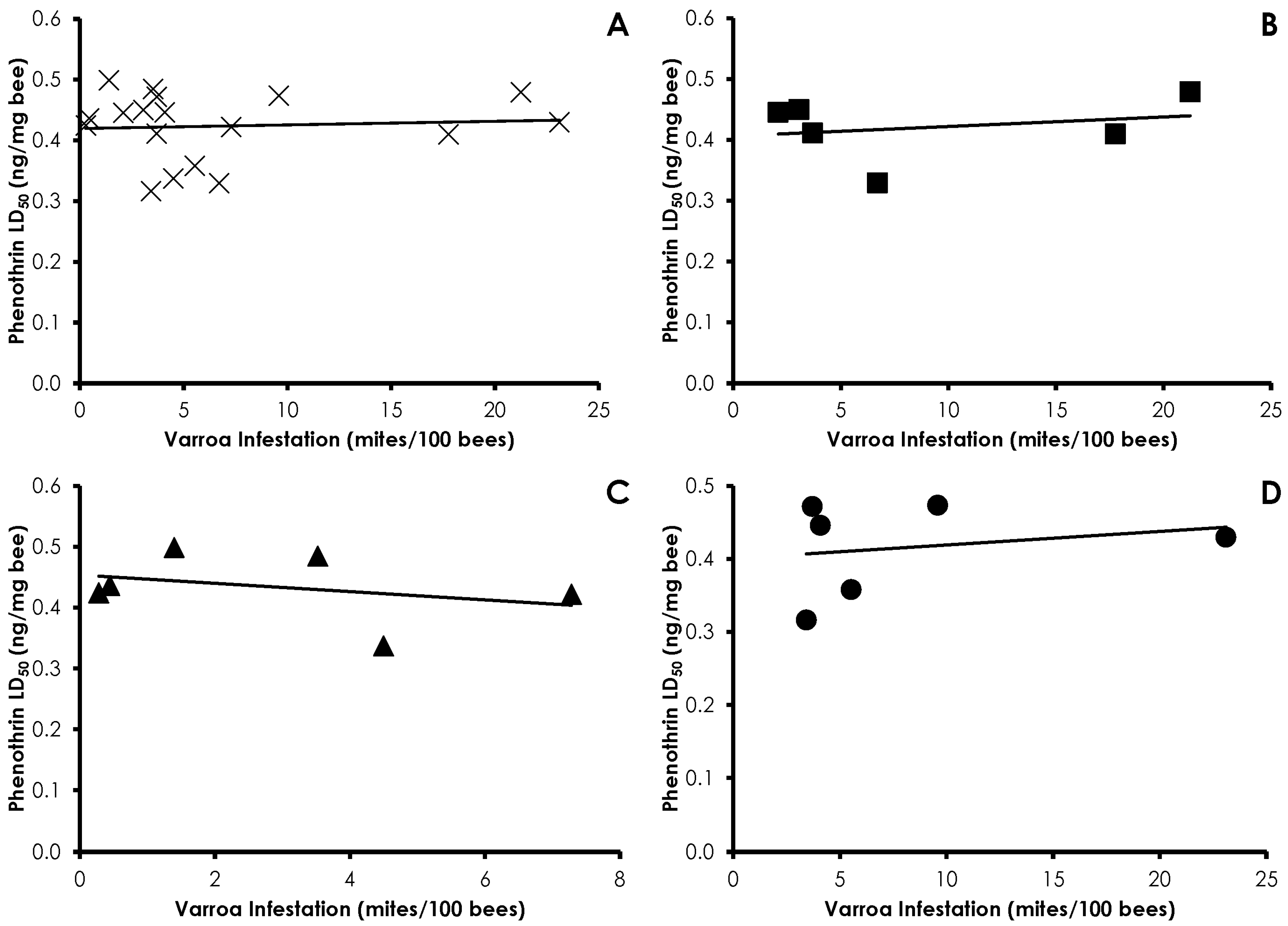
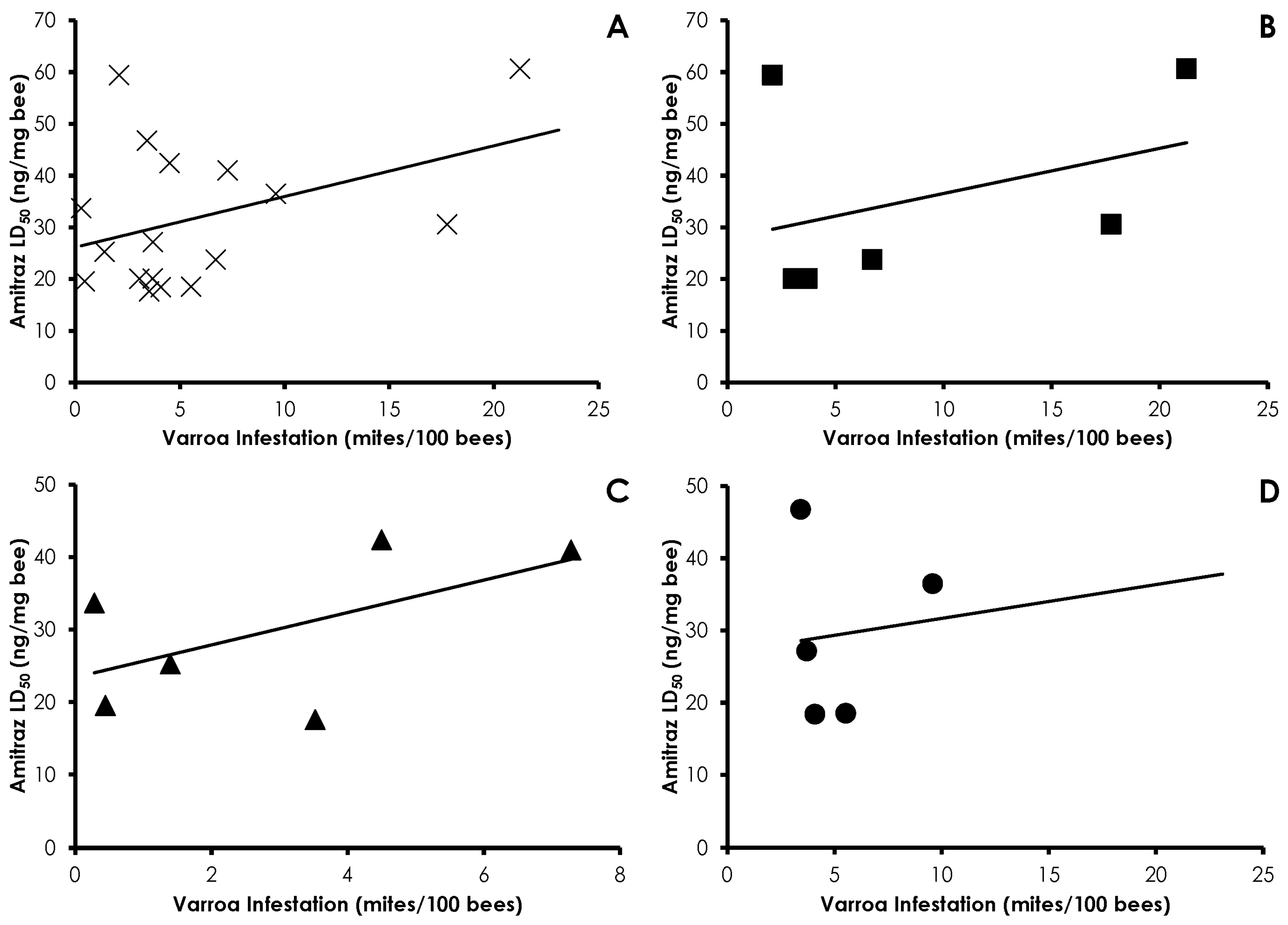
3.4. Interactions of Varroa Infestation, Pesticide Sensitivity, and Weight
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
References
- Finley, J.; Camazine, S.; Frazier, M. The epidemic of honey bee colony losses during the 1995–1996 season. Am. Bee J. 1996, 136, 805–808. [Google Scholar]
- Siede, R.; Konig, M.; Buchler, R.; Failing, K.; Thiel, H.J. A real-time PCR based survey on acute bee paralysis virus in German bee colonies. Apidologie 2008, 39, 650–661. [Google Scholar] [CrossRef]
- Gregory, P.G.; Evans, J.D.; Rinderer, T.; De Guzman, L. Conditional immune-gene supression of honeybees parasitized by Varroa mites. J. Insect Sci. 2005, 5, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Cox-Foster, D.L. Impact of an ectoparasite on the immunity and pathology of an invertebrate: Evidence for host immunosuppression and viral amplification. Proc. Natl. Acad. Sci. USA 2005, 102, 7470–7475. [Google Scholar] [CrossRef] [PubMed]
- Navajas, M.; Migeon, A.; Alaux, C.; Martin-Magniette, M.L.; Robinson, G.E.; Evans, J.D.; Cros-Arteil, S.; Crauser, D.; Le Conte, Y. Differential gene expression of the honey bee Apis mellifera associated with Varroa destructor infection. BMC Genom. 2008, 9, 301–311. [Google Scholar] [CrossRef]
- Duay, P.; De Jong, D.; Engels, W. Weight loss in drone pupae (Apis mellifera) multiply infested by Varroa destructor mites. Apidologie 2003, 34, 61–65. [Google Scholar] [CrossRef]
- Bowen-Walker, P.L.; Gunn, A. The effect of the ectoparasitic mite, Varroa destructor, on adult worker honey bee (Apis mellifera) emergence weights, water, protein, carbohydrate, and lipid levels. Entomol. Exp. Appl. 2001, 101, 207–217. [Google Scholar] [CrossRef]
- Amdam, G.V.; Hartfelder, K.; Norberg, K.; Hagen, A.; Omholt, S.W. Altered physiology in worker honey bees (Hhymenoptera: Apidae) infested with the mite Varroa destructor (Acari: Varroaidae): A factor in colony loss during overwintering? J. Econ. Entomol. 2004, 97, 741–747. [Google Scholar] [CrossRef] [PubMed]
- Seehuus, S.C.; Norberg, K.; Gimsa, U.; Krekling, T.; Amdam, G.V. Reproductive protein protects functionally sterile honey bee workers from oxidative stress. Proc. Natl. Acad. Sci. USA 2005, 103, 962–967. [Google Scholar] [CrossRef] [PubMed]
- Dahlgren, L.; Johnson, R.M.; Siegfried, B.D.; Ellis, M.D. Comparative toxicity of acaricides to honey bee (Hymenoptera: Apidae) workers and queens. J. Econ. Entomol. 2012, 105, 1895–1902. [Google Scholar] [CrossRef] [PubMed]
- Alaux, C.; Dantec, C.; Parrinello, H.; Le Conte, Y. Nutrigenomics in honey bees: Digital gene expression analysis of pollen’s nutritive effects on healthy and Varroa-parasitized bees. BMC Genom. 2011, 12, 496–508. [Google Scholar] [CrossRef] [PubMed]
- Baxter, J.; Eischen, F.; Pettis, J.; Wilson, W.; Shimanuki, H. Detection of fluvalinate-resistant Varroa mites in US honey bees. Am. Bee J. 1998, 138, 291. [Google Scholar]
- Elzen, P.; Westervelt, D. Detection of coumaphos resistance in Varroa destructor in Florida. Am. Bee J. 2002, 142, 291–292. [Google Scholar]
- Elzen, P.; Baxter, J.; Spivak, M.; Wilson, W. Amitraz resistance in Varroa: New discovery in North America. Am. Bee J. 1999, 139, 362. [Google Scholar]
- Mullin, C.A.; Frazier, M.; Frazier, J.L.; Ashcraft, S.; Simonds, R.; van Engelsdorp, D.; Pettis, J.S. High levels of miticides and agrochemicals in North American apiaries: Implications for honey bee health. PLoS ONE 2010, 5, e9754. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.M.; Dahlgren, L.; Siegfried, B.D.; Ellis, M.D. Acaricide, fungicide and drug interactions in honey bees (Apis mellifera). PLoS ONE 2013, 8, e54092. [Google Scholar] [CrossRef] [PubMed]
- Rinkevich, F.D.; Margotta, J.W.; Pittman, J.M.; Danka, R.G.; Tarver, M.R.; Ottea, J.A.; Healy, K.B. Genetics, synergists, and age affect insecticide sensitivity in the honey bee, Apis mellifera. PLoS ONE 2015, 10, e0139841. [Google Scholar] [CrossRef] [PubMed]
- Harbo, J.R.; Harris, J.W. Effect of screen floors on populations of honey bees and parasitic mites (Varroa destructor). J. Apic. Res. 2004, 43, 114–117. [Google Scholar] [CrossRef]
- Calderone, N.W. Evaluation of drone brood removal for management of Varroa destructor (Acari: Varroidae) in colonies of Apis mellifera (Hymenoptera: Apidae) in the northeastern United States. J. Econ. Entomol. 2005, 98, 645–650. [Google Scholar] [CrossRef] [PubMed]
- Aliano, N.P.; Ellis, M.D. A strategy for using powdered sugar to reduce Varroa populations in honey bee colonies. J. Apic. Res. 2005, 44, 54–57. [Google Scholar] [CrossRef]
- Delaplane, K.S.; Berry, J.A.; Skinner, J.A.; Parkman, J.P.; Hood, W.M. Integrated pest management against Varroa destructor reduces colony mite levels and delays treatment threshold. J. Apic. Res. 2005, 44, 157–162. [Google Scholar] [CrossRef]
- Rosenkranz, P.; Aumeier, P.; Ziegelmann, B. Biology and control of Varroa destructor. J. Invertebr. Pathol. 2010, 103, S96–S119. [Google Scholar] [CrossRef] [PubMed]
- Ellis, A.M.; Hayes, G.W.; Ellis, J.D. The efficacy of dusting honey bee colonies with powdered sugar to reduce Varroa mite populations. J. Apic. Res. 2009, 48, 72–76. [Google Scholar] [CrossRef]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Van Engelsdorp, D.; Meixner, M.D. A historical review of managed honey bee populations in Europe and the United States and the factors that may affect them. J. Invertebr. Pathol. 2010, 103, S80–S95. [Google Scholar] [CrossRef] [PubMed]
- Gallai, N.; Salles, J.M.; Settele, J.; Vaissiere, B.E. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 2009, 68, 810–821. [Google Scholar] [CrossRef] [Green Version]
- Guzman-Novoa, E.; Eccles, L.; Calvete, Y.; McGowan, J.; Kelly, P.G.; Correa-Benitez, A. Varroa destructor is the main culprit for the death and reduced populations of overwintered honey bee (Apis mellifera) colonies in Ontario, Canada. Apidologie 2010, 71, 443–450. [Google Scholar] [CrossRef]
- Boeking, O.; Genersch, E. Varroosis—The ongoing crisis in bee keeping. J. Verbrauch. Lebensm. 2008, 3, 221–228. [Google Scholar] [CrossRef]
- Berry, J.A.; Afik, O.; Nolan, M.P.; Delaplane, K.S. Revisiting powdered sugar for Varroa control on honey bees (Apis mellifera L.). J. Apic. Res. 2012, 51, 367–368. [Google Scholar] [CrossRef]
- Calderone, N.W. A three-year study of screened bottom boards in the northeast. Am. Bee J. 2007, 147, 436–437. [Google Scholar]
- Charriere, J.D.; Imdorf, A.; Bachofen, B.; Tschan, A. The removal of capped drone brood: An effective means of reducing the infestation of Varroa in honey bee colonies. Bee World 2003, 84, 117–124. [Google Scholar] [CrossRef]
- Smirle, M.J.; Winston, M.L. Intercolony variation in pesticide detoxification by the honey bee (Hymenoptera: Apidae). J. Econ. Entomol. 1987, 80, 5–8. [Google Scholar] [CrossRef]
- Harbo, J.R.; Harris, J.W. Responses to Varroa by honey bees with different levels of Varroa-Sensitive Hygiene. J. Apic. Res. 2009, 48, 156–161. [Google Scholar] [CrossRef]
- Danka, R.G.; Harris, J.W.; Dodds, G.E. Selection of VSH-derived “pol-line” honey bees and evaluation of their Varroa-resistance characteristics. Apidologie 2015, 47, 1–8. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Month | Control | Amitraz | IPM | ||||||
|---|---|---|---|---|---|---|---|---|---|
| n | LD50 (95% CI) | Slope (SE) | n | LD50 (95% CI) | Slope (SE) | n | LD50 (95% CI) | Slope (SE) | |
| May | 308 | 0.45 (0.35–0.52) a123 | 2.6 (0.4) | 239 | 0.48 (0.45–0.52) a1 | 6.4 (0.7) | 229 | 0.47 (0.32–0.57) a123 | 3.1 (0.6) |
| June | 489 | 0.45 (0.42–0.46) a13 | 7.2 (0.5) | 576 | 0.42 (0.40–0.44) a2 | 7.6 (0.5) | 531 | 0.32 (0.29–0.34) b3 | 5.8 (0.6) |
| July | 448 | 0.41 (0.38–0.43) a1 | 5.3 (0.5) | 374 | 0.43 (0.40–0.48) a12 | 3.7 (0.5) | 329 | 0.45 (0.41–0.49) a1 | 5.0 (0.6) |
| August | 383 | 0.33 (0.29–0.35) b2 | 4.7 (0.6) | 303 | 0.50 (0.46–0.54) a1 | 4.8 (0.6) | 543 | 0.35 (0.34–0.38) b2 | 4.7 (0.4) |
| September | 297 | 0.48 (0.45–0.51) a3 | 7.3 (0.7) | 252 | 0.42 (0.39–0.45) b12 | 6.3 (0.7) | 284 | 0.47 (0.44–0.50) ab1 | 6.7 (0.7) |
| October | 362 | 0.41 (0.37-0.44) a1 | 4.3 (0.6) | 464 | 0.34 (0.31–0.36) b3 | 4.8 (0.5) | 73 | 0.43 (0.31–0.52) ab123 | 4.7 (1.4) |
| Month | Control | Amitraz | IPM | ||||||
|---|---|---|---|---|---|---|---|---|---|
| n | LD50 (95% CI) | Slope (SE) | n | LD50 (95% CI) | Slope (SE) | n | LD50 (95% CI) | Slope (SE) | |
| May | 373 | 20.0 (18.4–21.8) b3 | 4.9 (0.4) | 339 | 17.6 (15.8–19.6) b5 | 3.7 (0.3) | 320 | 27.1 (24.7–29.7) a4 | 4.2 (0.4) |
| June | 380 | 59.4 (51.5–72.9) a1 | 3.4 (0.4) | 416 | 33.7 (31.1–36.6) b23 | 4.1 (0.4) | 368 | 46.8 (42.0–53.5) a12 | 3.5 (0.4) |
| July | 429 | 20.1 (18.5–21.9) a3 | 4.2 (0.3) | 238 | 19.5 (17.6–21.8) a5 | 5.0 (0.5) | 218 | 18.4 (16.3–20.9) a3 | 4.4 (0.5) |
| August | 324 | 23.8 (19.9–28.5) ab3 | 2.2 (0.3) | 328 | 25.3 (22.7–28.1) a4 | 3.5 (0.3) | 568 | 18.5 (16.7–20.5) b3 | 2.6 (0.2) |
| September | 327 | 60.6 (55.7–5.8) a1 | 4.3 (0.5) | 180 | 41.0 (35.8–46.1) b123 | 4.0 (0.6) | 255 | 36.4 (31.1–41.2) b2 | 4.0 (0.5) |
| October | 321 | 30.6 (28.5–32.3) b2 | 9.1 (1.3) | 350 | 42.4 (38.9–46.3) a12 | 4.4 (0.5) | NA | NA | NA |
| Month | Control | Amitraz | IPM | ||||||
|---|---|---|---|---|---|---|---|---|---|
| n | LC50 (95% CI) | Slope (SE) | n | LC50 (95% CI) | Slope (SE) | n | LC50 (95% CI) | Slope (SE) | |
| May | 308 | 1.12 (0.78–1.43) b3 | 2.4 (0.3) | 269 | 0.48 (0.09–0.93) b4 | 1.5 (0.3) | 293 | 2.78 (2.37–3.16) a1 | 4.0 (0.5) |
| June | 280 | 0.81 (0.51–1.10) a3 | 1.9 (0.3) | 460 | 0.35 (0.09–0.64) ab4 | 1.2 (0.2) | 479 | 0.07 (0.001–0.25) b5 | 1.0 (0.2) |
| July | 357 | 1.20 (1.07–1.37) a3 | 3.3 (0.3) | 253 | 1.14 (1.04–1.28) a3 | 5.6 (0.7) | 276 | 1.39 (1.22–1.65) a34 | 3.7 (0.4) |
| August | 325 | 2.19 (1.95–2.49) ab2 | 3.7 (0.4) | 287 | 2.47 (2.24–2.71) a1 | 5.8 (1.0) | 416 | 1.97 (1.81–2.15) b23 | 4.5 (0.4) |
| September | 370 | 3.25 (2.94–3.55) a1 | 4.3 (0.4) | 356 | 2.29 (1.95–2.62) b1 | 2.8 (0.3) | 340 | 2.59 (2.23–2.97) ab1 | 2.6 (0.3) |
| October | 296 | 2.62 (2.33–2.93) a2 | 4.0 (0.4) | 447 | 1.75 (1.60–1.90) b2 | 5.3 (0.4) | 54 | 1.60 (0.89–2.04) b234 | 7.3 (2.2) |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rinkevich, F.D.; Danka, R.G.; Healy, K.B. Influence of Varroa Mite (Varroa destructor) Management Practices on Insecticide Sensitivity in the Honey Bee (Apis mellifera). Insects 2017, 8, 9. https://doi.org/10.3390/insects8010009
Rinkevich FD, Danka RG, Healy KB. Influence of Varroa Mite (Varroa destructor) Management Practices on Insecticide Sensitivity in the Honey Bee (Apis mellifera). Insects. 2017; 8(1):9. https://doi.org/10.3390/insects8010009
Chicago/Turabian StyleRinkevich, Frank D., Robert G. Danka, and Kristen B. Healy. 2017. "Influence of Varroa Mite (Varroa destructor) Management Practices on Insecticide Sensitivity in the Honey Bee (Apis mellifera)" Insects 8, no. 1: 9. https://doi.org/10.3390/insects8010009