
Morphometric Modifications in Canthon quinquemaculatus Castelnau 1840 (Coleoptera: Scarabaeinae): Sublethal Effects of Transgenic Maize?

Programa de Pós Graduação em Ecologia, Departamento de Ecologia e Zoologia, Universidade Federal de Santa Catarina, Florianópolis 88040-900, Brasil
*
Author to whom correspondence should be addressed.
Insects 2017, 8(4), 115; https://doi.org/10.3390/insects8040115
Submission received: 21 August 2017
/
Revised: 27 September 2017
/
Accepted: 18 October 2017
/
Published: 21 October 2017
Abstract
:The effects of transgenic compounds on non-target organisms remain poorly understood, especially in native insect species. Morphological changes (e.g., changes in body size and shape) may reflect possible responses to environmental stressors, like transgenic toxins. The dung beetle Canthon quinquemaculatus (Coleoptera: Scarabaeinae) is a non-target species found in transgenic crops. We evaluated whether C. quinquemaculatus individuals inhabiting corn fields cultivated with different seed types (conventional, creole and transgenic) present modifications in body shape compared to individuals inhabiting adjacent native forest fragments. We collected C. quinquemaculatus specimens across an agricultural landscape in southern Brazil, during the summer of 2015. Six populations were sampled: three maize crop populations each under a different seed type, and three populations of adjacent forests. After sampling, specimens were subjected to morphometric analyses to discover differences in body shape. We chose fifteen landmarks to describe body shape, and morphometric data were tested with Procrustes ANOVA and Discriminant Analysis. We found that body shape did not differ between individuals collected in conventional and creole crops with their respective adjacent forests (p > 0.05); however, transgenic crop populations differed significantly from those collected in adjacent forests (p < 0.05). Insects in transgenic maize are more oval and have a retraction in the abdominal region, compared with the respective adjacent forest, this result shows the possible effect of transgenic crops on non-target species. This may have implications for the ecosystem service of organic matter removal, carried out by these organisms.
1. Introduction
Body shape is directly associated with several important ecological aspects, such as high-speed displacement capabilities, and consequently predator–prey interactions [1,2]. Shape characteristics can give advantages to males in sexual selection, having greater chances of mating [3]. Modifications in body shape offer advantages to certain types of behavior, such as foraging [4,5], reproduction [6] and habitat use [7]. Body morphology is also linked with community structuring since differences in shape are usually associated with different feeding and nesting strategies. Consequently, shape can be associated with better adapted competitors with improved foraging strategies in a particular community [8].
In addition to providing important information on species ecology, body shape may also indicate the effects of possible environmental stressors on individuals. These environmental stressors may be natural, such as ultraviolet radiation [9], or may be anthropic, like pollution [10]. Morphological alterations may imply possible ecological consequences, such as reduction in feed and displacement capacities [11,12]. Thus, in the face of environmental disturbances, body shape alterations may be classified as sublethal effects, and can affect an individual’s fitness [9,13]. Sublethal effects are defined as the physiological or behavioral changes in individuals exposed to chemical compounds [14]. These changes reduce individual fitness and have already been described in various non-target insects such as butterflies [15], bees [16] and dung beetles [17].
Sublethal effects of chemical compounds associated with agricultural practices have been observed in dung beetles. Dung beetles fed with feces containing Ivermectin (a chemical product used to control parasites in cattle) presented behavioral changes, increased developmental time, reduced fecundity and oviposition, and reduced cephalic capsule size [18,19,20]. Behavioral and physiological changes were also described in dung beetles fed with cattle feces containing transgenic maize. When fed with transgenic products, dung beetles showed lower dispersion velocity and alterations in the immune system [21]. These sublethal effects decrease the individual’s fitness, leading to abnormalities in their development, behavior and morphology. Fitness reduction may be evidenced by changes in individual size [22] or shape, particularly in holometabolous insects where the adult stage is greatly influenced by larval development. Therefore, larvae that develop in poor environments, with few resources, or under some kind of specific crop system (e.g., transgenic crops or under insecticidal and fungicidal effect) may result in adults presenting changes in shape and size.
Maize (Zea mays L.) is the second largest crop production in the world [23]. Maize production is associated, among other factors, with seed differences. The use of genetically modified seeds is growing globally, reaching approximately 180 million hectares in 2014 [24]. Since the beginning of its implementation in the mid-1980s, questions about their environmental safety have been raised [25]. Some studies have described the effectiveness and apparent safety of transgenic maize in reducing economically important insect populations and not harming non-target organisms [26,27]. On the other hand, several studies have shown target insect resistance development [28], accumulation of transgenic toxin compounds in the food chain [29], and lethal effects on non-target organisms [30]. Conventional seed varieties are severely affected by insects of economic importance, since the combination of food abundance with lack of natural enemies (due to native vegetation suppression) favors overpopulation of phytophagous insects, mainly Lepidoptera larvae. Creole seeds (landraces) normally are less productive than other seeds; however, they have increased genetic variability, and more resistance to phytophagous insects and specific environmental conditions [31]. These landraces can be productive even in unfavorable conditions [32], and represent a genetic heritage since they are selected by the farmers themselves, through the selection of more adapted individuals [33].
Canthon quinquemaculatus Castelnau 1840 is a non-target species that feeds on decaying material, and thus is engaged in nutrient cycling, promoting environmental services regulating physico-chemical soil properties [34,35]. C. quinquemaculatus is a widespread species throughout South America and it feeds on feces and carcasses of mammals and birds. These animals may consume different types of maize. Bt toxin can be transferred through the trophic chain: individuals of Orius majusculus (an insect predator) contained Bt toxins when feeding on spider mites reared on transgenic maize [36]. Thus, through the trophic chain these Bt toxins can accumulate and reach C. quinquemaculatus populations. In addition to the accumulation of transgenic toxins, individuals may suffer from different types of inputs, such as insecticides, which are used in maize crops. These inputs are greater in transgenic and conventional seeds, and are extremely low in creole seeds. Under the assumption that changes in individual shape indicate possible effects on non-target species physiology, the aim of this study was to evaluate whether C. quinquemaculatus individuals present morphometric differences in populations found within different maize crops (conventional, creole, and transgenic seeds) when compared with populations found in adjacent control populations inhabiting forest remnants.
2. Materials and Methods
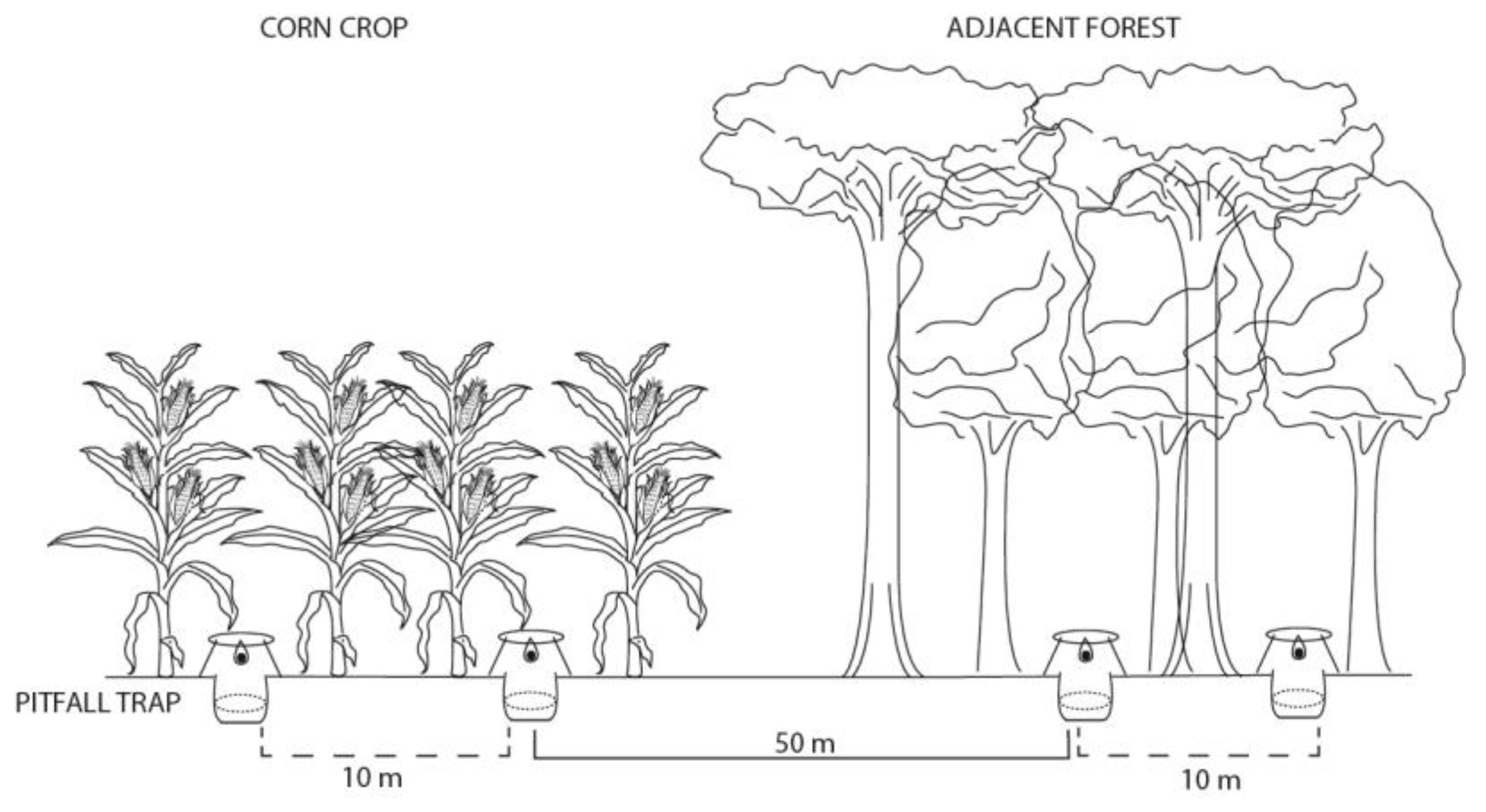
Population sampling occurred in the region of São Miguel do Oeste, Santa Catarina State, Brazil (26°43′31″ S, 53°31′05″ W). The region is composed of a mosaic of Atlantic forest fragments and maize crops. Local climate is humid subtropical (Cfa) according to the Köppen classification, with an annual average temperature between 16.3 °C and 17.9 °C, and rainfall between 1790 mm and 2280 mm [37]. Samples were collected during the summer of 2015 in maize crops with different profiles: five areas of conventional maize, five areas of creole maize, and five areas of transgenic maize. Furthermore, 15 adjacent forest remnants, one for each crop area, were sampled in order to have an independently replicated control population for each type of crop. The insects were collected using ten pitfall traps per area, five baited with human feces (10 g) and five with pig carrion (10 g) during 48 h. The traps were spaced 10 m apart within the same area, and a minimum distance of 50 m was established between crop traps and forest fragments to avoid interference (Appendix A Figure A1). Captured insects were fixed in 70% alcohol and deposited in the entomological collection of the Federal University of Santa Catarina (UFSC).
From a total of 231 specimens of C. quinquemaculatus collected, we used 157 individuals for morphometric analyses. We tried to balance the proportion between males and females in the most equitable way possible. Thus, we measured 12 specimens from conventional maize crops (6♀, 6♂); 17 from creole crops (11♀, 6♂); 28 from transgenic crops (14♀, 14♂); 36 from forest fragments adjacent to conventional crops (18♀, 18♂); 30 from forest fragments adjacent to creole crops (10♀, 20♂); and 34 from forests adjacent to transgenic crops (17♀, 17♂).
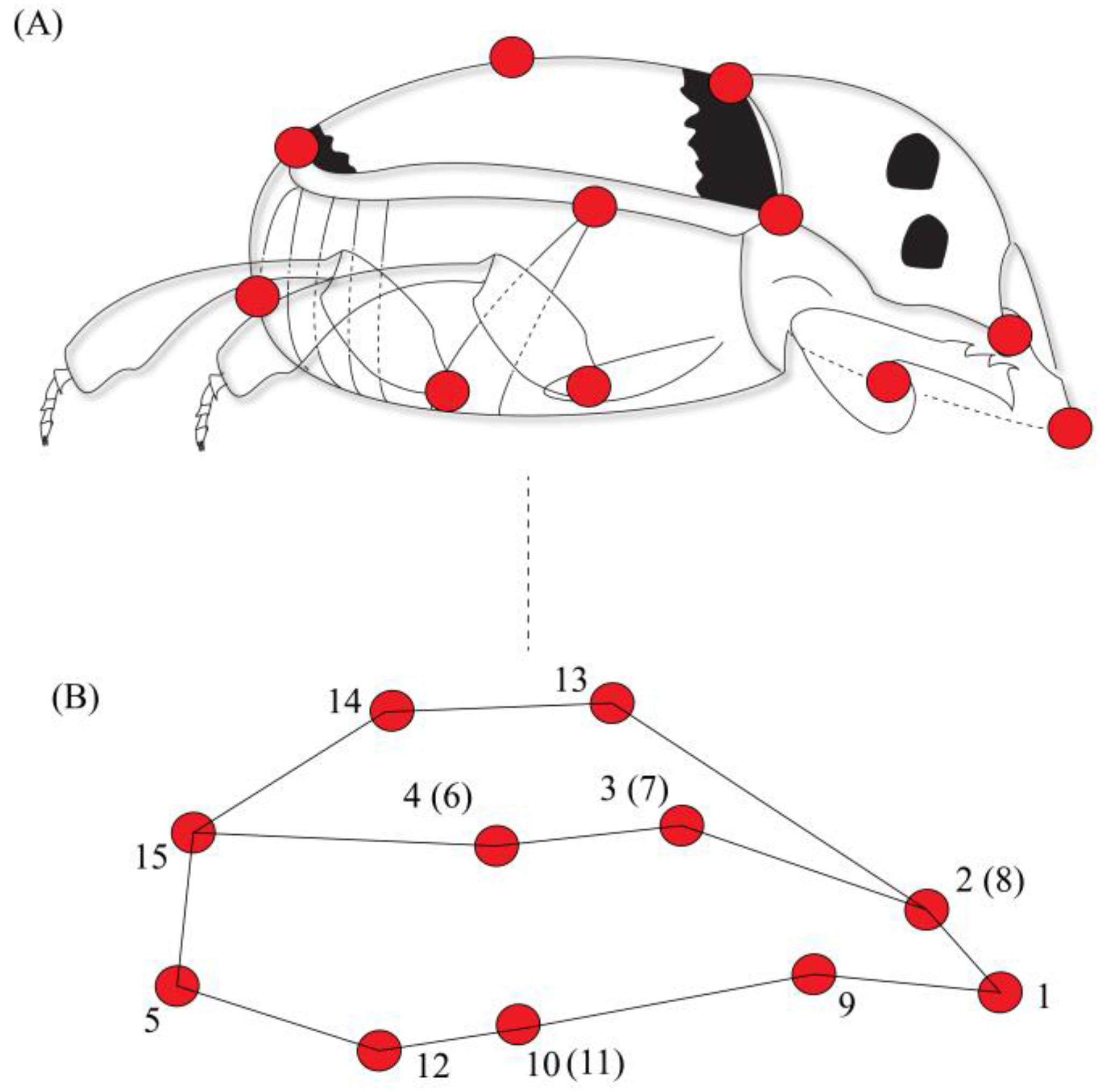
We defined the body shape through the placement of 15 anatomical landmarks. These 15 landmarks were chosen because they capture all morphological variations describing body shape in both anteroposterior and ventral dorsal axis, and these landmarks have already been successfully used to describe dung beetle body shape [8]. Each landmark corresponded to a point in space defined by three-dimensional cartesian coordinates (x, y, z) [38] as follows: (1) anterior margin of the head; (2) eye position; (3) division between the pronotum and the elytra; (4) division between the thorax and the abdomen; (5) posterior margin of the abdomen; (9) point of insertion of the anterior legs; (10 and 11) points of insertion of the central legs; (12) point of insertion of the posterior legs; (13) anterior point of convergence between elytra; (14) central point (mid-line) of convergence between elytra; and 15) posterior margin (along the mid-line) of the elytra. Points 6, 7 and 8 corresponded to points 4, 3 and 2 for the other side of the body, respectively [8]. Insects were photographed using Canon T3i camera in dorsal, ventral and lateral view; therefore, body shape was captured in three dimensions (Figure 1). A standard protocol for photographing specimens was established: insects were photographed on millimeter paper on a fixed surface to minimize the position effects. The camera was attached to a tripod and kept at a distance of 10 cm from each specimen at a perpendicular angle. Each individual had a tag, and for digitalization of the landmarks the individuals were mixed, so the marking was blind.
Coordinates from each landmark underwent Generalized Procrustes Analysis (GPA). The GPA is a three-step multivariate technique that assesses body shape eliminating the effects of size, position and orientation. The first step of the GPA is to center each landmark at the cartesian origin, eliminating the position effect. Afterwards, the landmark coordinates are scaled, eliminating the effect of size. Lastly, the landmark coordinates are rotated around the origin, which removes the orientation effect. Through these three steps, size and orientation data are removed leaving only body shape information [39]. We performed a canonical variable analysis (CVA) to view each individual’s position in the multivariate space. The overall shape difference was primarily tested with a Procrustes ANOVA, and to test the differences between maize crop populations and adjacent forest populations we performed a discriminant analysis (DA) based on the Mahalanobis distance. The reliability of a population’s individuals was determined by cross-validation to calculate the correct percentage of classification of the individuals in the original population, based only on the morphometric variation. Anatomical landmarks were inserted into specimens with the softwares TpsDig 2.16 [40] and tpsUtil1.53 [41]. The GPA, CVA and DA were performed in Morpho J 1.05d software [42].
3. Results
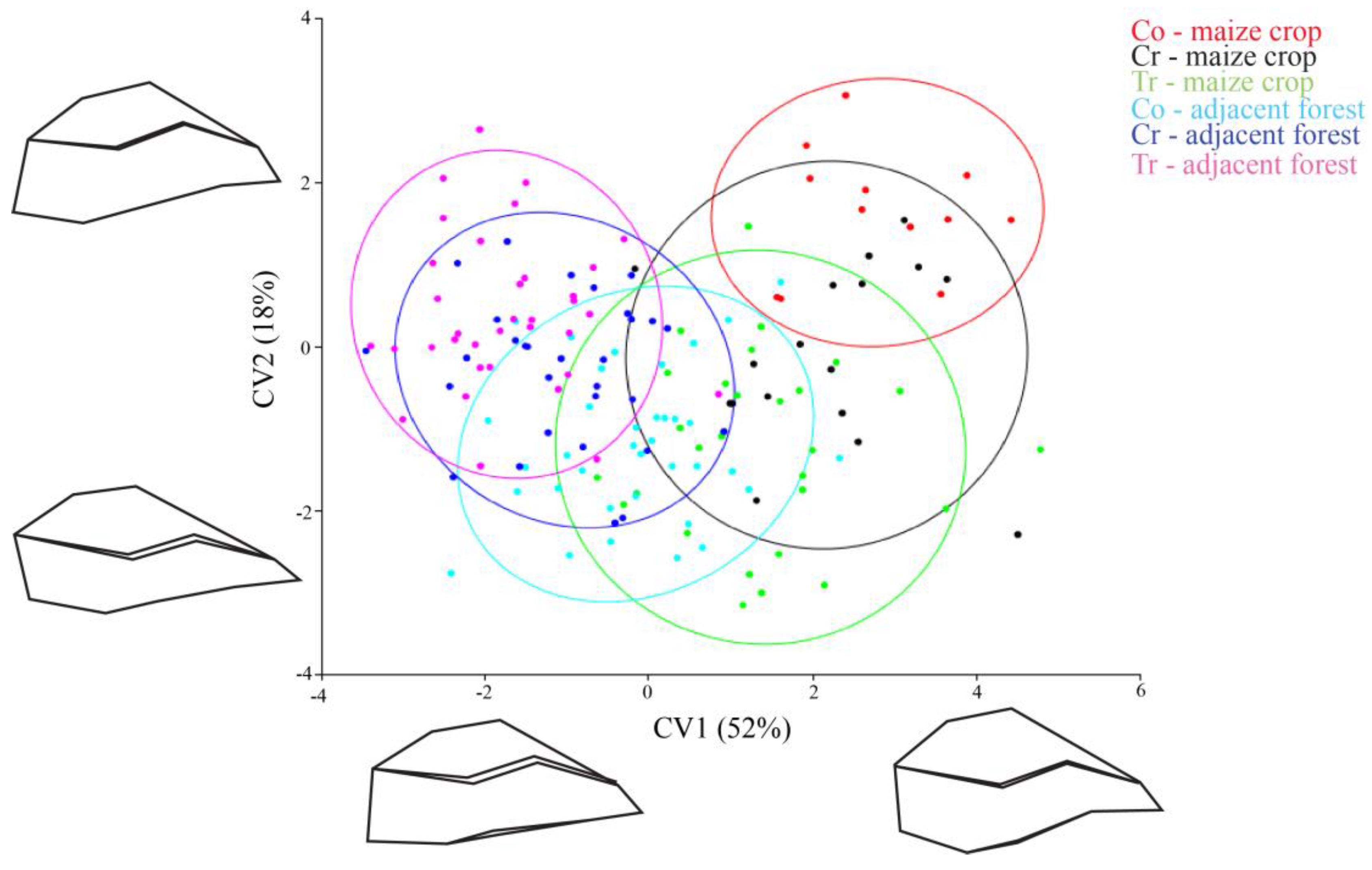
The shape of the individuals was assessed from the 15 landmarks and each specimen was represented by a point in Figure 2. Together, the first (CV1) and second (CV2) canonical variables in the CVA explained 70% of the observed body shape variation. The CVA showed that maize crop populations have mostly positive values in CV1 (red, black and green points in the Figure 2). On the other hand, forest fragment populations were mostly negative (light blue, blue and pink) and inferred a shape distinction between the insects that inhabit maize crops and those from the adjacent forests.
The CV1 (which accounted for 52% of the variance) organized individuals in a flat (negative scores) to oval (positive scores) body shape gradient; forest insects corresponded to a flat body shape and maize crop insects to an oval body shape (see the population’s shape design in both corners of Figure 2). Populations with positive CV1 (maize crop populations) (body shape in the right corner of Figure 2) had the landmarks of the dorsal region displaced above average shape, and landmark 5 (corresponding to abdominal region) displaced towards the body center. Together, these landmark displacements indicated a pronounced oval shape and an abdominal contraction (Figure 2 right corner). Populations with negative scores in CV1 (forest fragment populations) had landmark 5 (abdominal region) dislocated away from body center, and had the landmarks of the dorsal region displaced toward the body center. These rearrangements resulted in a flattened body shape (Figure 2, left corner).
Regarding CV2, there was no clear distinction between insects, insects with positive scores had dorsal region landmarks displaced towards the body center, and the ventral region landmarks dislocated away from the body center. These landmark displacements indicated a retraction on the anterior–posterior axis of body (Figure 2, designs at the top of CV2). Insects with negative scores had the dorsal region landmarks displaced away from the body center, and the ventral region landmarks dislocated towards the body center. This landmark configuration indicated an extension of the body’s anterior–posterior axis (Figure 2, designs at bottom of CV2).
The cross-validation tests indicated that an average of 75% (ranging between 58% and 90%) of individuals were correctly allocated to their respective populations. Maize crop populations had a high validation (ranging between 64% and 90%), including transgenic maize crop populations with 82% (Table 1).
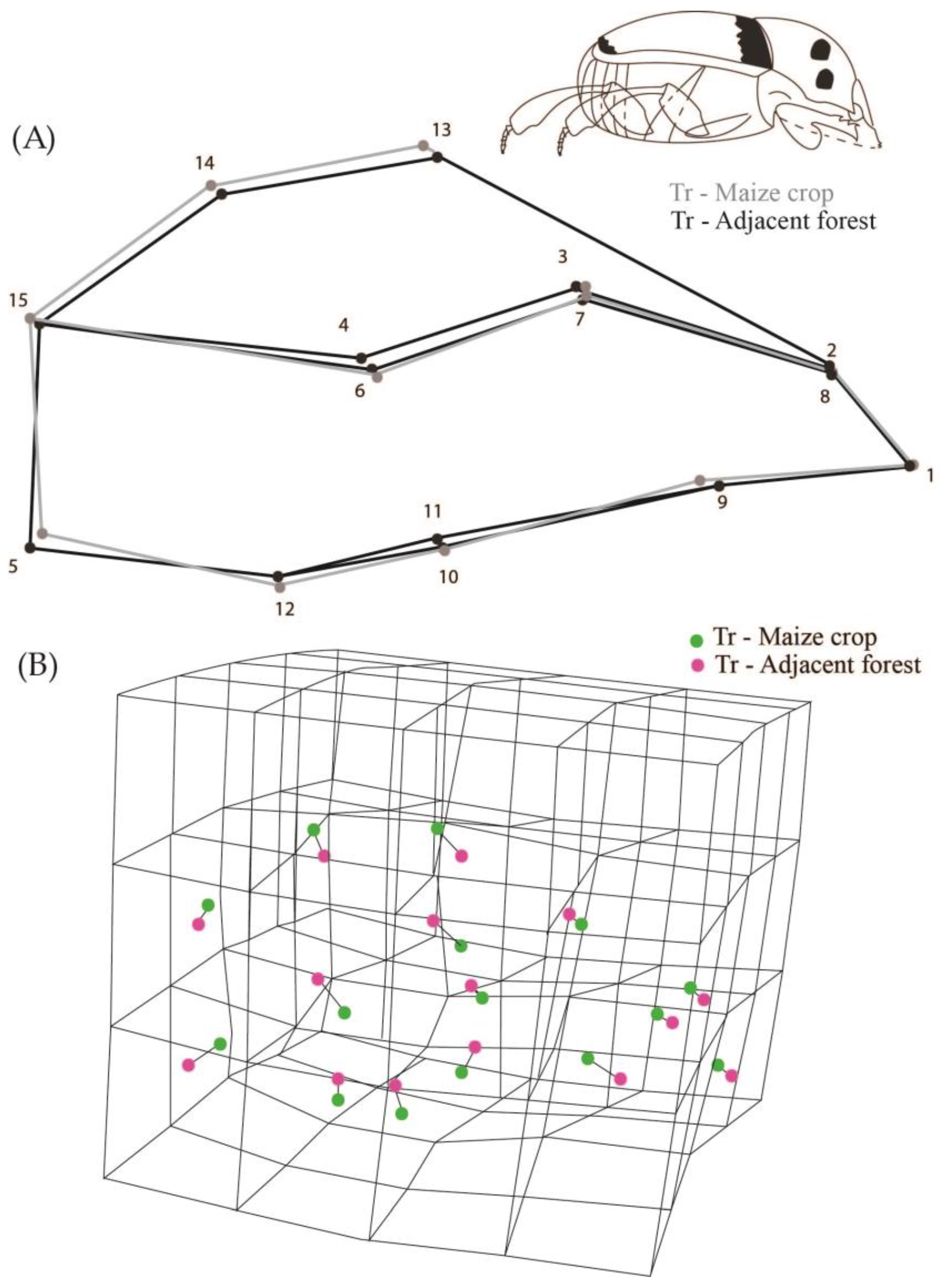
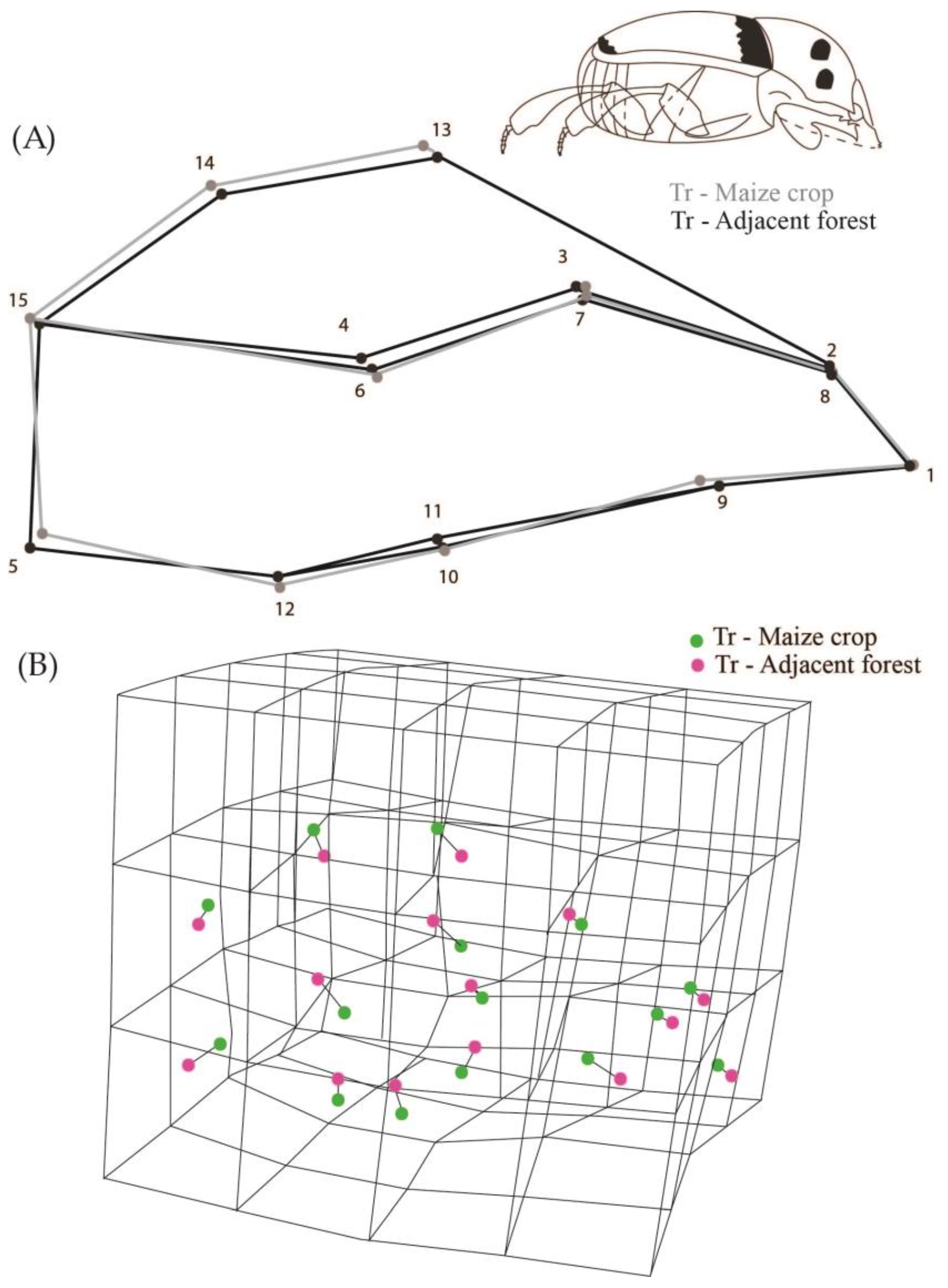
While analyzing all the populations we observed that they were different in relation to body shape (Procrustes ANOVA, F = 2.28, Pillai’s Trace = 2.03, p < 0.0001). We observed that the shape of the populations of creole and conventional crops were not significantly different from their respective adjacent forest areas (DA, p > 0.05). However, populations from transgenic maize crops differed in body shape from the populations of adjacent forests (DA, p = 0.0002) (Figure 3). The transgenic maize crop populations have the dorsal region landmarks higher up than the average shape of insects from adjacent forests (landmarks 13, 14 and 15 in Figure 3), and landmark 5 shifted to the center of the body. Consequently, this configuration indicates that populations of transgenic crops had a more oval shape and an abdominal retraction compared to individuals from adjacent forest populations (Figure 3).
4. Discussion
We observed different body shape patterns for C. quinquemaculatus individuals living in different habitats; populations found in transgenic maize crops presented significant morphometric modifications when compared to adjacent forest fragments. To date no published studies have reported changes in the entire body shape in response to the use of transgenic organisms in an agricultural environment. Studies with Diabrotica virgifera virgifera C. (Coleoptera: Chrysomelidae) collected in different types of maize crop, including transgenic crops, showed that the head of males inhabiting in transgenic maize was significantly smaller than in other treatments [43]. Similarly, individuals of Papilio machaon L. (Lepidoptera: Papilionidae) fed with transgenic pollen had lower body mass [22]. These results indicated that immature stages (larvae) could be exposed to sublethal doses of toxic transgenic maize, resulting in smaller adults. While implications of body size as a result of environmental effects are well documented [44], ecological implications of changes in body shape remain poorly known.
Although we did not detect changes in the body size of C. quinquemaculatus, we found differences in adult morphometrics, leading us to consider that the larvae or adults may have suffered effects derived from feeding on transgenic toxins; furthermore, laboratory experiments are needed to test for differences in the morphology of adults fed with transgenic toxins. With respect to body size, several studies failed to detect changes in insect populations grown in transgenic crops [45,46]. Inability of body size and mass to detect transgenic effects on non-target insect species suggests that both variables are less sensitive than shape, and using body shape to detect sublethal effects may be more efficient and informative. Changes in shape might have a major genetic influence compared to changes in size and mass, which are mainly driven by environmental factors [47].
Changes in body shape or in the shape of functional traits (e.g., wings, legs) can greatly affect some ecological attributes like sexual selection [48], adaptation [49], competition [8], development [50], and predation [51], causing significant changes in important populational parameters such as density, morphological variation, fitness, evolution and survival. Very few studies have evaluated changes in body shape or in the shape of bodily structures in response to transgenic compounds. Rahman et al. [52] observed alterations to skull shape in transgenic tilapia fish (Oreochromis niloticus). Studies with Pacific Salmon (Oncorhynchus spp.) showed changes in the shape of the head, tail and abdominal regions in response to a transgenic insertion [53]. Studies with transgenic Coho Salmon (O. kisutch) showed body shape differences between transgenic individuals and wild individuals even if they developed at the same temperatures [12]. This shape difference observed in Oncorhynchus spp. can affect its ability to swim, escape from predators and migrate. Similarly, Dunham et al. [54] found body shape differences in transgenic common carp (Cyprinus carpio). Furthermore, animals fed with transgenic organisms showed morphometric alterations; rats fed with transgenic potatoes presented shape alterations in the ileum [55]. Shape alterations in genetically modified organisms have also been described for plants; for example, Armon et al. [56] found differences in the leaf shape of wild and genetically modified Arabidopsis sp. specimens, suggesting that gene manipulation may interfere with growth regulating systems.
We have indications that transgenic compounds accumulating in the trophic chain affect the dung beetles, causing abnormalities in their physiology and behavior [21]. This pattern may be a product of a sum of factors, like historical management or use of different kinds of insecticides. Habes et al. [57] demonstrated morphometric alterations in Blatella germanic ovaries after insecticide application. Moreover, the management associated with transgenic crops alters the structure of the dung beetle community, reducing species abundance [30]. Therefore, the results from C. quinquemaculatus have given us another clue as to how transgenic compounds may be affecting dung beetles. As observed for C. quinquemaculatus, changes in insect morphometrics may lead to a decrease in insect fitness in response to the use of transgenics.
5. Conclusions
We have shown that the transgenic maize populations of C. quinquemaculatus are associated with changes in body shape, which may be a possible sub-lethal effect of the transgenic crop in this non-target species. These effects may have repercussions on the ecosystem functions performed by these insects, such as organic matter removal and regulation of soil physico-chemical properties.
Acknowledgments
We are grateful to Paulo Emilio Lovato (Department of Rural Engineering/UFSC) for coordinating the project “Fortalecimento das condições de produção e oferta de sementes de milho para a produção orgânica e agroecológica do Sul do Brasil” (CNPq Call 048/13). VMA would like to thank CAPES (Ministry of Education of Brazil) for a PhD Grant and MIMH would like to thank CNPq (Proc. 309030/2013-7) for a Research Productivity Grant.
Author Contributions
Both authors contributed equally in this work.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
Figure A1.
Experimental design scheme used for the captured individuals of C. quinquemaculatus in maize crops and adjacent forests of Atlantic Forest in the region of São Miguel do Oeste, SC, Brazil.
Figure A1.
Experimental design scheme used for the captured individuals of C. quinquemaculatus in maize crops and adjacent forests of Atlantic Forest in the region of São Miguel do Oeste, SC, Brazil.

References
- Burns, J.G.; di Nardo, P.; Rodd, F.H. The role of predation in variation in body shape in guppies Poecilia reticulata: A comparison of field and common garden phenotypes. J. Fish. Biol. 2009, 75, 1144–1157. [Google Scholar] [CrossRef] [PubMed]
- Dayton, G.H.; Saenz, D.; Baum, K.A.; Langerhans, R.B.; DeWitt, T.J. Body shape, burst speed and escape behavior of larval anurans. Oikos 2005, 111, 582–591. [Google Scholar] [CrossRef]
- Rowe, L.; Arnqvist, G. Sexual selection and the evolution of genital shape and complexity in water striders. Evolution 2012, 66, 40–54. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.A. Ecological morphology of lacustrine threespine stickleback Gasterosteus aculeatus L. (Gastoresteidae) body shape. Biol. J. Linn. Soc. 1997, 61, 3–50. [Google Scholar] [CrossRef]
- Keast, A.; Webb, D. Mouth and body form relative to feeding ecology in the fish fauna of a small lake, lake Opinicon, Ontario. J. Fish. Res. Board Can. 1966, 23, 1845–1874. [Google Scholar] [CrossRef]
- Forsman, A.; Shine, R. Parallel geographic variation in body shape and reproductive life history within the Australian scincid lizard Lampropholis delicata. Funct. Ecol. 1995, 9, 818–828. [Google Scholar] [CrossRef]
- Vanhooydonck, B.; Damme, R.V. Evolutionary relationships between body shape and habitat use in lacertid lizards. Evol. Ecol. Res. 1999, 1, 785–805. [Google Scholar]
- Hernández, M.I.M.; Monteiro, L.R.; Favila, M.E. The role of body size and shape in understanding competitive interactions within a community of Neotropical dung beetles. J. Insect Sci. 2011, 11, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Bernal, M.H.; Alton, L.A.; Cramp, R.L.; Franklin, C.E. Does simultaneous UV-B exposure enhance the lethal and sub-lethal effects of aquatic hypoxia on developing anuran embryos and larvae? J. Comp. Physiol. B 2011, 181, 973–980. [Google Scholar] [CrossRef] [PubMed]
- Primost, M.A.; Bigatti, G.; Márquez, F. Shell shape as indicator of pollution in marine gastropods affect by imposex. Mar. Freshw. Res. 2015, 67, 1948–1954. [Google Scholar] [CrossRef]
- Chan, K.Y.K.; Grünbaum, D.; O’Donnell, M.J. Effects of ocean-acidification-induced morphological changes on larval swimming. J. Exp. Biol. 2011, 214, 3857–3867. [Google Scholar] [CrossRef] [PubMed]
- Lõhmus, M.; Susdström, L.F.; Björklund, M.; Devlin, R.H. Genotype-temperature interaction in the regulation of development, growth, and morphometrics in wild-type, and growth-hormone transgenic coho salmon. PLoS ONE 2010, 5, e9980. [Google Scholar] [CrossRef] [PubMed]
- Woodley, S.K.; Matters, B.M.; Yates, E.K.; Relyea, R.A. Exposure to sublethal concentrations of a pesticide or predator cues induces changes in brain architecture in larval amphibians. Oecologia 2015, 179, 655–665. [Google Scholar] [CrossRef] [PubMed]
- Desneux, N.; Decourtye, A.; Delpuech, J.M. The sublethal effects of pesticides on benefical arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef] [PubMed]
- Pecenka, J.R.; Lundegren, J.G. Non-target effects of clothianidin on monarch butterflies. Sci. Nat. 2015, 102, 1–4. [Google Scholar] [CrossRef]
- Dively, G.P.; Embrey, M.S.; Kamel, A.; Hawthorne, D.J.; Pettis, J.S. Assessment of Chronic Sublethal Effects of Imidacloprid on Honey Bee Colony Health. PLoS ONE 2015, 10, e0118748. [Google Scholar] [CrossRef]
- Verdú, J.R.; Cortez, V.; Ortiz, A.J.; González-Rodríguez, E.; Martinez-Pinna, J.; Lumaret, J.P.; Lobo, J.M.; Numa, C.; Sánchez-Piñero, F. Low doses of ivermectin cause sensory and locomotor disorders in dung beetles. Sci. Rep. 2015, 5, 13912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sommer, C.; Nielsen, B.O. Larvae of the dung beetle Onthophagus gazella F. (Col., Scarabaeidae) exposed to lethal and sublethal ivermectin concentrations. J. Appl. Entomol. 1992, 114, 502–509. [Google Scholar] [CrossRef]
- Rosales, M.C.; Martínez, M.I.; López-Collado, J.; Vargas-Mendoza, M.; González-Hernández, H.; Fajersson, P. Effect of ivermectin on the survival and fecundity of Euoniticellus intermedius (Coleoptera: Scarabaeidae). Rev. Biol. Trop. 2012, 60, 333–345. [Google Scholar]
- Pérez-Cogollo, L.C.; Rodriguez-Vivas, R.I.; Delfín-González, H.; Reyes-Novelo, E.; Ojeda-Chi, M.M. Lethal and sublethal effects of Ivermectin on Onthophagus landolti (Coleoptera: Scarabaeidae). Environ. Entomol. 2015, 44, 1634–1640. [Google Scholar] [CrossRef] [PubMed]
- Campos, R.C. Influência do cultivo de milho transgênico em organismos não-alvo (Coleoptera: Scarabaeinae) e da ingestão indireta através da cadeia trófica. Ph.D. Thesis, Federal University of Santa Catarina, Florianópolis, Brazil, July 2016. [Google Scholar]
- Lang, A.; Vojtech, E. The effects of pollen consumption of transgenic Bt maize on the common swallowtail, Papilio machaon L. (Lepidoptera, Papilionidae). Basic Appl. Ecol. 2006, 7, 296–306. [Google Scholar] [CrossRef]
- Food and Agriculture Organization (FAO). Food and Agriculture Organization of the United Nations. Available online: http://www.fao.org/faostat (accessed on 27 September 2017).
- International Service for the Acquisition of Agri-biotech Applications (ISAAA). Global Status of Commercialized Biotech/GM Crops: 2014. Brief 49–2014: Executive Summary. Available online: http://isaaa.org (accessed on 20 November 2016).
- Ceccarelli, S. GM Crops, Organic Agriculture and Breeding for Sustainability. Sustainability 2014, 6, 4273–4286. [Google Scholar] [CrossRef]
- Zwahlen, C.; Hilbeck, A.; Nentwig, W. Field decomposition of transgenic Bt maize residue and the impact on non-target soil invertebrates. Plant Soil 2007, 300, 245–257. [Google Scholar] [CrossRef]
- Daly, T.; Buntin, G.D. Effect of Bacillus thuringiensis transgenic corn for Lepidopteran control on nontarget arthropods. Environ. Entomol. 2005, 34, 1292–1301. [Google Scholar] [CrossRef]
- Gassmann, A.J.; Petzold-Maxwell, J.L.; Keweshan, R.S.; Dunbar, M.W. Field-evolved resistance to Bt maize by Western Corn Rootworm. PLoS ONE 2011, 6, e22629. [Google Scholar] [CrossRef] [PubMed]
- Hilbeck, A.; Moar, W.J.; Pusztai-Carey, M.; Filipini, A.; Bigler, F. Toxicity of Bacillus thuringiensis Cry1Ab toxin to the predator Chrysoperla carnea (Neuroptera, Chrysopidae). Environ. Entomol. 1998, 27, 1255–1263. [Google Scholar] [CrossRef]
- Campos, R.C.; Hernández, M.I.M. The importance of maize management on dung beetle communities in Atlantic Forest fragments. PLoS ONE 2015, 10, e0145000. [Google Scholar] [CrossRef] [PubMed]
- Sandri, C.A.; Tofanelli, M.B.D. Milho crioulo: Uma alternativa para retabilidade no campo. Pesqui. Agropecu. Trop. 2008, 38, 59–61. [Google Scholar]
- Spaner, D.; Brathwaite, R.A.I.; Mather, D.E. Comparison of open-pollinated stress-tolerant and landrace maize for production under stress conditions in Trinidad. Maydica 1995, 40, 331–337. [Google Scholar]
- Ferreira, J.M.; Moreira, R.M.P.; Hidalgo, J.A.F. Capacidade combinatória e heterose em populaçoes de milho crioulo. Cienc. Rural 2008, 39, 332–339. [Google Scholar] [CrossRef]
- Halffter, G.; Matthews, E.G. The natural history of dung beetles of the subfamily Scarabaeinae (Coleoptera: Scarabaeidae). Folia Entomol. Mex. 1966, 12–14, 1–312. [Google Scholar]
- Cantil, L.F.; Sánchez, M.V.; Dinghi, P.A.; Genise, J.F. Food relocation behavior, nests, and brood balls of Canthon quinquemaculatus Laporte de Castelnau (Coleoptera: Scarabaeidae: Scarabaeinae). Coleopt. Bull. 2014, 68, 199–208. [Google Scholar] [CrossRef]
- Obrist, L.B.; Dutton, A.; Albajes, R.; Bigler, F. Exposure of arthropod predators to Cry1Ab toxin in Bt maize fields. Ecol. Entomol. 2006, 31, 143–154. [Google Scholar] [CrossRef]
- Thomé, V.M.R.; Zampieri, S.; Braga, H.J.; Pandolfo, C.; Silva, V.P.; Bacic, I.; Laus, J.; Soldateli, D.; Gebler, E.; Ore, J.D.; et al. Zoneamento Agroecológico e Socioeconômico de Santa Catarina; Epagri: Florianópolis, Brazil, 1999. Available online: http://ciram.epagri.sc.gov.br (accessed on 3 May 2016).
- Bookstein, F.L. Morphometric Tools for Landmark Data: Geometry and Biology, 1st ed.; Cambridge University Press: Cambridge, UK, 1991; 455p, ISBN 0521585988. [Google Scholar]
- Rohlf, F.J.; Slice, D. Extensions of the Procrustes method for optimal superimposition of landmarks. Syst. Zool. 1990, 39, 40–59. [Google Scholar] [CrossRef]
- Rohlf, F.J. TpsDig, Version 2.16. Department of Ecology and Evolution, State University of New York at Stony Brook, 2010. Available online: http://life.bio.sunysb.edu/morph/bibr28 (accessed on 20 May 2016).
- Rohlf, F.J. TpsUtil, File Utility Program. Version 1.53. Department of Ecology and Evolution, State University of New York at Stony Brook, 2012. Available online: http://life.bio.sunysb.edu/morph (accessed on 25 May 2016).
- Klingenberg, C.P. MorphoJ: An integrated software package for geometric morphometrics. Mol. Ecol. Resour. 2011, 11, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Murphy, A.F.; Seiter, N.J.; Krupke, C.H. The impact of Bt maize as a natal host on adult head capsule width in field populations of western corn rootworm. Entomol. Exp. Appl. 2011, 139, 8–16. [Google Scholar] [CrossRef]
- Peters, R.H. The Ecological Implications of Body Size, 1st ed.; Cambridge University Press: Cambridge, UK, 1983; 344p, ISBN 052128886X. [Google Scholar]
- Al-Deeb, M.A.; Wilde, G.E. Effect of bt corn expressing the Cry3Bb1 toxin on western corn rootworm (Coleoptera: Chrysomelidae) Biology. J. Kansas Entomol. Soc. 2005, 78, 142–152. [Google Scholar] [CrossRef]
- Lundgren, J.G.; Wiedenmann, R.N. Coleopteran-specific Cry3Bb toxin from transgenic corn pollen does not affect the fitness of a nontarget Species, Coleomegilla maculata DeGeer (Coleoptera: Coccinellidae). Environ. Entomol. 2002, 31, 1213–1218. [Google Scholar] [CrossRef]
- Cavicchi, S.; Giorgi, G.; Mochi, M. Investigation on early divergence between populations of Drosophila melanogaster kept at different temperatures. Genetica 1978, 48, 81–87. [Google Scholar] [CrossRef]
- Marsteller, S.; Adams, D.C.; Collyer, M.L.; Condon, M. Six cryptic species on a single species of host plant: Morphometric evidence for possible reproductive character displacement. Ecol. Entomol. 2009, 34, 66–73. [Google Scholar] [CrossRef]
- Alves, V.M.; Moura, M.O.; de Carvalho, C.J.B. Wing shape is influenced by environmental variability in Polietina orbitalis (Stein) (Diptera: Muscidae). Rev. Bras. Entomol. 2016, 60, 150–156. [Google Scholar] [CrossRef]
- Winans, G.A.; Nishioka, R.S. A multivariate description of change in body shape of coho salmon (Oncorhynchus kisutch) during smoltification. Aquaculture 1987, 66, 235–245. [Google Scholar] [CrossRef]
- McCollum, S.A.; Leimberger, J.D. Predator-induced morphological changes in an amphibian: Predation by dragonflies affects tadpole shape and color. Oecologia 1997, 109, 615–621. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Mak, R.; Ayad, H.; Smith, A.; Maclean, N. Expression of a novel piscine growth hormone gene results in growth enhancement in transgenic tilapia (Oreochromis niloticus). Transgenic Res. 1998, 7, 357–370. [Google Scholar] [CrossRef] [PubMed]
- Ostenfeld, T.H.; McLean, E.; Devlin, R.H. Transgenesis change body and head shape in Pacific salmon. J. Fish Biol. 1998, 52, 850–854. [Google Scholar] [CrossRef]
- Dunham, R.A.; Chatakondi, N.; Nichols, A.J.; Kucuktas, H.; Chen, T.T.; Powers, D.A.; Weetle, J.D.; Cummins, K.; Lovell, R.T. Effect of rainbow trout growth hormone complementary DNA on body shape, carcass yeld, and carcass composition of F1 and F2 transgenic common carp (Cyprinus carpio). Mar. Biotechnol. 2002, 4, 604–611. [Google Scholar] [CrossRef] [PubMed]
- Fares, N.H.; El-Sayed, A.K. Fine structural changes in the ileum of mice fed on endotoxin-treated potatoes and transgenic potatoes. Nat. Toxins 1998, 6, 219–233. [Google Scholar] [CrossRef]
- Armon, S.; Yanai, O.; Sharon, E. Quantitative phenotyping of leaf margins in three dimensions, demonstrated on KNOTTED and TCP trangenics in Arabidopsis. J. Exp. Bot. 2014, 65, 2071–2077. [Google Scholar] [CrossRef] [PubMed]
- Habes, D.; Messiad, R.; Gouasmia, S.; Grib, L. Effects of an inorganic insecticide (boric acid) against Blatella germanica: Morphometric measurements and biochemical composition of ovaries. Afr. J. Biotechnol. 2013, 12, 2492–2497. [Google Scholar]
Figure 1.
Landmarks (red dots) used in shape analysis of C. quinquemaculatus. (A) Lateral view; (B) Graphical representation of body shape based on 15 landmarks, adapted from Hernández et al. [8]. Landmarks in parentheses correspond to the same region on the other side of the body.
Figure 1.
Landmarks (red dots) used in shape analysis of C. quinquemaculatus. (A) Lateral view; (B) Graphical representation of body shape based on 15 landmarks, adapted from Hernández et al. [8]. Landmarks in parentheses correspond to the same region on the other side of the body.
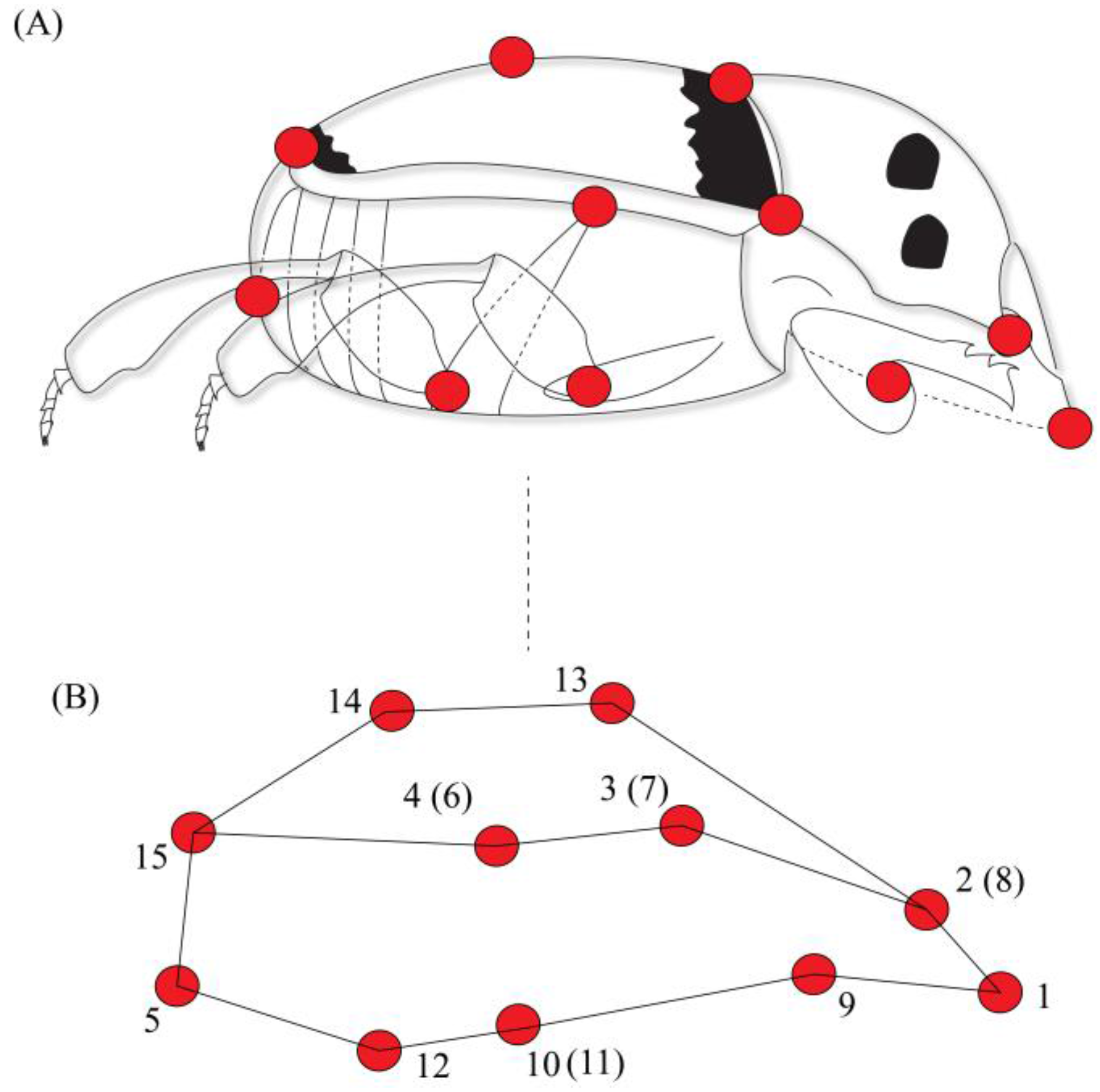
Figure 2.
Position of C. quinquemaculatus populations in morphometric space formed by two main axes of Canonical Variable Analysis, collected in the region of São Miguel do Oeste, SC, Brazil. Conventional (Co), Creole (Cr) and Transgenic (Tr); ellipses indicate a 95% confidence interval.
Figure 2.
Position of C. quinquemaculatus populations in morphometric space formed by two main axes of Canonical Variable Analysis, collected in the region of São Miguel do Oeste, SC, Brazil. Conventional (Co), Creole (Cr) and Transgenic (Tr); ellipses indicate a 95% confidence interval.
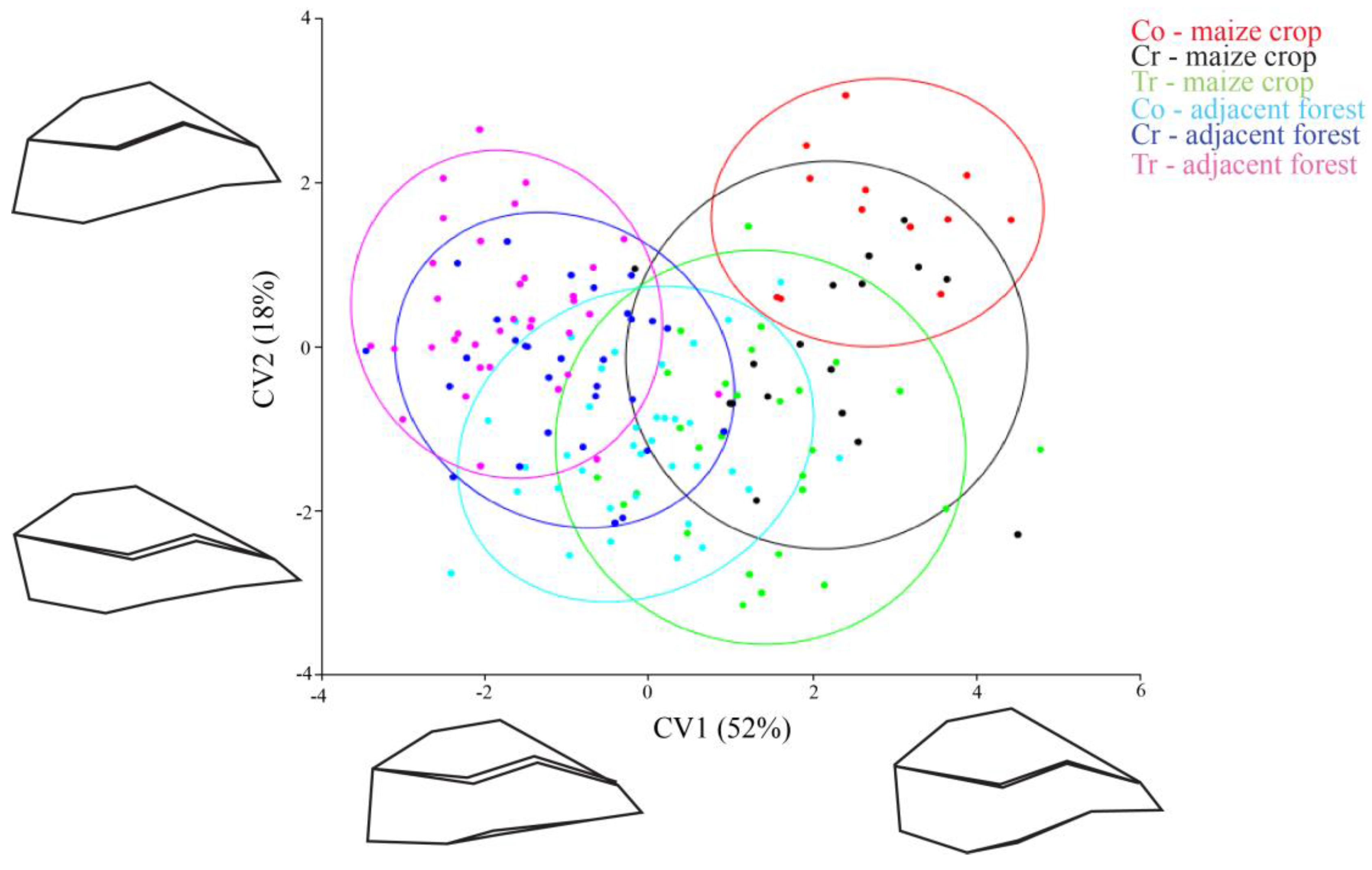
Figure 3.
(A) 2D Graphical reconstruction of C. quinquemaculatus body shape. Gray lines show body shape of transgenic maize crop populations, and black lines show adjacent forest populations; (B) 3D Graphical reconstruction of C. quinquemaculatus body shape. Green dots: transgenic maize populations. Pink dots: adjacent forest populations.
Figure 3.
(A) 2D Graphical reconstruction of C. quinquemaculatus body shape. Gray lines show body shape of transgenic maize crop populations, and black lines show adjacent forest populations; (B) 3D Graphical reconstruction of C. quinquemaculatus body shape. Green dots: transgenic maize populations. Pink dots: adjacent forest populations.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Percentage of correct classification of C. quinquemaculatus populations based on body morphology and discriminant analysis (DA).
Table 1.
Percentage of correct classification of C. quinquemaculatus populations based on body morphology and discriminant analysis (DA).
| Population | Allocation Value (%) |
|---|---|
| Conventional—Maize crop | 90.66 |
| Creole—Maize crop | 64.70 |
| Transgenic—Maize crop | 82.14 |
| Conventional—Adjacent forest | 58.33 |
| Creole—Adjacent forest | 73.33 |
| Transgenic—Adjacent forest | 82.85 |
| Overall classification accuracy | 75.33 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Alves, V.M.; Hernández, M.I.M. Morphometric Modifications in Canthon quinquemaculatus Castelnau 1840 (Coleoptera: Scarabaeinae): Sublethal Effects of Transgenic Maize? Insects 2017, 8, 115. https://doi.org/10.3390/insects8040115
AMA Style
Alves VM, Hernández MIM. Morphometric Modifications in Canthon quinquemaculatus Castelnau 1840 (Coleoptera: Scarabaeinae): Sublethal Effects of Transgenic Maize? Insects. 2017; 8(4):115. https://doi.org/10.3390/insects8040115
Chicago/Turabian StyleAlves, Victor Michelon, and Malva Isabel Medina Hernández. 2017. "Morphometric Modifications in Canthon quinquemaculatus Castelnau 1840 (Coleoptera: Scarabaeinae): Sublethal Effects of Transgenic Maize?" Insects 8, no. 4: 115. https://doi.org/10.3390/insects8040115
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.