
Chemical Compounds and Bioactivity of Aqueous Extracts of Alibertia spp. in the Control of Plutella xylostella L. (Lepidoptera: Plutellidae)



Abstract
:1. Introduction
2. Materials and Methods
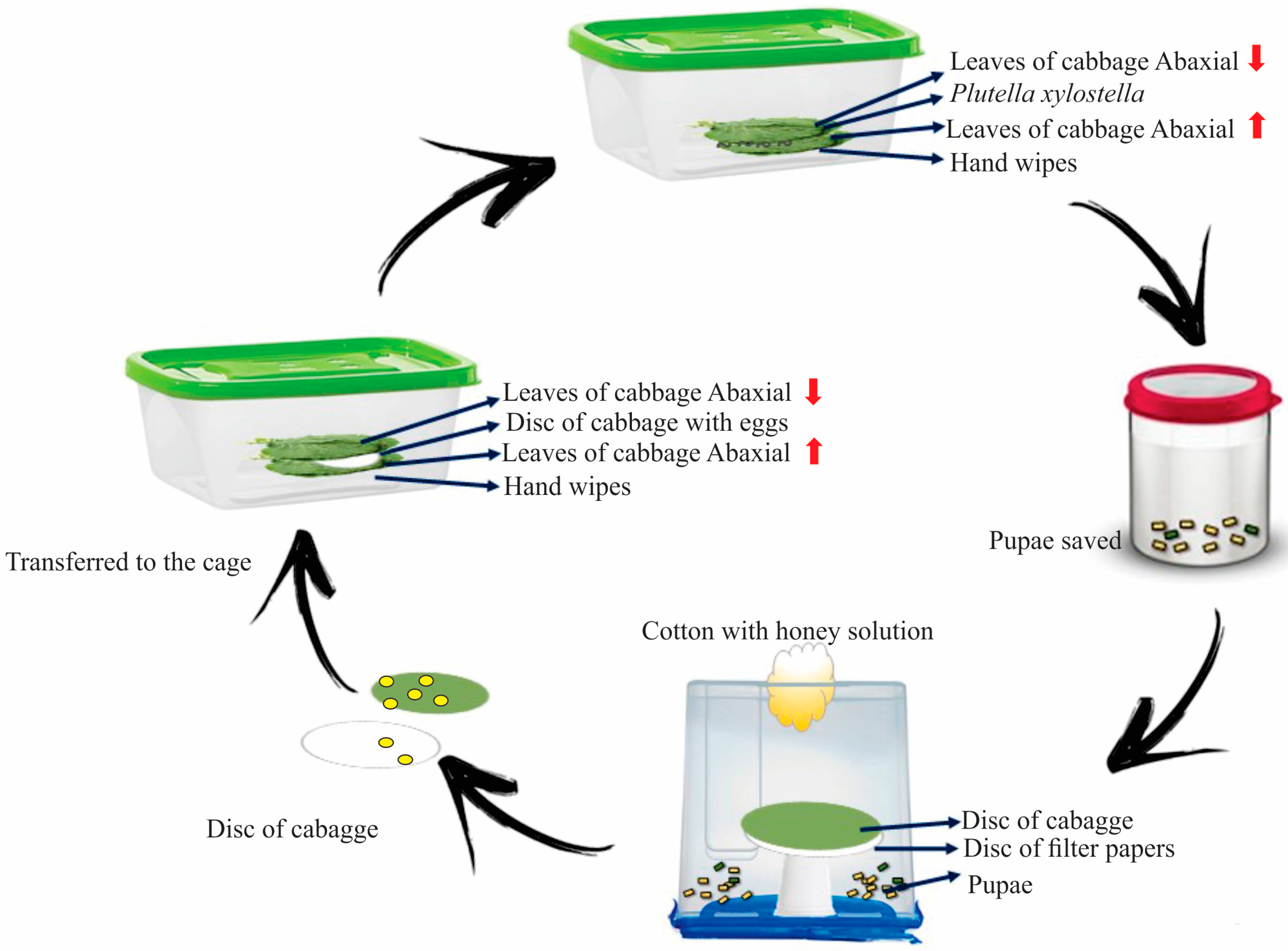
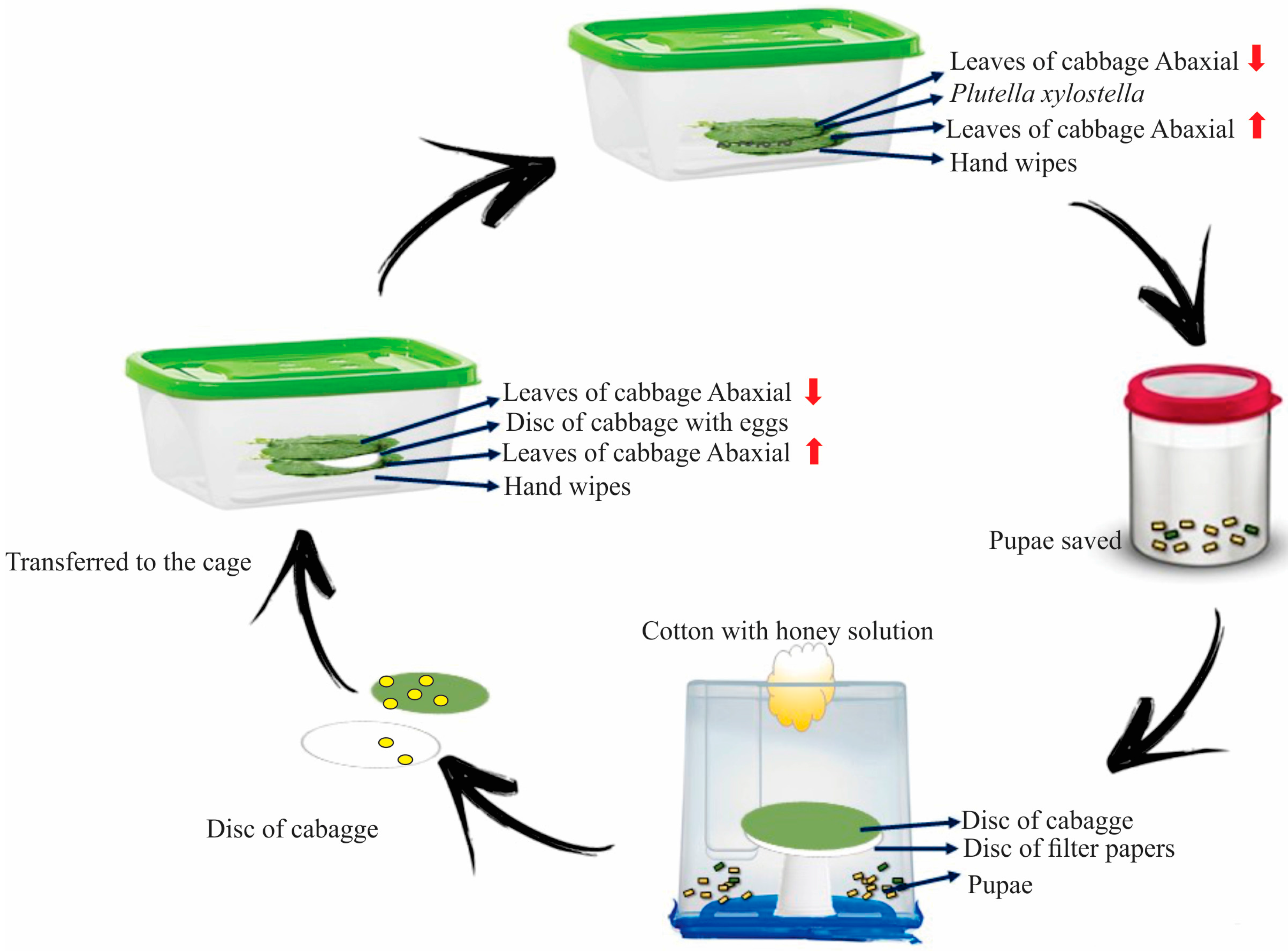
2.1. Rearing of Plutella xylostella
2.2. Preparation of Aqueous Extracts
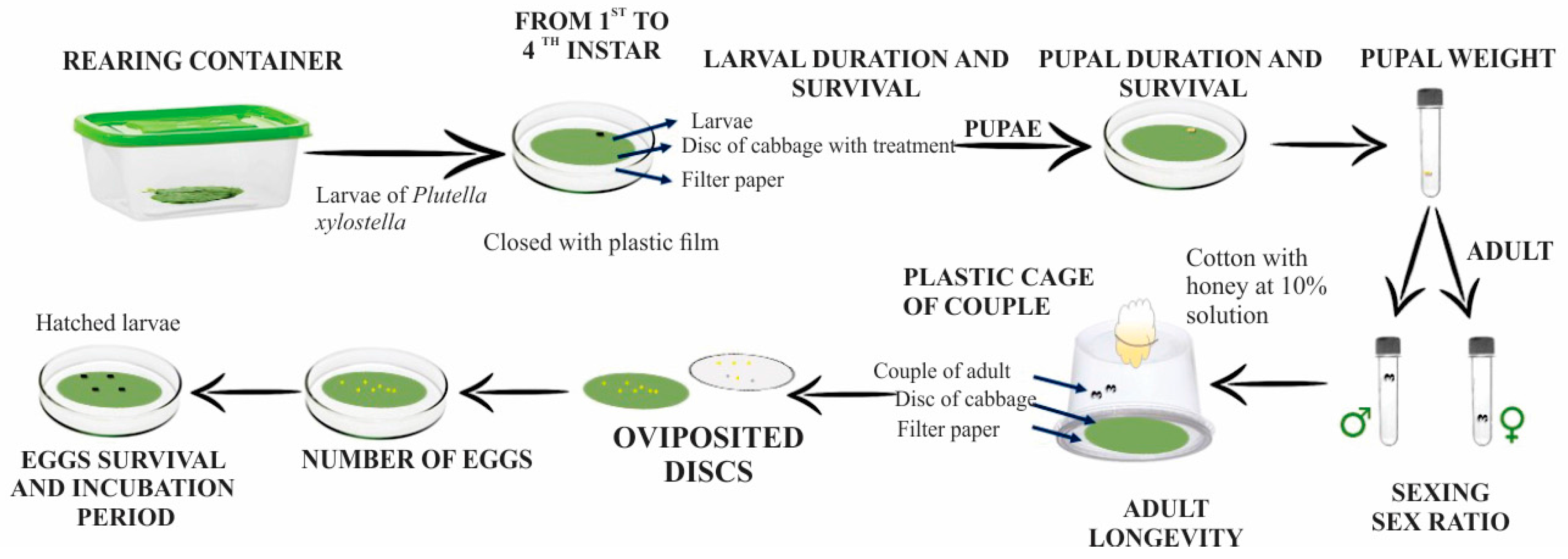
2.3. Bioactivity of A. edulis, A. intermedia, and A. sessilis Aqueous Extracts against P. xylostella
2.4. Identification of Compounds by High-Performance Liquid Chromatography (HPLC)
3. Results
3.1. Effect of the Extracts on the Development and Reproduction of P. xylostella
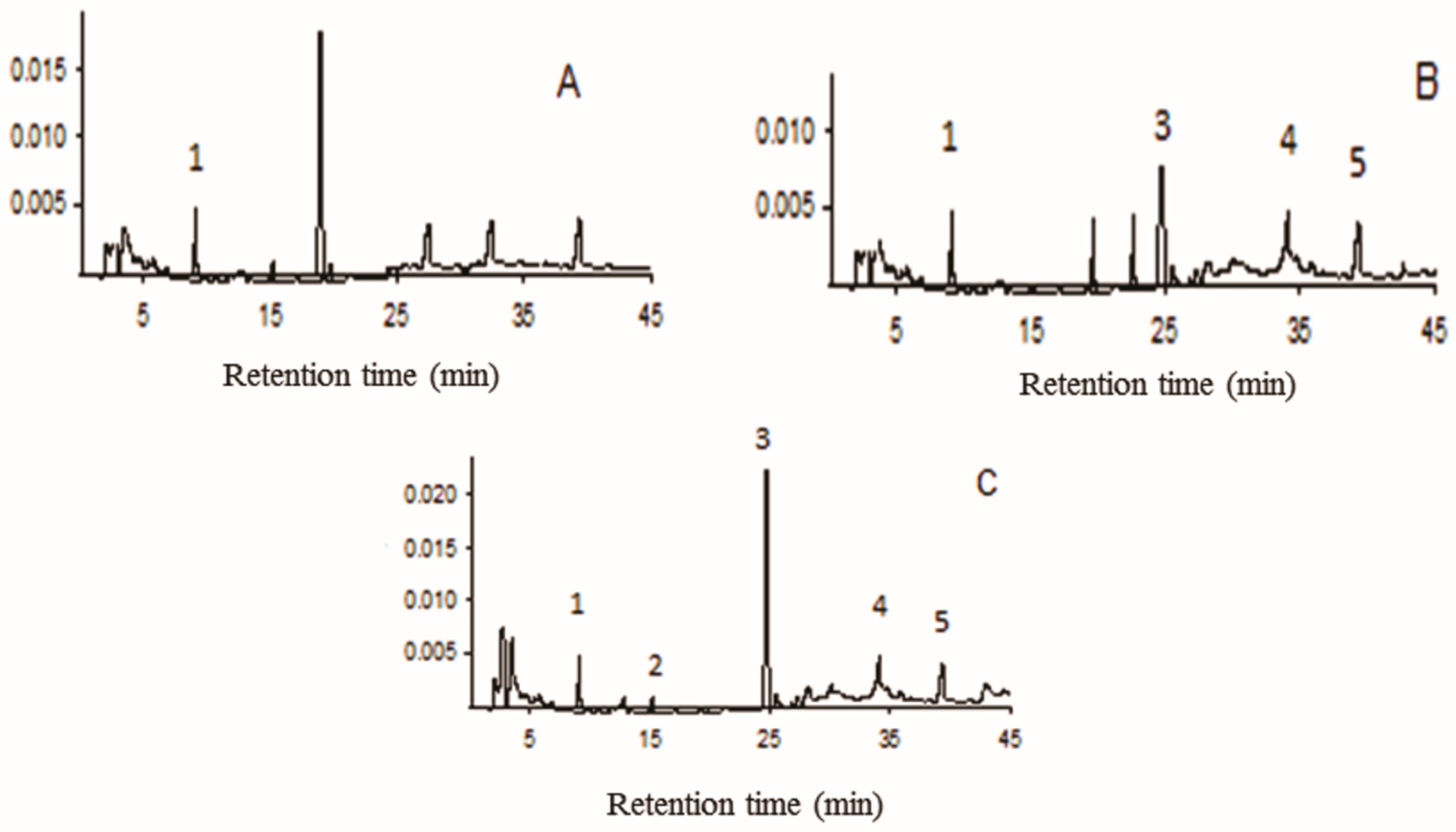
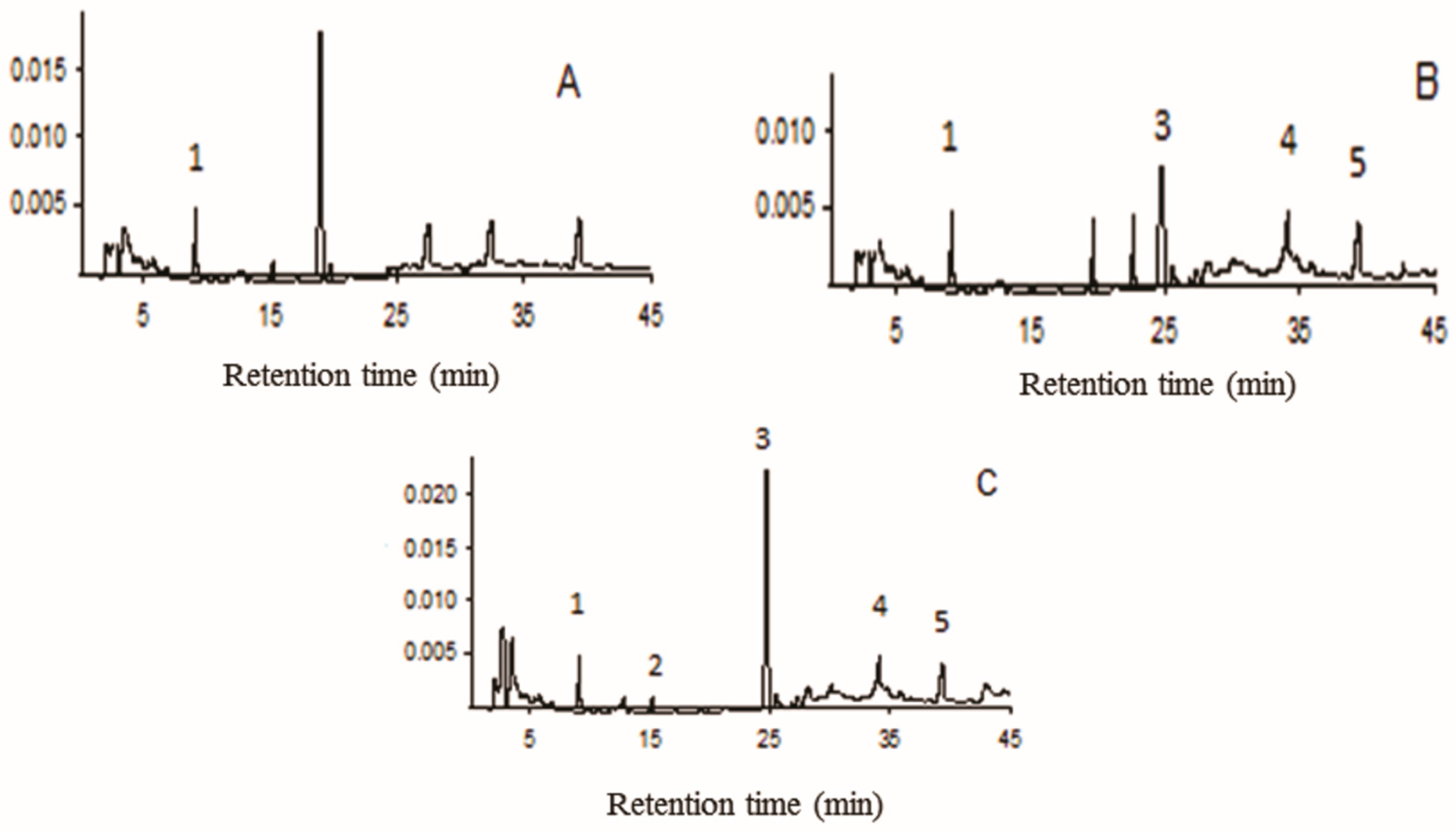
3.2. Compounds in Aqueous Extracts
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Furlong, M.J.; Wright, D.J.; Dosdall, L.M. Diamondback moth ecology and management: Problems, progress, and prospects. Annu. Rev. Entomol. 2013, 58, 517–541. [Google Scholar] [CrossRef] [PubMed]
- Zalucki, M.P.; Shabbir, A.; Silva, R.; Adamson, D.; Shu-Sheng, L.; Furlong, M.J. Estimating the economic cost of one of the world’s major insect pests, Plutella xylostella: Just how long is a piece of string? J. Econ. Entomol. 2012, 105, 1115–1129. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, A.J.; Endersby, N.M.; Ridland, P.M.; Zhang, J.; Neal, M. Effects of cultivar on oviposition preference, larval feeding and development time of diamondback moth, Plutella xylostella (L.) (Lepidoptera: Plutellidae), on some Brassica oleracea vegetables in Victoria. Aust. J. Entomol. 2005, 44, 284–287. [Google Scholar] [CrossRef]
- Talekar, N.S.; Shelton, A.M. Biology, ecology, and management of the diamondback moth. Annu. Rev. Entomol. 1993, 38, 275–301. [Google Scholar] [CrossRef]
- Pérez, C.J.; Alvarado, P.; Narváez, C.; Hernández, L.; Vanegas, H.; Hruska, A.; Shelton, A.M. Assessment of insecticide resistance in five insect pests attacking field and vegetable crops in Nicaragua. J. Econ. Entomol. 2000, 93, 1779–1787. [Google Scholar] [CrossRef] [PubMed]
- Castelo Branco, M.; Gatehouse, A.G. Insecticide resistance in Plutella xylostella (L.) (Lepidoptera: Yponomeutidae) in the Federal District, Brazil. An. Soc. Entomol. Bras. 1997, 26, 75–79. [Google Scholar] [CrossRef]
- Santos, V.C.; Siqueira, H.A.A.; Silva, J.E.; Farias, M.J.D.C. Insecticide resistance in populations of the diamondback moth, Plutella xylostella (L.) (Lepidoptera: Plutellidae), from the state of Pernambuco, Brazil. Neotrop. Entomol. 2011, 40, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Amoabeng, B.W.; Gurr, G.M.; Gitau, C.W.; Nicol, H.I.; Munyakazi, L.; Stevenson, P.C. Tri-Trophic insecticidal effects of African plants against cabbage pests. PLoS ONE 2013. [Google Scholar] [CrossRef]
- Wei, X.; Zhang, X.; Shen, D.; Wang, H.; Wu, Q.; Lu, P.; Qiu, Y.; Song, J.; Zhang, Y.; Li, X. Transcriptome analysis of Barbarea vulgaris infested with Diamondback Moth (Plutella xylostella) Larvae. PLoS ONE 2013. [Google Scholar] [CrossRef]
- Dong, X.; Zhai, Y.; Hu, M.; Zhong, G.; Huang, W.; Zheng, Z.; Han, P. Proteomic and properties analysis of botanical insecticide rhodojaponin III-Induced response of the Diamondback Moth, Plutella xyllostella (L.). PLoS ONE 2013. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, L.; Xu, X.; Freed, S.; Gao, Y.; Yu, J.; Wang, S.; Ju, W.; Zhang, Y.; Jin, F. Cecropins from Plutella xylostella and their interaction with Metarhizium anisopliae. PLoS ONE 2015. [Google Scholar] [CrossRef] [PubMed]
- Amoabeng, B.W.; Gurr, G.M.; Gitau, C.W.; Stevenson, P.C. Cost: Benefit analysis of botanical insecticide use in cabbage: Implications for smallholder farmers in developing countries. Crop Prot. 2014, 57, 71–76. [Google Scholar] [CrossRef]
- Kudom, A.A.; Mensah, B.A.; Botchey, M.A. Aqueous neem extract versus neem powder on Culex quinquefasciatus: Implications for control in anthropogenic habitats. J. Insect Sci. 2011, 11, 142. [Google Scholar] [CrossRef] [PubMed]
- Ladhari, A.; Laarif, A.; Omezzine, F.; Haouala, R. Effect of the extracts of the spiderflower, Cleome arabica, on feeding and survival of larvae of the cotton leafworm, Spodoptera littoralis. J. Insect Sci. 2013, 13, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Schmutterer, H. Potential of azadirachtin-containing pesticides for integrated pest control in developing and industrialized countries. J. Insect Physiol. 1988, 34, 713–719. [Google Scholar] [CrossRef]
- Mordue, A.J.; Blackwell, A. Azadirachtin: An update. J. Insect Physiol. 1993, 39, 903–924. [Google Scholar] [CrossRef]
- Deletre, E.; Martin, T.; Campagne, P.; Bourguet, D.; Cadin, A.; Menut, C.; Bonafos, R.; Chandre, F. Repellent, irritant and toxic effects of 20 plant extracts on adults of the malaria vector Anopheles gambiae Mosquito. PLoS ONE 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krinski, D.; Massaroli, A.; Machado, M. Potencial inseticida de plantas da família Annonaceae. Rev. Bras. Frutic. 2014, 36, 225–242. (In Portuguese) [Google Scholar] [CrossRef]
- Couto, I.F.S.; Fuchs, M.L.; Pereira, F.F.; Mauad, M.; Scalon, S.P.Q.; Dresch, D.M.; Mussury, R.M. Feeding preference of Plutella xylostella for leaves treated with plant extracts. An. Acad. Bras. Cienc. 2016, 88, 1781–1789. [Google Scholar] [CrossRef] [PubMed]
- Luciano, J.H.S.; Lima, M.A.S.; Souza, E.B.; Silveira, E.R. Chemical constituents of Alibertia myrciifolia Spruce ex K. Schum. Biochem. Syst. Ecol. 2004, 32, 1227–1229. [Google Scholar] [CrossRef]
- Schripsema, J.; Dagnina, D.; Grosman, G. Alcalóides indólicos. In Farmacognosia da Planta ao Medicamento, 5th ed.; Simões, C.M.O., Schenkel, E.P., Gosmann, G., Mello, J.C.P., Mentz, L.A., Petrovick, P.R., Eds.; Editora da UFRGS/UFSC: Porto Alegre/Florianópolis, Brasil, 2004; pp. 819–846. (In Portuguese) [Google Scholar]
- Keutgen, A.J.; Pawelzik, E. Modifications of strawberry fruit antioxidant pools and fruit quality under NaCl stress. J. Agric. Food Chem. 2007, 55, 4066–4072. [Google Scholar] [CrossRef] [PubMed]
- Bolzani, V.S.; Trevisan, L.M.V.; Young, M.C.M. Caffeic acid esters and triterpenes of Alibertia macrophylla. Phytochemistry 1991, 30, 2089–2091. [Google Scholar] [CrossRef]
- Olea, R.S.G.; Roque, N.F.; Bolzani, V.S. Acylated flavonol glycosides and terpenoids from the leaves of Alibertia sessilis. J. Braz. Chem. Soc. 1997, 8, 257–259. [Google Scholar] [CrossRef]
- Moura, V.M.; Santos, D.P.; Santin, S.M.O.; Carvalho, J.E.; Foglio, M.A. Constituintes químicos de Galianthe brasiliensis (Rubiaceae). Quim. Nova 2006, 29, 452–455. (In Portuguese) [Google Scholar] [CrossRef]
- Lebrini, M.; Robert, F.; Roos, C. Alkaloids extract from Palicourea guianensis plant as corrosion inhibitor for C38 steel in 1 M hydrochloric acid medium. Int. J. Electrochem. Sci. 2011, 6, 847–859. [Google Scholar]
- Ling, S.K.; Komorita, A.; Tanaka, T.; Fujioka, T.; Mihashi, K.; Kouno, I. Iridoids and anthraquinones from the Malaysian medicinal plant, Saprosma scortechinii (Rubiaceae). Chem. Pharm. Bull. 2002, 50, 1035–1040. [Google Scholar] [CrossRef] [PubMed]
- Silva, V.C.; Silva, G.H.; Bolzani, V.S.; Lopes, M.N. Isolation of lignans glycosides from Alibertia sessilis (Vell.) K. Schum. (Rubiaceae) by preparative high-performance liquid chromatography. Eclet. Quim. 2006, 31, 55–58. [Google Scholar] [CrossRef]
- Silva, W.C.; Pinheiro, C.C.S.; Rodrigues, J.M.G.; Souza, H.E.M.; Ribeiro, J.D. Avaliação do efeito tóxico de extratos de Palicourea marcgravii St. Hil. (Rubiaceae) sobre Aetalion sp. (Hemiptera: Aetalionidae) em laboratório. R. Bras. Bioci. 2009, 7, 129–133. (In Portuguese) [Google Scholar]
- Gonzaga, A.D.; Ribeiro, J.D.; Vieira, M.F.; Alécio, M.R. Toxidez de três concentrações de erva-de-rato (Palicourea marcgravii A. St.-Hill) e Manipueira (Manihot esculenta Crantz) em pulgão verde dos Citros (Aphis spiraecola Patch) em casa de vegetação. R. Bras. Bioc. 2007, 5, 55–56. (In Portuguese) [Google Scholar]
- Gao, G.; Lu, Z.; Tao, S.; Zhang, S.; Wang, F. Triterpenoid saponins with antifeedant activities from stem bark of Catunaregam spinosa (Rubiaceae) against Plutella xylostella (Plutellidae). Carbohydr. Res. 2011, 346, 2200–2205. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, M. Botanical pesticides: Past, present and future. In Insecticides of Plant Origin, 1st ed.; Arnason, J.T., Philogene, B.J.R., Morand, P., Eds.; America Chemical Society: Washington, DC, USA, 1989; pp. 1–10. [Google Scholar]
- Barros, R.; Thuler, R.T.; Pereira, F.F. Técnica de criação de Plutella xylostella (L. 1758) (Lepidoptera: Yponomeutidae). In Técnicas de Criação de Pragas de Importância Agrícola, em Dietas Naturais, 1st ed.; Pratissoli, D., Ed.; Edufes: Vitória, Brasil, 2012; pp. 65–84. (In Portuguese) [Google Scholar]
- Matias da Silva, R.; Fioratti, C.A.G.; Silva, G.B.; Cardoso, C.A.L.; Miranda, L.O.; Mauad, M.; Mussury, R.M. Mussury, R.M. Antibiose do extrato foliar de Duguetia furfuracea sobre Plutella xylostella (Lepidoptera: Plutellidae). In Temas Atuais em Ecologia Comportamental e Interações, Anais do II BecInt–Behavioral Ecology and Interactions Symposium, 1st ed.; Calixto, E.S., Toreza-Silingardi, H.M., Eds.; Editora Composer: Uberlândia, MG, Brasil, 2017; pp. 52–69. (In Portuguese) [Google Scholar]
- Duffey, S.S.; Isman, M.B. Inhibition of insect larval growth by phenolics in glandular trichomes of tomato leaves. Experientia 1981, 37, 575–576. [Google Scholar] [CrossRef]
- Isman, M.B.; Duffey, S.S. Toxicity of tomato phenolic compounds to the fruitworm, Heliothis zea. Entomol. Exp. Appl. 1982, 31, 370–376. [Google Scholar] [CrossRef]
- Klocke, J.A.; Kubo, I. Symposium: Insect Behavioral Ecology—′90: Defense of plants throügh regulation of insect feeding behavior. Fla. Entomol. 1991, 74, 18–23. [Google Scholar] [CrossRef]
- Summers, C.B.; Felton, G.W. Pro oxidant effects of phenolic acids on the generalist herbivore Helicoverpa zea (Lepidoptera: Noctuidae): Potential mode of action for phenolic compounds in plant anti-herbivore chemistry. Insect Biochem. Mol. Biol. 1994, 24, 943–953. [Google Scholar] [CrossRef]
- Pan, L.; Ren, L.; Chen, F.; Feng, Y.; Luo, Y. Antifeedant Activity of Ginkgo Biloba secondary metabolites against Hyphantria cunea Larvae: Mechanisms and applications. PLoS ONE 2016. [Google Scholar] [CrossRef] [PubMed]
- Rani, P.U.; Ravibabu, M.V. Allelochemicals in castor (Ricinus communis L.) plants and their impact on pest larval feeding as anti-herbivore defensive. Allelopath. J. 2011, 27, 263–275. [Google Scholar]
- Berenbaum, M. Adaptive significance of midgut pH in larval lepidoptera. Am. Nat. 1980, 115, 138–146. [Google Scholar] [CrossRef]
- Appel, H.M. Phenolics in ecological interactions: The importance of oxidation. J. Chem. Ecol. 1993, 19, 1521–1552. [Google Scholar] [CrossRef] [PubMed]
- Santiago, R.; Malvar, R.A.; Baamonde, M.D.; Revilla, P.; Souto, X.C. Free phenols in maize pith and their relationship with resistance to Sesamia nonagrioides (Lepidoptera: Noctuidae) attack. J. Econ. Entomol. 2005, 98, 1349–1356. [Google Scholar] [CrossRef] [PubMed]
- Arnason, J.T.; Gale, J.; Conilh de Beyssac, B.; Sen, A.; Miller, S.S.; Philogene, B.J.R.; Lambert, J.D.H.; Fulcher, R.G.; Serratos, A.; Mihm, J. Role of phenolics in resistance of maize grain to the stored grain insects, Prostephanus truncatus (Horn) and Sitophilus zeamais (Motsch.). J. Stored Prod. Res. 1992, 28, 119–126. [Google Scholar] [CrossRef]
- Tavares, W.S.; Pereira, A.I.A.; Freitas, S.S.; Serrão, J.E.; Zanuncio, J.C. The chemical exploration of Dimorphandra mollis (Fabaceae) in Brazil, with emphasis on insecticidal response: A review. J. Sci. Ind. Res. 2014, 73, 465–468. [Google Scholar]
- Piubelli, G.C.; Hoffmann-Campo, C.B.; Moscardi, F.; Miyakubo, S.H.; Oliveira, M.C.N. Baculovirus-resistant Anticarsia gemmatalis responds differently to dietary rutin. Entomol. Exp. Appl. 2006, 119, 53–60. [Google Scholar] [CrossRef]
- Hoffmann-Campo, C.B.; Ramos Neto, J.A.; Oliveira, M.C.N.; Oliveira, L.J. Detrimental effect of rutin on Anticarsia gemmatalis. Pesqui. Agropecu. Bras. 2006, 41, 1453–1459. [Google Scholar] [CrossRef]
- Stamp, N.E.; Skrobola, C.M. Failure to avoid rutin diets results in altered food utilization and reduced growth rate of Manduca sexta larvae. Entomol. Exp. Appl. 1993, 68, 127–142. [Google Scholar] [CrossRef]
- Silva, T.R.F.B.; Almeida, A.C.S.; Moura, T.L.; Silva, A.R.; Freitas, S.S.; Jesus, F.G. Effect of the flavonoid rutin on the biology of Spodoptera frugiperda (Lepidoptera: Noctuidae). Acta Sci. Agron. 2016, 38, 165–170. [Google Scholar] [CrossRef]
- Gazzoni, D.L.; Hulsmeyer, A.; Hoffmann-Campo, C.B. Efeito de diferentes doses de rutina e de quercetina na biologia de Anticarsia gemmatalis. Pesqui. Agropecu. Bras. 1997, 32, 673–681. (In Portuguese) [Google Scholar]
- Tabashnik, B.E. Plant secondary compounds as oviposition deterrents for cabbage butterfly, Pieris rapae (Lepidoptera: Pieridae). J. Chem. Ecol. 1987, 13, 309–316. [Google Scholar] [CrossRef] [PubMed]
- War, A.R.; Paulraj, M.G.; Ahmad, T.; Buhroo, A.A.; Hussain, B.; Ignacimuthu, S.; Sharma, H.C. Mechanisms of plant defense against insect herbivores. Plant Signal Behav. 2012, 7, 1306–1320. [Google Scholar] [CrossRef] [PubMed]
- Goławska, S.; Łukasik, I.; Kapusta, I.; Janda, B. Do the contents of luteolin, tricin, and chrysoeriol glycosides in Alfalfa (Medicago sativa L.) affect the behavior of Pea Aphid (Acyrthosiphon pisum)? Pol. J. Environ. Stud. 2012, 21, 1613–1619. [Google Scholar]
- Simmonds, M.S.J. Flavonoid-insect interactions: Recent advances in our knowledge. Phytochemistry 2003, 64, 21–30. [Google Scholar] [CrossRef]
- Goławska, S.; Sprawka, I.; Lukasik, I.; Golawski, A. Are naringenin and quercetin useful chemicals in pest-management strategies? J. Pest Sci. 2014, 87, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Kashiwagi, T.; Horibata, Y.; Mekuria, D.B.; Tebayashi, S.I.; Kim, C.S. Ovipositional deterrent in the sweet pepper, Capsicum annuum, at the Mature Stage against Liriomyza trifolii (Burgess). Biosci. Biotechnol. Biochem. 2005, 69, 1831–1835. [Google Scholar] [CrossRef] [PubMed]
- Diaz Napal, G.N.; Defagó, M.T.; Valladares, G.R.; Palacios, S.M. Response of Epilachna paenulata to two flavonoids, pinocembrin and quercetin, in a comparative study. J. Chem. Ecol. 2010, 36, 898–904. [Google Scholar] [CrossRef] [PubMed]
- Diaz Napal, G.N.; Palacios, S.M. Bioinsecticidal effect of the flavonoids pinocembrin and quercetin against Spodoptera frugiperda. J. Pest Sci. 2015, 88, 629–635. [Google Scholar] [CrossRef]
- Tringali, C.; Spatafora, C.; Cali, V.; Simmonds, M.S.J. Antifeedant constituents from Fagara macrophylla. Fitoterapia 2001, 72, 538–543. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Larval Duration (Days) | Larval Survival (%) | Pupal Duration (Days) | Pupal Survival (%) | Pupal Weight (mg) |
|---|---|---|---|---|---|
| Control | 5.86 ± 0.21 b * n = 50 | 96.67 ± 4.3 a n = 50 | 6.08 ± 0.17 a n = 46 | 98.49 ± 4.44 a n = 46 | 5.26 ± 0.3 a n = 46 |
| A. edulis | 4.72 ± 0.15 c n = 50 | 92.46 ± 3.3 a n = 50 | 5.67 ± 0.12 a n = 44 | 97.73 ± 3.59 a n = 44 | 4.61 ± 0.4 ab n = 44 |
| A. intermedia | 8.97 ± 0.19 a n = 50 | 56.41 ± 6.5 b n = 50 | 3.45 ± 0.12 b n = 28 | 96.34 ± 5.83 a n = 28 | 3.88 ± 0.2 bc n = 28 |
| A. sessilis | 8.21 ± 0.13 a n = 50 | 49.80 ± 4.5 b n = 50 | 2.36 ± 0.64 b n = 25 | 37.19 ± 11.7 b n = 11 | 3.46 ± 0.2 c n = 11 |
| CV (%) | 3.9 | 21.64 | 16.4 | 30.55 | 20.6 |
| Treatment | Longevity of Males (Days) | Longevity of Females (Days) | Fecundity | Survival of Eggs (%) |
|---|---|---|---|---|
| Control | 13.26 ± 1.41 b * n = 8 | 12.43 ± 1.19 a n = 8 | 198.08 ± 9.44 a | 91.75 ± 1.14 a |
| A. edulis | 19.37 ± 1.14 a n = 8 | 12.81± 0.66 a n = 8 | 187.57 ± 17.53 a | 71.04 ± 4.07 ab |
| A. intermedia | 14.66 ± 0.86 b n = 8 | 12.39 ± 0.90 a n = 8 | 86.69 ± 18.46 b | 66.24 ± 9.53 b |
| A. sessilis | 10.49 ± 0.50 b n = 2 | 8.97 ± 1.0 a n = 2 | 95.38 ± 6.5 ab | 77.41 ± 3.95 ab |
| CV (%) | 10.15 | 10.1 | 20.36 | 21.97 |
| Peaks | Compound | Retention Time (min) | Concentration (mg/g ± SD) A | Concentration (mg/g ± SD) B | Concentration (mg/g ± SD) C |
|---|---|---|---|---|---|
| 1 | Caffeic acid | 10.1 | 51.8 ± 0.1 | 48.9 ± 0.3 | 47.8 ± 0.2 |
| 2 | p-Coumaric acid | 15.2 | - | - | 10.7 ± 0.4 |
| 3 | Rutin | 24.7 | - | 88.9 ± 0.3 | 178.9 ± 0.6 |
| 4 | Quercetin | 34.5 | - | 25.7 ± 0.1 | 23.9 ± 0.1 |
| 5 | Luteolin | 39.7 | - | 32.5 ± 0.2 | 33.1 ± 0.1 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peres, L.L.S.; Sobreiro, A.I.; Couto, I.F.S.; Silva, R.M.; Pereira, F.F.; Heredia-Vieira, S.C.; Cardoso, C.A.L.; Mauad, M.; Scalon, S.P.Q.; Verza, S.S.; et al. Chemical Compounds and Bioactivity of Aqueous Extracts of Alibertia spp. in the Control of Plutella xylostella L. (Lepidoptera: Plutellidae). Insects 2017, 8, 125. https://doi.org/10.3390/insects8040125
Peres LLS, Sobreiro AI, Couto IFS, Silva RM, Pereira FF, Heredia-Vieira SC, Cardoso CAL, Mauad M, Scalon SPQ, Verza SS, et al. Chemical Compounds and Bioactivity of Aqueous Extracts of Alibertia spp. in the Control of Plutella xylostella L. (Lepidoptera: Plutellidae). Insects. 2017; 8(4):125. https://doi.org/10.3390/insects8040125
Chicago/Turabian StylePeres, Lucas L. S., Ana I. Sobreiro, Irys F. S. Couto, Rosicléia M. Silva, Fabricio F. Pereira, Silvia C. Heredia-Vieira, Claudia A. L. Cardoso, Munir Mauad, Silvana P. Q. Scalon, Sandra S. Verza, and et al. 2017. "Chemical Compounds and Bioactivity of Aqueous Extracts of Alibertia spp. in the Control of Plutella xylostella L. (Lepidoptera: Plutellidae)" Insects 8, no. 4: 125. https://doi.org/10.3390/insects8040125