Oxygen Consumption and Acoustic Activity of Adult Callosobruchus maculatus (F.) (Coleoptera: Chrysomelidae: Bruchinae) during Hermetic Storage



Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Rearing and Grain Infestation
2.2. Acoustic Probe Installation
2.3. Oxygen Monitoring
2.4. Estimation of Available Oxygen in the Experimental Glass Jars
2.5. Estimation of Available Oxygen and Initial Infestation Level in a PICS Bag
2.6. Acoustic Activity Monitoring
2.7. Signal Processing
2.8. Statistical Analysis
3. Results
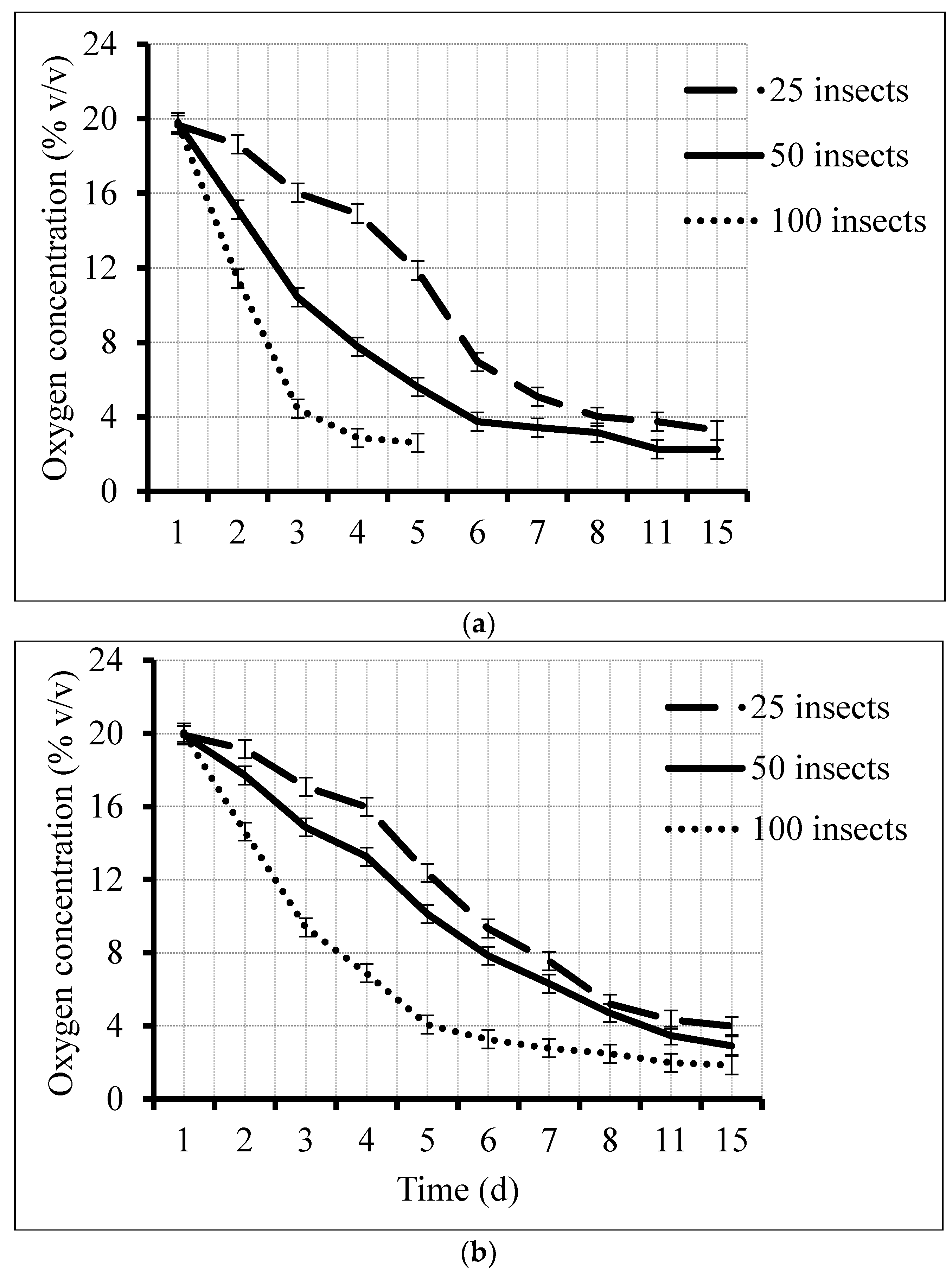
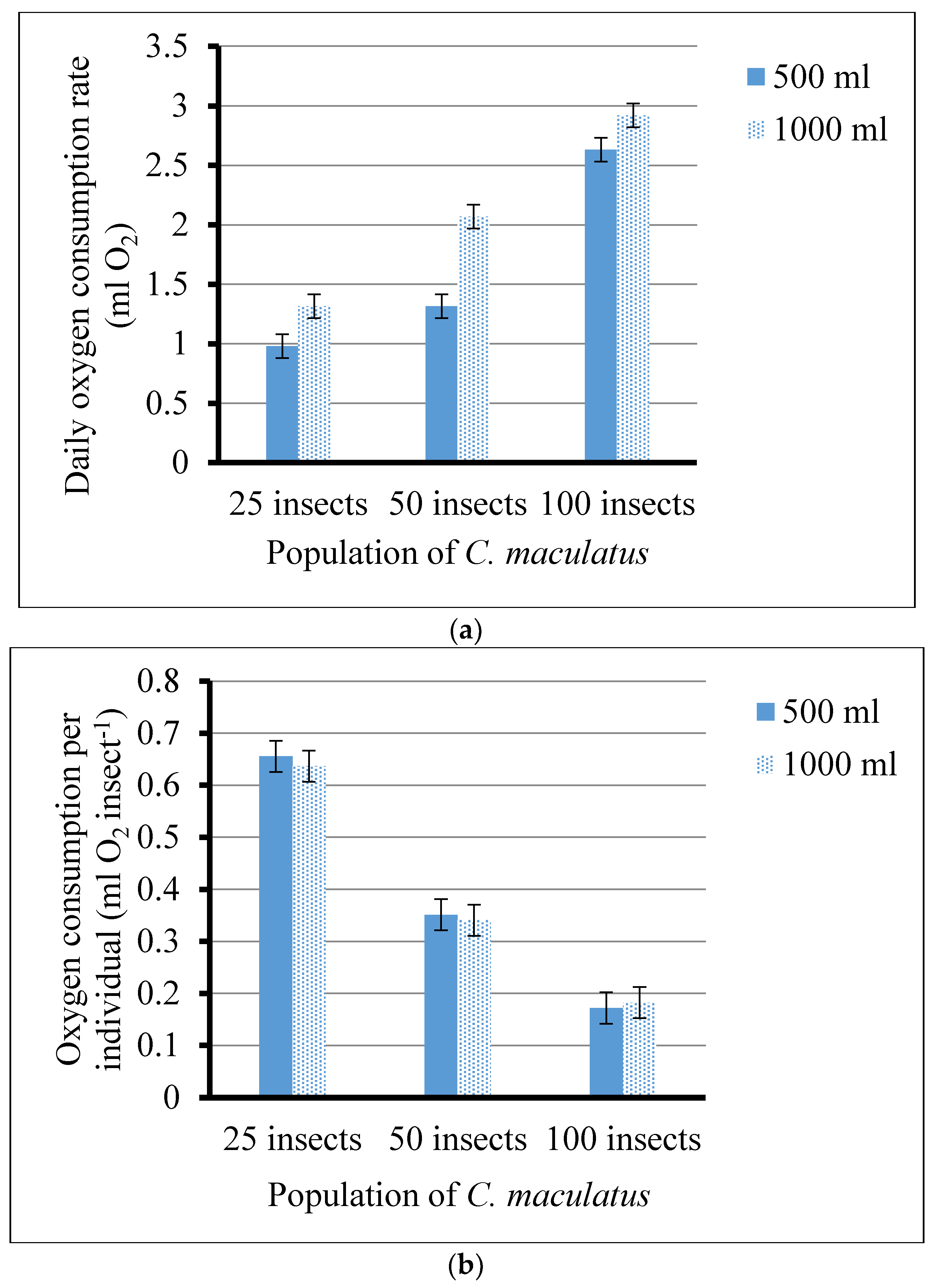
3.1. Oxygen Depletion Patterns and Rates
3.2. Temperature and Relative Humidity Variation
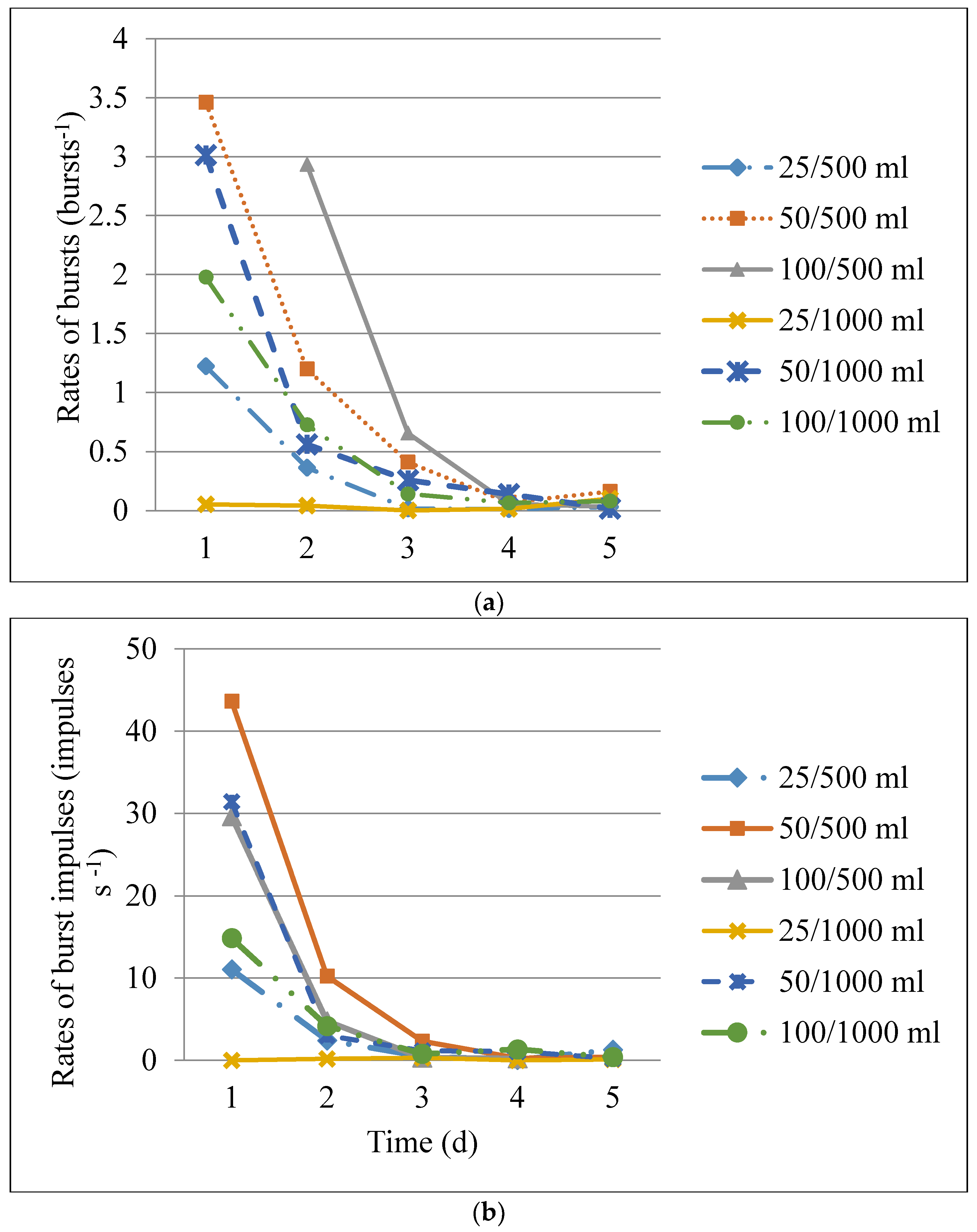
3.3. Acoustic Activity of C. maculatus during Hermetic Storage
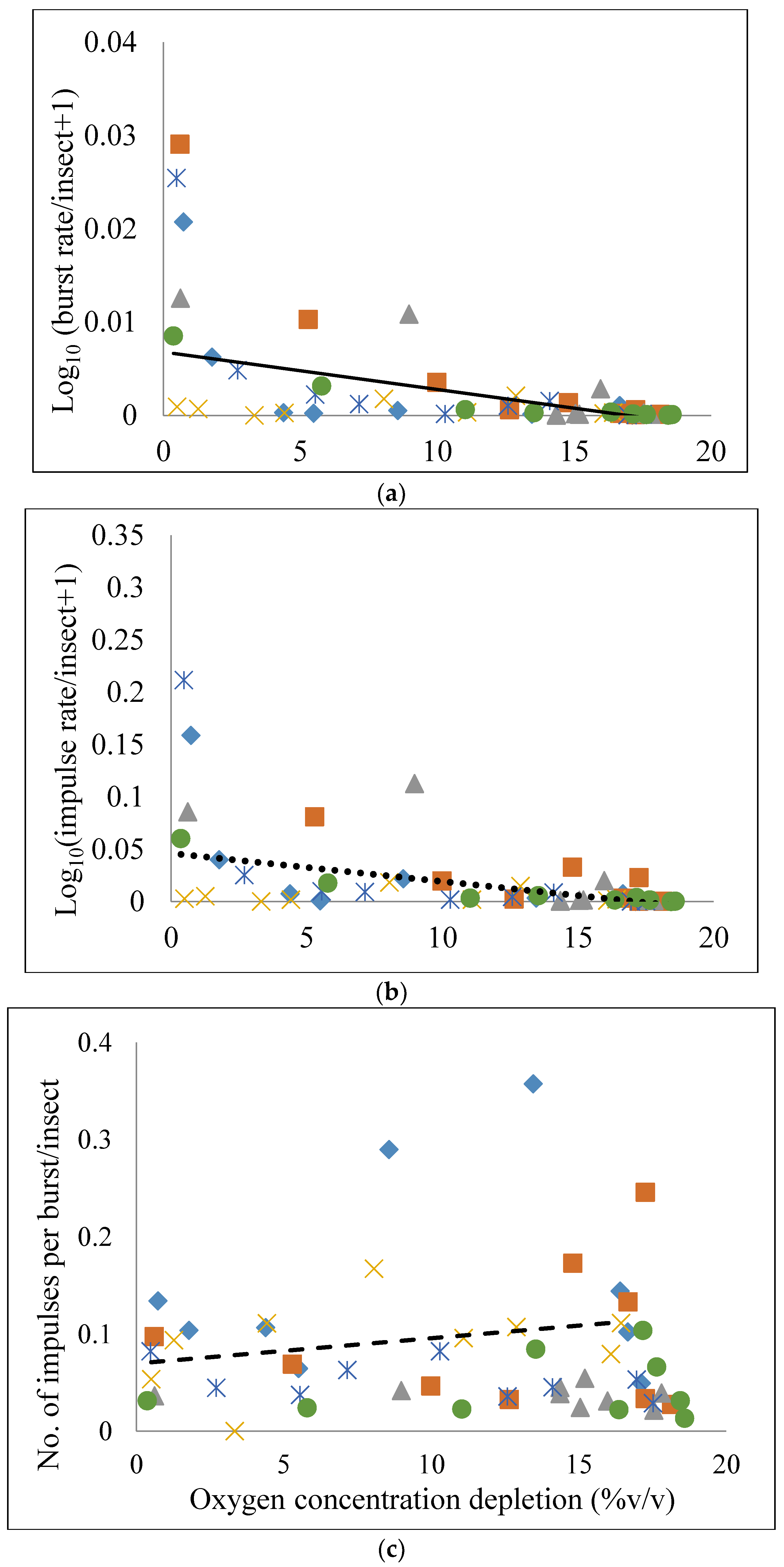
3.4. Regression Analysis
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Nedumaran, S.; Abinaya, P.; Jyosthnaa, P.; Shraavya, B.; Rao, P.; Bantilan, C. Grain Legumes Production, Consumption and Trade Trends in Developing Countries; Working Paper; Series Number 60; International Crops Research Institute for the Semi-Arid Tropics: Patancheru, Telangana, India, 2015; p. 64. [Google Scholar]
- Singh, R.J. (Ed.) Genetic Resources, Chromosome Engineering, and Crop Improvement: Vegetable Crops; CRC Press: Boca Raton, FL, USA, 2006; Volume 3. [Google Scholar]
- Langyintuo, A.S.; Lowenberg-DeBoer, J.; Faye, M.; Lambert, D.; Ibro, G.; Moussa, B.; Ntoukam, G. Cowpea supply and demand in West and Central Africa. Field Crops Res. 2003, 82, 215–231. [Google Scholar] [CrossRef]
- Carlos, G. Cowpea: Post Harvest Operation; Food and Agricultural Organization of United Nations: Rome, Italy, 2000. [Google Scholar]
- Hill, S.B. Pest Control—Cultural Control of Insects. Cultural Methods of Pest, Primarily Insect Control; EAP Ecological Agricultural Projects; Publication-58; Macdonald College of McGill University: Sainte-Anne-de-Bellevue, QC, Canada, 1989. [Google Scholar]
- Mbata, G.N.; Hetz, S.K.; Reichmuth, C.; Adler, C. Tolerance of pupae and pharate adults of Callosobruchus subinnotatus Pic (Coleoptera: Bruchidae) to modified atmospheres: A function of metabolic rate. J. Insect Physiol. 2000, 46, 145–151. [Google Scholar] [CrossRef]
- Seck, D.; Lognay, G.; Haubruge, E.; Marlier, M.; Gaspar, C. Alternative protection of cowpea seeds against Callosobruchus maculatus (F.) (Coleoptera: Bruchidae) using hermetic storage alone or in combination with Boscia senegalensis (Pers.) Lam ex Poir. J. Stored Prod. Res. 1996, 32, 39–44. [Google Scholar] [CrossRef]
- Ntoukam, G.; Murdock, L.L.; Shade, R.E.; Kitch, L.W.; Endondo, C.; Ousmane, B.; Wolfson, J. Managing insect Pests of cowpea in storage. In Proceedings of the Midcourse 2000 Research Meeting of Bean/Cowpea, Dakar, Senegal, 9–14 April 2000; pp. 3–4. [Google Scholar]
- Johnson, J.A.; Valero, K.A. Use of Commercial Freezers to Control Cowpea Weevil, Callosobruchus maculatus (Coleoptera: Bruchidae), in Organic Garbanzo Beans. J. Econ. Entomol. 2003, 96, 1952–1957. [Google Scholar] [CrossRef] [PubMed]
- Baoua, I.B.; Margam, V.; Amadou, L.; Murdock, L.L. Performance of Triple Bagging Hermetic Technology for Postharvest Storage of Cowpea Grain in Niger. J. Stored Prod. Res. 2012, 51, 81–85. [Google Scholar] [CrossRef]
- Bailey, S.W. Air-Tight Storage of Grain; Its Effects on Insect Pests. I. Calandra granaria L. (Coleoptera, Curculionidae). Aust. J. Agric. Res. 1955, 6, 33–51. [Google Scholar] [CrossRef]
- Murdock, L.L.; Baoua, I.B. On Purdue Improved Cowpea Storage (PICS) technology: Background, mode of action, future prospects. J. Stored Prod. Res. 2014, 58, 3–11. [Google Scholar] [CrossRef]
- Baoua, I.B.; Amadou, L.; Baributsa, D.; Murdock, L.L. Triple bag hermetic technology for post-harvest preservation of Bambara groundnut (Vigna subterranea (L.) Verdc.). J. Stored Prod. Res. 2014, 58, 48–52. [Google Scholar] [CrossRef]
- Murdock, L.L.; Margam, V.; Baoua, I.; Balfe, S.; Shade, R.E. Death by desiccation: Effects of hermetic storage on cowpea bruchids. J. Stored Prod. Res. 2012, 49, 166–170. [Google Scholar] [CrossRef]
- Baributsa, D.; Fletcher-Timmons, H. Purdue Improved Crop Storage (PICS) Newsletter; PICS Program Partnerships, Department of Entomology, Purdue University: West Lafayette, IN, USA, 2017; p. 2. [Google Scholar]
- Hashem, M.Y.; Risha, E.S.M.; El-Sherif, S.I.; Ahmed, S.S. The effect of modified atmospheres, an alternative to methyl bromide, on the susceptibility of immature stages of angoumois grain moth Sitotroga cerealella (Olivier) (Lepidoptera: Gelechiidae). J. Stored Prod. Res. 2012, 50, 57–61. [Google Scholar] [CrossRef]
- Mbata, G.N.; Phillips, T.W. Effects of temperature and exposure time on mortality of stored-product insects exposed to low pressure. J. Econ. Entomol. 2001, 94, 1302–1307. [Google Scholar] [CrossRef] [PubMed]
- Navarro, S.; Dias, R.; Donahaye, E. Induced tolerance of Sitophilus oryzae adults to carbon dioxide. J. Stored Prod. Res. 1985, 21, 207–213. [Google Scholar] [CrossRef]
- Ofuya, T.I.; Reichmuth, C. Effect of relative humidity on the susceptibility of Callosobruchus maculatus (Fabricius) (Coleoptera: Bruchidae) to two modified atmospheres. J. Stored Prod. Res. 2002, 38, 139–146. [Google Scholar] [CrossRef]
- Yan, Y.; Williams, S.B.; Murdock, L.L.; Baributsa, D. Hermetic storage of wheat and maize flour protects against red flour beetle (Tribolium castaneum Herbst). PLoS ONE 2017, 12, e0185386. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Lei, J.; Fox, C.W.; Johnston, J.S.; Zhu-Salzman, K. Comparison of life history and genetic properties of cowpea bruchid strains and their response to hypoxia. J. Insect Physiol. 2015, 75, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Adler, C.; Corinth, H.G.; Reichmuth, C. Modified atmospheres. In Alternatives to Pesticides in Stored-Product IPM; Springer: New York, NY, USA, 2000; pp. 105–146. [Google Scholar]
- Njoroge, A.W.; Mankin, R.W.; Smith, B.W.; Baributsa, D. Effects of Hermetic Storage on Adult Sitophilus oryzae L. (Coleoptera: Curculionidae) Acoustic Activity Patterns and Mortality. J. Econ. Entomol. 2017, 110, 2707–2715. [Google Scholar] [CrossRef] [PubMed]
- Mankin, R.W.; Mizrach, A.; Hetzroni, A.; Levsky, S.; Nakache, Y.; Soroker, V. Temporal and spectral features of sounds of wood-boring beetle larvae: Identifiable patterns of activity enable improved discrimination from background noise. Fla. Entomol. 2008, 91, 241–248. [Google Scholar] [CrossRef]
- Kiobia, D.O.; Tumbo, S.D.; Cantillo, J.; Rohde, B.B.; Mallikarjunan, P.K.; Mankin, R.W. Characterization of sounds in maize produced by internally feeding insects: Investigations to develop inexpensive devices for detection of Prostephanus truncatus (Coleoptera: Bostrichidae) and Sitophilus zeamais (Coleoptera: Curculionidae) in small-scale storage facilities in sub-Saharan Africa. Fla. Entomol. 2015, 98, 405–409. [Google Scholar] [CrossRef]
- Shade, R.E.; Furgason, E.S.; Murdock, L.L. Detection of hidden insect infestations by feeding-generated ultrasonic signals. Am. Entomol. 1990, 36, 231–235. [Google Scholar] [CrossRef]
- Devereau, A.D.; Gudrups, I.; Appleby, J.H.; Credland, P.F. Automatic, rapid screening of seed resistance in cowpea, Vigna unguiculata (L.) Walpers, to the seed beetle Callosobruchus maculatus (F.) (Coleoptera: Bruchidae) using acoustic monitoring. J. Stored Prod. Res. 2003, 39, 117–129. [Google Scholar] [CrossRef]
- Baributsa, D.; Baoua, I.; Murdock, L.L. Purdue Improved Crop Storage (PICS) Bag: Size Matters; Purdue University, Department of Entomology: West Lafayette, IN, USA, 2013. [Google Scholar]
- Carter, S.; Gartner, S.; Haines, M.; Olmstead, A.; Sutch, R.; Wright, G. Historical Statistics of the United States, Millennial Edition; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Silim Nahdy, M.; Ellis, R.H.; Silim, S.N.; Smith, J. Field infestation of pigeonpea (Cajanus cajan (L.) Millsp.) by Callosobruchus chinensis (L.) in Uganda. J. Stored Prod. Res. 1998, 34, 207–216. [Google Scholar] [CrossRef]
- Bioacoustics Research Program. Raven Lite: Interactive Sound Analysis Software; Version 2.0; The Cornell Lab of Ornithology: Ithaca, NY, USA, 2016; Available online: http://www.birds.cornell.edu/raven (accessed on 13 January 2017).
- Njoroge, A.W.; Affognon, H.; Mutungi, C.; Richter, U.; Hensel, O.; Rohde, B.; Mankin, R.W. Bioacoustics of Acanthoscelides obtectus (Coleoptera: Chrysomelidae: Bruchinae) on Phaseolus vulgaris (Fabaceae). Fla. Entomol. 2017, 100, 109–115. [Google Scholar] [CrossRef]
- Njoroge, A.W.; Affognon, H.; Mutungi, C.; Rohde, B.; Richter, U.; Hensel, O.; Mankin, R.W. Frequency and time pattern differences in acoustic signals produced by Prostephanus truncatus (Horn) (Coleoptera: Bostrichidae) and Sitophilus zeamais (Motschulsky) (Coleoptera: Curculionidae) in stored maize. J. Stored Prod. Res. 2016, 69, 31–40. [Google Scholar] [CrossRef]
- Mankin, R.W.; Brandhorst-Hubbard, J.; Flanders, K.L.; Zhang, M.; Crocker, R.L.; Lapointe, S.L.; Weaver, D.K. Eavesdropping on insects hidden in soil and interior structures of plants. J. Econ. Entomol. 2000, 93, 1173–1182. [Google Scholar] [CrossRef] [PubMed]
- Herrick, N.J.; Mankin, R.W.; Dosunmu, O.G.; Kairo, M.T.K. Ecology and detection of the red palm weevil, Rhynchophorus ferrugineus (Coleoptera: Curculionidae), and related weevils for the protection of palm tree species in the United States. In Proceedings of the Colloque Méditerranéen sur les Ravageurs des Palmiers, Nice, France, 16–18 January 2013. [Google Scholar]
- Mankin, R.W.; Hagstrum, D.W.; Smith, M.T.; Roda, A.L.; Kairo, M.T.K. Perspective and promise: A century of insect acoustic detection and monitoring. Am. Entomol. 2011, 57, 30–44. [Google Scholar] [CrossRef]
- Jalinas, J.; Güerri-Agulló, B.; Mankin, R.W.; López-Follana, R.; Lopez-Llorca, L.V. Acoustic assessment of Beauveria bassiana (Hypocreales: Clavicipitaceae) effects on Rhynchophorus ferrugineus (Coleoptera: Dryophthoridae) larval activity and mortality. J. Econ. Entomol. 2015, 108, 444–453. [Google Scholar] [CrossRef] [PubMed]
- StataCorp. Stata Statistical Software: Release 12; StataCorp LP: College Station, TX, USA, 2011. [Google Scholar]
- SAS Institute Inc. Base SAS® 9.4. Procedures Guide: Statistical Procedures, 2nd ed.; SAS Institute Inc.: Cary, NC, USA, 2013. [Google Scholar]
- Jalinas, J.; Güerri-Agulló, B.; Dosunmu, O.G.; Lopez Llorca, L.V.; Mankin, R.W. Acoustic activity cycles of Rhynchophorus ferrugineus (Coleoptera: Dryophthoridae) early instars after Beauveria bassiana (Hypocreales: Clavicipitaceae) treatments. Ann. Entomol. Soc. Am. 2017, 110, 551–557. [Google Scholar] [CrossRef]
- Ofuya, T.I.; Reichmuth, C. Effect of level of seed infestation on mortality of larvae and pupae of Callosobruchus maculatus (F.) (Coleoptera: Bruchidae) in some controlled atmospheres. J. Stored Prod. Res. 1994, 30, 75–78. [Google Scholar] [CrossRef]
- Quellhorst, H.E.; Williams, S.B.; Murdock, L.L.; Baributsa, D. Cumulative oxygen consumption during development of two postharvest insect pests: Callosobruchus maculatus Fabricius and Plodia interpunctella Hübner. J. Stored Prod. Res. 2018. [Google Scholar] [CrossRef]
- Williams, S.B.; Murdock, L.L.; Kharel, K.; Baributsa, D. Grain size and grain depth restrict oxygen movement in leaky hermetic containers and contribute to protective effect. J. Stored Prod. Res. 2016, 69, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Williams, S.B.; Baributsa, D.; Murdock, L.L. Hypoxia Treatment of Callosobruchus maculatus Females and Its Effects on Reproductive Output and Development of Progeny Following Exposure. Insects 2016, 7, 26. [Google Scholar] [CrossRef] [PubMed]
- Mitcham, E.; Martin, T.; Zhou, S. The mode of action of insecticidal controlled atmospheres. Bull. Entomol. Res. 2006, 96, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.; Alexander, C.; Lowenberg-DeBoer, J. Profitability of hermetic Purdue Improved Crop Storage (PICS) Bags for African Common Bean Producers; Department of Agricultural Economics, Purdue University: West Lafayette, IN, USA, 2011; pp. 1–29. [Google Scholar]
- Baributsa, D.; Lowenberg-DeBoer, J.; Murdock, L.; Moussa, B. Profitable chemical-free cowpea storage technology for smallholder farmers in Africa: Opportunities and challenges. In Proceedings of the 10th International Working Conference on Stored Product Protection, Berlin, Germany, 27 June–2 July 2010; Volume 27, pp. 1046–1052. [Google Scholar]
- Baributsa, D.; Abdoulaye, T.; Lowenberg-DeBoer, J.; Dabiré, C.; Moussa, B.; Coulibaly, O.; Baoua, I. Market building for post-harvest technology through large-scale extension efforts. J. Stored Prod. Res. 2014, 58, 59–66. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Residual Oxygen Level (%) * | ||||
|---|---|---|---|---|---|
| Day 1 | Day 2 | Day 3 | Day 4 | Day 5 | |
| 25 insects/1000 mL | 19.91 ± 0.04 a | 19.14 ± 0.31 a | 17.08 ± 0.25 a | 15.98 ± 0.28 a | 12.35 ± 0.79 a |
| 25 insects/500 mL | 19.68 ± 0.77 a | 18.63 ± 0.97 ab | 16.02 ± 1.10 ab | 14.92 ± 2.24 ab | 11.85± 3.37 ab |
| 50 insects/1000 mL | 19.93 ± 0.08 a | 17.71 ± 0.72 b | 14.86 ± 0.39 b | 13.26 ± 0.36 b | 10.12± 0.52 b |
| 50 insects/500 mL | 19.80 ± 0.34 a | 15.12 ± 1.56 c | 10.40 ± 1.16 c | 7.76 ± 0.94 c | 5.61 ± 1.02 c |
| 100 insects/1000 mL | 20.05 ± 0.14 a | 14.63 ± 1.38 c | 9.37 ± 0.53 c | 6.87 ± 0.87 c | 4.06 ± 0.63 c |
| 100 insects/500 mL | 19.79 ± 0.66 a | 11.43 ± 1.74 d | 4.44 ± 1.56 d | 2.86 ± 1.03 d | 2.61 ± 1.30 d |
| Parameter | Df | F | p |
|---|---|---|---|
| Rates of bursts | |||
| Treatment | 4 | 15.48 | <0.001 |
| Storage time | 8 | 23.29 | <0.001 |
| Treatment × Storage time | 22 | 5.34 | <0.001 |
| Rates of impulses in bursts | |||
| Treatment | 4 | 13.36 | 0.0008 |
| Storage time | 8 | 16.97 | 0.0203 |
| Treatment × Storage time | 22 | 4.55 | 0.0033 |
| Impulses per burst | |||
| Treatment | 4 | 3.96 | <0.001 |
| Storage time | 8 | 1.73 | <0.001 |
| Treatment × Storage time | 22 | 2.06 | <0.001 |
| Treatment | Daily Callosobruchus maculatus Mean Activity (Bursts s−1) * | ||||
|---|---|---|---|---|---|
| Day 1 | Day 2 | Day 3 | Day 4 | Day 5 | |
| 25 insects/500 mL | 0.40 ± 0.12 a | 0.12 ± 0.03 a | 0.02 ± 0.01 a | 0.0 ± 0.0 a | 0.01 ± 0.01 a |
| 25 insects/1000 mL | 0.02 ± 0.01 b | 0.01 ± 0.004 b | 0.0 ± 0.0 a | 0.01± 0.001 a | 0.03 ± 0.02 a |
| 50 insects/500 mL | 1.15 ± 0.22 ac | 0.40 ± 0.07 c | 0.14 ± 0.03 b | 0.02 ± 0.004 b | 0.05 ± 0.01 a |
| 50 insects/1000 mL | 1.01 ± 0.20 c | 0.19 ± 0.06 a | 0.09 ± 0.02 ab | 0.05 ± 0.02 b | 0.01 ± 0.003 a |
| 100 insects/500 mL | 0.83 ± 0.19 ac | 0.98 ± 0.14 a | 0.22 ± 0.03 c | 0.02 ± 0.01 b | 0.01 ± 0.003 a |
| 100 insects/1000 mL | 0.65 ± 0.19 ac | 0.24 ± 0.04 a | 0.05 ± 0.02 ab | 0.02 ± 0.01 b | 0.03 ± 0.01 a |
| Measurement | Intercept ± SEM | t | p > t | Slope ± SEM | t | p > t |
|---|---|---|---|---|---|---|
| Br (Equation (1)) | 9.44 ± 1.412 | 6.69 | <0.001 | −0.585 ± 0.111 | −5.27 | <0.001 |
| Ir (Equation (2)) | 75.33 ± 12.62 | 5.97 | <0.001 | −4.598 ± 0.993 | −4.62 | <0.001 |
| Ipb (Equation (3)) | 7075.6 ± 2097 | 3.37 | 0.0014 | 215.5 ± 165.13 | 1.30 | 0.1976 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Njoroge, A.W.; Mankin, R.W.; Smith, B.W.; Baributsa, D. Oxygen Consumption and Acoustic Activity of Adult Callosobruchus maculatus (F.) (Coleoptera: Chrysomelidae: Bruchinae) during Hermetic Storage. Insects 2018, 9, 45. https://doi.org/10.3390/insects9020045
Njoroge AW, Mankin RW, Smith BW, Baributsa D. Oxygen Consumption and Acoustic Activity of Adult Callosobruchus maculatus (F.) (Coleoptera: Chrysomelidae: Bruchinae) during Hermetic Storage. Insects. 2018; 9(2):45. https://doi.org/10.3390/insects9020045
Chicago/Turabian StyleNjoroge, Anastasia W., Richard W. Mankin, Bradley W. Smith, and Dieudonne Baributsa. 2018. "Oxygen Consumption and Acoustic Activity of Adult Callosobruchus maculatus (F.) (Coleoptera: Chrysomelidae: Bruchinae) during Hermetic Storage" Insects 9, no. 2: 45. https://doi.org/10.3390/insects9020045