Effect of the Topical Repellent para-Menthane-3,8-diol on Blood Feeding Behavior and Fecundity of the Dengue Virus Vector Aedes aegypti


Abstract
:1. Introduction
2. Materials and Methods
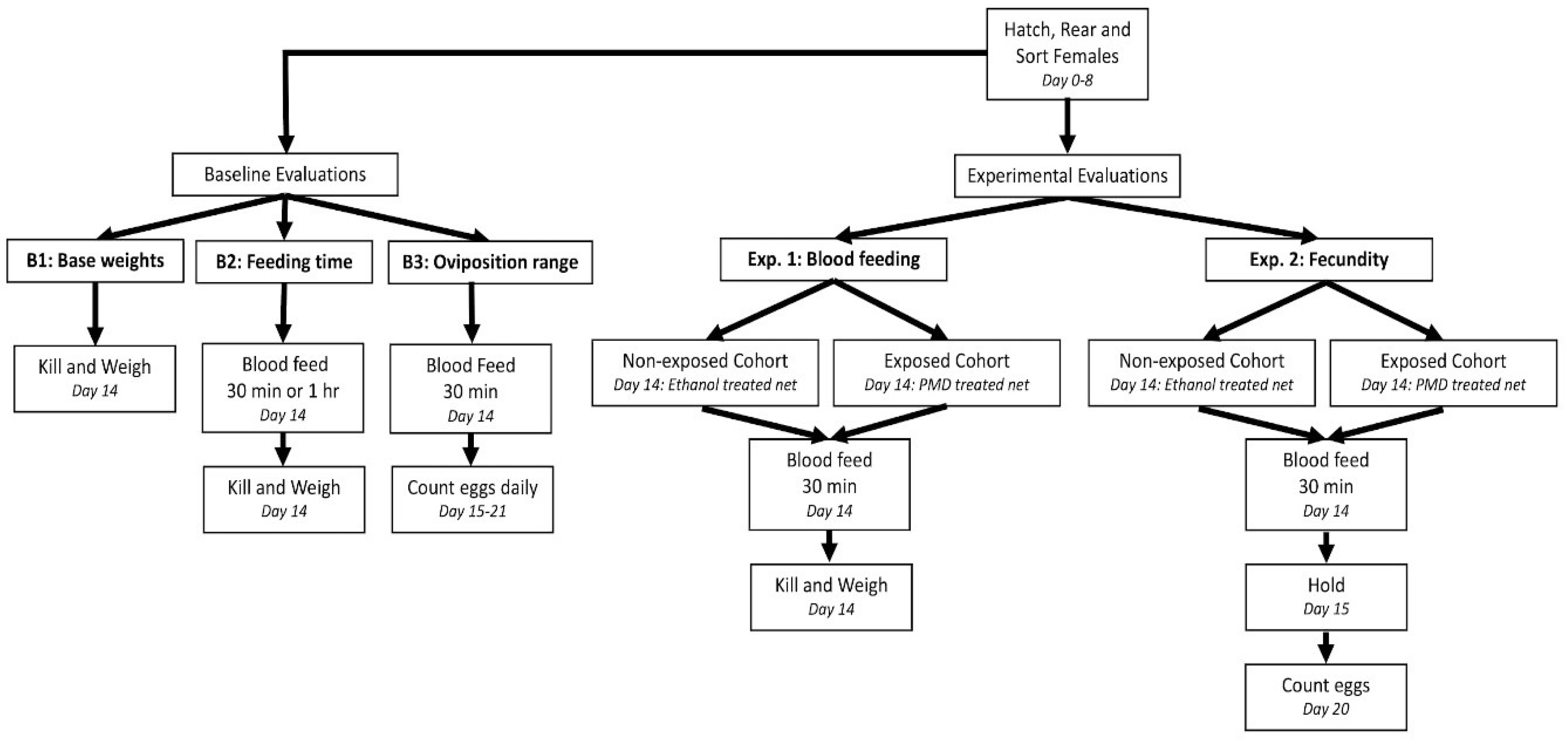
2.1. Experimental Design
2.2. Mosquito Cohorts
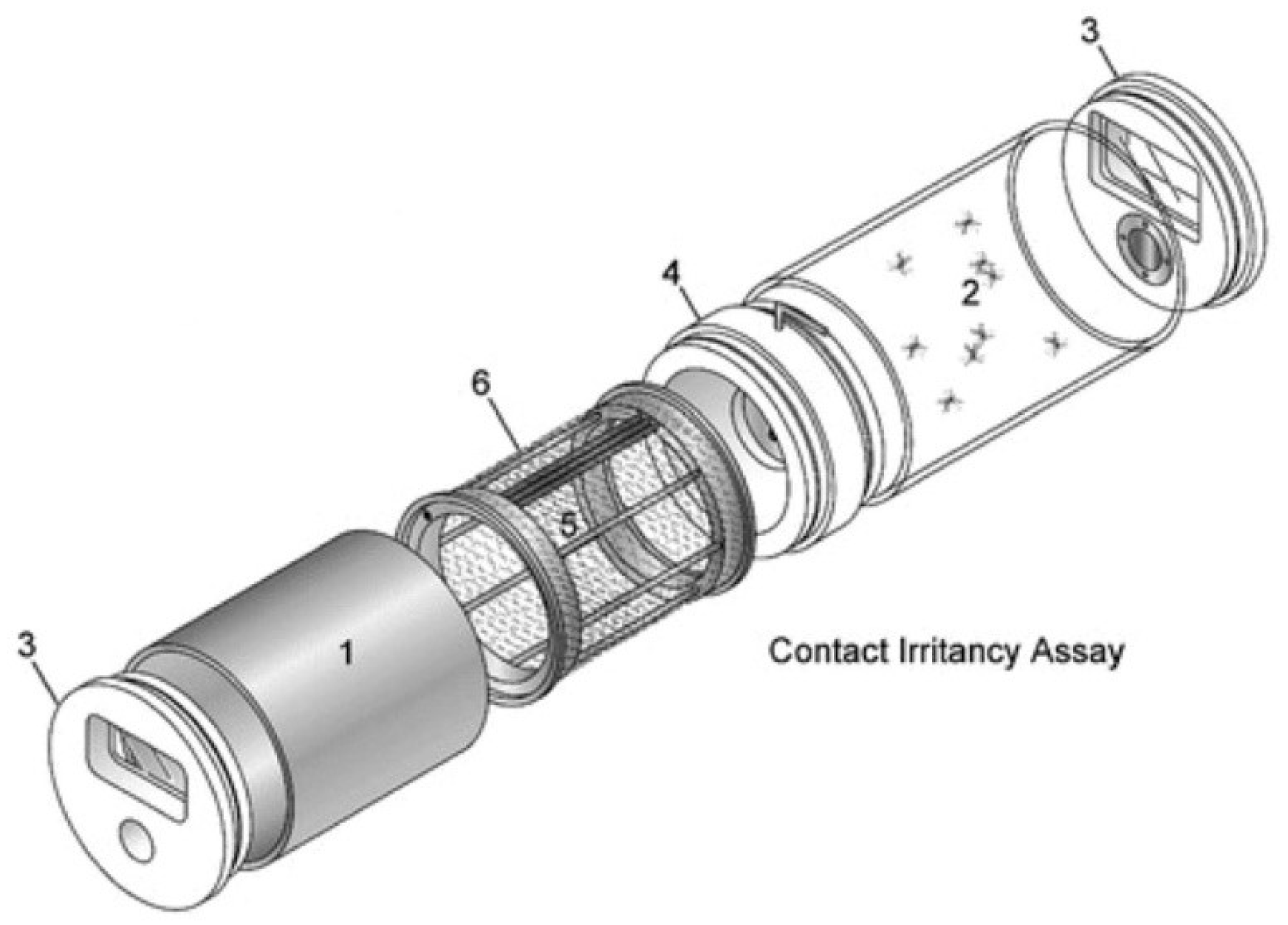
2.3. Chemical Exposure
2.4. Experiment 1—Blood-Feeding Behavior
2.5. Experiment 2—Fecundity
2.6. Data Analysis
3. Results
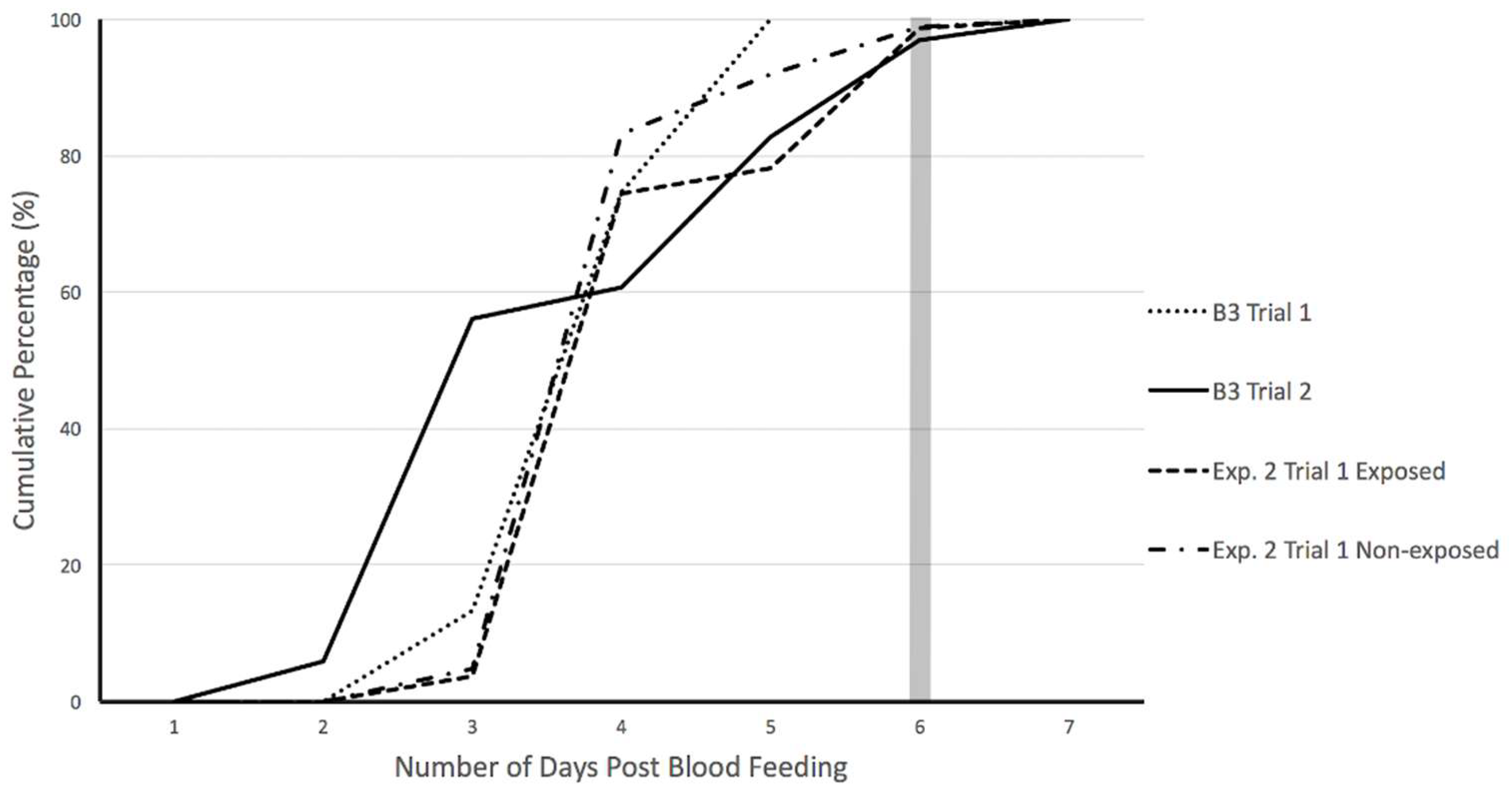
3.1. Baseline Trials
3.2. Experimental Trials
3.2.1. Experiment 1—Blood-Feeding Behavior
3.2.2. Experiment 2—Fecundity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- WHO. Dengue and Severe Dengue. Available online: http://www.who.int/mediacentre/factsheets/fs117/en/ (accessed on 28 March 2017).
- World Health Organization, Department of Control of Neglected Tropical Diseases. Guidelines for Laboratory and Field-Testing of Long-Lasting Insecticidal Nets; WHO Press: Geneva, Switzerland, 2013; ISBN 978-92-4-150527-7. [Google Scholar]
- Benelli, G.; Romano, D. Mosquito vectors of Zika virus. Entomologia Generalis 2017, 36, 309–318. [Google Scholar] [CrossRef]
- Rassmussen, S.A.; Jamieson, D.J.; Honein, M.A.; Petersen, L.R. Zika virus and birth defects—Reviewing the evidence for causality. N. Engl. J. Med. 2016, 374, 1981–1987. [Google Scholar] [CrossRef] [PubMed]
- WHO. Questions and Answers on Dengue Vaccines. Available online: http://www.who.int/immunization/research/development/dengue_q_and_a/en/ (accessed on 28 March 2017).
- Kwek, S.S.; Watanabe, S.; Chan, K.R.; Ong, E.Z.; Tan, H.C.; Ng, W.C.; Nguyen, M.T.X.; Gan, E.S.; Zhang, S.L.; Chan, K.W.K.; et al. A systematic approach to the development of a safe live attenuated Zika vaccine. Nat. Commun. 2018, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Hemingway, J. The role of vector control in stopping the transmission of malaria: Threats and opportunities. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Global Strategy for Dengue Prevention and Control 2012–2020; WHO Press: Geneva, Switzerland, 2012; ISBN 9789241504034. [Google Scholar]
- Benelli, G.; Mehlhorn, H. Declining malaria, rising of dengue and Zika virus: Insights for mosquito vector control. Parasitol. Res. 2016, 115, 1747–1754. [Google Scholar] [CrossRef] [PubMed]
- Ditzen, M.; Pellegrino, M.; Vosshall, L.B. Insect odorant receptors are molecular targets of the insect repellent DEET. Science 2008, 319, 1838–1842. [Google Scholar] [CrossRef] [PubMed]
- Pellegrinino, M.; Steinbach, N.; Stensmyr, M.C.; Hansson, B.S.; Vosshall, L.B. A natural polymorphism alters odour and DEET sensitivity in an insect odorant receptor. Nature 2011, 478, 511–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanczyk, N.M.; Brookfield, J.F.Y.; Ignell, R.; Logan, J.G.; Field, L.M. Behavioral insensitivity to DEET in Aedes aegypti is a genetically determined trait residing in changes in sensillum function. Proc. Natl. Acad. Sci. USA 2010, 107, 8575–8580. [Google Scholar] [CrossRef] [PubMed]
- Stanczyk, N.M.; Brookfield, J.F.Y.; Field, L.M.; Logan, J.G. Aedes aegypti Mosquitoes Exhibit Decreased Repellency by DEET following Previous Exposure. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.J.; Hill, N.; Ruiz, C.; Cameron, M.M. Field evaluation of traditionally used plant-based Insect repellents and fumigants against the malaria vector Anopheles darlingi in Riberalta, Bolivian Amazon. J. Med. Entomol. 2007, 44, 624–630. [Google Scholar] [CrossRef] [PubMed]
- Maia, M.F.; Moore, S.J. Plant-based insect repellents: A review of their efficacy, development and testing. Malar. J. 2011, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carroll, S.P.; Loye, J. PMD, a Registered Botanical Mosquito Repellent with Deet-Like Efficacy. J. Am. Mosq. Control Assoc. 2006, 22, 507–514. [Google Scholar] [CrossRef]
- Uzzan, B.; Konate, L.; Diop, A.; Nicolas, P.; Dia, I.; Dieng, Y.; Izri, A. Efficacy of four insect repellents against mosquito bites: A double-blind randomized placebo-controlled field study in Senegal. Fundam. Clin. Pharmacol. 2009, 23, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Barnard, D.R.; Xue, R. Laboratory evaluation of mosquito repellents against Aedes albopictus, Culex nigripalpus, and Ochlerotatus triseriatus (Diptera: Culicidae). J. Med. Entomol. 2004, 41, 726–730. [Google Scholar] [CrossRef] [PubMed]
- Solomon, B.; Gebre-Mariam, T.; Asres, K. Mosquito repellent actions of the essential oils of Cymbopogon citratus, Cymbopogon nardus and Eucalyptus citriodora: Evaluation and formulation studies. J. Essent. Bear. Plants 2012, 15, 766–773. [Google Scholar] [CrossRef]
- Peterson, C.; Coats, J. Insect repellents—Past, present and future. Pestic. Outlook 2001, 12, 154–158. [Google Scholar] [CrossRef]
- Hao, H.; Wei, J.; Dai, J.; Du, J. Host-seeking and blood-feeding behavior of Aedes albopictus (Diptera: Culicidae) exposed to vapors of geraniol, citral, citronellal, eugenol, or anisaldehyde. J. Med. Entomol. 2008, 45, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.B.; Grieco, J.P.; Apperson, C.S.; Schal, C.; Ponnusamy, L.; Wesson, D.M.; Achee, N.L. Effect of spatial repellent exposure on dengue vector attraction to oviposition sites. PLoS Negl Trop Dis. 2016, 10, e0004850. [Google Scholar] [CrossRef] [PubMed]
- Sugiharto, V.A.; Grieco, J.P.; Murphy, J.R.; Olsen, C.H.; Colacicco-Mayhugh, M.G.; Stewart, V.A.; Achee, N.L.; Turell, M.J. Effects of preexposure to DEET on the downstream blood-feeding behaviors of Aedes aegypti (Diptera: Culicidae) mosquitoes. J. Med. Entomol. 2016, 53, 1100–1104. [Google Scholar] [CrossRef] [PubMed]
- Achee, N.L.; Foggie, T. Standard Operating Procedures: Rearing Aedes aegypti for the HITSS and Box Laboratory Assays. Available online: https://www.usuhs.edu/sites/default/files/media/pmb/pdf/insectarysop.pdf (accessed on 21 September 2017).
- Shiao, S.; Hansen, I.A.; Zhu, J.; Sieglaff, D.H.; Raikhel, A.S. Juvenile hormone connects larval nutrition with target of rapamycin signaling in the mosquito Aedes aegypti. J. Insect Physiol. 2008, 54, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Achee, N.L. Behavioral Modifying Compounds for Disease Vector Control. Available online: https://www.usuhs.mil/sites/default/files/media/pmb/pdf/htsstrainingmanual.pdf (accessed on 21 September 2017).
- Grieco, J.P.; Achee, N.L.; Sardelis, M.R.; Chauhan, K.R.; Roberts, D.R. A novel high-throughput screening system to evaluate the behavioral response of adult mosquitoes to chemicals. J. Am. Mosq. Control Assoc. 2005, 21, 404–411. [Google Scholar] [CrossRef]
- Putnam, J.L.; Scott, T.W. Blood-Feeding Behavior of Dengue-2 Virus-Infected Aedes Aegypti. Am. J. Trop. Med. Hyg. 1995, 52, 225–227. [Google Scholar] [CrossRef] [PubMed]
- Scott, T.W.; Chow, E.; Strickman, D.; Kittayapong, P.; Wirtz, R.A.; Lorenz, L.H.; Edman, J.D. Blood-feeding patterns of Aedes aegypti (Diptera: Culicidae) collected in a rural Thai village. J. Med. Entomol. 1993, 30, 922–927. [Google Scholar] [CrossRef] [PubMed]
- Esteva, L.; Vargas, C. Analysis of a dengue disease transmission model. Math. Biosci. 1998, 150, 131–151. [Google Scholar] [CrossRef]
- Lambrechts, L.; Paaijmans, K.P.; Fansiri, T.; Carrington, L.B.; Kramer, L.D.; Thomas, M.B.; Scott, T.W.; Beaty, B.J. Impact of daily temperature fluctuations on dengue virus transmission by Aedes aegypti. Proc. Natl. Acad. Sci. USA 2011, 108, 7460–7465. [Google Scholar] [CrossRef] [PubMed]
- Zwiebel, L.J.; Takken, W. Olfactory regulation of mosquito–host interactions. Insect Biochem. Mol. Biol. 2004, 34, 645–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, B.T.; Brewster, C.C.; Paulson, S.L. La Crosse virus infection alters blood feeding behavior in Aedes triseriatus and Aedes albopictus (Diptera: Culicidae). J. Med. Entomol. 2012, 49, 1424–1429. [Google Scholar] [CrossRef] [PubMed]
- Klowden, M.J.; Lea, A.O. Abdominal distention terminates subsequent host-seeking behaviour of Aedes aegypti following a blood meal. J. Insect Physiol. 1979, 25, 583–585. [Google Scholar] [CrossRef]
- Nentwig, G.; Frohberger, S.; Sonneck, R. Evaluation of clove oil, icaridin, and transfluthrin for spatial repellent effects in Three Tests Systems Against the Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 2017, 54, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Pilitt, D.R.; Jones, J.C. A qualitative method for estimating the degree of engorgement of Aedes aegypti adults. J. Med. Entomol. 1972, 9, 334–337. [Google Scholar] [CrossRef] [PubMed]
- Milali, M.P.; Sikulu-Lord, M.T.; Govella, N.J. Bites before and after bedtime can carry a high risk of human malaria infection. Malar. J. 2017, 16, 91. [Google Scholar] [CrossRef] [PubMed]
- Xue, R.; Barnard, D.R.; Ali, A. Influence of multiple blood meals on gonotrophic dissociation and fecundity in Aedes albopictus. J. Am. Mosq. Control Assoc. 2009, 25, 504–507. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization, Department of Control of Neglected Tropical Diseases. Handbook for Integrated Vector Management; WHO Press: Geneva, Switzerland, 2013; ISBN 978-92-4-150280-1. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exposed | Non-Exposed | p-Value | |
|---|---|---|---|
| % blood-fed 2 ± SEM | 38.1 (±1.84) n = 700 | 49.1 (±1.89) n = 700 | <0.001 |
| Weight blood-fed 2 . (mg) ± SD | 5.24 (±0.91) n = 267 | 5.31 (±0.96) n = 344 | 0.2394 |
| Weight non-blood-fed 2 (mg) ± SD | 2.47 (±0.47) n = 433 | 2.44 (±0.47) n = 356 | |
| % oviposited 3 ± SEM | 55.5 (±3.70) n = 180 | 58.9 (±3.67) n = 180 | 0.594 |
| No. eggs 3 ± SD | 62.0 (±4.00) n = 100 | 54.0 (±4.00) n = 106 | 0.141 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.; Choi, D.B.; Liu, F.; Grieco, J.P.; Achee, N.L. Effect of the Topical Repellent para-Menthane-3,8-diol on Blood Feeding Behavior and Fecundity of the Dengue Virus Vector Aedes aegypti. Insects 2018, 9, 60. https://doi.org/10.3390/insects9020060
Lee J, Choi DB, Liu F, Grieco JP, Achee NL. Effect of the Topical Repellent para-Menthane-3,8-diol on Blood Feeding Behavior and Fecundity of the Dengue Virus Vector Aedes aegypti. Insects. 2018; 9(2):60. https://doi.org/10.3390/insects9020060
Chicago/Turabian StyleLee, Jugyeong, Diane B. Choi, Fang Liu, John P. Grieco, and Nicole L. Achee. 2018. "Effect of the Topical Repellent para-Menthane-3,8-diol on Blood Feeding Behavior and Fecundity of the Dengue Virus Vector Aedes aegypti" Insects 9, no. 2: 60. https://doi.org/10.3390/insects9020060