
Epigenetic Regulation of a Heat-Activated Retrotransposon in Cruciferous Vegetables
by
 ,
,
Kosuke Nozawa
1, 
Yuki Kawagishi
1,
Akira Kawabe
2,
Mio Sato
1,
Yukari Masuta
3,
Atsushi Kato
3 and
Hidetaka Ito
3,* 1
Graduate School of Life Science, Hokkaido University, Kita10 Nishi8, Kita-ku, Sapporo, Hokkaido 060-0810, Japan
2
Faculty of Life Sciences, Kyoto Sangyo University, Kamigamo Motoyama Kita-ku, Kyoto 603-8555, Japan
3
Faculty of Science, Hokkaido University, Kita10 Nishi8, Kita-ku, Sapporo, Hokkaido 060-0810, Japan
*
Author to whom correspondence should be addressed.
Epigenomes 2017, 1(1), 7; https://doi.org/10.3390/epigenomes1010007
Submission received: 10 May 2017
/
Revised: 26 May 2017
/
Accepted: 31 May 2017
/
Published: 7 June 2017
(This article belongs to the Special Issue Plant Epigenome Dynamics)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Transposable elements (TEs) are highly abundant in plant genomes. Environmental stress is one of the critical stimuli that activate TEs. We analyzed a heat-activated retrotransposon, named ONSEN, in cruciferous vegetables. Multiple copies of ONSEN-like elements (OLEs) were found in all of the cruciferous vegetables that were analyzed. The copy number of OLE was high in Brassica oleracea, which includes cabbage, cauliflower, broccoli, Brussels sprout, and kale. Phylogenic analysis demonstrated that some OLEs transposed after the allopolyploidization of parental Brassica species. Furthermore, we found that the high copy number of OLEs in B. oleracea appeared to induce transpositional silencing through epigenetic regulation, including DNA methylation. The results of this study would be relevant to the understanding of evolutionary adaptations to thermal environmental stress in different species.
1. Introduction
Retrotransposons are major components of most plant genomes and are among the main sources of genetic diversity. They act as perpetual agents of mutagenicity because of their amplification and mobility [1]. Retrotransposons are also known to be involved in the regulation of gene expression as well as in cellular response to stress [2]. The differences in the copy numbers of transposons contribute to the differences in the genome sizes among species [3].
Retrotransposon can transpose and amplify its copy number through an RNA intermediate that is reverse transcribed into an extrachromosomal DNA and is integrated into the nuclear genome. Based on their structure, retrotransposons are divided in two groups: members of one group contain long terminal repeats (LTRs) on both their ends, and the second group involves non-LTR retrotransposons or long interspersed nuclear elements (LINEs). Retrotransposons contain two open-reading frames that encode gag and pol genes, respectively. The pol gene includes protease, reverse transcriptase (RT), RNase H, and integrase domains that are necessary for retrotransposition. Retrotransposon families can be typically recognized based on the similarities in their sequences and by the structure of the gene coding regions that allows phylogenetic and evolutionary analyses of the different families of retrotransposons.
Most of the transposons are transcriptionally controlled by epigenetic regulations of the host genome through DNA methylation and histone modifications [4,5,6]. However, despite the tight regulation, eukaryotes carry a large number of transposons [7]. We previously reported a heat-activated retrotransposon, named ONSEN, in Arabidopsis thaliana [8]. ONSEN is a Ty1/Copia like retrotransposon with its LTRs containing a cluster of four nGAAn motifs that form the heat-responsive element (HRE) [9]. When exposed to heat stress, a heat shock factor A2 binds to the HRE and triggers its transcriptional activity. We also found that the heat-activated ONSEN was transposed in a mutant that was deficient in small RNA biosynthesis as well as in plants regenerated from callus [8,10]. Interestingly, in A. thaliana, ONSEN preferentially inserts within the genes [8]. The transcriptional activation of ONSEN and its movement could affect the heat responsiveness of the flanking genes. We previously demonstrated that a gene close to a new ONSEN integration site became responsive to heat stress, suggesting that ONSEN insertion could introduce a new gene network. The changes in the expression patterns of genes caused by ONSEN activation could contribute to the specific environmental adaptation of the host plants. For instance, the transposition of ONSEN generated a mutation in an abscisic acid (ABA) responsive gene, resulting in an ABA-insensitive phenotype in A. thaliana [11].
Comparisons of the ONSEN transposon family across species are important for understanding the evolutionary adaptations of retrotransposons and the host plants to environmental stresses. Brassicaceae is among the largest plant families containing 338 genera and about 3700 species. It includes several important vegetable crops, oil seed plants, and crops that provide condiments and fodder [12]. The present study describes the analysis of the organization and heat-induced activation of the ONSEN family in several Brassicaceae species.
2. Results
2.1. Copy Number of ONSEN-Like Elements
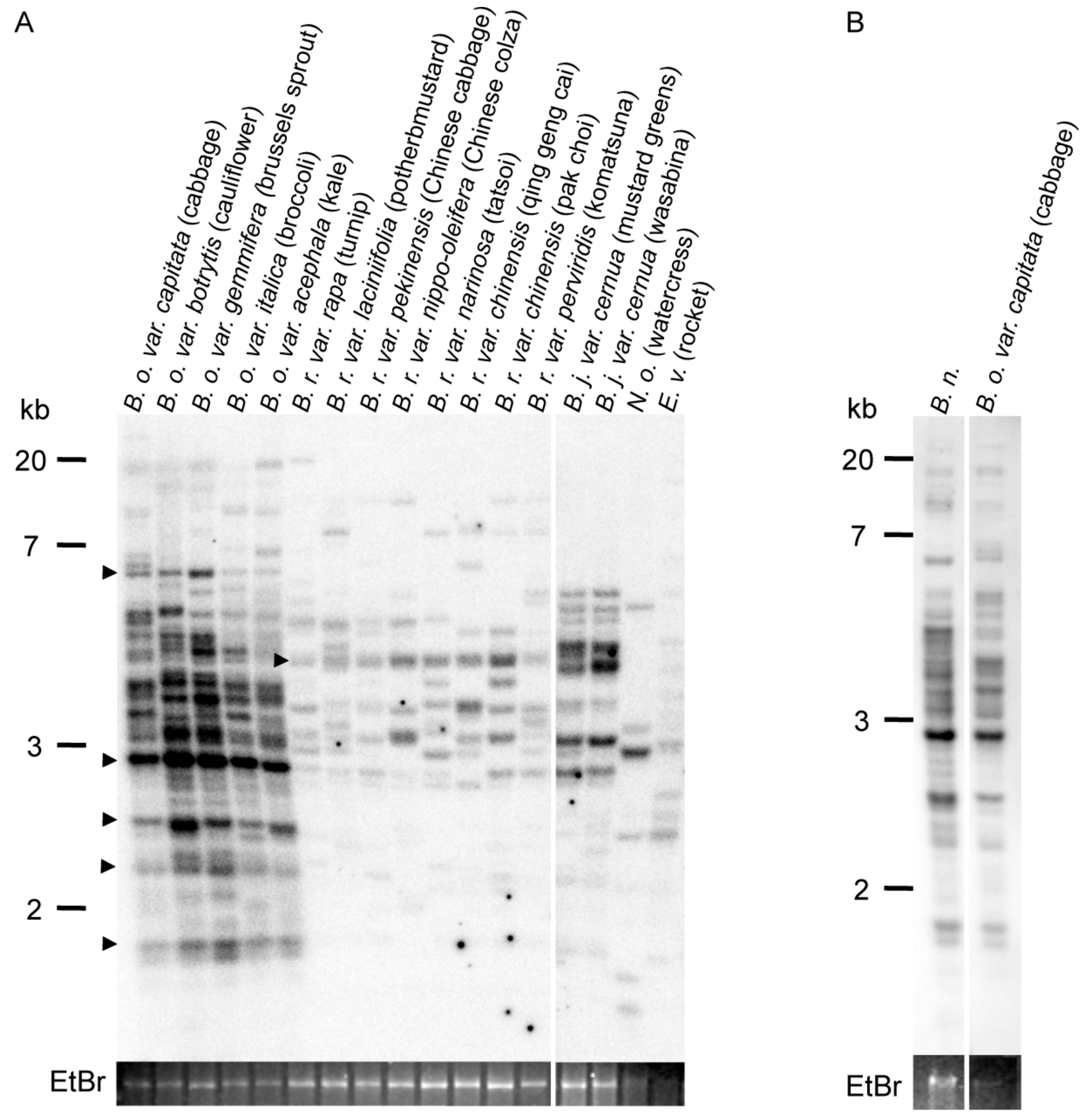
To examine the copy number of ONSEN-like elements (OLEs) in cruciferous vegetables, we analyzed 17 commercial croppings of Brassicaceae species. The Southern blot analysis showed that OLEs were present in all the analyzed cruciferous vegetables, whereas their copy numbers varied among species (Figure 1A). The copy number of OLE was abundant in Brassica oleracea, which includes cabbage, cauliflower, broccoli, Brussels sprout, and kale (Figure 1A). A common band was detected among the same species of B. oleracea, B. rapa, and B. juncea, respectively (Figure 1A). To analyze the phylogenic relationships within the Brassica species, we also analyzed the copy number of OLE in B. napus. The results showed that the copy number of OLE in B. napus was highest among the analyzed species (Figure 1B); this is consistent with the information from the whole genome sequence data of Brassica species (see Materials and Methods). There were 4–6 copies of OLEs in B. rapa, 8 copies in B. nigra, 10 copies in B. juncea, 29–62 copies in B. oleracea, and 129 copies in B. napus (34 from A genome chromosome and 78 from C genome chromosome), based on the presence of RT region (which occupied at least 50% of the core domain region).
2.2. Heat-Activation of OLE in Cruciferous Vegetables
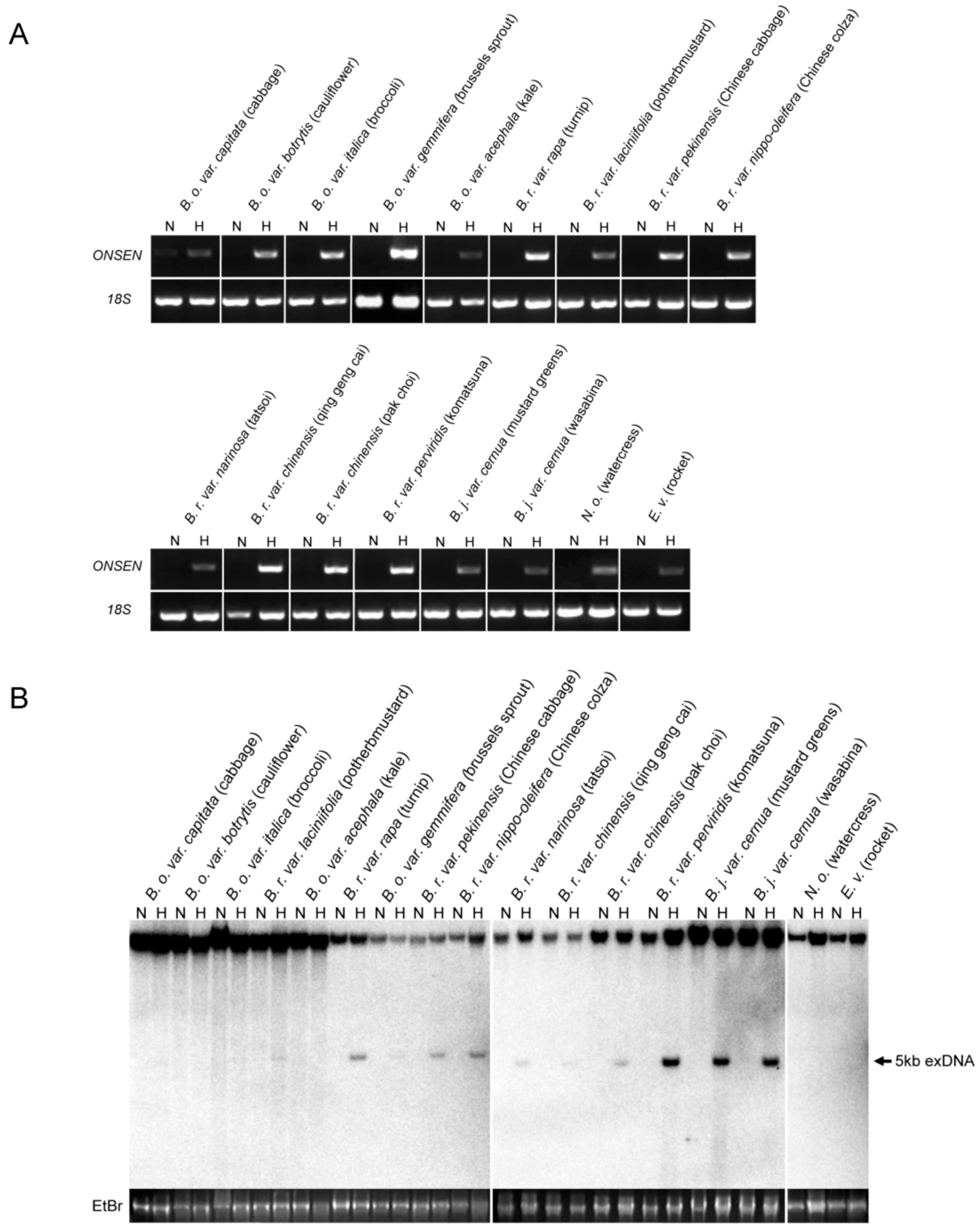
To understand the heat-responsiveness of ONSEN in cruciferous vegetables, we searched the structure of HREs within the ONSEN LTRs. None of the analyzed cruciferous vegetables contained the A. thaliana-like HREs; however, some HRE-like motifs were conserved within their LTRs (Figure S1). To examine the heat-activation of OLEs in 17 species of cruciferous vegetables, we analyzed the transcripts by Reverse Transcription Polymerase Chain Reaction (RT-PCR). The OLE transcript was detected in all the analyzed species subjected to heat stress (Figure 2A). For testing the transpositional activity of OLEs in cruciferous vegetables, we analyzed the extrachromosomal DNA that was synthesized by reverse transcription when the mRNA of an OLE was transcribed upon heat activation. The Southern blot analysis showed that the full length extrachromosomal DNA was detected in some species (Figure 2B). The result suggested that in some species OLEs were intact and might be transposable by heat stress.
2.3. Phylogenic Analysis of OLEs in Cruciferous Vegetables
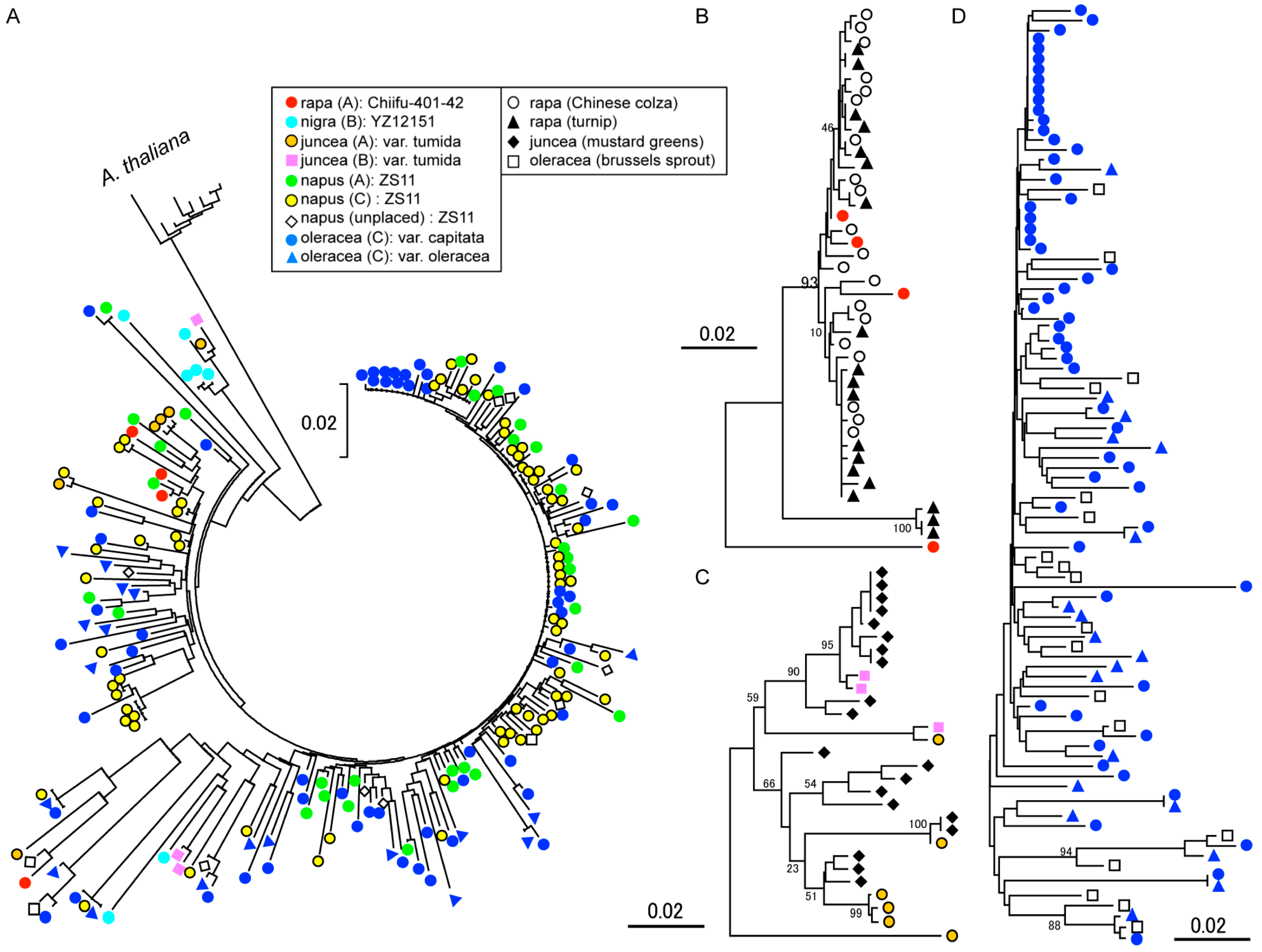
The sequences of the RT core domain region were retrieved from the whole genome sequence data of Brassica species. The phylogenetic relationship of the RT region showed several rapid amplifications of the copies, especially in B. oleracea and B. napus (Figure 3A). For B. napus, copies from A genome sometimes clustered with B. oleracea sequences, suggesting transposition to A genome after allopolyploidization. Although there were several genome specific clusters in the tree, species or genome specificities were weak, possibly because of high conservation of the RT regions. To analyze the evolutionary history of OLEs in cruciferous vegetables, we also cloned and sequenced an RT gene of OLE from four varieties of Brassica species, including B. rapa var. nippo-oleifera (rape blossoms), B. rapa var. rapa (turnip), B. juncea var. cernua (mustard greens), and B. oleracea var. gemmifera (Brussels sprout). The sequences from the different varieties (or individuals) were present in a different cluster different from those of the copies from genomic sequences (Figure 3B–D). Although identical sequences could be obtained from the same copy and some copies could not be sequenced because of PCR based cloning, amplification and degradation occurred in each variety and individual after speciation.
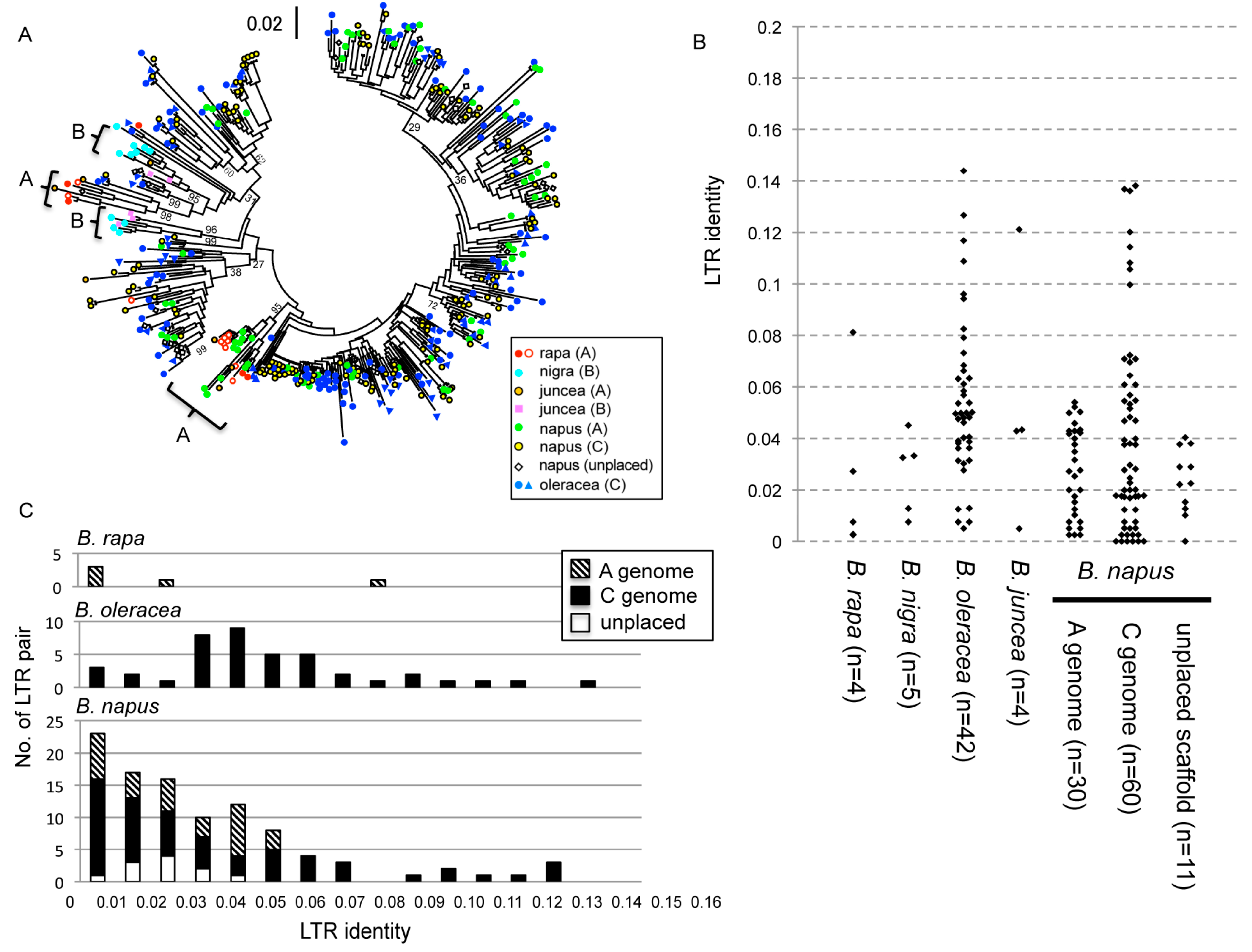
To understand the detailed evolutionary relationship, the LTR region (less conserved compared to the RT region) was used for phylogenetic analyses. The phylogenetic tree showed several clear clusters of the originated genome (Figure 4A). Brassica juncea, which originated by allopolyploidization of A and B genome species, almost had complete relation of the phylogenetic and genomic position, except that one copy from the A genome chromosome clustered with the B genome copies from B. nigra and B. juncea. This result suggests relatively weak transposability of A and B genome copies. Brassica napus has large number of copies that could have originated from either A or C genomes. The distribution of the phylogenetic positions indicated that the B. napus copies were clustered with B. oleracea copies but not with B. rapa copies. The A genome of B. napus even had copies similar to that of the B. oleracea C genome, suggesting the amplification of C genome copies after polyploidization to transpose into A genome chromosomes. The identities of the LTR pair, which could represent the age of insertion, showed different patterns among the species (Figure 4B). Although the copy number in B. oleracea was 5-times higher than that in B. rapa and B. nigra, the insertion age of B. oleracea copies varied and very old copies were still present in the genome. As predicted from the phylogenetic relationship, the B. napus A genome included relatively younger copies where all the LTR pairs had identities less than 0.06. In contrast, B. napus C genome included old copies possibly inherited from the C genome donor species. The age distribution clearly showed recent amplification in B. napus where the A genome chromosomes had relatively young copies and very young copies were amplified even in the C genome chromosomes (Figure 4C).
2.4. Epigenetic Regulation of OLE in Cruciferous Vegetables
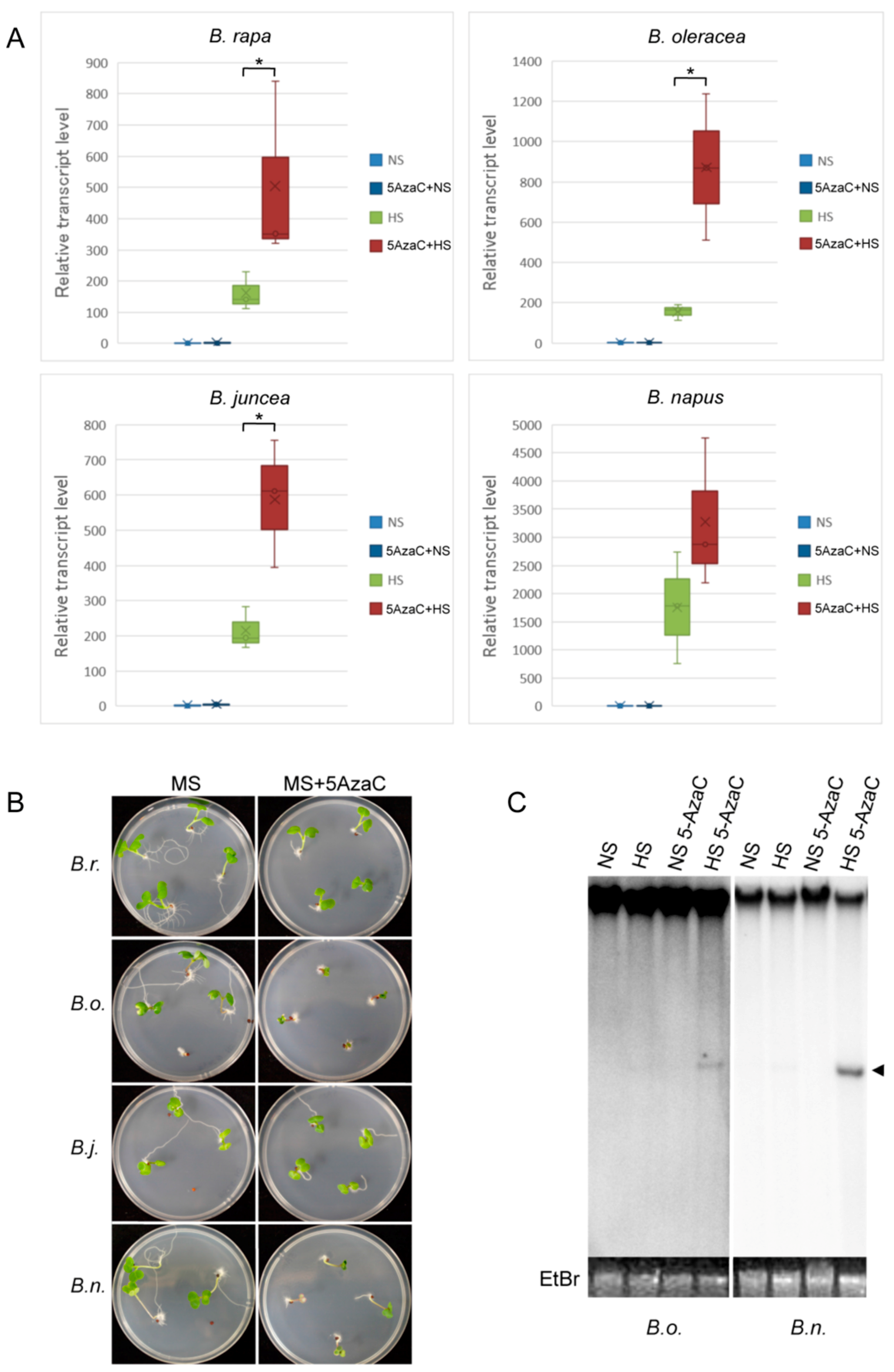
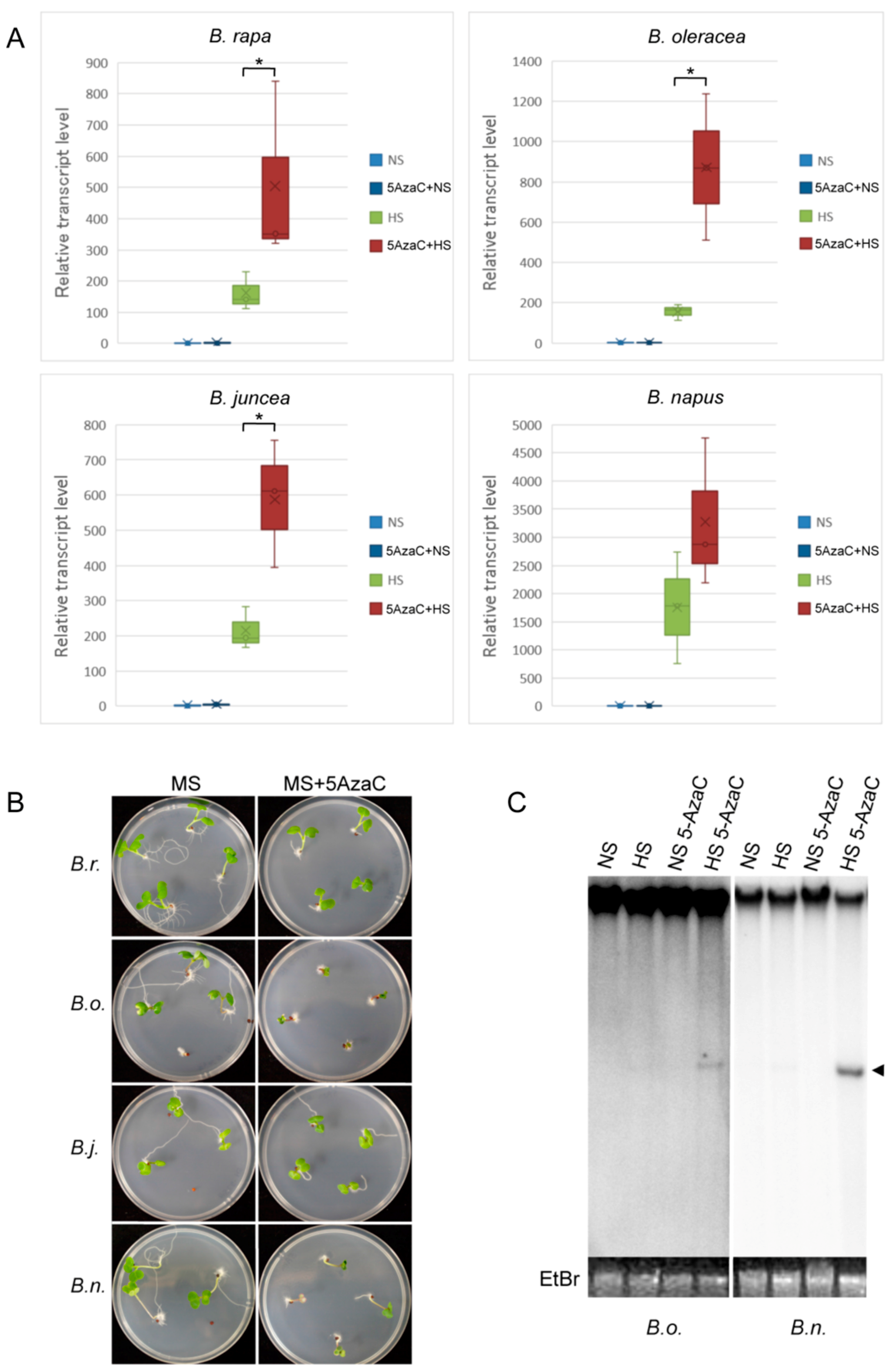
To analyze the epigenetic regulation of OLE, we analyzed the heat-activation of OLE after treatment of the plants with a DNA methylation inhibitor, 5-aza-2′-deoxycytidine (5AzaC). The results showed that the transcript level was upregulated in all the analyzed cruciferous vegetables (Figure 5A). This indicated that the expression of OLE was regulated by DNA methylation. Interestingly, 5AzaC-treated B. oleracea and B. napus showed growth inhibition in young seedlings (Figure 5B). In this study, we found that extrachromosomal DNA was not detected in most of the B. oleracea species (Figure 2B). To analyze the participation of DNA methylation in transpositional regulation of OLE, we analyzed the accumulation of extrachromosomal DNA in cruciferous vegetables treated with 5AzaC under heat stress. The results showed that the extrachromosomal DNA was detected in the heat-stressed plants treated with 5AzaC (Figure 5C).
3. Discussion
Transposable elements are highly conserved among the plant species. Some TEs have a function to regulate the stress-responsive genes in plants [13,14,15,16,17]. In this study, we focused on a heat-activated retrotransposon in cruciferous vegetables. All of the cruciferous vegetables analyzed in this study were observed to have a conserved element that was homologous to a heat-activated retrotransposon in Arabidopsis, named ONSEN. We previously reported the presence of ONSEN related copies in the cross-related species of Brassicaceae, forming a cluster with other species in the phylogenetic tree [18]. Pietzenuk et al. analyzed the common and conserved trait of HREs of ONSEN in Brassicaceae [19]. They showed that HREs in ONSEN was conserved over millions of year and evolved from a proto-HRE that was present in the evolution of Brassicaceae although most of them are species-specific. They mentioned that gain of HREs and the heat-activation does not always provide a selective advantage for TEs however the heat activation may increase the probability of survival during the co-evolution of hosts and TEs. This study demonstrated that OLEs were conserved among the more distant species. Two types of HREs were conserved in OLEs although some of them are lower efficiency suggesting that some HREs are not sufficient to trigger heat-induced activation of OLEs.
The level of transcript and the synthesized extrachromosomal DNA of OLEs did not always correlate. This indicated that some OLEs have lost their mobility due to the non-functional transcript of reverse transcriptase that is necessary to synthesize the extrachromosomal DNA. The copy number of OLE in B. oleracea was high, although the transcriptional level was lower than that in the other cruciferous vegetables. It is possible that the increased OLE copies were suppressed by an epigenetic mechanism in B. oleracea.
In Arabidopsis, the increased copy number of an LTR retrotransposon was induced by transcriptional gene silencing through RNA-directed DNA methylation (RdDM) [20]. In Brassica species, the copy number of OLE in B. oleracea was much higher than that in B. rapa and B. juncea. The accumulation of OLE in B. oleracea may induce epigenetic regulation in B. oleracea. The growth inhibition observed in B. oleracea and B. napus might be caused by an ectopic activation of TEs although there is a possibility that those species might be more sensitive to the toxicity of 5AzaC that negatively affect cell cycle progression. It is worthy of analysis to check the 5AzaC-responsive transcripts and the closeness of the transposon to the gene to see whether these TEs could stimulate hypomethylation-responsive gene expression.
The extrachromosomal DNA is synthesized as an intermediate of retrotransposition and is necessary for transpositional activation of LTR retrotransposon. In the present study, the up regulation of the transcript level and the accumulation of extrachromosomal DNA of OLEs subjected to heat suggested that OLEs could transpose in cruciferous vegetables upon exposure to heat stress. Furthermore, the extrachromosomal DNA was accumulated in B. oleracea and B. napus treated with 5AzaC under heat stress. The result indicated that the retrotransposition of some OLEs in B. oleracea and B. napus might be regulated post-transcriptionally by epigenetic regulation that involved DNA methylation.
Genomic changes could occur during the hybridization of two species that might cause genome-shock stress [21]. The interspecific hybridization has been shown to induce the activation of some transposons that provides an opportunity to investigate the evolution of transposons and the consequences of transposition [22,23]. Many Brassica species are polyploid and have evolved by genome duplications. In the present study, the copy number of OLEs was observed to vary among the species and was found to be abundant in B. oleracea and B. napus, indicating that OLEs might contribute to gene diversity in the highly duplicated Brassica genome. It is worth analyzing the regulatory factors that affect the copy number of OLEs during the evolutional history of Brassica species.
4. Materials and Methods
4.1. Plant Material and Stress Treatments
The seeds of cruciferous vegetables, including B. oleracea var. capitata (cabbage), B. oleracea var. botrytis (cauliflower), B. oleracea var. gemmifera (Brussels sprout), B. oleracea var. italica (broccoli), B. oleracea var. acephala (kale), B. rapa var. rapa (turnip), B. rapa var. lancifolia (potherb mustard), B. rapa var. pekinensis (Chinese cabbage), B. rapa var. nippo-oleifera (rape blossoms), B. rapa var. narinosa (tatsoi), B. rapa var. chinensis (qing geng cai), B. rapa var. perviridis (komatsuna), B. juncea var. cernua (mustard greens), and B. juncea var. cernua (wasabina), Nasturtium officinale (watercress), and Eruca vesicaria (rocket) were procured from SAKATA Seed Corporation (Yokohama, Japan). Brassica napus (rapeseed) was obtained from Takii & Company, Limited (Kyoto, Japan). The plants were grown on Murashige and Skoog (MS) medium with continuous light at 21 °C. The heat stress treatment was conducted using 2-week-old seedlings that were subjected to a temperature shift of 24 h at 37 °C. After the heat treatment, the plants were transplanted to soil for further growth at 21 °C under continuous light conditions. For 5AzaC treatment, the plants were grown on MS supplemented with 100 µM 5AzaC (Wako, Osaka, Japan).
4.2. Southern Blot Analysis
The genomic DNA was isolated using Nucleon PhytoPure DNA extraction kit (GE Healthcare Life Science, Chicago, IL, USA). The Southern blotting was performed as described previously [24]. The hybridization signals were detected using a radiolabeled ONSEN-specific probe (Supplementary Table S1) that was generated using Megaprime DNA Labeling System (GE Healthcare Life Science) in a hybridization buffer [25].
4.3. RT-PCR
Total RNA was extracted from whole seedlings using TRI Reagent (Sigma Aldrich, St. Louis, MO, USA), according to the manufacturer’s instructions. For RT-PCR and real-time RT-PCR, approximately 3–5 µg of the total RNA was treated with RQ1 RNase-free DNase (Promega, Madison, WI, USA) and reverse-transcribed using the ReverTraAce qPCR RT Kit (Toyobo, Osaka, Japan) with a random primer. Polymerase chain reaction was performed using TaKaRA Ex Taq (TaKaRA, Shiga, Japan) and primers kwgs_ATRS_RVT2-F (5′-TGGGAGTTAACTTCACTTCCA-3′) and kwgs_ATRS_RVT2-R (5′-CGCATTCCATTGGTGTACAA-3′); the reaction conditions were as follows: 5 min at 94 °C; 30 cycles of 94 °C (30 s), 55 °C (30 s), and 72 °C (1 min); 7 min at 72 °C. Real-time RT-PCR was performed using Applied Biosystems 7300 Real Time PCR System (Thermo Fischer Scientific, Waltham, Massachusetts, USA) with Thunderbird SYBR qPCR Mix (Toyobo, Osaka, Japan). Three biological repetitions were performed, and the standard deviation was calculated. The DNA was quantitated using a standard curve and was normalized to the amount of 18S rDNA.
4.4. Sequence Analysis
The RT of OLE from the genome of cruciferous vegetables was amplified by PCR. The PCR primers (Supplementary Table S1) were designed based on the RT of ONSEN from A. thaliana genome (TAIR10 Whole genome). The PCR fragments were sequenced after cloning into pGEM-T Easy Vector (Promega) (Supplementary Figure S2).
The whole genome sequences of B. rapa [26], B. nigra [27], B. oleracea [28,29], B. juncea [27], and B. napus [30] were used. The reverse transcriptase core domain region from the A. thaliana ONSEN copies was used as a query in the homology search performed using the BLAST server of NCBI and Phytozome ver 10 [31]. The sequences showing at least 50% homology were retained for alignment. The aligned sequences were then checked manually to delete the sequences with more than 100 bp ambiguous or missing sites.
The LTR sequences were obtained from the regions flanking the RT sequence in the whole genome sequences. LTR finder [32] was used for searching the LTR candidates in the 5-kb region flanking the RT sequence. The candidate LTR sequences were then searched in the flanking regions of the LTR finder-negative sequences. The obtained LTRs were aligned and the p-distance was calculated for the pair of LTRs. The phylogenetic trees for the RT regions and LTR were constructed by the neighbor-joining method with JC distances. The bootstrap probabilities were calculated by 500 replications. All the phylogenetic analyses were done by MEGA ver 7.0 [33].
Supplementary Materials
The following are available online at https://www.mdpi.com/2075-4655/1/1/7/s1. Figure S1: The structure of HREs within the ONSEN LTRs. Figure S2. A reverse transcriptase (RT) of OLE from the genome of cruciferous vegetables. Table S1: Primer sequences.
Acknowledgments
We thank So Nakagawa for bioinformatic assistance. This work was supported by Suharakinennzaidann; Cooperative Research Grant of the Plant Transgenic Design Initiative, Gene Research Center, the University of Tsukuba, NIG-JOINT (32A2017); the Joint Research Program of Arid Land Research Center, Tottori University (28C2002); and a Grant-in-Aid for scientific Research on Innovative Areas (JP15H05960).
Author Contributions
K.N. performed most of the experiments and analyzed data; Y.K., M.S., and Y.M. contributed with experimental work; A.k.K. contributed phylogenic analysis; A.k.K. and A.t.K. supervised the work and designed experimental approaches; and A.k.K. and H.I. wrote the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kumar, A.; Bennetzen, J.L. Plant retrotransposons. Annu. Rev. Genet. 1999, 33, 479–532. [Google Scholar] [CrossRef] [PubMed]
- SanMiguel, P.; Gaut, B.S.; Tikhonov, A.; Nakajima, Y.; Bennetzen, J.L. The paleontology of intergene retrotransposons of maize. Nat. Genet. 1998, 20, 43–45. [Google Scholar] [CrossRef] [PubMed]
- Vitte, C.; Bennetzen, J.L. Analysis of retrotransposon structural diversity uncovers properties and propensities in angiosperm genome evolution. Proc. Natl. Acad. Sci. USA 2006, 103, 17638–17643. [Google Scholar] [CrossRef] [PubMed]
- Henderson, I.R.; Jacobsen, S.E. Epigenetic inheritance in plants. Nature 2007, 447, 418–424. [Google Scholar] [CrossRef] [PubMed]
- Lisch, D. Epigenetic Regulation of Transposable Elements in Plants. Annu. Rev. Plant Biol. 2009, 60, 43–66. [Google Scholar] [CrossRef] [PubMed]
- Slotkin, R.K.; Martienssen, R. Transposable elements and the epigenetic regulation of the genome. Nat. Rev. Genet. 2007, 8, 272–285. [Google Scholar] [CrossRef] [PubMed]
- Feschotte, C.; Jiang, N.; Wessler, S.R. Plant transposable elements: Where genetics meets genomics. Nat. Rev. Genet. 2002, 3, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Ito, H.; Gaubert, H.; Bucher, E.; Mirouze, M.; Vaillant, I.; Paszkowski, J. An siRNA pathway prevents transgenerational retrotransposition in plants subjected to stress. Nature 2011, 472, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Cavrak, V.V.; Lettner, N.; Jamge, S.; Kosarewicz, A.; Bayer, L.M.; Scheid, O.M. How a Retrotransposon Exploits the Plant's Heat Stress Response for Its Activation. PLoS Genet. 2014, 10. [Google Scholar] [CrossRef] [PubMed]
- Masuta, Y.; Nozawa, K.; Takagi, H.; Yaegashi, H.; Tanaka, K.; Ito, T.; Saito, H.; Kobayashi, H.; Matsunaga, W.; Masuda, S.; et al. Inducible Transposition of a Heat-Activated Retrotransposon in Tissue Culture. Plant Cell Physiol. 2017, 58, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Ito, H.; Kim, J.M.; Matsunaga, W.; Saze, H.; Matsui, A.; Endo, T.A.; Harukawa, Y.; Takagi, H.; Yaegashi, H.; Masuta, Y.; et al. A Stress-Activated Transposon in Arabidopsis Induces Transgenerational Abscisic Acid Insensitivity. Sci. Rep. 2016, 6, 23181. [Google Scholar] [CrossRef] [PubMed]
- Warwick, S.I.; Francis, A.; Al-Shehbaz, I.A. Brassicaceae: Species checklist and database on CD-Rom. Plant Syst. Evol. 2006, 259, 249–258. [Google Scholar] [CrossRef]
- Maumus, F.; Allen, A.E.; Mhiri, C.; Hu, H.H.; Jabbari, K.; Vardi, A.; Grandbastien, M.A.; Bowler, C. Potential impact of stress activated retrotransposons on genome evolution in a marine diatom. BMC Genom. 2009, 10, 624. [Google Scholar] [CrossRef] [PubMed]
- Naito, K.; Zhang, F.; Tsukiyama, T.; Saito, H.; Hancock, C.N.; Richardson, A.O.; Okumoto, Y.; Tanisaka, T.; Wessler, S.R. Unexpected consequences of a sudden and massive transposon amplification on rice gene expression. Nature 2009, 461, 1130–1134. [Google Scholar] [CrossRef] [PubMed]
- Lisch, D. How important are transposons for plant evolution? Nat. Rev. Genet. 2013, 14, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Butelli, E.; Licciardello, C.; Zhang, Y.; Liu, J.J.; Mackay, S.; Bailey, P.; Reforgiato-Recupero, G.; Martin, C. Retrotransposons Control Fruit-Specific, Cold-Dependent Accumulation of Anthocyanins in Blood Oranges. Plant Cell 2012, 24, 1242–1255. [Google Scholar] [CrossRef] [PubMed]
- Makarevitch, I.; Waters, A.J.; West, P.T.; Stitzer, M.; Hirsch, C.N.; Ross-Ibarra, J.; Springer, N.M. Transposable elements contribute to activation of maize genes in response to abiotic stress. PLoS Genet. 2015, 11, e1004915. [Google Scholar] [CrossRef]
- Ito, H.; Yoshida, T.; Tsukahara, S.; Kawabe, A. Evolution of the ONSEN retrotransposon family activated upon heat stress in Brassicaceae. Gene 2013, 518, 256–261. [Google Scholar] [CrossRef] [PubMed]
- Pietzenuk, B.; Markus, C.; Gaubert, H.; Bagwan, N.; Merotto, A.; Bucher, E.; Pecinka, A. Recurrent evolution of heat-responsiveness in Brassicaceae COPIA elements. Genome Biol. 2016, 17, 209. [Google Scholar] [CrossRef] [PubMed]
- Mari-Ordonez, A.; Marchais, A.; Etcheverry, M.; Martin, A.; Colot, V.; Voinnet, O. Reconstructing de novo silencing of an active plant retrotransposon. Nat. Genet. 2013, 45, 1029–1039. [Google Scholar] [CrossRef] [PubMed]
- Grandbastien, M.A. Activation of plant retrotransposons under stress conditions. Trends Plant Sci. 1998, 3, 181–187. [Google Scholar] [CrossRef]
- Liu, B.; Wendel, J.F. Retrotransposon activation followed by rapid repression in introgressed rice plants. Genome 2000, 43, 874–880. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.P.; Si, Y.; Hanson, R.E.; Crane, C.F.; Price, H.J.; Stelly, D.M.; Wendel, J.F.; Paterson, A.H. Dispersed repetitive DNA has spread to new genomes since polyploid formation in cotton. Genome Res. 1998, 8, 479–492. [Google Scholar] [CrossRef] [PubMed]
- Miura, A.; Kato, M.; Watanabe, K.; Kawabe, A.; Kotani, H.; Kakutani, T. Genomic localization of endogenous mobile CACTA family transposons in natural variants of Arabidopsis thaliana. Mol. Genet. Genom. 2004, 270, 524–532. [Google Scholar] [CrossRef] [PubMed]
- Church, G.M.; Gilbert, W. Genomic sequencing. Proc. Natl. Acad. Sci. USA 1984, 81, 1991–1995. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.W.; Wang, H.Z.; Wang, J.; Sun, R.F.; Wu, J.; Liu, S.Y.; Bai, Y.Q.; Mun, J.H.; Bancroft, I.; Cheng, F.; et al. The genome of the mesopolyploid crop species Brassica rapa. Nat. Genet. 2011, 43, 1035–1039. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Liu, D.; Wang, X.; Ji, C.; Cheng, F.; Liu, B.; Hu, Z.; Chen, S.; Pental, D.; Ju, Y.; et al. The genome sequence of allopolyploid Brassica juncea and analysis of differential homoeolog gene expression influencing selection. Nat. Genet. 2016, 48, 1225–1232. [Google Scholar] [CrossRef] [PubMed]
- Parkin, I.A.; Koh, C.; Tang, H.; Robinson, S.J.; Kagale, S.; Clarke, W.E.; Town, C.D.; Nixon, J.; Krishnakumar, V.; Bidwell, S.L.; et al. Transcriptome and methylome profiling reveals relics of genome dominance in the mesopolyploid Brassica oleracea. Genome Biol. 2014, 15, R77. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Liu, Y.; Yang, X.; Tong, C.; Edwards, D.; Parkin, I.A.; Zhao, M.; Ma, J.; Yu, J.; Huang, S.; et al. The Brassica oleracea genome reveals the asymmetrical evolution of polyploid genomes. Nat. Commun. 2014, 5, 3930. [Google Scholar] [CrossRef] [PubMed]
- Chalhoub, B.; Denoeud, F.; Liu, S.; Parkin, I.A.; Tang, H.; Wang, X.; Chiquet, J.; Belcram, H.; Tong, C.; Samans, B.; et al. Plant genetics. Early allopolyploid evolution in the post-Neolithic Brassica napus oilseed genome. Science 2014, 345, 950–953. [Google Scholar] [CrossRef] [PubMed]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Wang, H. LTR_FINDER: An efficient tool for the prediction of full-length LTR retrotransposons. Nucleic Acids Res. 2007, 35, W265–W268. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Southern blot analysis for determining the copies of ONSEN-like element (OLE). Southern blot analysis for determining the copies of ONSEN-like element (OLE) in 17 cruciferous vegetables (A) and B. napus (B). The genomic DNA was digested with SspI and hybridized with an ONSEN-specific probe. The arrow in (A) indicates the conserved copy in the same species. A gel stained with ethidium bromide (EtBr) is shown at the bottom of each panel as a loading control.
Figure 1.
Southern blot analysis for determining the copies of ONSEN-like element (OLE). Southern blot analysis for determining the copies of ONSEN-like element (OLE) in 17 cruciferous vegetables (A) and B. napus (B). The genomic DNA was digested with SspI and hybridized with an ONSEN-specific probe. The arrow in (A) indicates the conserved copy in the same species. A gel stained with ethidium bromide (EtBr) is shown at the bottom of each panel as a loading control.
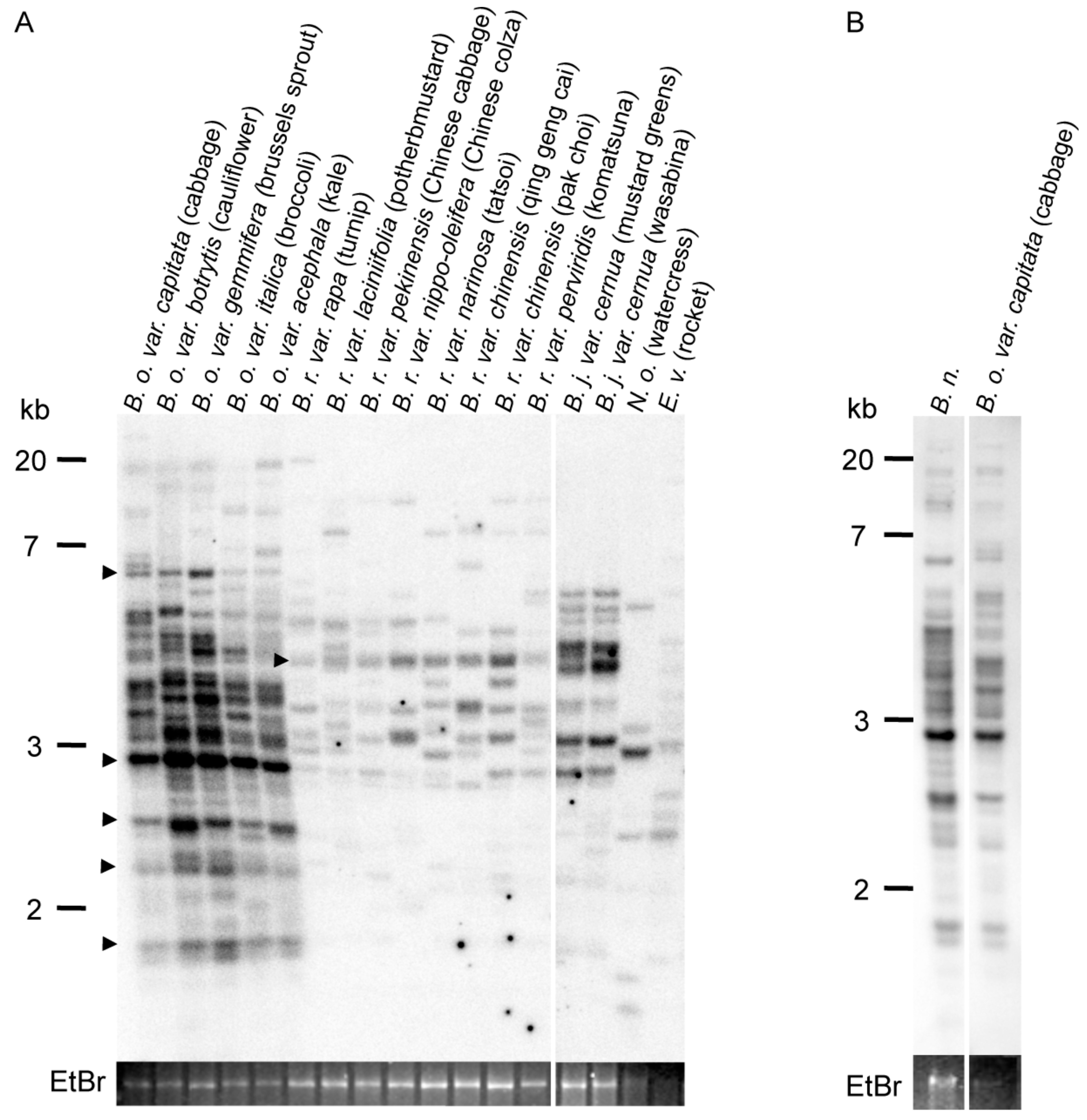
Figure 2.
Heat-activation of OLE in cruciferous vegetables (A) RT-PCR of ONSEN-like elements (OLEs) in 17 Cruciferous vegetables. 18S rDNA was used as a control. (B) Southern blot of non-digested DNA for detecting the extrachromosomal DNA (5 kb) of OLEs in cruciferous vegetables. A gel stained with ethidium bromide (EtBr) is shown at the bottom of each panel as a loading control.
Figure 2.
Heat-activation of OLE in cruciferous vegetables (A) RT-PCR of ONSEN-like elements (OLEs) in 17 Cruciferous vegetables. 18S rDNA was used as a control. (B) Southern blot of non-digested DNA for detecting the extrachromosomal DNA (5 kb) of OLEs in cruciferous vegetables. A gel stained with ethidium bromide (EtBr) is shown at the bottom of each panel as a loading control.
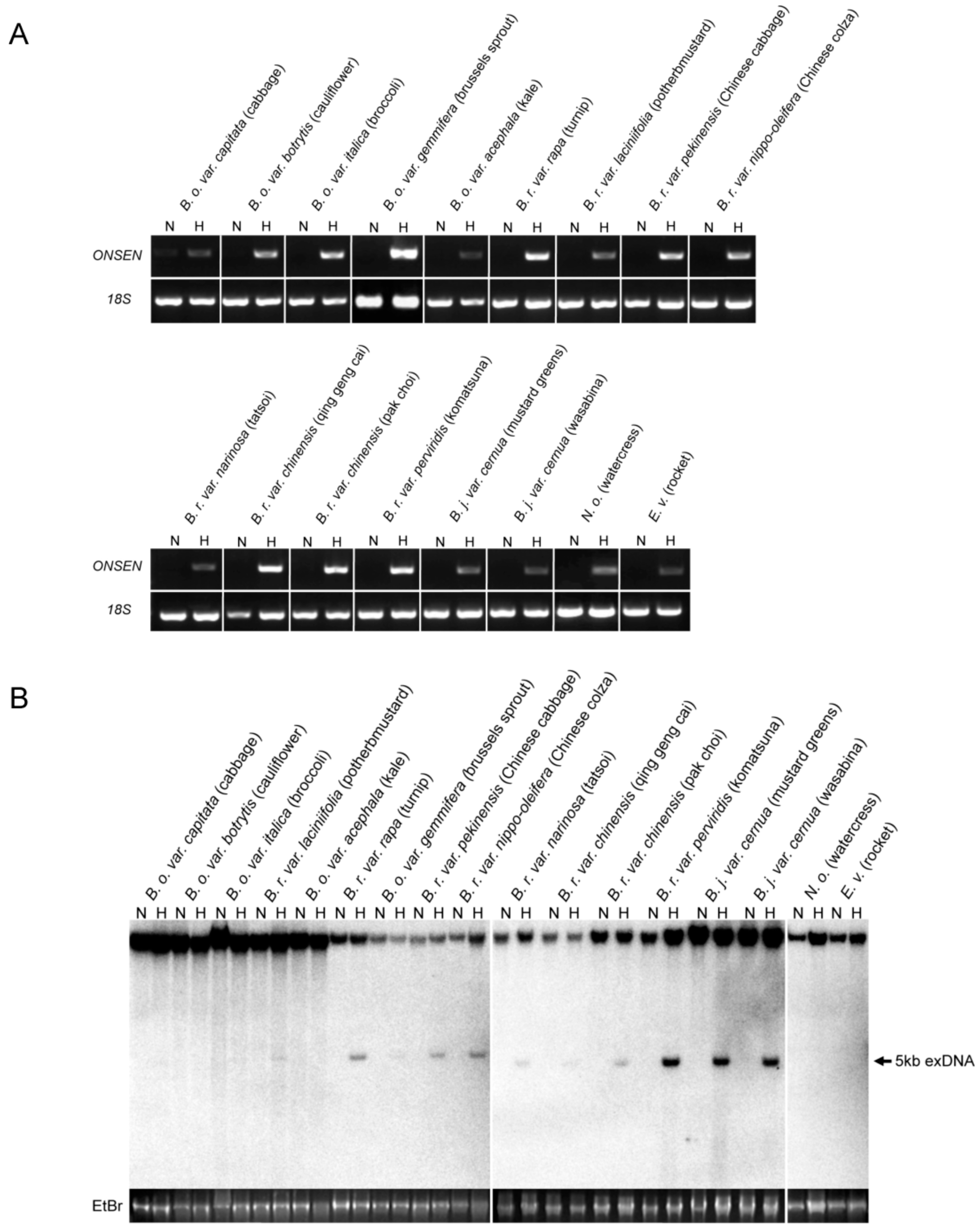
Figure 3.
Phylogenetic relationship of the reverse transcriptase region in Brassica species. The phylogenetic relationship was represented by a tree constructed using the neighbor-joining method. All the trees are shown in the same scale; the scale bars are shown beside each tree. The description of marker is shown in the middle at the top of the figure. The A. thaliana sequences were also included for whole genome sequence based analyses. The bootstrap values of the major clades are indicated beside the branches. (A) Sequences from the whole genome of Brassica species, (B) B. rapa, (C) B. juncea, (D) B. oleracea.
Figure 3.
Phylogenetic relationship of the reverse transcriptase region in Brassica species. The phylogenetic relationship was represented by a tree constructed using the neighbor-joining method. All the trees are shown in the same scale; the scale bars are shown beside each tree. The description of marker is shown in the middle at the top of the figure. The A. thaliana sequences were also included for whole genome sequence based analyses. The bootstrap values of the major clades are indicated beside the branches. (A) Sequences from the whole genome of Brassica species, (B) B. rapa, (C) B. juncea, (D) B. oleracea.
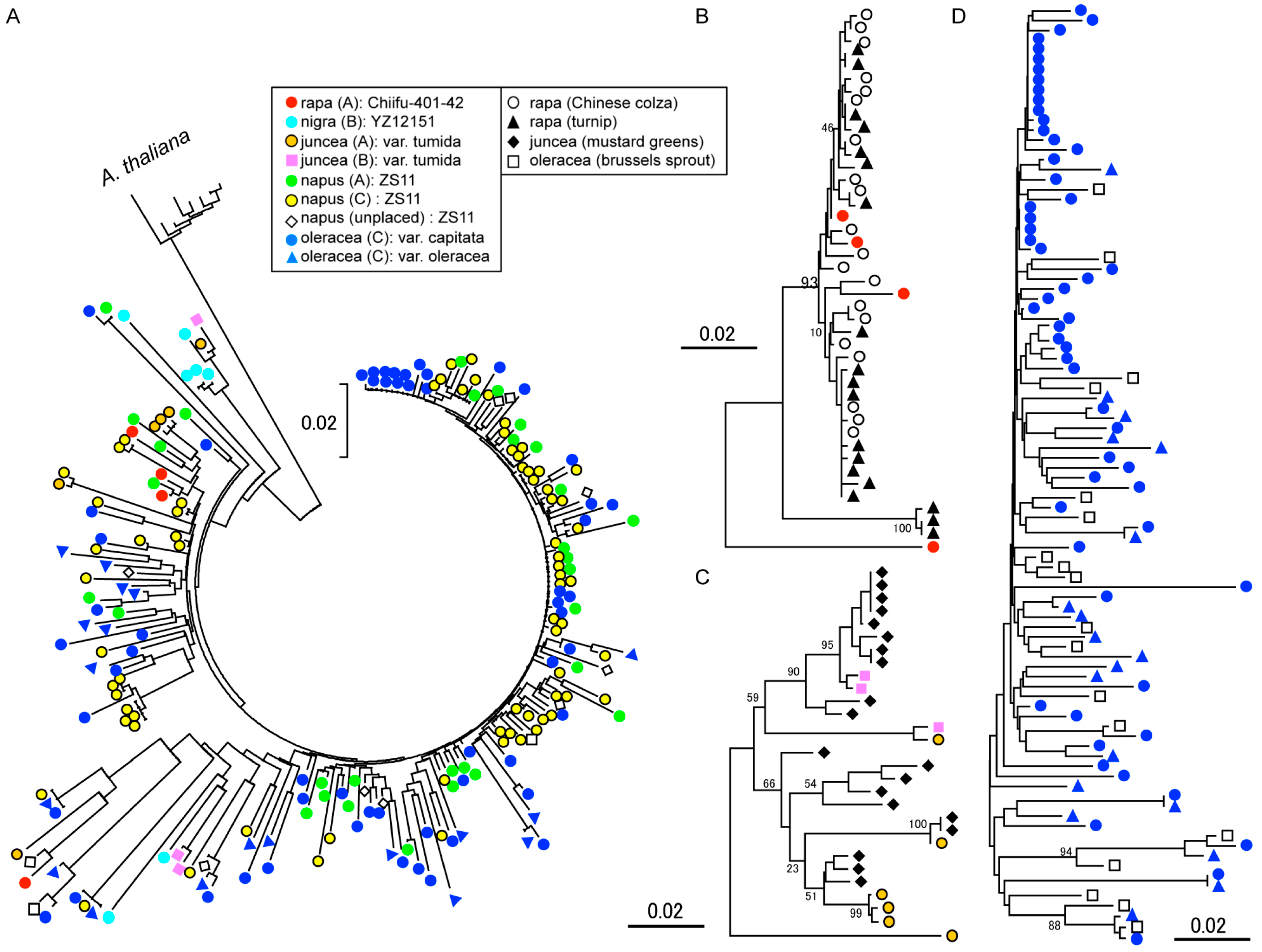
Figure 4.
Evolutionary analyses of long terminal repeat (LTR) sequences. (A) A phylogenetic tree is shown. The description of marker is shown in the bottom at the right of the tree. The scale bar is shown in the middle at the top. For B. rapa and B. oleracea, copies from the different genome surveys are indicated with different marks. The bootstrap values for the major clades are shown beside the branches. (B) Scatter plot of LTR identities. The LTR identities are shown for each species. The number of pairs is shown in the parentheses. (C) Distribution of LTR identities for B. rapa, B. oleracea, and B. napus. The locations of genomes are indicated as A: right hatched bar, C: filled bar, unplaced scaffold: open bar.
Figure 4.
Evolutionary analyses of long terminal repeat (LTR) sequences. (A) A phylogenetic tree is shown. The description of marker is shown in the bottom at the right of the tree. The scale bar is shown in the middle at the top. For B. rapa and B. oleracea, copies from the different genome surveys are indicated with different marks. The bootstrap values for the major clades are shown beside the branches. (B) Scatter plot of LTR identities. The LTR identities are shown for each species. The number of pairs is shown in the parentheses. (C) Distribution of LTR identities for B. rapa, B. oleracea, and B. napus. The locations of genomes are indicated as A: right hatched bar, C: filled bar, unplaced scaffold: open bar.

Figure 5.
Epigenetic regulation of OLE in cruciferous vegetables (A) Relative transcript levels of ONSEN-like element (OLEs) with or without 5AzaC treatment in four Brassica species. NS: non-stressed, HS: heat stressed. Asterisks mark significant differences (p < 0.05). (B) Young seedlings with or without the 5AzaC treatment in four Brassica species. (C) Southern blot of non-digested DNA for detecting the extrachromosomal DNA with or without the 5AzaC treatment in B. oleracea (B.O.) and B. napus (B. n.). NS: non-stressed, HS: heat stressed. The arrowhead indicates the 5 kb exDNA.
Figure 5.
Epigenetic regulation of OLE in cruciferous vegetables (A) Relative transcript levels of ONSEN-like element (OLEs) with or without 5AzaC treatment in four Brassica species. NS: non-stressed, HS: heat stressed. Asterisks mark significant differences (p < 0.05). (B) Young seedlings with or without the 5AzaC treatment in four Brassica species. (C) Southern blot of non-digested DNA for detecting the extrachromosomal DNA with or without the 5AzaC treatment in B. oleracea (B.O.) and B. napus (B. n.). NS: non-stressed, HS: heat stressed. The arrowhead indicates the 5 kb exDNA.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Nozawa, K.; Kawagishi, Y.; Kawabe, A.; Sato, M.; Masuta, Y.; Kato, A.; Ito, H. Epigenetic Regulation of a Heat-Activated Retrotransposon in Cruciferous Vegetables. Epigenomes 2017, 1, 7. https://doi.org/10.3390/epigenomes1010007
AMA Style
Nozawa K, Kawagishi Y, Kawabe A, Sato M, Masuta Y, Kato A, Ito H. Epigenetic Regulation of a Heat-Activated Retrotransposon in Cruciferous Vegetables. Epigenomes. 2017; 1(1):7. https://doi.org/10.3390/epigenomes1010007
Chicago/Turabian StyleNozawa, Kosuke, Yuki Kawagishi, Akira Kawabe, Mio Sato, Yukari Masuta, Atsushi Kato, and Hidetaka Ito. 2017. "Epigenetic Regulation of a Heat-Activated Retrotransposon in Cruciferous Vegetables" Epigenomes 1, no. 1: 7. https://doi.org/10.3390/epigenomes1010007