The Metabolic and Ecological Interactions of Oxalate-Degrading Bacteria in the Mammalian Gut

Abstract
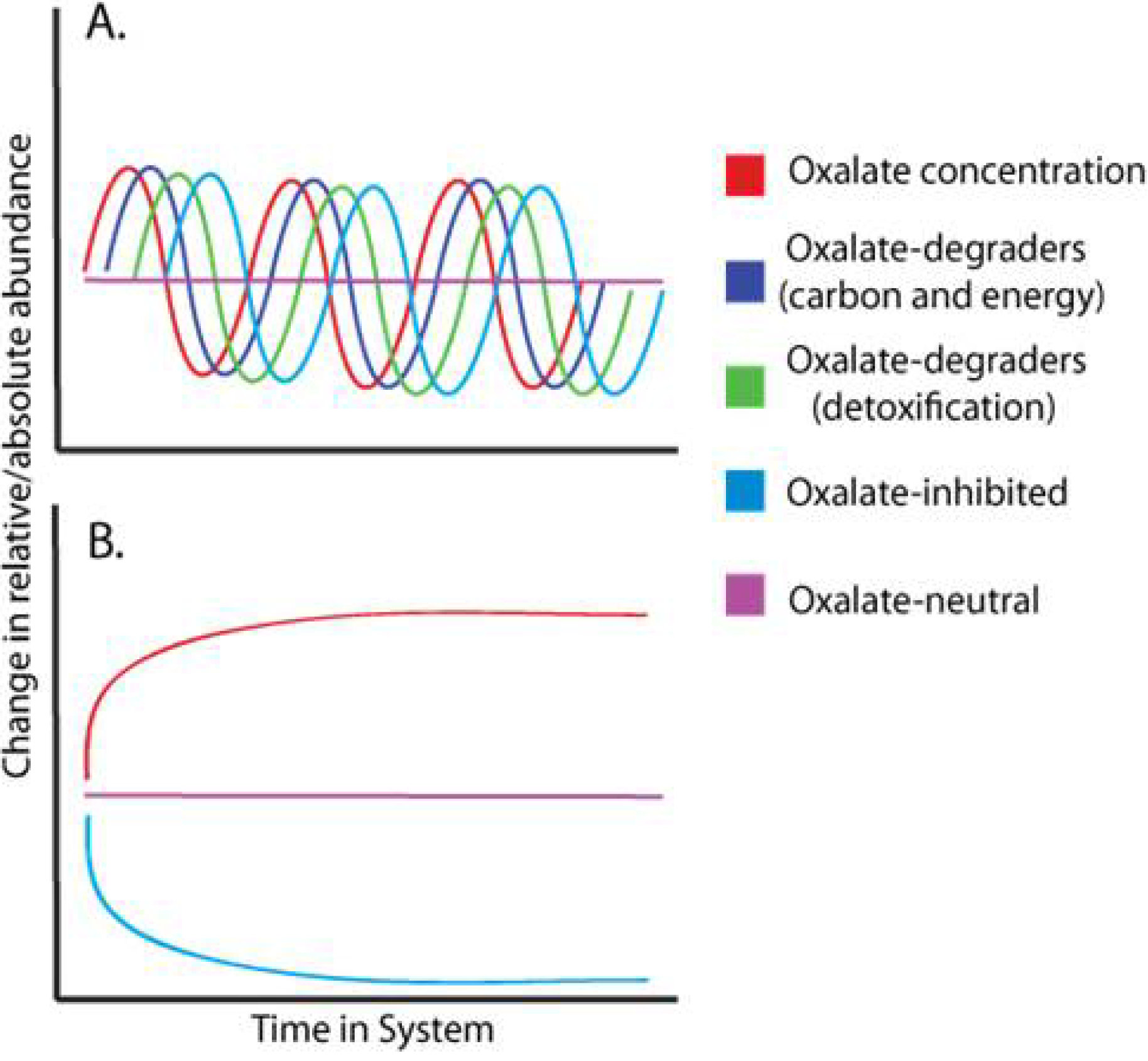
:1. Introduction
2. The Flow of Oxalate in the Body
3. Microbial Oxalate Degradation

{kind=link}
| Organism | Source | Pathway | References |
|---|---|---|---|
| Oxalobacter formigenes | Various mammals | carbon/energy | [60,65] |
| Eggerthella lenta | Human stool | N/A | [61,66] |
| Enterococcus gallinarum | Woodrat feces | detoxification | [64] |
| Enterococcus faecium | Canine feces | N/A | [67] |
| Enterococcus faecalis | Human stool, canine feces | carbon/energy | [62,67] |
| Provendencia rettgeri | Human stool | N/A | [68] |
| Streptococcus thermophilus | Probiotic | detoxification | [63] |
| Lactobacillus plantarum | Probiotic, canine/feline feces | detoxification | [69,70] |
| Lactobacillus gasseri | Probiotic, woodrat gut | detoxification | [64,70] |
| Lactobacillus casei | Probiotic | detoxification | [70,71,72] |
| Lactobacillus acidophilus | Human stool | detoxification | [73] |
| Lactobacillus rhamnosus | Probiotic | detoxification | [70] |
| Lactobacillus salviarius | Probiotic | detoxification | [70] |
| Lactobacillus johnsonii | Woodrat gut | detoxification | [64] |
| Bifidobacterium infantis | Probiotic | detoxification | [63] |
| Bifidobacterium animalis | Human stool | detoxification | [70] |
| Clostridium sporogenes | Woodrat feces | detoxification | [64] |
| Leuconostoc lactis | Canine/feline feces | N/A | [69] |
| Leuconostoc mesenteroides | Canine feces | N/A | [72] |
4. The Gut Ecosystem Mediates Microbial Oxalate Degradation
5. Ecological Interactions within the Gut Microbiota in Response to Oxalate

6. Oxalate-Degrading Bacteria as Probiotics
7. Evolutionary Considerations of Oxalate-Degrading Bacteria
8. Conclusions
Acknowledgments
Conflicts of Interest
References
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Hooper, L.V.; Littman, D.R.; Macpherson, A.J. Interactions between the gut microbiota and the immune system. Science 2012, 336, 1268–1273. [Google Scholar] [CrossRef]
- Nicholson, J.K.; Holmes, E.; Kinross, J.; Burcelin, R.; Gibson, G.; Jia, W.; Petterson, S. Host-gut microbiota metabolic interactions. Science 2012, 336, 1262–1267. [Google Scholar] [CrossRef]
- Guarner, F.; Bourdet-Sicard, R.; Brandtzaeg, P.; Gill, H.S.; McGuirk, P.; van Eden, W.; Versalovic, J.; Weinstock, J.V.; Rook, G.A.M. Mechanisms of disease: The hygiene hypothesis revisited. Nat. Rev. Gastro. Hepat. 2006, 3, 275–284. [Google Scholar]
- O’Hara, A.M.; Shanahan, F. The gut microbiota as a forgotten organ. EMBO 2006, 7, 688–693. [Google Scholar] [CrossRef]
- Dethlefsen, L.; McFall-Ngai, M.; Relman, D.A. An ecological and evolutionary perspective on human-microbe mutualism and disease. Nature 2007, 449, 811–818. [Google Scholar] [CrossRef]
- Martin, F.P.J.; Wang, Y.; Sprenger, N.; Yap, I.K.S.; Lundstedt, T.; Lek, P.; Rezzi, S.; Ramadan, Z.; van Bladeren, P.; Fay, L.B.; et al. Probiotic modulation of symbiotic gut microbial-host metabolic interactions in a humanized microbiome mouse model. Mol. Syst. Biol. 2008, 4, 1–15. [Google Scholar]
- Kaufman, D.W.; Kelly, J.P.; Curhan, G.C.; Anderson, T.E.; Dretler, S.P.; Preminger, G.M.; Cave, D.R. Oxalobacter formigenes may reduce the risk of calcium oxalate kidney stones. J. Am. Soc. Nephro. 2008, 19, 1197–1203. [Google Scholar] [CrossRef]
- Round, J.L.; Mazmanian, S.K. The gut microbiota shapes intestinal immune responses during health and disease. Nat. Rev. Immun. 2009, 9, 313–323. [Google Scholar] [CrossRef]
- Gómez-Hurtado, I.; Santacruz, A.; Peiró, G.; Zapater, P.; Gutiérrez, A.; Pérez-Mateo, M.; Sanz, Y.; Francés, R. Gut microbiota dysbiosis is associated with inflammation and bacterial translocation in mice with CCL4-induced fibrosis. PLoS One 2011. [Google Scholar] [CrossRef]
- Baker, P.I.; Love, D.R.; Ferguson, L.R. Role of gut microbiota in Crohn’s disease. Expert Rev. Gastroenterol. Hepatol. 2009, 3, 535–546. [Google Scholar] [CrossRef]
- Vijay-Kumar, M.; Aitken, J.D.; Carvalho, F.A.; Cullender, T.C.; Mwangi, S.; Srinivasan, S.; Sitaraman, S.V.; Knight, R.; Ley, R.E.; Gewirtz, A.T. Metabolic syndrome and altered gut microbiota in mice lacking toll-like receptor 5. Science 2010, 328, 228–231. [Google Scholar] [CrossRef]
- Musso, G.; Gambino, R.; Cassader, M. Interactions between gut microbiota and host metabolism predisposing to obesity and diabetes. Ann. Rev. Med. 2011, 62, 361–380. [Google Scholar] [CrossRef]
- Chung, W.G.; Roh, H.K.; Kim, H.M.; Cha, Y.N. Involvement of CYP3A1, 2B1 and 2E1 in C-8 hydroxylation and CYP1A2 and flavin-containing monooxygenase in N-demethylation of caffeine: Identified by using inducer treated rat liver microsomes that are characterized with testosterone metabolic patterns. Chem. Biol. Interact. 1998, 113, 1–14. [Google Scholar] [CrossRef]
- Holmes, R.P.; Goodman, H.O.; Assimos, D.G. Contribution of dietary oxalate to urinary oxalate excretion. Kidney Int. 2001, 59, 270–276. [Google Scholar] [CrossRef]
- Forbey, J.S.; Harvey, A.L.; Huffman, M.A.; Provenza, F.D.; Sullivan, R.; Tasdemir, D. Exploitation of secondary metabolites by animals: A response to homeostatic challenges. Int. Comp. Biol. 2009, 49, 314–328. [Google Scholar] [CrossRef]
- Forbey, J.S.; Dearing, M.D.; Gross, E.M.; Orians, C.M.; Sotka, E.E.; Foley, W.J. A pharm-ecological approach of terrestrial and aquatic plant-herbivore interactions. J. Chem. Ecol. 2013, 39, 465–480. [Google Scholar] [CrossRef]
- Torregrossa, A.M.; Dearing, M.D. Nutritional ecology of mammals: Regulated intake of plant secondary compounds. Funct. Ecol. 2009, 23, 48–56. [Google Scholar] [CrossRef]
- Dearing, M.D.; Foley, W.J.; McClean, S. The influence of plant secondary metabolites on the nutritional ecology of herbivorous terrestrial vertebrates. Ann. Rev. Ecol. Evol. Syst. 2005, 36, 169–189. [Google Scholar] [CrossRef]
- Freeland, W.J.; Jansen, D.H. Strategies in herbivory by mammals. Am. Nat. 1974, 108, 269–288. [Google Scholar]
- Karasov, W.H.; Carey, H.V. Metabolic teamwork between gut microbes and host. Microbe 2009, 4, 323–328. [Google Scholar]
- Jones, R.J.; Megarrity, R.G. Successful transfer of DHP-degrading bacteria from Hawaiian goats to Australian ruminants to overcome toxicity of Leucaena. Aust. Vet. J. 1986, 63, 259–262. [Google Scholar] [CrossRef]
- Sasaki, E.; Shimada, T.; Osawa, R.; Nishitani, Y.; Spring, S.; Lang, E. Isolation of tannin-degrading bacteria from feces of the Japanese large wood mouse, Apodemus speciosus, feeding on tannin-rich acorns. Syst. Appl. Microbiol. 2005, 28, e358. [Google Scholar] [CrossRef]
- Shimada, T.; Saitoh, T.; Sasaki, E.; Nishitani, Y.; Osawa, R. Role of tannin-binding salivary proteins and tannase-producing bacteria in the acclimation of the Japanese wood mouse to acorn tannins. J. Chem. Ecol. 2006, 32, 1165–1180. [Google Scholar] [CrossRef]
- Rodríguez, H.; de la Rivas, B.; Gómez-Cordovés, C.; Muñez, R. Degradation of tannic acid by cell-free extracts of Lactobacillus planarum. Food Chem. 2008, 107, 664–670. [Google Scholar] [CrossRef] [Green Version]
- Hiura, D.; Hashidoko, Y.; Kobayashi, Y.; Tahara, S. Effective degradation of tannic acid by immobilized rumen microbes of a sika deer (Cervus nippon yesoensis) in winter. Anim. Feed. Sci.Tech. 2010, 155, 1–8. [Google Scholar] [CrossRef]
- Ley, R.E.; Hamady, M.; Lozupone, C.; Turnbaugh, P.; Ramey, R.R.; Bircher, J.S.; Schegel, M.L.; Tucker, T.A.; Schrenzel, M.D.; Knight, R.; et al. Evolution of mammals and their gut microbes. Science 2008, 320, 1647–1651. [Google Scholar] [CrossRef]
- Muegge, B.D.; Kuczynski, J.; Knights, D.; Clemente, J.C.; González, A.; Fontana, L.; Henrissat, B.; Knight, R.; Gordon, J.I. Diet drives convergence in gut microbiome functions across mammalian phylogeny and within humans. Science 2011, 332, 970–973. [Google Scholar] [CrossRef]
- Conyers, R.A.J.; Bals, R.; Rofe, A.M. The relation of clinical catastrophes, endogenous oxalate production, and urolithiasis. Clin. Chem. 1990, 36, 1717–1730. [Google Scholar]
- Holmes, R.P.; Kennedy, M. Estimation of the oxalate content of foods and daily oxalate intake. Kidney Int. 2000, 57, 1662–1667. [Google Scholar] [CrossRef]
- Jaeger, P.; Robertson, W.G. Role of dietary intake and intestinal absorption of oxalate in calcium stone formation. Nephron Physiol. 2004, 98, 64–71. [Google Scholar] [CrossRef]
- Franceschi, V.R.; Nakata, P.A. Calcium oxalate in plants: Formation and function. Ann. Rev. Plant Biol. 2005, 56, 41–71. [Google Scholar] [CrossRef]
- Knight, J.; Jiang, J.; Assimos, D.G.; Holmes, R.P. Hydroxyproline ingestion and urinary oxalate and glyoxalate excretion. Kidney Int. 2006, 70, 1929–1934. [Google Scholar]
- Massey, L.K. Food oxalate: Factors affecting measurement, biological variation, and bioavailability. J. Am. Diet. Assoc. 2007, 107, 1191–1194. [Google Scholar] [CrossRef]
- Taylor, E.N.; Curhan, G.C. Determinants of 24-hour urinary oxalate excretion. Clin. J. Am. Soc. Nephrol. 2008, 3, 1453–1460. [Google Scholar] [CrossRef]
- Hess, B.; Jost, C.; Zipperle, L.; Takkinen, R.; Jaegar, P. High-calcium intake abolishes hyperoxaluria and reduces urinary crystallization during a 20-fold normal oxalate load in humans. Nephrol. Dial. Transpl. 1998, 13, 2241–2247. [Google Scholar] [CrossRef]
- Aslani, M.R.; Movassaghi, A.R.; Najarnezhad, V.; Pirouz, H.J.; Bami, M.H. Acute oxalate intoxication associated to ingestion of eshnan (Seidlitzia rosmarinus) in sheep. Trop. Anim. Health Prod. 2011, 43, 1065–1068. [Google Scholar] [CrossRef]
- James, L.F.; Butcher, J.E. Halogeton poisoning of sheep: Effect of high level of oxalate intake. J. Anim. Sci. 1972, 35, 1233–1238. [Google Scholar]
- Concon, J.M. Food Toxicology—Principles and Concepts; Marcel Dekker: New York, NY, USA, 1988. [Google Scholar]
- Amoroso, A.; Pirulli, D.; Florian, F.; Puzzer, D.; Boniotto, M.; Crovella, S.; Zezlina, S.; Spano, A.; Mazzola, G.; Savoldi, S.; et al. AGXT gene mutations and their influence on clinical heterogeneity of type 1 primary hyperoxaluria. J. Am. Soc. Nephrol. 2001, 12, 2072–2079. [Google Scholar]
- Siener, R.; Ebert, D.; Nicolay, C.; Hesse, A. Dietary risk factors for hyperoxaluria in calcium oxalate stone formers. Kidney Int. 2003, 63, 1037–1043. [Google Scholar] [CrossRef]
- Coe, F.L.; Evan, A.; Worcester, E. Kidney stone disease. J. Clin. Invest. 2005, 115, 2598–2608. [Google Scholar] [CrossRef]
- Hodgkinson, A. Oxalic Acid in Biology and Medicine; Academic Press: New York, NY, USA, 1977. [Google Scholar]
- Hoppe, B.; Beck, B.; Gatter, N.; von Unruh, G.; Tischer, A.; Hesse, A.; Laube, N.; Kaul, P.; Sidhu, H. Oxalobacter formigenes: A potential tool for the treatment of primary hyperoxaluria type I. Kidney Int. 2006, 70, 1305–1311. [Google Scholar] [CrossRef]
- Sidhu, H.; Schmidt, M.E.; Cornelius, J.G.; van Thamilsel, S.; Khan, S.R.; Hesse, A.; Peck, A.B. Direct correlation between hyperoxaluria/oxalate stone disease and the absence of the gastrointestinal tract-dwelling bacterium Oxalobacter formigenes: Possible prevention by gut recolonization or enzyme replacement therapy. J. Am. Soc. Nephrol. 1999, 10, 334–340. [Google Scholar]
- Sidhu, H.; Allison, M.J.; May Chow, J.O.; Clark, A.; Peck, A.B. Rapid reversal of hyperoxaluria in a rat model after probiotic administration of Oxalobacter formigenes. J. Urol. 2001, 166, 1487–1491. [Google Scholar] [CrossRef]
- Justice, K.E. Oxalate digestability in Neotoma albigula and Neotoma mexicana. Oecologia 1985, 67, 231–234. [Google Scholar] [CrossRef]
- Ruiz, N.; Ward, D.; Saltz, D. Calcium oxalate crystals in leaves of Pancratium sickenbergeri: Constitutive or induced defense? Funct. Ecol. 2002, 16, 99–105. [Google Scholar] [CrossRef]
- Palgi, N.; Ronen, Z.; Pinshow, B. Oxalate balance in fat sand rats feeding on high and low calcium diets. J. Comp. Phys. B 2008, 178, 617–622. [Google Scholar] [CrossRef]
- Noonan, S.C.; Savage, G.P. Oxalate content of foods and its effect on humans. Asia Pacific J. Clin. Nutr. 1999, 8, 64–74. [Google Scholar] [CrossRef]
- Contreras-Padilla, M.; Pérez-Torrero, E.; Hernández-Urbiola, M.I.; Hernández-Quevedo, G.; del Real, A.; Rivera-Muñoz, E.M.; Rodríguez-García, M.E. Evaluation of oxalates and calcium in nopal pads (Opuntia ficus-indica var. redonda) at different maturity stages. J. Food Comp. Anal. 2011, 24, 38–43. [Google Scholar] [CrossRef]
- Hatch, M.; Cornelius, J.; Allison, M.; Sidhu, H.; Peck, A.; Freel, R.W. Oxalobacter sp. reduces urinary oxalate excretion by promoting enteric oxalate secretion. Kidney Int. 2006, 69, 691–698. [Google Scholar] [CrossRef]
- Caspary, W.F.; Tönissen, J.; Lankisch, P.G. “Enteral” hyperoxaluria: Effect of cholestyramine, calcium, neomycin, and bile acids on intestinal oxalate absorption in man. Acta Hepatogastroenterol (Stuttg) 1977, 24, 193–200. [Google Scholar]
- Hanes, D.A.; Weaver, C.M.; Heaney, R.P.; Wastney, M. Absorption of calcium oxalate does not require dissociation in rats. J. Nutr. 1999, 129, 170–173. [Google Scholar]
- Hatch, M.; Gjymishka, A.; Salido, E.C.; Allison, M.J.; Freel, R.W. Enteric oxalate elimination is induced and oxalate is normalized in a mouse model of primary hyperoxaluria following intestinal colonization with Oxalobacter. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 300, 461–469. [Google Scholar] [CrossRef]
- Hatch, M.; Freel, R.W. The roles and mechanisms of intestinal oxalate transport in oxalate homeostasis. Semin. Nephrol. 2008, 28, 143–151. [Google Scholar] [CrossRef]
- Königsberger, E.; Königsberger, L.C. Thermodynamic modeling of crystal deposition in humans. Pure Appl. Chem. 2001, 73, 785–797. [Google Scholar] [CrossRef]
- Allison, M.J.; Littledike, E.T.; James, L.F. Changes in ruminal oxalate degradation rates associated with adaptation to oxalate ingestion. J. Anim. Sci. 1977, 53, 1173–1179. [Google Scholar]
- Daniel, S.L.; Hartman, P.A.; Allison, M.J. Microbial degradation of oxalate in the gastrointestinal tract of rats. Appl. Environ. Microbiol. 1987, 53, 1793–1797. [Google Scholar]
- Allison, M.J.; Dawson, K.A.; Mayberry, W.R.; Foss, J.G. Oxalobacter formigene gen. nov., sp. nov.: Oxalate-degrading anaerobes that inhabit the gastrointestinal tract. Arch. Microbiol. 1985, 141, 1–7. [Google Scholar]
- Ito, H.; Miura, N.; Masai, M.; Yamamoto, K.; Hara, T. Reduction of oxalate content of foods by the oxalate degrading bacterium, Eubacterium lentum WYH-1. Int. J. Urol. 1996, 3, 31–34. [Google Scholar] [CrossRef]
- Hokama, S.; Honma, Y.; Toma, C.; Ogawa, Y. Oxalate-degrading Enterococcus faecalis. Microbiol. Immunol. 2000, 44, 235–240. [Google Scholar] [CrossRef]
- Campieri, C.; Campieri, M.; Bertuzzi, V.; Swennen, E.; Matteuzzi, D.; Stefoni, S.; Pirovano, F.; Centi, C.; Ulisse, S.; Famularo, G.; et al. Reduction of oxaluria after an oral course of lactic acid bacteria at high concentration. Kidney Int. 2001, 60, 1097–1105. [Google Scholar] [CrossRef]
- Miller, A.W.; Kohl, K.D.; Dearing, M.D. Microenvironments of the gut harbor distinct consortia of oxalate-degrading bacteria. Appl. Environ. Microbiol. 2013. submitted. [Google Scholar]
- Allison, M.J.; Cook, H.M.; Milne, D.B.; Gallagher, S.; Clayman, R.V. Oxalate degradation by gastrointestinal bacteria from humans. J. Nut. 1986, 116, 455–460. [Google Scholar]
- Kageyama, A.; Benno, Y.; Nakase, T. Phylogenetic evidence for the transfer of Eubacterium lentum to the genus Eggerthella as Eggerthella lenta gen. nov., comb. nov. Int. J. Syst. Evol. Microbiol. 1999, 49, 1725–1732. [Google Scholar] [CrossRef]
- Ren, Z.; Pan, C.; Jiang, L.; Wu, C.; Liu, Y.; Zhong, Z.; Ran, L.; Ren, F.; Chen, X.; Wang, Y.; Zhu, Y.; Huang, K. Oxalate-degrading capabilities of lactic acid bacteria in canine feces. Vet. Microbiol. 2011, 152, 368–373. [Google Scholar] [CrossRef]
- Hokama, S.; Toma, C.; Iwanaga, M.; Morozumi, M.; Sagaya, K.; Ogawa, Y. Oxalate-degrading Providencia rettgeri isolated from human stools. Int. J. Urol. 2005, 12, 533–538. [Google Scholar] [CrossRef]
- Kwak, C.; Jeong, B.C.; Ku, J.H.; Kim, H.H.; Lee, J.J.; Huh, C.S.; Baek, Y.J.; Lee, S.E. Prevention of nephrolithiasis by Lactobacillus in stone-forming rats: A preliminary study. Urol. Res. 2006, 34, 265–270. [Google Scholar] [CrossRef]
- Turroni, S.; Vitali, B.; Bendazzoli, C.; Candela, M.; Gotti, R.; Federici, F.; Pirovano, F.; Brigidi, P. Oxalate consumption by Lactobacilli: Evaluation of oxalyl-CoA decarboxylase and formyl-CoA transferase activity in Lactobacillus acidophilus. J. Appl. Microbiol. 2007, 103, 1600–1607. [Google Scholar] [CrossRef]
- Ferraz, R.R.N.; Marques, N.C.; Froeder, L.; Menon, V.B.; Siliano, P.R.; Baxmann, A.C.; Heilberg, I.P. Effects of Lactobacillus casei and Bifidobacterium breve on urinary oxalate excretion in nephrolithiasis patients. Urol. Res. 1999, 37, 95–100. [Google Scholar]
- Weese, J.S.; Weese, H.E.; Yuricek, L.; Rousseau, J. Oxalate degradation by intestinal lactic acid bacteria in dogs and cats. Vet. Microbiol. 2004, 101, 161–166. [Google Scholar] [CrossRef]
- Ruan, Z.S.; Anantharam, V.; Crawford, I.T.; Ambudkar, S.V.; Rhee, S.Y.; Allison, M.J.; Maloney, P.C. Identification, purification, and reconstitution of OxlT, the oxalate: formate antiporter protein of Oxalobacter formigenes. J. Biol. Chem. 1992, 267, 10537–10543. [Google Scholar]
- Baetz, A.L.; Allison, M.J. Purification and characterization of oxalyl-coenzyme A decarboxylase from Oxalobacter formigenes. J. Bacteriol. 1989, 171, 2605–2608. [Google Scholar]
- Baetz, A.L.; Allison, M.J. Purification and characterization of formyl-coenzyme A transferase from Oxalobacter formigenes. J. Bacteriol. 1990, 172, 3537–3540. [Google Scholar]
- Lung, H.Y.; Cornelius, J.; Peck, A.B. Cloning and expression of the oxalyl-CoA decarboxylase gene from the bacterium, Oxalobacter formigenes: Prospects for gene therapy to control Ca-oxalate kidney stone formation. Am. J. Kidney Dis. 1991, 17, 381–385. [Google Scholar]
- Sidhu, H.; Enatska, L.; Ogden, S.; Williams, W.N.; Allison, M.J.; Peck, A.B. Evaluating children in the Ukraine for colonization with the intestinal bacterium Oxalobacter formigenes, using a polymerase chain reactor-based detection system. Mol. Diagn. 1997, 2, 89–97. [Google Scholar] [CrossRef]
- Turroni, S.; Bendazzoli, C.; Dipalo, S.C.F.; Candela, M.; Vitali, B.; Gotti, R.; Brigidi, P. Oxalate-degrading activity in Bifidobacterium animalis subsp. lactis: Impact of acidic conditions on the transcriptional levels of the oxalyl-CoA decarboxylase and formyl-CoA transferase genes. Appl. Environ. Microbiol. 2010, 76, 5609–5620. [Google Scholar]
- Ley, R.E.; Lozupone, C.; Hamady, M.; Knight, R.; Gordon, J.I. Worlds within worlds: Evolution of the vertebrate gut microbiota. Nat. Rev. Microbiol. 2008, 6, 776–788. [Google Scholar] [CrossRef]
- Karasov, W.H.; Martinez del Rio, C. Physiological Ecology; Princeton University Press: Princeton, NJ, USA, 2007. [Google Scholar]
- Hook, S.E.; Steele, M.A.; Northwood, K.S.; Dijkstra, J.; France, J.; Wright, A.D.G.; McBride, B.W. Impact of subacute ruminal acidosis adaptation and recovery on the density and diversity of bacteria in the rumen of cows. FEMS Microbiol. Ecol. 2011, 78, 275–284. [Google Scholar] [CrossRef]
- Ley, R.E.; Peterson, D.A.; Gordon, J.I. Ecological and evolutionary forces shaping microbial diversity in the human intestines. Cell 2006, 124, 837–848. [Google Scholar] [CrossRef]
- Whitman, W.B.; Coleman, D.C.; Wiebe, W.J. Prokaryotes: The unseen majority. Proc. Natl. Acad. Sci. USA 1998, 95, 6578–6583. [Google Scholar] [CrossRef]
- Azcarate-Peril, M.A.; Bruno-Bárcena, J.M.; Hassan, H.M.; Klaenhammer, T.R. Transcriptional and functional analysis of oxalyl-coenzyme A (CoA) decarboxylase and formyl-coA transferase genes from Lactobacillus acidophilus. Appl. Environ. Microbiol. 2006, 72, 1891–1899. [Google Scholar] [CrossRef]
- Kuhner, C.H.; Hartman, P.A.; Allison, M.J. Generation of a proton motive force by the anaerobic oxalate-degrading Oxalobacter formigenes. Appl. Environ. Microbiol. 1996, 62, 2494–2500. [Google Scholar]
- Shirley, E.K.; Schmidt-Nielsen, K. Oxalate metabolism in the pack rat, sand rat, hamster, and white rat. J. Nutr. 1967, 91, 496–502. [Google Scholar]
- Knight, J.; Deora, R.; Assimos, D.G.; Holmes, R.P. The genetic composition of Oxalobacter formigenes and its relationship to colonization and calcium oxalate stone disease. Urol. Res. 2013, 41, 187–196. [Google Scholar]
- Belenguer, A.; Ben Bati, M.; Hervás, G.; Toral, P.G.; Yáñez-Ruiz, D.R.; Frutos, P. Impact of oxalic acid on rumen function and bacterial community in sheep. Animal 2013, 7, 940–947. [Google Scholar] [CrossRef] [Green Version]
- Allison, M.J.; Cook, H.M. Oxalate degradation by microbes of the large bowel of herbivores: The effect of dietary oxalate. Science 1981, 212, 675–676. [Google Scholar]
- Duncan, A.J.; Frutos, P.; Young, S.A. Rates of oxalate degradation in the rumen of sheep and goats in response to different levels of oxalic acid administration. Anim. Sci. 1997, 65, 451–455. [Google Scholar] [CrossRef]
- Nicholson, J.K.; Holmes, E.; Wilson, I.D. Gut microorganisms, mammalian metabolism, and personalized health care. Nat. Rev. Microbiol. 2005, 3, 431–438. [Google Scholar] [CrossRef]
- Lieske, J.C.; Tremaine, W.J.; de Simone, C.; O’Connor, H.M.; Li, X.; Bergstralh, E.J.; Goldfarb, D.S. Diet, but not oral probiotics, effectively reduces urinary oxalate excretion and calcium oxalate supersaturation. Kidney Int. 2010, 78, 1178–1185. [Google Scholar] [CrossRef]
- Duncan, S.H.; Richardson, A.J.; Kaul, P.; Homes, R.P.; Allison, M.J.; Stewart, C.S. Oxalobacter formigenes and its potential role in human health. Appl. Environ. Microbiol. 2002, 68, 3841–3847. [Google Scholar] [CrossRef]
- Kelly, J.P.; Curhan, G.C.; Cave, D.R.; Anderson, T.E.; Kaufman, D.W. Factors related to colonization with Oxalobacter formigenes in U.S. adults. J. Endourol. 2011, 25, 673–679. [Google Scholar] [CrossRef]
- Kharlamb, V.; Schelker, J.; Francois, F.; Jiang, J.; Holmes, R.P.; Goldfarb, D.S. Oral antibiotic treatment of Helicobacter pylori leads to persistently reduced intestinal colonization rates with Oxalobacter formigenes. J. Endourol. 2011, 25, 1781–1785. [Google Scholar] [CrossRef]
- Lange, J.N.; Wood, K.D.; Wong, H.; Otto, R.; Mufarrij, P.W.; Knight, J.; Akpinar, H.; Holmes, R.P.; Assimos, D.G. Sensitivity of human strains of Oxalobacter formigenes to commonly prescribed antibiotics. Urology 2012, 79, 1286–1289. [Google Scholar] [CrossRef]
- Sidhu, H.; Hoppe, B.; Hesse, A.; Tenbrock, K.; Bromme, S.; Rietschel, E.; Peck, A.B. Absence of Oxalobacter formigenes in cystic fibrosis patients: A risk factor for hyperoxaluria. Lancet 1998, 352, 1026–1029. [Google Scholar] [CrossRef]
- Lieske, J.C.; Goldfarb, D.S.; de Simone, C.; Regnier, C. Use of a probiotic to decrease enteric hyperoxaluria. Kidney Int. 2005, 68, 1244–1249. [Google Scholar] [CrossRef]
- Palgi, N.; Taliesnik, H.; Pinshow, B. Elimination of oxalate by fat sand rats (Psammomys obesus): Wild and laboratory-bred animals compared. Comp. Biochem. Phys. A 2008, 149, 197–202. [Google Scholar] [CrossRef]
- Borody, T.J.; Eloise, F.; Warren, S.M.; Leis, R.S.; Ori, A.; Siarakas, S. Bacteriotherapy using fecal flora: Toying with human motions. J. Clin. Gastroenterol. 2004, 38, 475–483. [Google Scholar]
- Huebner, E.; Surawicz, C.M. Probiotics in the prevention and treatment of gastrointestinal infections. Gastroenterol. Clin. North Am. 2006, 35, 355–365. [Google Scholar] [CrossRef]
- Gough, E.; Shaikh, H.; Manges, A.R. Systematic review of intestinal microbiota transplantation (fecal bacteriotherapy) for recurrent Clostridium difficile infection. Clin. Infect. Dis. 2011, 53, 994–1002. [Google Scholar] [CrossRef]
- Reid, G.; Younes, J.A.; van der Mei, H.C.; Gloor, G.B.; Knight, R.; Busscher, H.J. Microbiota restoration: Natural and supplemented recovery of human microbial communities. Nat. Rev. Microbiol. 2011, 9, 27–38. [Google Scholar] [CrossRef]
- Rawls, J.F.; Mahowald, M.A.; Ley, R.E.; Gordon, J.I. Reciprocal gut microbiota transplants from zebrafish and mice to germ-free recipients reveal host habitat selection. Cell 2006, 127, 423–433. [Google Scholar] [CrossRef]
- Pang, X.; Hua, X.; Yang, Q.; Ding, D.; Che, C.; Cui, L.; Jia, W.; Buchell, P.; Zhao, L. Inter-species transplantation of gut microbiota from humans to pigs. ISME J. 2007, 1, 156–162. [Google Scholar] [CrossRef]
- Hooper, L.V.; Midtvedt, T.; Gordon, J.I. How host-microbial interactions shape the nutrient environment of the mammalian intestine. Ann. Rev. Nutr. 2002, 22, 283–307. [Google Scholar] [CrossRef]
- Smillie, C.S.; Smith, M.B.; Friedman, J.; Cordero, O.T.; David, L.A.; Alm, E.J. Ecology drives a global network of gene exchange connecting the human microbiome. Nature 2011, 480, 241–244. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Miller, A.W.; Dearing, D. The Metabolic and Ecological Interactions of Oxalate-Degrading Bacteria in the Mammalian Gut. Pathogens 2013, 2, 636-652. https://doi.org/10.3390/pathogens2040636
Miller AW, Dearing D. The Metabolic and Ecological Interactions of Oxalate-Degrading Bacteria in the Mammalian Gut. Pathogens. 2013; 2(4):636-652. https://doi.org/10.3390/pathogens2040636
Chicago/Turabian StyleMiller, Aaron W., and Denise Dearing. 2013. "The Metabolic and Ecological Interactions of Oxalate-Degrading Bacteria in the Mammalian Gut" Pathogens 2, no. 4: 636-652. https://doi.org/10.3390/pathogens2040636