Biomolecular Mechanisms of Pseudomonas aeruginosa and Escherichia coli Biofilm Formation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Adhesion in the Gram-Negative Bacteria Pseudomonas aeruginosa and Escherichia coli
2.1. Pseudomonas aeruginosa Adhesion and Accumulation
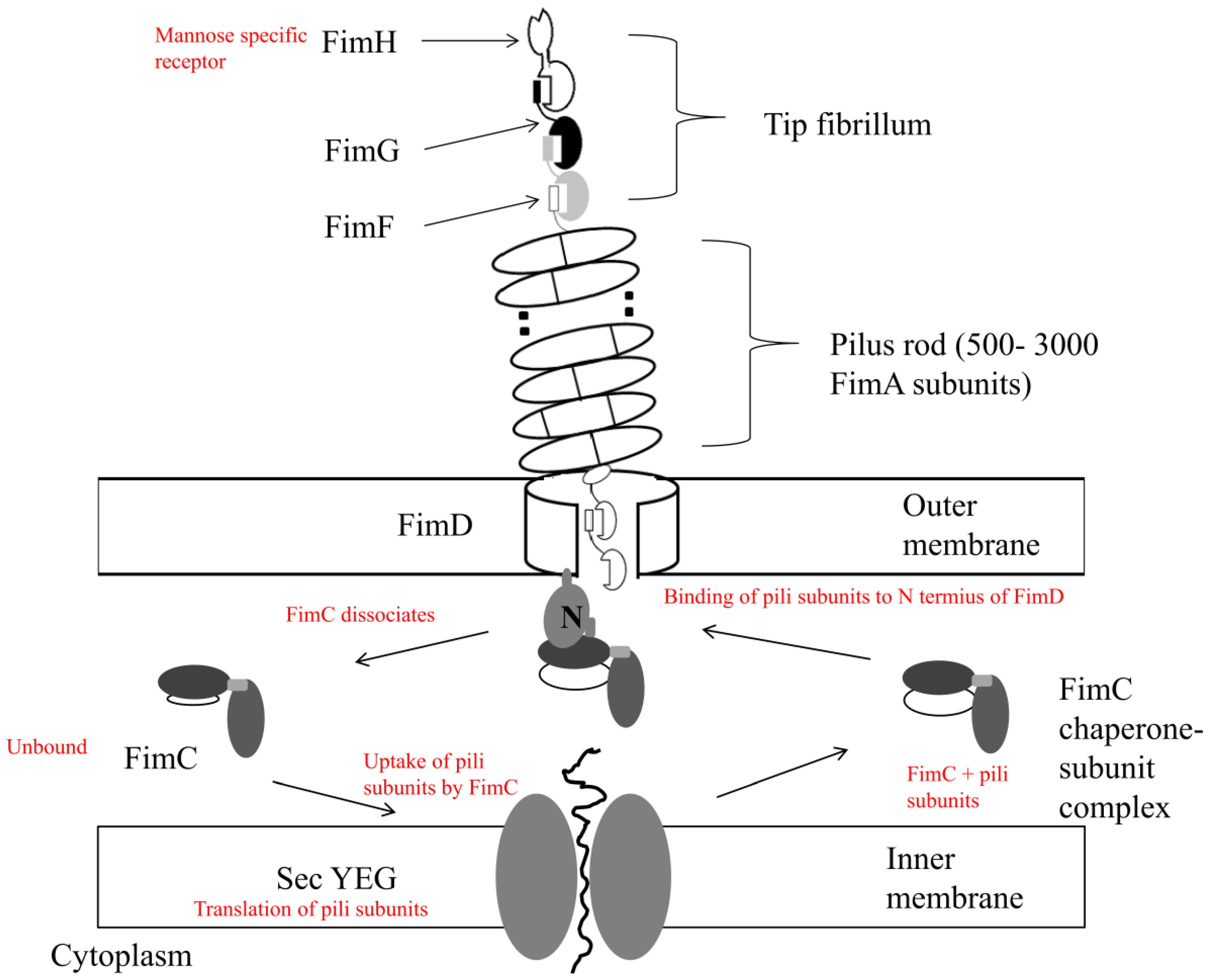
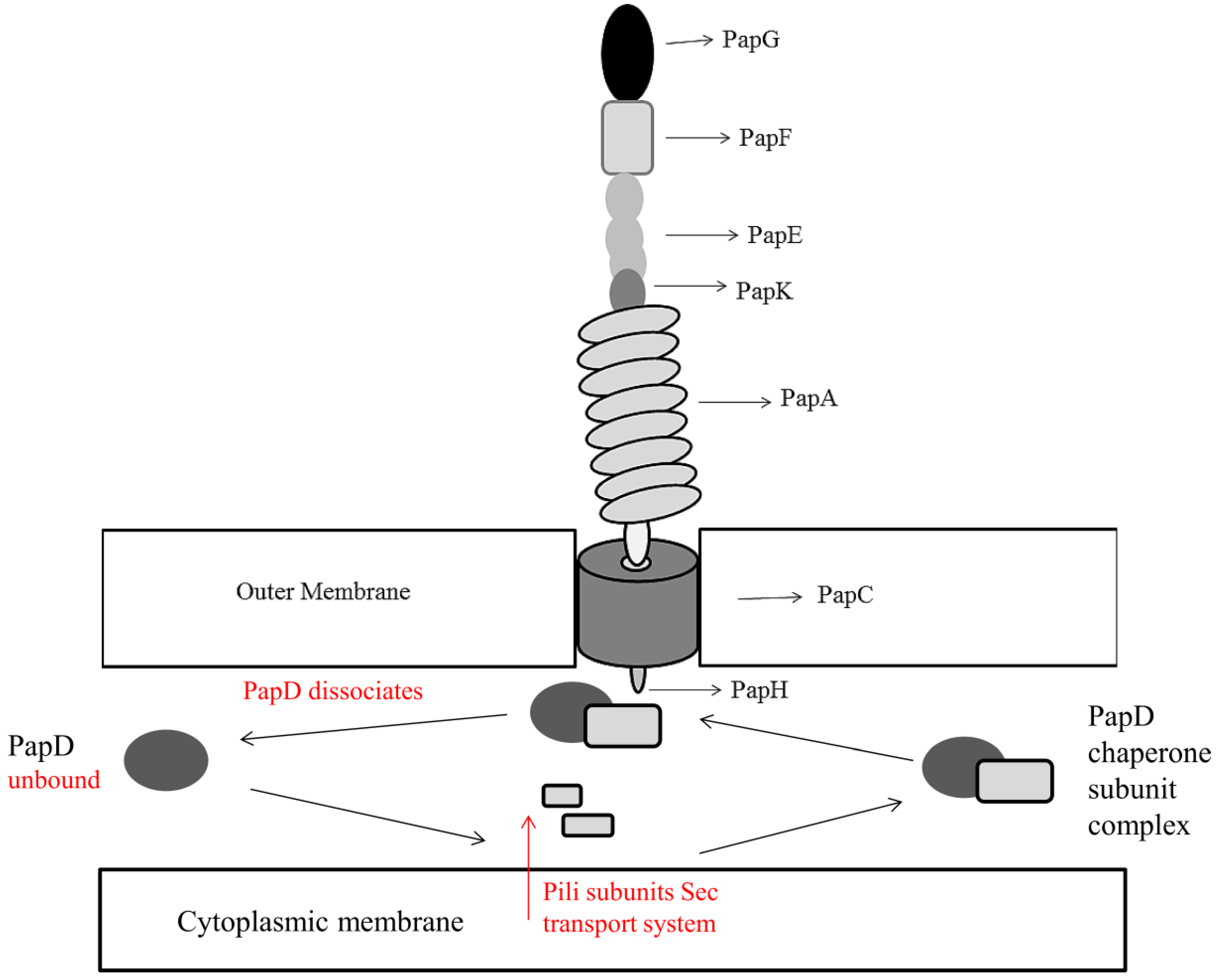
2.2. Escherichia Coli Adhesion and Accumulation


3. Biofilm Maturation in Pseudomonas Aeruginosa and Escherichia Coli
3.1. Pseudomonas aeruginosa Biofilm Maturation: Production of Exopolysaccharides via the Polysaccharide Synthesis Locus (Psl), Pellicle Formation (Pel), and Alginic Acid Synthesis
3.1.1. Production of the Psl and Pel Exopolysaccharides by Non-Mucoid Pseudomonas aeruginosa
3.1.2. Production of the Exopolysaccharide Alginic Acid by Mucoid Pseudomonas aeruginosa

3.2. Escherichia coli Biofilm Maturation
4. Quorum Sensing in Gram-Negative Bacteria: Pseudomonas aeruginosa and Escherichia coli
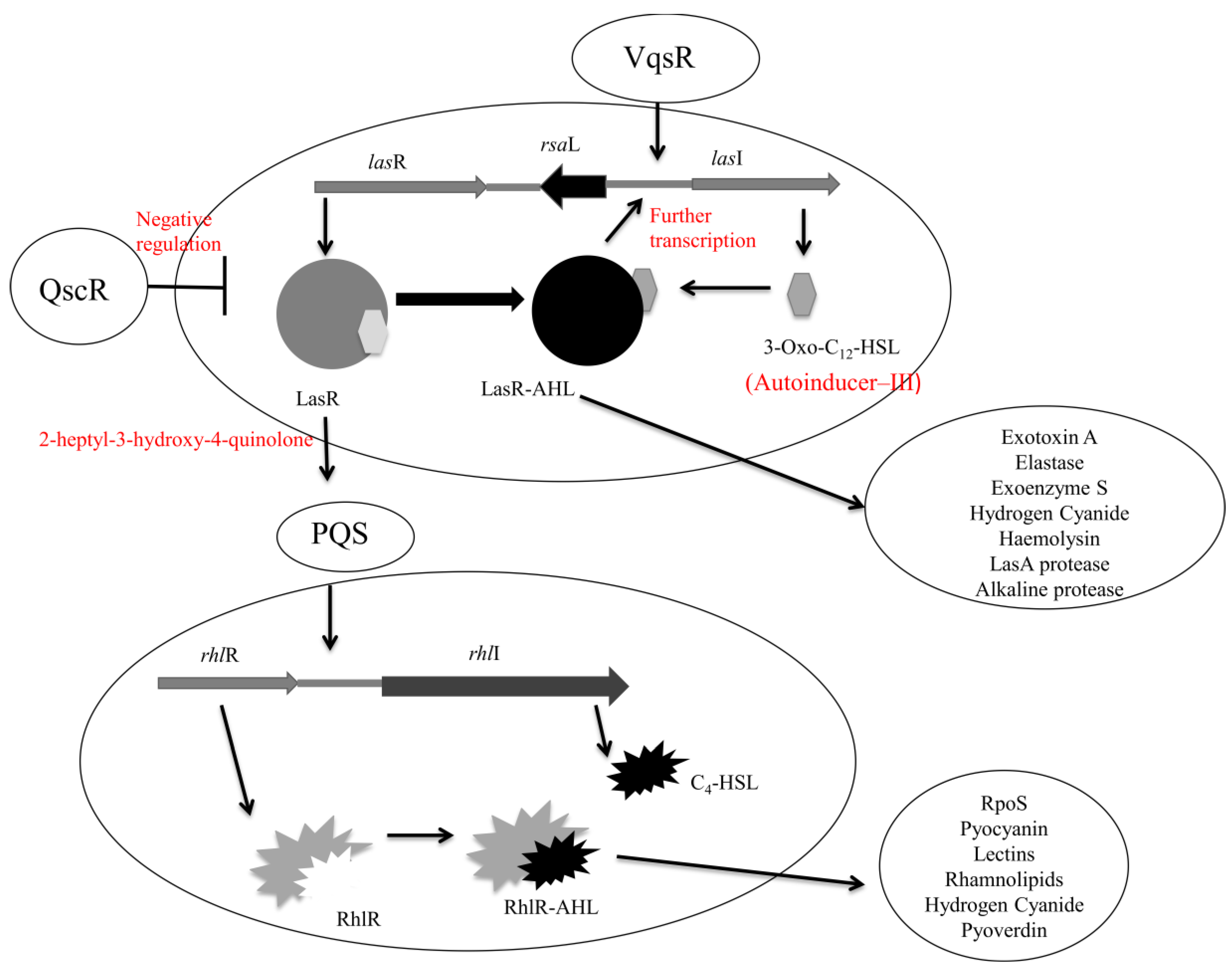
4.1. Quorum Sensing in Pseudomonas aeruginosa

4.2. RpoS and Quorum Sensing
4.3. Quorum Sensing in Escherichia coli
4.3.1. sdiA Quorum Sensing System
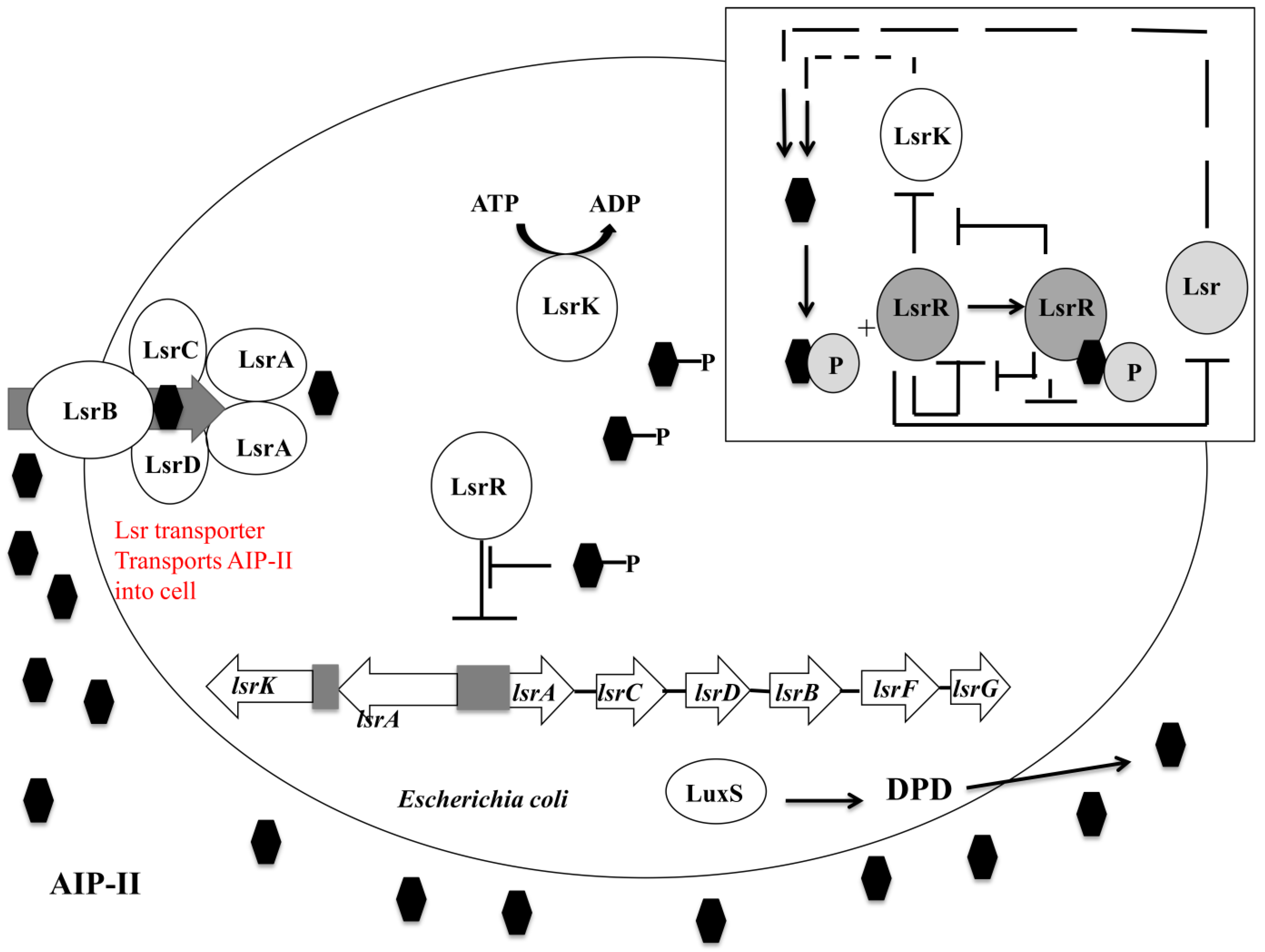
4.3.2. luxS Quorum Sensing System
 DPD: 4,5-dihydroxy-2,3-pentanedione. Adapted from Li, 2007 [184].
DPD: 4,5-dihydroxy-2,3-pentanedione. Adapted from Li, 2007 [184].
DPD: 4,5-dihydroxy-2,3-pentanedione. Adapted from Li, 2007 [184].
DPD: 4,5-dihydroxy-2,3-pentanedione. Adapted from Li, 2007 [184].
4.3.3. Autoinducer-III/Epinephrine/Norepinephrine and Indole Quorum Sensing Systems
5. Conclusions
Author Contributions
Conflicts of Interest
References
- Christensen, L.D.; Moser, C.; Jensen, P.O.; Rasmussen, T.B.; Christophersen, L.; Kjelleberg, S.; Kumar, N.; Hoiby, N.; Givskov, M.; Bjarnsholt, T. Impact of Pseudomonas aeruginosa Quorum Sensing on Biofilm Persistence in an in Vivo Intraperitoneal Foreign-Body Infection Model. Microbiology 2007, 153, 2312–2320. [Google Scholar] [CrossRef]
- Cole, S.J.; Records, A.R.; Orr, M.W.; Linden, S.B.; Lee, V.T. Catheter-Associated Urinary Tract Infection by Pseudomonas aeruginosa is Mediated by Exopolysaccharide-Independent Biofilms. Infect. Immun. 2014, 82, 2048–2058. [Google Scholar] [CrossRef]
- Guggenbichler, J.P.; Assadian, O.; Boeswald, M.; Kramer, A. Incidence and Clinical Implication of Nosocomial Infections Associated with Implantable Biomaterials—Catheters, Ventilator-Associated Pneumonia, Urinary Tract Infections. GMS Krankenhhyg. Interdiszip. 2011, 6, 1–19. [Google Scholar]
- Ramos, G.P.; Rocha, J.L.; Tuon, F.F. Seasonal Humidity may Influence Pseudomonas aeruginosa Hospital-Acquired Infection Rates. Int. J. Infect. Dis. 2013, 17, e757–e761. [Google Scholar] [CrossRef]
- Hoiby, N.; Ciofu, O.; Bjarnsholt, T. Pseudomonas aeruginosa Biofilms in Cystic Fibrosis. Future Microbiol. 2010, 5, 1663–1674. [Google Scholar] [CrossRef]
- Jacobsen, S.M.; Stickler, D.J.; Mobley, H.L.; Shirtliff, M.E. Complicated Catheter-Associated Urinary Tract Infections due to Escherichia coli and Proteus mirabilis. Clin. Microbiol. Rev. 2008, 21, 26–59. [Google Scholar] [CrossRef]
- Foxman, B.; Brown, P. Epidemiology of Urinary Tract Infections: Transmission and Risk Factors, Incidence, and Costs. Infect. Dis. Clin. N. Am. 2003, 17, 227–241. [Google Scholar] [CrossRef]
- Laverty, G.; Gorman, S.P.; Gilmore, B.F. Biomolecular Mechanisms of Staphylococcal Biofilm Formation. Future Microbiol. 2013, 8, 509–524. [Google Scholar] [CrossRef] [Green Version]
- Sutherland, I.W. Microbial Polysaccharides from Gram-negative Bacteria. Int. Dairy J. 2001, 11, 663–674. [Google Scholar] [CrossRef]
- Lejeune, P. Contamination of Abiotic Surfaces: What a Colonizing Bacterium Sees and how to Blur it. Trends Microbiol. 2003, 11, 179–184. [Google Scholar] [CrossRef]
- O’Toole, G.A.; Kolter, R. Flagellar and Twitching Motility are Necessary for Pseudomonas aeruginosa Biofilm Development. Mol. Microbiol. 1998, 30, 295–304. [Google Scholar] [CrossRef]
- Dunne, W.M., Jr. Bacterial Adhesion: Seen any Good Biofilms Lately? Clin. Microbiol. Rev. 2002, 15, 155–166. [Google Scholar] [CrossRef]
- Bohn, Y.S.; Brandes, G.; Rakhimova, E.; Horatzek, S.; Salunkhe, P.; Munder, A.; van Barneveld, A.; Jordan, D.; Bredenbruch, F.; Haussler, S.; et al. Multiple Roles of Pseudomonas aeruginosa TBCF10839 PilY1 in Motility, Transport and Infection. Mol. Microbiol. 2009, 71, 730–747. [Google Scholar] [CrossRef]
- Strom, M.S.; Nunn, D.N.; Lory, S. Posttranslational Processing of Type IV Prepilin and Homologs by PilD of Pseudomonas aeruginosa. Methods Enzymol. 1994, 235, 527–540. [Google Scholar] [CrossRef]
- Mattick, J.S. Type IV Pili and Twitching Motility. Ann. Rev. Microbiol. 2002, 56, 289–314. [Google Scholar] [CrossRef]
- Nunn, D.; Bergman, S.; Lory, S. Products of Three Accessory Genes, pilB, pilC, and pilD, are Required for Biogenesis of Pseudomonas aeruginosa Pili. J. Bacteriol. 1990, 172, 2911–2919. [Google Scholar]
- Alm, R.A.; Hallinan, J.P.; Watson, A.A.; Mattick, J.S. Fimbrial Biogenesis Genes of Pseudomonas aeruginosa: pilW and pilX Increase the Similarity of Type 4 Fimbriae to the GSP Protein-Secretion Systems and pilY1 Encodes a Gonococcal PilC Homologue. Mol. Microbiol. 1996, 22, 161–173. [Google Scholar] [CrossRef]
- Murray, T.S.; Kazmierczak, B.I. Pseudomonas aeruginosa Exhibits Sliding Motility in the Absence of Type IV Pili and Flagella. J. Bacteriol. 2008, 190, 2700–2708. [Google Scholar] [CrossRef]
- Makin, S.A.; Beveridge, T.J. The Influence of A-Band and B-Band Lipopolysaccharide on the Surface Characteristics and Adhesion of Pseudomonas aeruginosa to Surfaces. Microbiology 1996, 142, 299–307. [Google Scholar] [CrossRef]
- Darzins, A.; Russell, M.A. Molecular Genetic Analysis of Type-4 Pilus Biogenesis and Twitching Motility using Pseudomonas aeruginosa as a Model System-a Review. Gene 1997, 192, 109–115. [Google Scholar] [CrossRef]
- Barken, K.B.; Pamp, S.J.; Yang, L.; Gjermansen, M.; Bertrand, J.J.; Klausen, M.; Givskov, M.; Whitchurch, C.B.; Engel, J.N.; Tolker-Nielsen, T. Roles of Type IV Pili, Flagellum-Mediated Motility and Extracellular DNA in the Formation of Mature Multicellular Structures in Pseudomonas aeruginosa Biofilms. Environ. Microbiol. 2008, 10, 2331–2343. [Google Scholar] [CrossRef]
- Wentworth, J.S.; Austin, F.E.; Garber, N.; Gilboa-Garber, N.; Paterson, C.A.; Doyle, R.J. Cytoplasmic Lectins Contribute to the Adhesion of Pseudomonas aeruginosa. Biofouling 1991, 4, 99–104. [Google Scholar] [CrossRef]
- Tielker, D.; Hacker, S.; Loris, R.; Strathmann, M.; Wingender, J.; Wilhelm, S.; Rosenau, F.; Jaeger, K.E. Pseudomonas aeruginosa Lectin LecB is Located in the Outer Membrane and is Involved in Biofilm Formation. Microbiology 2005, 151, 1313–1323. [Google Scholar] [CrossRef]
- Adam, J.; Pokorna, M.; Sabin, C.; Mitchell, E.P.; Imberty, A.; Wimmerova, M. Engineering of PA-IIL Lectin from Pseudomonas aeruginosa—Unravelling the Role of the Specificity Loop for Sugar Preference. BMC Struct. Biol. 2007, 7, 36. [Google Scholar] [CrossRef]
- Mewe, M.; Tielker, D.; Schonberg, R.; Schachner, M.; Jaeger, K.E.; Schumacher, U. Pseudomonas aeruginosa Lectins I and II and their Interaction with Human Airway Cilia. J. Laryngol. Otol. 2005, 119, 595–599. [Google Scholar]
- Hauber, H.P.; Schulz, M.; Pforte, A.; Mack, D.; Zabel, P.; Schumacher, U. Inhalation with Fucose and Galactose for Treatment of Pseudomonas aeruginosa in Cystic Fibrosis Patients. Int. J. Med. Sci. 2008, 5, 371–376. [Google Scholar]
- Kolomiets, E.; Swiderska, M.A.; Kadam, R.U.; Johansson, E.M.; Jaeger, K.E.; Darbre, T.; Reymond, J.L. Glycopeptide Dendrimers with High Affinity for the Fucose-Binding Lectin LecB from Pseudomonas aeruginosa. ChemMedChem 2009, 4, 562–569. [Google Scholar] [CrossRef]
- Winzer, K.; Falconer, C.; Garber, N.C.; Diggle, S.P.; Camara, M.; Williams, P. The Pseudomonas aeruginosa Lectins PA-IL and PA-IIL are Controlled by Quorum Sensing and by RpoS. J. Bacteriol. 2000, 182, 6401–6411. [Google Scholar] [CrossRef]
- Hull, R.A.; Gill, R.E.; Hsu, P.; Minshew, B.H.; Falkow, S. Construction and Expression of Recombinant Plasmids Encoding Type 1 Or D-Mannose-Resistant Pili from a Urinary Tract Infection Escherichia coli Isolate. Infect. Immun. 1981, 33, 933–938. [Google Scholar]
- Baorto, D.M.; Gao, Z.; Malaviya, R.; Dustin, M.L.; van der Merwe, A.; Lublin, D.M.; Abraham, S.N. Survival of FimH-Expressing Enterobacteria in Macrophages Relies on Glycolipid Traffic. Nature 1997, 389, 636–639. [Google Scholar] [CrossRef]
- Connell, I.; Agace, W.; Klemm, P.; Schembri, M.; Marild, S.; Svanborg, C. Type 1 Fimbrial Expression Enhances Escherichia coli Virulence for the Urinary Tract. Proc. Natl. Acad. Sci. USA 1996, 93, 9827–9832. [Google Scholar] [CrossRef]
- Mobley, H.L.; Chippendale, G.R.; Tenney, J.H.; Hull, R.A.; Warren, J.W. Expression of Type 1 Fimbriae may be Required for Persistence of Escherichia coli in the Catheterized Urinary Tract. J. Clin. Microbiol. 1987, 25, 2253–2257. [Google Scholar]
- Cormio, L.; Vuopio-Varkila, J.; Siitonen, A.; Talja, M.; Ruutu, M. Bacterial Adhesion and Biofilm Formation on various Double-J Stents in Vivo and in Vitro. Scand. J. Urol. Nephrol. 1996, 30, 19–24. [Google Scholar] [CrossRef]
- Vetsch, M.; Puorger, C.; Spirig, T.; Grauschopf, U.; Weber-Ban, E.U.; Glockshuber, R. Pilus Chaperones Represent a New Type of Protein-Folding Catalyst. Nature 2004, 431, 329–333. [Google Scholar] [CrossRef]
- Sauer, F.G.; Barnhart, M.; Choudhury, D.; Knight, S.D.; Waksman, G.; Hultgren, S.J. Chaperone-Assisted Pilus Assembly and Bacterial Attachment. Curr. Opin. Struct. Biol. 2000, 10, 548–556. [Google Scholar] [CrossRef]
- Gossert, A.D.; Bettendorff, P.; Puorger, C.; Vetsch, M.; Herrmann, T.; Glockshuber, R.; Wuthrich, K. NMR Structure of the Escherichia coli Type 1 Pilus Subunit FimF and its Interactions with Other Pilus Subunits. J. Mol. Biol. 2008, 375, 752–763. [Google Scholar] [CrossRef]
- Capitani, G.; Eidam, O.; Glockshuber, R.; Grutter, M.G. Structural and Functional Insights into the Assembly of Type 1 Pili from Escherichia coli. Microbes Infect. 2006, 8, 2284–2290. [Google Scholar] [CrossRef]
- Lugmaier, R.A.; Schedin, S.; Kuhner, F.; Benoit, M. Dynamic Restacking of Escherichia coli P-Pili. Eur. Biophys. J. 2008, 37, 111–120. [Google Scholar] [CrossRef]
- Jacob-Dubuisson, F.; Heuser, J.; Dodson, K.; Normark, S.; Hultgren, S. Initiation of Assembly and Association of the Structural Elements of a Bacterial Pilus Depend on Two Specialized Tip Proteins. EMBO J. 1993, 12, 837–847. [Google Scholar]
- Baga, M.; Norgren, M.; Normark, S. Biogenesis of E. coli Pap Pili: PapH, a Minor Pilin Subunit Involved in Cell Anchoring and Length Modulation. Cell 1987, 49, 241–251. [Google Scholar] [CrossRef]
- Jones, C.H.; Danese, P.N.; Pinkner, J.S.; Silhavy, T.J.; Hultgren, S.J. The Chaperone-Assisted Membrane Release and Folding Pathway is Sensed by Two Signal Transduction Systems. EMBO J. 1997, 16, 6394–6406. [Google Scholar] [CrossRef]
- Thanassi, D.G.; Saulino, E.T.; Lombardo, M.J.; Roth, R.; Heuser, J.; Hultgren, S.J. The PapC Usher Forms an Oligomeric Channel: Implications for Pilus Biogenesis Across the Outer Membrane. Proc. Natl. Acad. Sci. USA 1998, 95, 3146–3151. [Google Scholar]
- Anantha, R.P.; Stone, K.D.; Donnenberg, M.S. Effects of bfp Mutations on Biogenesis of Functional Enteropathogenic Escherichia coli Type IV Pili. J. Bacteriol. 2000, 182, 2498–2506. [Google Scholar] [CrossRef]
- Strom, M.S.; Lory, S. Structure-Function and Biogenesis of the Type IV Pili. Ann. Rev. Microbiol. 1993, 47, 565–596. [Google Scholar] [CrossRef]
- Ramer, S.W.; Schoolnik, G.K.; Wu, C.Y.; Hwang, J.; Schmidt, S.A.; Bieber, D. The Type IV Pilus Assembly Complex: Biogenic Interactions among the Bundle-Forming Pilus Proteins of Enteropathogenic Escherichia coli. J. Bacteriol. 2002, 184, 3457–3465. [Google Scholar] [CrossRef]
- Sauer, F.G.; Knight, S.D.; Waksman and, G.J.; Hultgren, S.J. PapD-Like Chaperones and Pilus Biogenesis. Semin. Cell Dev. Biol. 2000, 11, 27–34. [Google Scholar] [CrossRef]
- Verger, D.; Miller, E.; Remaut, H.; Waksman, G.; Hultgren, S. Molecular Mechanism of P Pilus Termination in Uropathogenic Escherichia coli. EMBO Rep. 2006, 7, 1228–1232. [Google Scholar] [CrossRef]
- Mu, X.Q.; Bullitt, E. Structure and Assembly of P-Pili: A Protruding Hinge Region used for Assembly of a Bacterial Adhesion Filament. Proc. Natl. Acad. Sci. USA 2006, 103, 9861–9866. [Google Scholar] [CrossRef]
- Mu, X.Q.; Jiang, Z.G.; Bullitt, E. Localization of a Critical Interface for Helical Rod Formation of Bacterial Adhesion P-Pili. J. Mol. Biol. 2005, 346, 13–20. [Google Scholar] [CrossRef]
- Olsen, A.; Arnqvist, A.; Hammar, M.; Sukupolvi, S.; Normark, S. The RpoS Sigma Factor Relieves H-NS-Mediated Transcriptional Repression of csgA, the Subunit Gene of Fibronectin-Binding Curli in Escherichia coli. Mol. Microbiol. 1993, 7, 523–536. [Google Scholar] [CrossRef]
- Arnqvist, A.; Olsen, A.; Normark, S. Sigma S-Dependent Growth-Phase Induction of the csgBA Promoter in Escherichia coli can be Achieved in Vivo by Sigma 70 in the Absence of the Nucleoid-Associated Protein H-NS. Mol. Microbiol. 1994, 13, 1021–1032. [Google Scholar] [CrossRef]
- Romling, U.; Rohde, M.; Olsen, A.; Normark, S.; Reinkoster, J. AgfD, the Checkpoint of Multicellular and Aggregative Behaviour in Salmonella typhimurium Regulates at Least Two Independent Pathways. Mol. Microbiol. 2000, 36, 10–23. [Google Scholar] [CrossRef]
- Brown, P.K.; Dozois, C.M.; Nickerson, C.A.; Zuppardo, A.; Terlonge, J.; Curtiss, R., 3rd. MlrA, a Novel Regulator of Curli (AgF) and Extracellular Matrix Synthesis by Escherichia coli and Salmonella enterica Serovar Typhimurium. Mol. Microbiol. 2001, 41, 349–363. [Google Scholar] [CrossRef]
- Gerstel, U.; Park, C.; Romling, U. Complex Regulation of csgD Promoter Activity by Global Regulatory Proteins. Mol. Microbiol. 2003, 49, 639–654. [Google Scholar] [CrossRef]
- Gerstel, U.; Romling, U. Oxygen Tension and Nutrient Starvation are Major Signals that Regulate agfD Promoter Activity and Expression of the Multicellular Morphotype in Salmonella typhimurium. Environ. Microbiol. 2001, 3, 638–648. [Google Scholar] [CrossRef]
- Olsen, A.; Arnqvist, A.; Hammar, M.; Normark, S. Environmental Regulation of Curli Production in Escherichia coli. Infect. Agents Dis. 1993, 2, 272–274. [Google Scholar]
- Romling, U.; Sierralta, W.D.; Eriksson, K.; Normark, S. Multicellular and Aggregative Behaviour of Salmonella typhimurium Strains is Controlled by Mutations in the agfD Promoter. Mol. Microbiol. 1998, 28, 249–264. [Google Scholar] [CrossRef]
- Dorel, C.; Vidal, O.; Prigent-Combaret, C.; Vallet, I.; Lejeune, P. Involvement of the Cpx Signal Transduction Pathway of E. coli in Biofilm Formation. FEMS Microbiol. Lett. 1999, 178, 169–175. [Google Scholar] [CrossRef]
- Cegelski, L.; Pinkner, J.S.; Hammer, N.D.; Cusumano, C.K.; Hung, C.S.; Chorell, E.; Aberg, V.; Walker, J.N.; Seed, P.C.; Almqvist, F.; et al. Small-Molecule Inhibitors Target Escherichia coli Amyloid Biogenesis and Biofilm Formation. Nat. Chem. Biol. 2009, 5, 913–919. [Google Scholar] [CrossRef]
- Dorel, C.; Lejeune, P.; Rodrigue, A. The Cpx System of Escherichia coli, a Strategic Signaling Pathway for Confronting Adverse Conditions and for Settling Biofilm Communities? Res. Microbiol. 2006, 157, 306–314. [Google Scholar] [CrossRef]
- Landini, P. Cross-Talk Mechanisms in Biofilm Formation and Responses to Environmental and Physiological Stress in Escherichia coli. Res. Microbiol. 2009, 160, 259–266. [Google Scholar] [CrossRef]
- Danese, P.N.; Oliver, G.R.; Barr, K.; Bowman, G.D.; Rick, P.D.; Silhavy, T.J. Accumulation of the Enterobacterial Common Antigen Lipid II Biosynthetic Intermediate Stimulates degP Transcription in Escherichia coli. J. Bacteriol. 1998, 180, 5875–5884. [Google Scholar]
- Prigent-Combaret, C.; Brombacher, E.; Vidal, O.; Ambert, A.; Lejeune, P.; Landini, P.; Dorel, C. Complex Regulatory Network Controls Initial Adhesion and Biofilm Formation in Escherichia coli Via Regulation of the csgD Gene. J. Bacteriol. 2001, 183, 7213–7223. [Google Scholar] [CrossRef]
- De Wulf, P.; Kwon, O.; Lin, E.C. The CpxRA Signal Transduction System of Escherichia coli: Growth-Related Autoactivation and Control of Unanticipated Target Operons. J. Bacteriol. 1999, 181, 6772–6778. [Google Scholar]
- Becker, G.; Klauck, E.; Hengge-Aronis, R. Regulation of RpoS Proteolysis in Escherichia coli: The Response Regulator RssB is a Recognition Factor that Interacts with the Turnover Element in RpoS. Proc. Natl. Acad. Sci. USA 1999, 96, 6439–6444. [Google Scholar] [CrossRef]
- Beloin, C.; Valle, J.; Latour-Lambert, P.; Faure, P.; Kzreminski, M.; Balestrino, D.; Haagensen, J.A.; Molin, S.; Prensier, G.; Arbeille, B.; Ghigo, J.M. Global Impact of Mature Biofilm Lifestyle on Escherichia coli K-12 Gene Expression. Mol. Microbiol. 2004, 51, 659–674. [Google Scholar]
- Hirakawa, H.; Inazumi, Y.; Masaki, T.; Hirata, T.; Yamaguchi, A. Indole Induces the Expression of Multidrug Exporter Genes in Escherichia coli. Mol. Microbiol. 2005, 55, 1113–1126. [Google Scholar]
- De Wulf, P.; McGuire, A.M.; Liu, X.; Lin, E.C. Genome-Wide Profiling of Promoter Recognition by the Two-Component Response Regulator CpxR-P in Escherichia coli. J. Biol. Chem. 2002, 277, 26652–26661. [Google Scholar] [CrossRef]
- Jubelin, G.; Vianney, A.; Beloin, C.; Ghigo, J.M.; Lazzaroni, J.C.; Lejeune, P.; Dorel, C. CpxR/OmpR Interplay Regulates Curli Gene Expression in Response to Osmolarity in Escherichia coli. J. Bacteriol. 2005, 187, 2038–2049. [Google Scholar] [CrossRef]
- Rampersaud, A.; Harlocker, S.L.; Inouye, M. The OmpR Protein of Escherichia coli Binds to Sites in the ompF Promoter Region in a Hierarchical Manner Determined by its Degree of Phosphorylation. J. Biol. Chem. 1994, 269, 12559–12566. [Google Scholar]
- Yamamoto, K.; Hirao, K.; Oshima, T.; Aiba, H.; Utsumi, R.; Ishihama, A. Functional Characterization in Vitro of all Two-Component Signal Transduction Systems from Escherichia coli. J. Biol. Chem. 2005, 280, 1448–1456. [Google Scholar]
- Ogasawara, H.; Yamada, K.; Kori, A.; Yamamoto, K.; Ishihama, A. Regulation of the Escherichia coli csgD Promoter: Interplay between Five Transcription Factors. Microbiology 2010, 156, 2470–2483. [Google Scholar] [CrossRef]
- Vidal, O.; Longin, R.; Prigent-Combaret, C.; Dorel, C.; Hooreman, M.; Lejeune, P. Isolation of an Escherichia coli K-12 Mutant Strain Able to Form Biofilms on Inert Surfaces: Involvement of a New ompR Allele that Increases Curli Expression. J. Bacteriol. 1998, 180, 2442–2449. [Google Scholar]
- Landini, P.; Zehnder, A.J. The Global Regulatory hns Gene Negatively Affects Adhesion to Solid Surfaces by Anaerobically Grown Escherichia coli by Modulating Expression of Flagellar Genes and Lipopolysaccharide Production. J. Bacteriol. 2002, 184, 1522–1529. [Google Scholar] [CrossRef]
- Evans, L.R.; Linker, A. Production and Characterization of the Slime Polysaccharide of Pseudomonas aeruginosa. J. Bacteriol. 1973, 116, 915–924. [Google Scholar]
- Chen, J.; Lee, S.M.; Mao, Y. Protective Effect of Exopolysaccharide Colanic Acid of Escherichia coli O157:H7 to Osmotic and Oxidative Stress. Int. J. Food Microbiol. 2004, 93, 281–286. [Google Scholar] [CrossRef]
- Zhao, K.; Liu, M.; Burgess, R.R. Adaptation in Bacterial Flagellar and Motility Systems: From Regulon Members to Foraging-Like Behavior in E. coli. Nucl. Acids Res. 2007, 35, 4441–4452. [Google Scholar] [CrossRef]
- Sperandio, V.; Torres, A.G.; Jarvis, B.; Nataro, J.P.; Kaper, J.B. Bacteria-Host Communication: The Language of Hormones. Proc. Natl. Acad. Sci. USA 2003, 100, 8951–8956. [Google Scholar]
- Winzer, K.; Williams, P. Escherichia coli Gets the Message. Nat. Med. 2003, 9, 1118–1119. [Google Scholar] [CrossRef]
- Vendeville, A.; Winzer, K.; Heurlier, K.; Tang, C.M.; Hardie, K.R. Making “Sense” of Metabolism: Autoinducer-2, LuxS and Pathogenic Bacteria. Nat. Rev. Microbiol. 2005, 3, 383–396. [Google Scholar] [CrossRef]
- Williams, P.; Winzer, K.; Chan, W.C.; Camara, M. Look Who’s Talking: Communication and Quorum Sensing in the Bacterial World. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2007, 362, 1119–1134. [Google Scholar] [CrossRef]
- Winzer, K.; Hardie, K.R.; Williams, P. Bacterial Cell-to-Cell Communication: Sorry, can’t Talk Now—Gone to Lunch! Curr. Opin. Microbiol. 2002, 5, 216–222. [Google Scholar] [CrossRef]
- Allesen-Holm, M.; Barken, K.B.; Yang, L.; Klausen, M.; Webb, J.S.; Kjelleberg, S.; Molin, S.; Givskov, M.; Tolker-Nielsen, T.A. Characterization of DNA Release in Pseudomonas aeruginosa Cultures and Biofilms. Mol. Microbiol. 2006, 59, 1114–1128. [Google Scholar] [CrossRef]
- Branda, S.S.; Vik, S.; Friedman, L.; Kolter, R. Biofilms: The Matrix Revisited. Trends Microbiol. 2005, 13, 20–26. [Google Scholar] [CrossRef]
- Matsukawa, M.; Greenberg, E.P. Putative Exopolysaccharide Synthesis Genes Influence Pseudomonas aeruginosa Biofilm Development. J. Bacteriol. 2004, 186, 4449–4456. [Google Scholar] [CrossRef]
- Deretic, V.; Govan, J.R.; Konyecsni, W.M.; Martin, D.W. Mucoid Pseudomonas aeruginosa in Cystic Fibrosis: Mutations in the muc Loci Affect Transcription of the algR and algD Genes in Response to Environmental Stimuli. Mol. Microbiol. 1990, 4, 189–196. [Google Scholar] [CrossRef]
- Stapper, A.P.; Narasimhan, G.; Ohman, D.E.; Barakat, J.; Hentzer, M.; Molin, S.; Kharazmi, A.; Hoiby, N.; Mathee, K. Alginate Production Affects Pseudomonas aeruginosa Biofilm Development and Architecture, but is Not Essential for Biofilm Formation. J. Med. Microbiol. 2004, 53, 679–690. [Google Scholar] [CrossRef]
- Doggett, R.G.; Harrison, G.M.; Stillwell, R.N.; Wallis, E.S. An Atypical Pseudomonas aeruginosa Associated with Cystic Fibrosis of the Pancreas. J. Pediatr. 1966, 68, 215–221. [Google Scholar] [CrossRef]
- Friedman, L.; Kolter, R. Two Genetic Loci Produce Distinct Carbohydrate-Rich Structural Components of the Pseudomonas aeruginosa Biofilm Matrix. J. Bacteriol. 2004, 186, 4457–4465. [Google Scholar] [CrossRef]
- Lee, V.T.; Matewish, J.M.; Kessler, J.L.; Hyodo, M.; Hayakawa, Y.; Lory, S. A Cyclic-Di-GMP Receptor Required for Bacterial Exopolysaccharide Production. Mol. Microbiol. 2007, 65, 1474–1484. [Google Scholar] [CrossRef]
- Byrd, M.S.; Sadovskaya, I.; Vinogradov, E.; Lu, H.; Sprinkle, A.B.; Richardson, S.H.; Ma, L.; Ralston, B.; Parsek, M.R.; Anderson, E.M.; et al. Genetic and Biochemical Analyses of the Pseudomonas aeruginosa Psl Exopolysaccharide Reveal Overlapping Roles for Polysaccharide Synthesis Enzymes in Psl and LPS Production. Mol. Microbiol. 2009, 73, 622–638. [Google Scholar] [CrossRef]
- Overhage, J.; Schemionek, M.; Webb, J.S.; Rehm, B.H. Expression of the psl Operon in Pseudomonas aeruginosa PAO1 Biofilms: PslA Performs an Essential Function in Biofilm Formation. Appl. Environ. Microbiol. 2005, 71, 4407–4413. [Google Scholar] [CrossRef]
- Wozniak, D.J.; Wyckoff, T.J.; Starkey, M.; Keyser, R.; Azadi, P.; O’Toole, G.A.; Parsek, M.R. Alginate is Not a Significant Component of the Extracellular Polysaccharide Matrix of PA14 and PAO1 Pseudomonas aeruginosa Biofilms. Proc. Natl. Acad. Sci. USA 2003, 100, 7907–7912. [Google Scholar]
- Friedman, L.; Kolter, R. Genes Involved in Matrix Formation in Pseudomonas aeruginosa PA14 Biofilms. Mol. Microbiol. 2004, 51, 675–690. [Google Scholar] [CrossRef]
- Stewart, P.S.; Franklin, M.J. Physiological Heterogeneity in Biofilms. Nat. Rev. Microbiol. 2008, 6, 199–210. [Google Scholar] [CrossRef]
- Ma, L.; Lu, H.; Sprinkle, A.; Parsek, M.R.; Wozniak, D.J. Pseudomonas aeruginosa Psl is a Galactose- and Mannose-Rich Exopolysaccharide. J. Bacteriol. 2007, 189, 8353–8356. [Google Scholar] [CrossRef]
- Rani, S.A.; Pitts, B.; Beyenal, H.; Veluchamy, R.A.; Lewandowski, Z.; Davison, W.M.; Buckingham-Meyer, K.; Stewart, P.S. Spatial Patterns of DNA Replication, Protein Synthesis, and Oxygen Concentration within Bacterial Biofilms Reveal Diverse Physiological States. J. Bacteriol. 2007, 189, 4223–4233. [Google Scholar] [CrossRef]
- DiGiandomenico, A.; Warrener, P.; Hamilton, M.; Guillard, S.; Ravn, P.; Minter, R.; Camara, M.M.; Venkatraman, V.; Macgill, R.S.; Lin, J.; et al. Identification of Broadly Protective Human Antibodies to Pseudomonas aeruginosa Exopolysaccharide Psl by Phenotypic Screening. J. Exp. Med. 2012, 209, 1273–1287. [Google Scholar] [CrossRef]
- Christen, B.; Christen, M.; Paul, R.; Schmid, F.; Folcher, M.; Jenoe, P.; Meuwly, M.; Jenal, U. Allosteric Control of Cyclic Di-GMP Signaling. J. Biol. Chem. 2006, 281, 32015–32024. [Google Scholar]
- Kulasakara, H.; Lee, V.; Brencic, A.; Liberati, N.; Urbach, J.; Miyata, S.; Lee, D.G.; Neely, A.N.; Hyodo, M.; Hayakawa, Y.; et al. Analysis of Pseudomonas aeruginosa Diguanylate Cyclases and Phosphodiesterases Reveals a Role for Bis-(3'-5')-Cyclic-GMP in Virulence. Proc. Natl. Acad. Sci. USA 2006, 103, 2839–2844. [Google Scholar] [CrossRef]
- Belas, R. Biofilms, flagella, and mechanosensing of surfaces by bacteria. Trends Microbiol. 2014. [Google Scholar] [CrossRef]
- Sintim, H.O.; Smith, J.A.; Wang, J.; Nakayama, S.; Yan, L. Paradigm Shift in Discovering Next-Generation Anti-Infective Agents: Targeting Quorum Sensing, c-Di-GMP Signaling and Biofilm Formation in Bacteria with Small Molecules. Future Med. Chem. 2010, 2, 1005–1035. [Google Scholar] [CrossRef]
- Castiglione, N.; Stelitano, V.; Rinaldo, S.; Giardina, G.; Caruso, M.; Cutruzzolà, F. Metabolism of Cyclic-Di-GMP in Bacterial Biofilms: From a General Overview to Biotechnological Applications. Indian J. Biotechnol. 2011, 10, 423–431. [Google Scholar]
- Irie, Y.; Borlee, B.R.; O’Connor, J.R.; Hill, P.J.; Harwood, C.S.; Wozniak, D.J.; Parsek, M.R. Self-Produced Exopolysaccharide is a Signal that Stimulates Biofilm Formation in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2012, 109, 20632–20636. [Google Scholar]
- Anantharaman, V.; Iyer, L.M.; Aravind, L. Presence of a Classical RRM-Fold Palm Domain in Thg1-Type 3'- 5'Nucleic Acid Polymerases and the Origin of the GGDEF and CRISPR Polymerase Domains. Biol. Direct 2010, 5, 43. [Google Scholar] [CrossRef]
- Mano, E.; Hyodo, M.; Sato, Y.; Ishihara, Y.; Ohta, M.; Hayakawa, Y. Synthesis of Cyclic Bis(3'-5')-2'-deoxyguanylic/guanylic Acid (c-dGpGp) and its Biological Activities to Microbes. ChemMedChem 2007, 2, 1410–1413. [Google Scholar] [CrossRef]
- Barraud, N.; Schleheck, D.; Klebensberger, J.; Webb, J.S.; Hassett, D.J.; Rice, S.A.; Kjelleberg, S. Nitric Oxide Signaling in Pseudomonas aeruginosa Biofilms Mediates Phosphodiesterase Activity, Decreased Cyclic Di-GMP Levels, and Enhanced Dispersal. J. Bacteriol. 2009, 191, 7333–7342. [Google Scholar] [CrossRef]
- Yan, H.; Wang, X.; KuoLee, R.; Chen, W. Synthesis and Immunostimulatory Properties of the Phosphorothioate Analogues of cdiGMP. Bioorg. Med. Chem. Lett. 2008, 18, 5631–5634. [Google Scholar] [CrossRef]
- Ghafoor, A.; Hay, I.D.; Rehm, B.H. Role of Exopolysaccharides in Pseudomonas aeruginosa Biofilm Formation and Architecture. Appl. Environ. Microbiol. 2011, 77, 5238–5246. [Google Scholar] [CrossRef]
- Whitchurch, C.B.; Tolker-Nielsen, T.; Ragas, P.C.; Mattick, J.S. Extracellular DNA Required for Bacterial Biofilm Formation. Science 2002, 295, 1487. [Google Scholar] [CrossRef]
- Ma, L.; Jackson, K.D.; Landry, R.M.; Parsek, M.R.; Wozniak, D.J. Analysis of Pseudomonas aeruginosa Conditional Psl Variants Reveals Roles for the Psl Polysaccharide in Adhesion and Maintaining Biofilm Structure Postattachment. J. Bacteriol. 2006, 188, 8213–8221. [Google Scholar] [CrossRef]
- Mulcahy, H.; Charron-Mazenod, L.; Lewenza, S. Extracellular DNA Chelates Cations and Induces Antibiotic Resistance in Pseudomonas aeruginosa Biofilms. PLoS Pathog. 2008, 4, e1000213. [Google Scholar] [CrossRef]
- May, T.B.; Shinabarger, D.; Maharaj, R.; Kato, J.; Chu, L.; DeVault, J.D.; Roychoudhury, S.; Zielinski, N.A.; Berry, A.; Rothmel, R.K. Alginate Synthesis by Pseudomonas aeruginosa: A Key Pathogenic Factor in Chronic Pulmonary Infections of Cystic Fibrosis Patients. Clin. Microbiol. Rev. 1991, 4, 191–206. [Google Scholar]
- Kumon, H.; Tomochika, K.; Matunaga, T.; Ogawa, M.; Ohmori, H. A Sandwich Cup Method for the Penetration Assay of Antimicrobial Agents through Pseudomonas Exopolysaccharides. Microbiol. Immunol. 1994, 38, 615–619. [Google Scholar] [CrossRef]
- DeVault, J.D.; Kimbara, K.; Chakrabarty, A.M. Pulmonary Dehydration and Infection in Cystic Fibrosis: Evidence that Ethanol Activates Alginate Gene Expression and Induction of Mucoidy in Pseudomonas aeruginosa. Mol. Microbiol. 1990, 4, 737–745. [Google Scholar] [CrossRef]
- Zielinski, N.A.; Maharaj, R.; Roychoudhury, S.; Danganan, C.E.; Hendrickson, W.; Chakrabarty, A.M. Alginate Synthesis in Pseudomonas aeruginosa: Environmental Regulation of the algC Promoter. J. Bacteriol. 1992, 174, 7680–7688. [Google Scholar]
- Gacesa, P. Bacterial Alginate Biosynthesis-Recent Progress and Future Prospects. Microbiology 1998, 144, 1133–1143. [Google Scholar] [CrossRef]
- Maharaj, R.; May, T.B.; Wang, S.K.; Chakrabarty, A.M. Sequence of the alg8 and alg44 Genes Involved in the Synthesis of Alginate by Pseudomonas aeruginosa. Gene 1993, 136, 267–269. [Google Scholar] [CrossRef]
- Boyd, A.; Chakrabarty, A.M. Role of Alginate Lyase in Cell Detachment of Pseudomonas aeruginosa. Appl. Environ. Microbiol. 1994, 60, 2355–2359. [Google Scholar]
- Robles-Price, A.; Wong, T.Y.; Sletta, H.; Valla, S.; Schiller, N.L. AlgX is a Periplasmic Protein Required for Alginate Biosynthesis in Pseudomonas aeruginosa. J. Bacteriol. 2004, 186, 7369–7377. [Google Scholar] [CrossRef]
- Douthit, S.A.; Dlakic, M.; Ohman, D.E.; Franklin, M.J. Epimerase Active Domain of Pseudomonas aeruginosa AlgG, a Protein that Contains a Right-Handed Beta-Helix. J. Bacteriol. 2005, 187, 4573–4583. [Google Scholar] [CrossRef]
- Franklin, M.J.; Ohman, D.E. Mutant Analysis and Cellular Localization of the AlgI, AlgJ, and AlgF Proteins Required for O Acetylation of Alginate in Pseudomonas aeruginosa. J. Bacteriol. 2002, 184, 3000–3007. [Google Scholar] [CrossRef]
- Franklin, M.J.; Douthit, S.A.; McClure, M.A. Evidence that the algI/algJ Gene Cassette, Required for O Acetylation of Pseudomonas aeruginosa Alginate, Evolved by Lateral Gene Transfer. J. Bacteriol. 2004, 186, 4759–4773. [Google Scholar] [CrossRef]
- Rehm, B.H.; Valla, S. Bacterial Alginates: Biosynthesis and Applications. Appl. Microbiol. Biotechnol. 1997, 48, 281–288. [Google Scholar] [CrossRef]
- Ramsey, D.M.; Wozniak, D.J. Understanding the Control of Pseudomonas aeruginosa Alginate Synthesis and the Prospects for Management of Chronic Infections in Cystic Fibrosis. Mol. Microbiol. 2005, 56, 309–322. [Google Scholar] [CrossRef]
- Gimmestad, M.; Sletta, H.; Ertesvag, H.; Bakkevig, K.; Jain, S.; Suh, S.J.; Skjak-Braek, G.; Ellingsen, T.E.; Ohman, D.E.; Valla, S. The Pseudomonas Fluorescens AlgG Protein, but Not its Mannuronan C-5-Epimerase Activity, is Needed for Alginate Polymer Formation. J. Bacteriol. 2003, 185, 3515–3523. [Google Scholar] [CrossRef]
- Shinabarger, D.; Berry, A.; May, T.B.; Rothmel, R.; Fialho, A.; Chakrabarty, A.M. Purification and Characterization of Phosphomannose Isomerase-Guanosine Diphospho-d-Mannose Pyrophosphorylase. A Bifunctional Enzyme in the Alginate Biosynthetic Pathway of Pseudomonas aeruginosa. J. Biol. Chem. 1991, 266, 2080–2088. [Google Scholar]
- Regni, C.; Naught, L.; Tipton, P.A.; Beamer, L.J. Structural Basis of Diverse Substrate Recognition by the Enzyme PMM/PGM from P. aeruginosa. Structure 2004, 12, 55–63. [Google Scholar] [CrossRef]
- Govan, J.R.; Deretic, V. Microbial Pathogenesis in Cystic Fibrosis: Mucoid Pseudomonas aeruginosa and Burkholderia. cepacia. Microbiol. Rev. 1996, 60, 539–574. [Google Scholar]
- DeVries, C.A.; Ohman, D.E. Mucoid-to-Nonmucoid Conversion in Alginate-Producing Pseudomonas aeruginosa often Results from Spontaneous Mutations in algT, Encoding a Putative Alternate Sigma Factor, and shows Evidence for Autoregulation. J. Bacteriol. 1994, 176, 6677–6687. [Google Scholar]
- Wozniak, D.J.; Sprinkle, A.B.; Baynham, P.J. Control of Pseudomonas aeruginosa algZ Expression by the Alternative Sigma Factor AlgT. J. Bacteriol. 2003, 185, 7297–7300. [Google Scholar] [CrossRef]
- Nikolskaya, A.N.; Galperin, M.Y. A Novel Type of Conserved DNA-Binding Domain in the Transcriptional Regulators of the AlgR/AgrA/LytR Family. Nucl. Acids Res. 2002, 30, 2453–2459. [Google Scholar] [CrossRef]
- Woolwine, S.C.; Wozniak, D.J. Identification of an Escherichia coli pepA Homolog and its Involvement in Suppression of the algB Phenotype in Mucoid Pseudomonas aeruginosa. J. Bacteriol. 1999, 181, 107–116. [Google Scholar]
- Leech, A.J.; Sprinkle, A.; Wood, L.; Wozniak, D.J.; Ohman, D.E. The NtrC Family Regulator AlgB, which Controls Alginate Biosynthesis in Mucoid Pseudomonas aeruginosa, Binds Directly to the algD Promoter. J. Bacteriol. 2008, 190, 581–589. [Google Scholar] [CrossRef]
- Baynham, P.J.; Wozniak, D.J. Identification and Characterization of AlgZ, an AlgT-Dependent DNA-Binding Protein Required for Pseudomonas aeruginosa algD Transcription. Mol. Microbiol. 1996, 22, 97–108. [Google Scholar] [CrossRef]
- Tart, A.H.; Blanks, M.J.; Wozniak, D.J. The AlgT-Dependent Transcriptional Regulator AmrZ (AlgZ) Inhibits Flagellum Biosynthesis in Mucoid, Nonmotile Pseudomonas aeruginosa Cystic Fibrosis Isolates. J. Bacteriol. 2006, 188, 6483–6489. [Google Scholar] [CrossRef]
- Mathee, K.; McPherson, C.J.; Ohman, D.E. Posttranslational Control of the algT (algU)-Encoded sigma22 for Expression of the Alginate Regulon in Pseudomonas aeruginosa and Localization of its Antagonist Proteins MucA and MucB (AlgN). J. Bacteriol. 1997, 179, 3711–3720. [Google Scholar]
- Schurr, M.J.; Yu, H.; Martinez-Salazar, J.M.; Boucher, J.C.; Deretic, V. Control of AlgU, a Member of the Sigma E-Like Family of Stress Sigma Factors, by the Negative Regulators MucA and MucB and Pseudomonas aeruginosa Conversion to Mucoidy in Cystic Fibrosis. J. Bacteriol. 1996, 178, 4997–5004. [Google Scholar]
- Rowen, D.W.; Deretic, V. Membrane-to-Cytosol Redistribution of ECF Sigma Factor AlgU and Conversion to Mucoidy in Pseudomonas aeruginosa Isolates from Cystic Fibrosis Patients. Mol. Microbiol. 2000, 36, 314–327. [Google Scholar] [CrossRef]
- Boucher, J.C.; Schurr, M.J.; Yu, H.; Rowen, D.W.; Deretic, V. Pseudomonas aeruginosa in Cystic Fibrosis: Role of mucC in the Regulation of Alginate Production and Stress Sensitivity. Microbiology 1997, 143, 3473–3480. [Google Scholar] [CrossRef]
- Wood, L.F.; Ohman, D.E. Independent Regulation of MucD, an HtrA-Like Protease in Pseudomonas aeruginosa, and the Role of its Proteolytic Motif in Alginate Gene Regulation. J. Bacteriol. 2006, 188, 3134–3137. [Google Scholar] [CrossRef]
- Goldberg, J.B.; Gorman, W.L.; Flynn, J.L.; Ohman, D.E. A Mutation in algN Permits Trans Activation of Alginate Production by algT in Pseudomonas Species. J. Bacteriol. 1993, 175, 1303–1308. [Google Scholar]
- Martin, D.W.; Schurr, M.J.; Mudd, M.H.; Deretic, V. Differentiation of Pseudomonas aeruginosa into the Alginate-Producing Form: Inactivation of mucB Causes Conversion to Mucoidy. Mol. Microbiol. 1993, 9, 497–506. [Google Scholar] [CrossRef]
- Prigent-Combaret, C.; Vidal, O.; Dorel, C.; Lejeune, P. Abiotic Surface Sensing and Biofilm-Dependent Regulation of Gene Expression in Escherichia coli. J. Bacteriol. 1999, 181, 5993–6002. [Google Scholar]
- Danese, P.N.; Pratt, L.A.; Kolter, R. Exopolysaccharide Production is Required for Development of Escherichia coli K12 Biofilm Architecture. J. Bacteriol. 2000, 182, 3593–3596. [Google Scholar] [CrossRef]
- Obadia, B.; Lacour, S.; Doublet, P.; Baubichon-Cortay, H.; Cozzone, A.J.; Grangeasse, C. Influence of Tyrosine-Kinase Wzc Activity on Colanic Acid Production in Escherichia coli K12 Cells. J. Mol. Biol. 2007, 367, 42–53. [Google Scholar] [CrossRef]
- Whitfield, C.; Roberts, I.S. Structure, Assembly and Regulation of Expression of Capsules in Escherichia coli. Mol. Microbiol. 1999, 31, 1307–1319. [Google Scholar] [CrossRef]
- Ebel, W.; Trempy, J.E. Escherichia coli RcsA, a Positive Activator of Colanic Acid Capsular Polysaccharide Synthesis, Functions to Activate its Own Expression. J. Bacteriol. 1999, 181, 577–584. [Google Scholar]
- Gervais, F.G.; Drapeau, G.R. Identification, Cloning, and Characterization of rcsF, a New Regulator Gene for Exopolysaccharide Synthesis that Suppresses the Division Mutation ftsZ84 in Escherichia coli K-12. J. Bacteriol. 1992, 174, 8016–8022. [Google Scholar]
- Lacroix, J.M.; Loubens, I.; Tempete, M.; Menichi, B.; Bohin, J.P. The mdoA Locus of Escherichia coli Consists of an Operon Under Osmotic Control. Mol. Microbiol. 1991, 5, 1745–1753. [Google Scholar] [CrossRef]
- Ebel, W.; Vaughn, G.J.; Peters, H.K., 3rd; Trempy, J.E. Inactivation of mdoH Leads to Increased Expression of Colanic Acid Capsular Polysaccharide in Escherichia coli. J. Bacteriol. 1997, 179, 6858–6861. [Google Scholar]
- Sailer, F.C.; Meberg, B.M.; Young, K.D. Beta-Lactam Induction of Colanic Acid Gene Expression in Escherichia coli. FEMS Microbiol. Lett. 2003, 226, 245–249. [Google Scholar] [CrossRef]
- Sledjeski, D.; Gottesman, S. A Small RNA Acts as an Antisilencer of the H-NS-Silenced rcsA Gene of Escherichia coli. Proc. Natl. Acad. Sci. USA 1995, 92, 2003–2007. [Google Scholar] [CrossRef]
- Vincent, C.; Doublet, P.; Grangeasse, C.; Vaganay, E.; Cozzone, A.J.; Duclos, B. Cells of Escherichia coli Contain a Protein-Tyrosine Kinase, Wzc, and a Phosphotyrosine-Protein Phosphatase, Wzb. J. Bacteriol. 1999, 181, 3472–3477. [Google Scholar]
- Grangeasse, C.; Cozzone, A.J.; Deutscher, J.; Mijakovic, I. Tyrosine Phosphorylation: An Emerging Regulatory Device of Bacterial Physiology. Trends Biochem. Sci. 2007, 32, 86–94. [Google Scholar] [CrossRef]
- Mijakovic, I.; Poncet, S.; Boel, G.; Maze, A.; Gillet, S.; Jamet, E.; Decottignies, P.; Grangeasse, C.; Doublet, P.; le Marechal, P.; Deutscher, J. Transmembrane Modulator-Dependent Bacterial Tyrosine Kinase Activates UDP-Glucose Dehydrogenases. EMBO J. 2003, 22, 4709–4718. [Google Scholar] [CrossRef]
- Lee, D.C.; Zheng, J.; She, Y.M.; Jia, Z. Structure of Escherichia coli Tyrosine Kinase Etk Reveals a Novel Activation Mechanism. EMBO J. 2008, 27, 1758–1766. [Google Scholar] [CrossRef]
- Peleg, A.; Shifrin, Y.; Ilan, O.; Nadler-Yona, C.; Nov, S.; Koby, S.; Baruch, K.; Altuvia, S.; Elgrably-Weiss, M.; Abe, C.M.; Knutton, S.; Saper, M.A.; Rosenshine, I. Identification of an Escherichia coli Operon Required for Formation of the O-Antigen Capsule. J. Bacteriol. 2005, 187, 5259–5266. [Google Scholar] [CrossRef]
- Lacour, S.; Bechet, E.; Cozzone, A.J.; Mijakovic, I.; Grangeasse, C. Tyrosine Phosphorylation of the UDP-Glucose Dehydrogenase of Escherichia coli is at the Crossroads of Colanic Acid Synthesis and Polymyxin Resistance. PLoS One 2008, 3, e3053. [Google Scholar]
- Raetz, C.R.; Reynolds, C.M.; Trent, M.S.; Bishop, R.E. Lipid A Modification Systems in Gram-negative Bacteria. Ann. Rev. Biochem. 2007, 76, 295–329. [Google Scholar] [CrossRef]
- Meredith, T.C.; Mamat, U.; Kaczynski, Z.; Lindner, B.; Holst, O.; Woodard, R.W. Modification of Lipopolysaccharide with Colanic Acid (M-Antigen) Repeats in Escherichia coli. J. Biol. Chem. 2007, 282, 7790–7798. [Google Scholar]
- Passador, L.; Cook, J.M.; Gambello, M.J.; Rust, L.; Iglewski, B.H. Expression of Pseudomonas aeruginosa Virulence Genes Requires Cell-to-Cell Communication. Science 1993, 260, 1127–1130. [Google Scholar]
- Ochsner, U.A.; Reiser, J. Autoinducer-Mediated Regulation of Rhamnolipid Biosurfactant Synthesis in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 1995, 92, 6424–6428. [Google Scholar] [CrossRef]
- Diggle, S.P.; Cornelis, P.; Williams, P.; Camara, M. 4-Quinolone Signalling in Pseudomonas aeruginosa: Old Molecules, New Perspectives. Int. J. Med. Microbiol. 2006, 296, 83–91. [Google Scholar] [CrossRef]
- Latifi, A.; Foglino, M.; Tanaka, K.; Williams, P.; Lazdunski, A. A Hierarchical Quorum-Sensing Cascade in Pseudomonas aeruginosa Links the Transcriptional Activators LasR and RhIR (VsmR) to Expression of the Stationary-Phase Sigma Factor RpoS. Mol. Microbiol. 1996, 21, 1137–1146. [Google Scholar]
- Wagner, V.E.; Bushnell, D.; Passador, L.; Brooks, A.I.; Iglewski, B.H. Microarray Analysis of Pseudomonas aeruginosa Quorum-Sensing Regulons: Effects of Growth Phase and Environment. J. Bacteriol. 2003, 185, 2080–2095. [Google Scholar] [CrossRef]
- Winson, M.K.; Camara, M.; Latifi, A.; Foglino, M.; Chhabra, S.R.; Daykin, M.; Bally, M.; Chapon, V.; Salmond, G.P.; Bycroft, B.W. Multiple N-Acyl-l-Homoserine Lactone Signal Molecules Regulate Production of Virulence Determinants and Secondary Metabolites in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 1995, 92, 9427–9431. [Google Scholar] [CrossRef]
- Davies, D.G.; Parsek, M.R.; Pearson, J.P.; Iglewski, B.H.; Costerton, J.W.; Greenberg, E.P. The Involvement of Cell-to-Cell Signals in the Development of a Bacterial Biofilm. Science 1998, 280, 295–298. [Google Scholar] [CrossRef]
- Hentzer, M.; Eberl, L.; Nielsen, J.; Givskov, M. Quorum Sensing: A Novel Target for the Treatment of Biofilm Infections. BioDrugs 2003, 17, 241–250. [Google Scholar] [CrossRef]
- Sakuragi, Y.; Kolter, R. Quorum-Sensing Regulation of the Biofilm Matrix Genes (pel) of Pseudomonas aeruginosa. J. Bacteriol. 2007, 189, 5383–5386. [Google Scholar] [CrossRef]
- Pearson, J.P.; Gray, K.M.; Passador, L.; Tucker, K.D.; Eberhard, A.; Iglewski, B.H.; Greenberg, E.P. Structure of the Autoinducer Required for Expression of Pseudomonas aeruginosa Virulence Genes. Proc. Natl. Acad. Sci. USA 1994, 91, 197–201. [Google Scholar]
- De Kievit, T.R.; Iglewski, B.H. Bacterial Quorum Sensing in Pathogenic Relationships. Infect. Immun. 2000, 68, 4839–4849. [Google Scholar] [CrossRef]
- Pesci, E.C.; Iglewski, B.H. The Chain of Command in Pseudomonas Quorum Sensing. Trends Microbiol. 1997, 5, 132–135. [Google Scholar] [CrossRef]
- Davey, M.E.; Caiazza, N.C.; O’Toole, G.A. Rhamnolipid Surfactant Production Affects Biofilm Architecture in Pseudomonas aeruginosa PAO1. J. Bacteriol. 2003, 185, 1027–1036. [Google Scholar] [CrossRef]
- Williams, P.; Camara, M. Quorum Sensing and Environmental Adaptation in Pseudomonas aeruginosa: A Tale of Regulatory Networks and Multifunctional Signal Molecules. Curr. Opin. Microbiol. 2009, 12, 182–191. [Google Scholar] [CrossRef]
- Beatson, S.A.; Whitchurch, C.B.; Semmler, A.B.; Mattick, J.S. Quorum Sensing is Not Required for Twitching Motility in Pseudomonas aeruginosa. J. Bacteriol. 2002, 184, 3598–3604. [Google Scholar] [CrossRef]
- Raina, S.; de Vizio, D.; Odell, M.; Clements, M.; Vanhulle, S.; Keshavarz, T. Microbial Quorum Sensing: A Tool Or a Target for Antimicrobial Therapy? Biotechnol. Appl. Biochem. 2009, 54, 65–84. [Google Scholar] [CrossRef]
- McKnight, S.L.; Iglewski, B.H.; Pesci, E.C. The Pseudomonas Quinolone Signal Regulates rhl Quorum Sensing in Pseudomonas aeruginosa. J. Bacteriol. 2000, 182, 2702–2708. [Google Scholar] [CrossRef]
- Diggle, S.P.; Winzer, K.; Chhabra, S.R.; Worrall, K.E.; Camara, M.; Williams, P. The Pseudomonas aeruginosa Quinolone Signal Molecule Overcomes the Cell Density-Dependency of the Quorum Sensing Hierarchy, Regulates rhl-Dependent Genes at the Onset of Stationary Phase and can be Produced in the Absence of LasR. Mol. Microbiol. 2003, 50, 29–43. [Google Scholar] [CrossRef]
- Pesci, E.C.; Milbank, J.B.; Pearson, J.P.; McKnight, S.; Kende, A.S.; Greenberg, E.P.; Iglewski, B.H. Quinolone Signaling in the Cell-to-Cell Communication System of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 1999, 96, 11229–11234. [Google Scholar]
- Aendekerk, S.; Diggle, S.P.; Song, Z.; Hoiby, N.; Cornelis, P.; Williams, P.; Camara, M. The MexGHI-OpmD Multidrug Efflux Pump Controls Growth, Antibiotic Susceptibility and Virulence in Pseudomonas aeruginosa Via 4-Quinolone-Dependent Cell-to-Cell Communication. Microbiology 2005, 151, 1113–1125. [Google Scholar] [CrossRef]
- Juhas, M.; Wiehlmann, L.; Huber, B.; Jordan, D.; Lauber, J.; Salunkhe, P.; Limpert, A.S.; von Gotz, F.; Steinmetz, I.; Eberl, L.; Tummler, B. Global Regulation of Quorum Sensing and Virulence by VqsR in Pseudomonas aeruginosa. Microbiology 2004, 150, 831–841. [Google Scholar] [CrossRef]
- Ledgham, F.; Ventre, I.; Soscia, C.; Foglino, M.; Sturgis, J.N.; Lazdunski, A. Interactions of the Quorum Sensing Regulator QscR: Interaction with itself and the Other Regulators of Pseudomonas aeruginosa LasR and RhlR. Mol. Microbiol. 2003, 48, 199–210. [Google Scholar] [CrossRef]
- Rampioni, G.; Bertani, I.; Zennaro, E.; Polticelli, F.; Venturi, V.; Leoni, L. The Quorum-Sensing Negative Regulator RsaL of Pseudomonas aeruginosa Binds to the lasI Promoter. J. Bacteriol. 2006, 188, 815–819. [Google Scholar] [CrossRef]
- Rampioni, G.; Polticelli, F.; Bertani, I.; Righetti, K.; Venturi, V.; Zennaro, E.; Leoni, L. The Pseudomonas Quorum-Sensing Regulator RsaL Belongs to the Tetrahelical Superclass of H-T-H Proteins. J. Bacteriol. 2007, 189, 1922–1930. [Google Scholar] [CrossRef]
- Hentzer, M.; Wu, H.; Andersen, J.B.; Riedel, K.; Rasmussen, T.B.; Bagge, N.; Kumar, N.; Schembri, M.A.; Song, Z.; Kristoffersen, P.; et al. Attenuation of Pseudomonas aeruginosa Virulence by Quorum Sensing Inhibitors. EMBO J. 2003, 22, 3803–3815. [Google Scholar] [CrossRef]
- Geske, G.D.; Wezeman, R.J.; Siegel, A.P.; Blackwell, H.E. Small Molecule Inhibitors of Bacterial Quorum Sensing and Biofilm Formation. J. Am. Chem. Soc. 2005, 127, 12762–12763. [Google Scholar] [CrossRef]
- Jorgensen, F.; Bally, M.; Chapon-Herve, V.; Michel, G.; Lazdunski, A.; Williams, P.; Stewart, G.S. RpoS-Dependent Stress Tolerance in Pseudomonas aeruginosa. Microbiology 1999, 145, 835–844. [Google Scholar] [CrossRef]
- Kojic, M.; Venturi, V. Regulation of rpoS Gene Expression in Pseudomonas: Involvement of a TetR Family Regulator. J. Bacteriol. 2001, 183, 3712–3720. [Google Scholar] [CrossRef]
- Whiteley, M.; Parsek, M.R.; Greenberg, E.P. Regulation of Quorum Sensing by RpoS in Pseudomonas aeruginosa. J. Bacteriol. 2000, 182, 4356–4360. [Google Scholar] [CrossRef]
- Medina, G.; Juarez, K.; Soberon-Chavez, G. The Pseudomonas aeruginosa rhlAB Operon is Not Expressed during the Logarithmic Phase of Growth Even in the Presence of its Activator RhlR and the Autoinducer N-Butyryl-Homoserine Lactone. J. Bacteriol. 2003, 185, 377–380. [Google Scholar] [CrossRef]
- Schuster, M.; Hawkins, A.C.; Harwood, C.S.; Greenberg, E.P. The Pseudomonas aeruginosa RpoS Regulon and its Relationship to Quorum Sensing. Mol. Microbiol. 2004, 51, 973–985. [Google Scholar] [CrossRef]
- Ahmer, B.M. Cell-to-Cell Signalling in Escherichia coli and Salmonella enterica. Mol. Microbiol. 2004, 52, 933–945. [Google Scholar] [CrossRef]
- Van Houdt, R.; Aertsen, A.; Moons, P.; Vanoirbeek, K.; Michiels, C.W. N-Acyl-l-Homoserine Lactone Signal Interception by Escherichia coli. FEMS Microbiol. Lett. 2006, 256, 83–89. [Google Scholar] [CrossRef]
- De Keersmaecker, S.C.; Sonck, K.; Vanderleyden, J. Let LuxS Speak Up in AI-2 Signaling. Trends Microbiol. 2006, 14, 114–119. [Google Scholar] [CrossRef]
- Walters, M.; Sperandio, V. Quorum Sensing in Escherichia coli and Salmonella. Int. J. Med. Microbiol. 2006, 296, 125–131. [Google Scholar] [CrossRef]
- Kanamaru, K.; Kanamaru, K.; Tatsuno, I.; Tobe, T.; Sasakawa, C. SdiA, an Escherichia coli Homologue of Quorum-Sensing Regulators, Controls the Expression of Virulence Factors in Enterohaemorrhagic Escherichia coli O157:H7. Mol. Microbiol. 2000, 38, 805–816. [Google Scholar] [CrossRef]
- Nealson, K.H.; Platt, T.; Hastings, J.W. Cellular Control of the Synthesis and Activity of the Bacterial Luminescent System. J. Bacteriol. 1970, 104, 313–322. [Google Scholar]
- Michael, B.; Smith, J.N.; Swift, S.; Heffron, F.; Ahmer, B.M. SdiA of Salmonella enterica is a LuxR Homolog that Detects Mixed Microbial Communities. J. Bacteriol. 2001, 183, 5733–5742. [Google Scholar] [CrossRef]
- Ravichandiran, V.; Shanmugam, K.; Solomon, A.P. Screening of SdiA Inhibitors from Melia Dubia Seeds Extracts Towards the Hold Back of Uropathogenic E. Coli Quorum Sensing-Regulated Factors. J. Med. Chem. 2013, 9, 819–827. [Google Scholar] [CrossRef]
- Chong, T.M.; Koh, C.L.; Sam, C.K.; Choo, Y.M.; Yin, W.F.; Chan, K.G. Characterization of Quorum Sensing and Quorum Quenching Soil Bacteria Isolated from Malaysian Tropical Montane Forest. Sensors 2012, 12, 4846–4859. [Google Scholar] [CrossRef]
- Bjarnsholt, T.; Jensen, P.O.; Rasmussen, T.B.; Christophersen, L.; Calum, H.; Hentzer, M.; Hougen, H.P.; Rygaard, J.; Moser, C.; Eberl, L.; et al. Garlic Blocks Quorum Sensing and Promotes Rapid Clearing of Pulmonary Pseudomonas aeruginosa Infections. Microbiology 2005, 151, 3873–3880. [Google Scholar] [CrossRef]
- Jha, B.; Kavita, K.; Westphal, J.; Hartmann, A.; Schmitt-Kopplin, P. Quorum Sensing Inhibition by Asparagopsis. taxiformis, a Marine Macro Alga: Separation of the Compound that Interrupts Bacterial Communication. Mar. Drugs 2013, 11, 253–265. [Google Scholar] [CrossRef]
- March, J.C.; Bentley, W.E. Quorum Sensing and Bacterial Cross-Talk in Biotechnology. Curr. Opin. Biotechnol. 2004, 15, 495–502. [Google Scholar] [CrossRef]
- Hardie, K.R.; Cooksley, C.; Green, A.D.; Winzer, K. Autoinducer 2 Activity in Escherichia coli Culture Supernatants can be Actively Reduced Despite Maintenance of an Active Synthase, LuxS. Microbiology 2003, 149, 715–728. [Google Scholar] [CrossRef]
- Xavier, K.B.; Bassler, B.L. Regulation of Uptake and Processing of the Quorum-Sensing Autoinducer AI-2 in Escherichia coli. J. Bacteriol. 2005, 187, 238–248. [Google Scholar] [CrossRef]
- Wang, L.; Li, J.; March, J.C.; Valdes, J.J.; Bentley, W.E. luxS-Dependent Gene Regulation in Escherichia coli K-12 Revealed by Genomic Expression Profiling. J. Bacteriol. 2005, 187, 8350–8360. [Google Scholar] [CrossRef]
- Taga, M.E.; Miller, S.T.; Bassler, B.L. Lsr-Mediated Transport and Processing of AI-2 in Salmonella typhimurium. Mol. Microbiol. 2003, 50, 1411–1427. [Google Scholar] [CrossRef]
- Li, J.; Attila, C.; Wang, L.; Wood, T.K.; Valdes, J.J.; Bentley, W.E. Quorum Sensing in Escherichia coli is Signaled by AI-2/LsrR: Effects on Small RNA and Biofilm Architecture. J. Bacteriol. 2007, 189, 6011–6020. [Google Scholar] [CrossRef]
- Robinson, L.S.; Ashman, E.M.; Hultgren, S.J.; Chapman, M.R. Secretion of Curli Fibre Subunits is Mediated by the Outer Membrane-Localized CsgG Protein. Mol. Microbiol. 2006, 59, 870–881. [Google Scholar] [CrossRef]
- Barnhart, M.M.; Chapman, M.R. Curli Biogenesis and Function. Ann. Rev. Microbiol. 2006, 60, 131–147. [Google Scholar] [CrossRef]
- DeLisa, M.P.; Wu, C.F.; Wang, L.; Valdes, J.J.; Bentley, W.E. DNA Microarray-Based Identification of Genes Controlled by Autoinducer 2-Stimulated Quorum Sensing in Escherichia coli. J. Bacteriol. 2001, 183, 5239–5247. [Google Scholar] [CrossRef]
- Schauder, S.; Shokat, K.; Surette, M.G.; Bassler, B.L. The LuxS Family of Bacterial Autoinducers: Biosynthesis of a Novel Quorum-Sensing Signal Molecule. Mol. Microbiol. 2001, 41, 463–476. [Google Scholar]
- Diaz, Z.; Xavier, K.B.; Miller, S.T. The Crystal Structure of the Escherichia coli Autoinducer-2 Processing Protein LsrF. PLoS One 2009, 4, e6820. [Google Scholar] [CrossRef]
- Xavier, K.B.; Miller, S.T.; Lu, W.; Kim, J.H.; Rabinowitz, J.; Pelczer, I.; Semmelhack, M.F.; Bassler, B.L. Phosphorylation and Processing of the Quorum-Sensing Molecule Autoinducer-2 in Enteric Bacteria. ACS Chem. Biol. 2007, 2, 128–136. [Google Scholar] [CrossRef]
- Kendall, M.M.; Rasko, D.A.; Sperandio, V. Global Effects of the Cell-to-Cell Signaling Molecules Autoinducer-2, Autoinducer-3, and Epinephrine in a luxS Mutant of Enterohemorrhagic Escherichia coli. Infect. Immun. 2007, 75, 4875–4884. [Google Scholar] [CrossRef]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef]
- Walters, M.; Sircili, M.P.; Sperandio, V. AI-3 Synthesis is Not Dependent on luxS in Escherichia coli. J. Bacteriol. 2006, 188, 5668–5681. [Google Scholar] [CrossRef]
- Sperandio, V.; Torres, A.G.; Giron, J.A.; Kaper, J.B. Quorum Sensing is a Global Regulatory Mechanism in Enterohemorrhagic Escherichia coli O157:H7. J. Bacteriol. 2001, 183, 5187–5197. [Google Scholar] [CrossRef]
- Freestone, P.P.; Lyte, M.; Neal, C.P.; Maggs, A.F.; Haigh, R.D.; Williams, P.H. The Mammalian Neuroendocrine Hormone Norepinephrine Supplies Iron for Bacterial Growth in the Presence of Transferrin Or Lactoferrin. J. Bacteriol. 2000, 182, 6091–6098. [Google Scholar] [CrossRef]
- Sperandio, V.; Torres, A.G.; Kaper, J.B. Quorum Sensing Escherichia Coli Regulators B and C (QseBC): A Novel Two-Component Regulatory System Involved in the Regulation of Flagella and Motility by Quorum Sensing in E. Coli. Mol. Microbiol. 2002, 43, 809–821. [Google Scholar] [CrossRef]
- Reading, N.C.; Rasko, D.A.; Torres, A.G.; Sperandio, V. The Two-Component System QseEF and the Membrane Protein QseG Link Adrenergic and Stress Sensing to Bacterial Pathogenesis. Proc. Natl. Acad. Sci. USA 2009, 106, 5889–5894. [Google Scholar]
- Sperandio, V.; Li, C.C.; Kaper, J.B. Quorum-Sensing Escherichia coli Regulator A: A Regulator of the LysR Family Involved in the Regulation of the Locus of Enterocyte Effacement Pathogenicity Island in Enterohemorrhagic E. coli. Infect. Immun. 2002, 70, 3085–3093. [Google Scholar] [CrossRef]
- Wang, D.; Ding, X.; Rather, P.N. Indole can Act as an Extracellular Signal in Escherichia coli. J. Bacteriol. 2001, 183, 4210–4216. [Google Scholar] [CrossRef]
- World Health Organization. Antimicrobial Resistance: Global Report on Surveillance 2014; WHO: Geneva, Switzerland, 2014. [Google Scholar]
- Ito, A.; Taniuchi, A.; May, T.; Kawata, K.; Okabe, S. Increased Antibiotic Resistance of Escherichia coli in Mature Biofilms. Appl. Environ. Microbiol. 2009, 75, 4093–4100. [Google Scholar] [CrossRef]
- O’Loughlin, C.T.; Miller, L.C.; Siryaporn, A.; Drescher, K.; Semmelhack, M.F.; Bassler, B.L. A Quorum-Sensing Inhibitor Blocks Pseudomonas aeruginosa Virulence and Biofilm Formation. Proc. Natl. Acad. Sci. USA 2013, 110, 17981–17986. [Google Scholar]
- Deep, A.; Chaudhary, U.; Gupta, V. Quorum Sensing and Bacterial Pathogenicity: From Molecules to Disease. J. Lab. Phys. 2011, 3, 4–11. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Laverty, G.; Gorman, S.P.; Gilmore, B.F. Biomolecular Mechanisms of Pseudomonas aeruginosa and Escherichia coli Biofilm Formation. Pathogens 2014, 3, 596-632. https://doi.org/10.3390/pathogens3030596
Laverty G, Gorman SP, Gilmore BF. Biomolecular Mechanisms of Pseudomonas aeruginosa and Escherichia coli Biofilm Formation. Pathogens. 2014; 3(3):596-632. https://doi.org/10.3390/pathogens3030596
Chicago/Turabian StyleLaverty, Garry, Sean P. Gorman, and Brendan F. Gilmore. 2014. "Biomolecular Mechanisms of Pseudomonas aeruginosa and Escherichia coli Biofilm Formation" Pathogens 3, no. 3: 596-632. https://doi.org/10.3390/pathogens3030596