Wild Mushroom Extracts as Inhibitors of Bacterial Biofilm Formation


 ,
,
Abstract
:1. Introduction
2. Results and Discussion

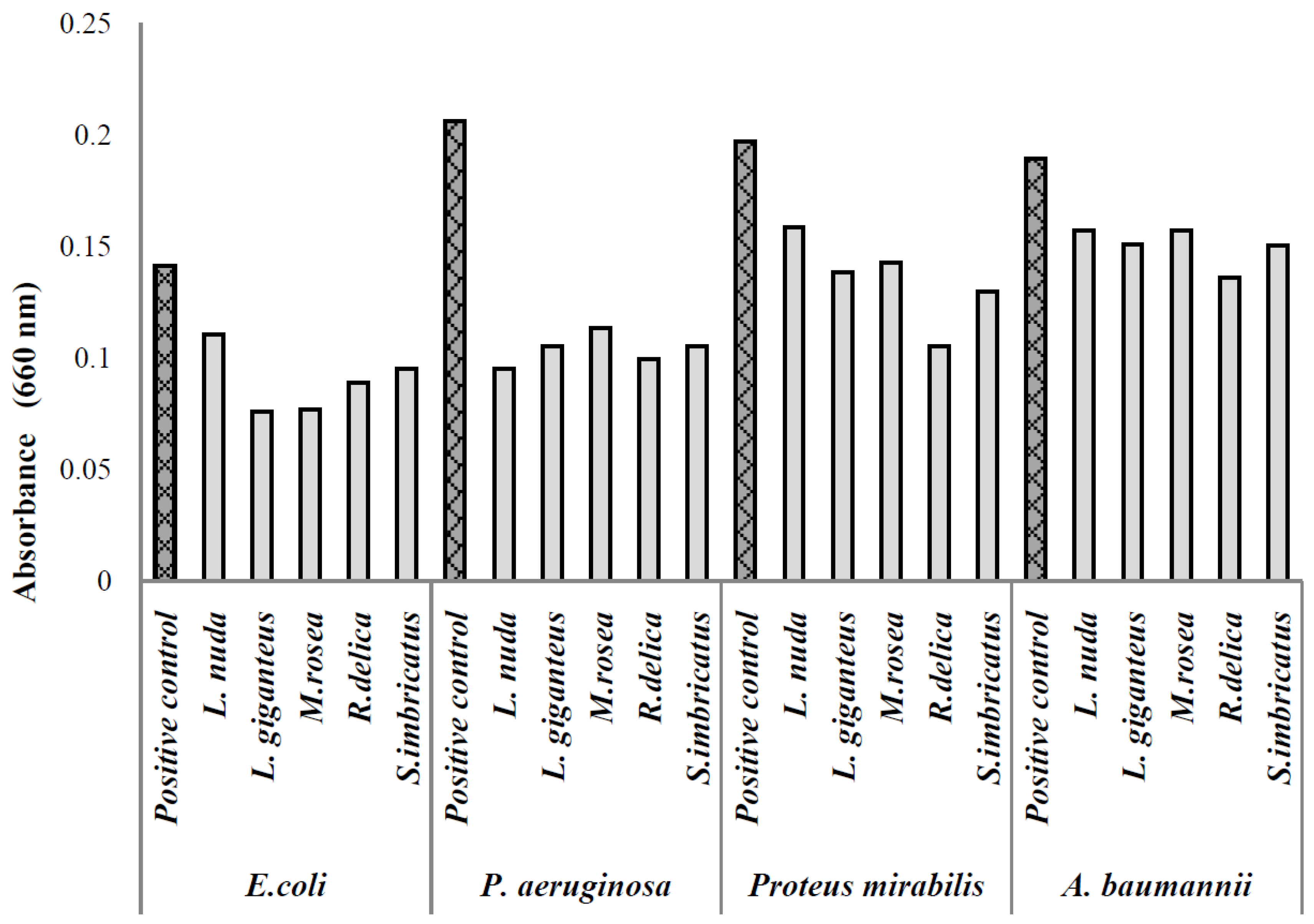{kind=link}
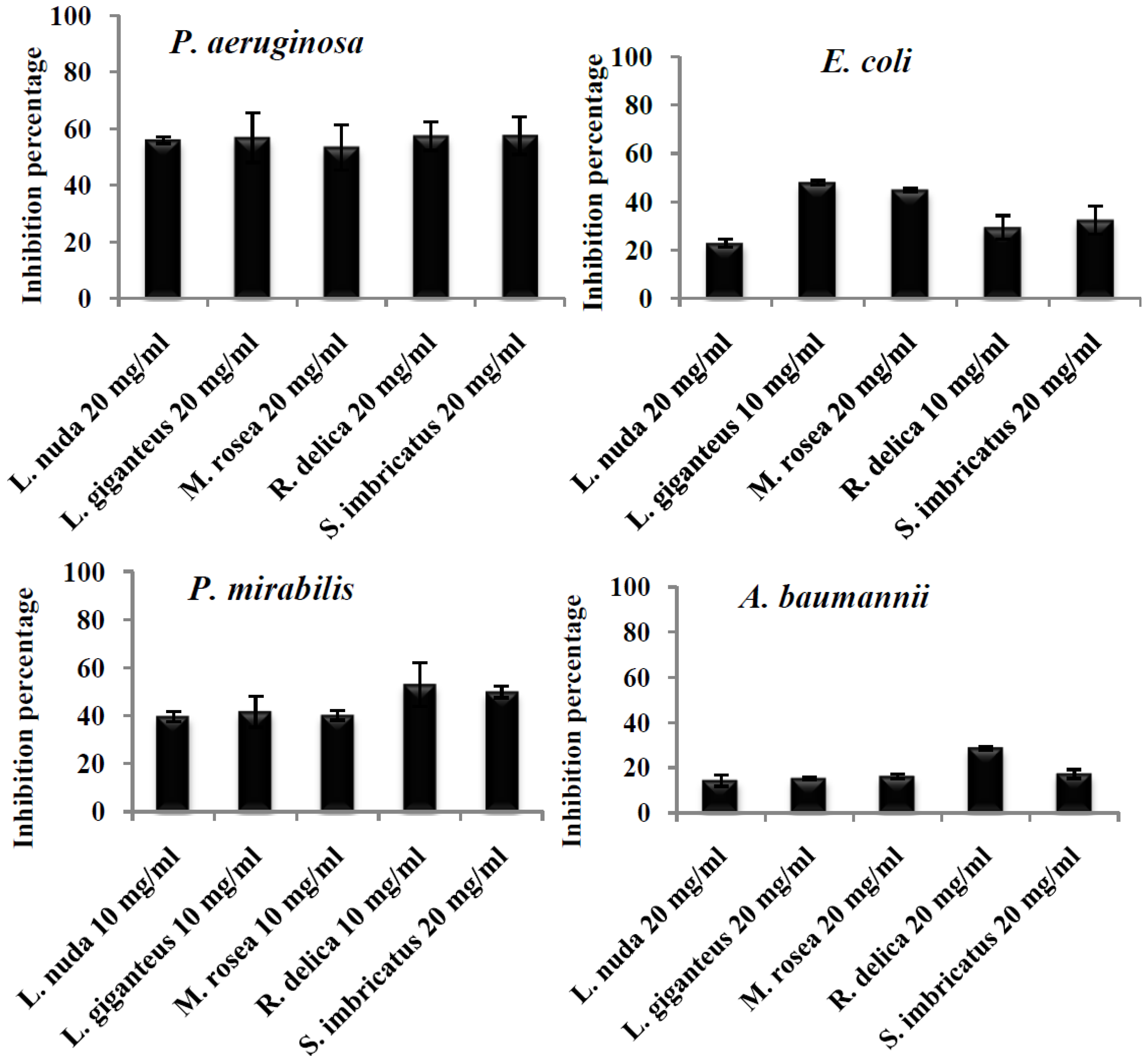{kind=link}
| Mushroom | Average Production Inhibition of Biofilm (%) | |||
|---|---|---|---|---|
| P. aeruginosa | E. coli | A. baumannii | P. mirabilis | |
| L. nuda | 55.99 ± 1.13 ª | 22.89 ± 1.53 c | 14.22 ± 2.52 b | 39.73 ± 2.17 b |
| L. giganteus | 56.79 ± 8.78 ª | 47.84 ± 0.93 ª | 15.12 ± 0.58 b | 41.72 ± 6.53 ab |
| M. rosea | 53.36 ± 8.02 ª | 44.88 ± 0.68 ª | 16.08 ± 1.05 b | 40.24 ± 2.08 ab |
| R. delica | 57.35 ± 5.01 ª | 29.37 ± 4.95 bc | 28.59 ± 0.70 ª | 60.31 ± 1.20 a |
| S. imbricatus | 57.49 ± 6.63 ª | 32.41 ± 5.86 b | 17.15 ± 1.97 b | 49.97 ± 2.53 ab |


| Mushroom | Phenolics (mg GAE/g MWE Extract) | Protocatechuic Acid (mg/kg dw) | p-Hydroxybenzoic Acid (mg/kg dw) | p-Coumaric Acid (mg/kg dw) | Gallic acid (mg/mL EE) | Caffeic Acid (mg/mL EE) | Catechin (mg/mL EE) | Rutin (mg/mL EE) |
|---|---|---|---|---|---|---|---|---|
| L. nuda 1,2 | 6.31 ± 0.13 a | 33.47 ± 0.50 | 29.31 ± 1.54 b | 3.75 ± 0.56 | - | - | - | |
| L. giganteus 3 | 6.29 ± 0.20 a | - | - | - | - | - | - | |
| M. rosea 3 | 3.56 ± 0.37 c | na | na | na | na | na | na | |
| R. delica 3,4 | 2.23 ± 0.18 d | - | - | - | 0.05 | 0.11 | 5.33 | 0.46 |
| S. imbricatus 1 | 3.76 ± 0.11 b | - | 33.19 ± 1.92 a | - | - | - | - |
3. Materials and Methods
3.1. Extracts Preparation
3.2. Bacterial Isolates
3.3. Isolates Identification and Antimicrobial Susceptibility Testing
3.4. Inhibition of Biofilm Formation
3.5. Toxicity Assay
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Centers for Disease Control and Prevention (CDC). Epidemiological Report Reporting on 2011 Surveillance Data and 2012 Epidemic Intelligence Data; Centers Disease Control and Prevention: Atlanta, GA, USA, 2013; p. 199.
- Akpaka, P.E.; Swanston, W.H.; Ihemere, H.N.; Correa, A.; Torres, J.A.; Tafur, J.D.; Montealegre, M.C.; Quinn, J.P.; Villegas, M.V. Emergence of KPC-producing Pseudomonas aeruginosa in Trinidad and Tobago. J. Clin. Microbiol. 2009, 47, 2670–2671. [Google Scholar] [CrossRef] [PubMed]
- Maki, D.G.; Masur, H.; McCormick, R.D.; Mermel, L.A.; Pearson, M.L.; Raad, I.I.; Randolph, A.; Weinstein, R.A. Guidelines for the prevention of intravascular catheter-related infections. Centers for disease control and prevention. Morb. Mortal. Wkly. Rep. 2002, 9, 1–29. [Google Scholar]
- Stahlhut, G.S.; Struve, C.; Krogfelt, A.K.; Reisner, A. Biofilm formation of Klebsiella pneumoniae on urethral catheters requires either type 1 or type 3 fimbriae. FEMS Immunol. Med. Microbiol. 2012, 65, 350–359. [Google Scholar] [CrossRef] [PubMed]
- Wasfi, R.; Abd El-Rahman, O.A.; Mansour, L.E.; Hanora, A.S.; Hashem, A.M.; Ashour, M.S. Antimicrobial activities against biofilm formed by Proteus mirabilis isolates from wound and urinary tract infections. Indian J. Med. Microbiol. 2012, 30, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Foxman, B.; Wu, J.; Farrer, E.C.; Goldberg, D.E.; Younger, J.G.; Xi, C. Early development of bacterial community diversity in emergently placed urinary catheters. BMC Res. Notes 2012, 5, 332. [Google Scholar] [CrossRef] [PubMed]
- Al-Mathkhury, H.J.F.; Ali, A.S.; Ghafil, J.A. Antagonistic effect of bacteriocin against urinary catheter associated Pseudomonas aeruginosa biofilm. N. Am. J. Med. Sci. 2011, 3, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Bonkat, G.; Widmer, A.F.; Rieken, M.; Merwe, A.; Braissant, O.; Müller, G.; Wyler, S.; Frei, R.; Gasser, T.C.; Bachmann, A. Microbial biofilm formation and catheter associated bacteriuria in patients with suprapubic catheterisation. World J. Urol. 2013, 31, 565–571. [Google Scholar] [CrossRef] [PubMed]
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial biofilms: A common cause of persistent infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.T.; Marsh, P.D. A review of biofilms and their role in microbial contamination of dental unit water systems (DUWS). Int. Biodeterior. Biodegrad. 2004, 54, 87–98. [Google Scholar] [CrossRef]
- Branda, S.S.; Vik, S.; Friedman, L.; Kolter, R. Biofilms: The matrix revisited. Trends Microbiol. 2005, 13, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Hall-Stoodley, L.; Stoodley, P. Evolving concepts in biofilm infections. Cell. Microbiol. 2009, 11, 1034–1043. [Google Scholar] [CrossRef] [PubMed]
- Flowers, R.H.K.J.; Schwenzer, R.F.; Kopel, M.J.; Fish, S.I.; Tucker, B.M. Efficacy of an attachable subcutaneous cuff for the prevention of intravascular catheter-related infection. A randomized controlled trial. JAMA 1989, 261, 878–883. [Google Scholar]
- Bach, A.H.; Eberhardt, A.; Frick, H.; Schmidt, B.W.; Bottinger; Martin, E. Efficacy of silver-coating central venous catheters in reducing bacterial colonization. Crit. Care Med. 1999, 27, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Donelli, G.; Francolini, I.; Piozzi, R.; di Rosa Marconi, W. New polymer-antibiotic systems to inhibit bacterial biofilm formation: A suitable approach to prevent central venous catheter-associated infections. J. Chemother. 2002, 14, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Maki, D.G.S.M.; Stolz, S.J.; Wheeler, L.A.; Mermel, L.A. Prevention of central-venous catheter-related bloodstream infection by use of an antiseptic- impregnated catheter. Ann. Int. Med. 1997, 127, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Marconi, W.A.; Galloppa, A.; Martinelli, A.; Piozzi, A. New polyurethane compositions able to bond high amounts of both albumin and heparin. II: Copolymers and polymer blends. Biomaterials 1996, 17, 1795–1802. [Google Scholar]
- Marconi, W.I.; Francolini, A.; Piozzi di Rosa, R. Antibiotic releasing urethane polymers for prevention of catheter related infections. J. Bioact. Compat. Polym. 2001, 16, 393–407. [Google Scholar] [CrossRef]
- Raad, I.R.; Darouiche, R.; Hachem, M.; Mansouri, M.; Bodey, G.P. The broad-spectrum activity and efficacy of catheters coated with minocyclin and rifampin. J. Infect. Dis. 1996, 173, 418–424. [Google Scholar] [CrossRef] [PubMed]
- Schierholz, J.M.; Lucas, L.J.; Rump, A.; Pulverer, G. Efficacy of silver-coated medical devices. J. Hosp. Infect. 1998, 40, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Sherertz, R.J.; Carruth, W.A.; Hampton, A.A.; Byron, M.P.; Solomon, D.D. Efficacy of antibiotic-coated catheters in preventing subcutaneous Staphylococcus aureus infection in rabbits. J. Infect. Dis. 1993, 167, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Tebbs, S.E.; Elliott, T.S.J. A novel antimicrobial central venous catheter impregnated with benzalkonium chloride. J. Antimicrob. Chemother. 1993, 31, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Donelli, G.; Francolini, I. Efficacy of antiadhesive, antibiotic and antiseptic coatings in preventing catheter-related infections: Review. J. Chemother. 2001, 13, 595–606. [Google Scholar] [CrossRef] [PubMed]
- Harvey, A. Strategies for discovering drugs from previously unexplored natural products. Drug Discov. Today 2000, 5, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Turkoglu, A.; Duru, M.E.; Mercan, N.; Kivrak, I.; Gezer, K. Antioxidant and antimicrobial activities of Laetiporus sulphurous (Bull.) Murrill. Food Chem. 2007, 101, 267–273. [Google Scholar]
- Gao, Y.H.; Tang, W.B.; Gao, H.; Chan, E.; Lan, J.; Li, X.; Zhou, S. Antimicrobial activity of the medicinal mushroom Ganoderma. Food Rev. Int. 2005, 21, 211–229. [Google Scholar] [CrossRef]
- Hatvani, N. Antibacterial effect of the culture fluid of Lentinus edodes mycelium grown in submerged liquid culture. Int. J. Antimicrob. Agents 2001, 17, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Alves, M.J.; Ferreira, I.C.F.R.; Martins, A.; Pintado, M. Antimicrobial activity of wild mushrooms extracts against clinical isolates resistant to different antibiotics. J. Appl. Microbiol. 2012, 113, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Alves, M.J.; Ferreira, I.C.F.R.; Froufe, H.J.C.; Abreu, R.M.V.; Martins, A.; Pintado, M. Antimicrobial activity of phenolic compounds identified in wild mushrooms, SAR analysis and docking studies. J. Appl. Microbiol. 2013, 115, 346–357. [Google Scholar] [CrossRef] [PubMed]
- Jagani, S.; Chelikani, R.; Kim, D.-S. Effects of phenol and natural phenolic compounds on biofilm formation by Pseudomonas aeruginosa. Biofouling 2009, 25, 321–324. [Google Scholar] [CrossRef] [PubMed]
- Soković, M.; Ćirić, A.; Glamočlija, J.; Nikolić, M.; van Griensven, J.L.D.L. Agaricus blazei hot water extract shows anti quorum sensing activity in the nosocomial human pathogen Pseudomonas aeruginosa. Molecules 2014, 19, 4189–4199. [Google Scholar]
- Signoretto, C.; Marchi, A.; Bertoncelli, A.; Burlacchini, G.; Papetti, A.; Pruzzo, C.; Zaura, E.; Lingström, P.; Ofek, I.; Pratten, J.; et al. The anti-adhesive mode of action of a purified mushroom (Lentinus edodes) extract with anticaries and antigingivitis properties in two oral bacterial phatogens. BMC Complement. Altern. Med. 2014, 14, 75. [Google Scholar] [CrossRef] [PubMed]
- Francolini, I.; Norris, P.; Piozzi, A.; Donelli, G.; Stoodley, P. Usnic acid, a natural antimicrobial agent able to inhibit bacterial biofilm formation on polymer surfaces. Antimicrob. Agents Chemother. 2004, 48, 4360–4365. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.-F.; Garo, E.; Goering, M.G.; Pasmore, M.; Yoo, H-D.; Esser, T.; Sestrich, J.; Cremin, P.A.; Hough, G.W.; Perrone, P.; et al. Bacterial Biofilm Inhibitors from Diospyros dendo. J. Nat. Prod. 2006, 69, 118–120. [Google Scholar]
- Borges, A.; Saavedra, M.J.; Simões, M. The activity of ferulic and gallic acids in biofilm prevention and controlo of pathogenic bacteria. Biofouling 2012, 7, 755–767. [Google Scholar] [CrossRef]
- Hancock, V.; Dahl, M.; Vejborg, R.M.; Klemm, P. Dietary plant components ellagic acid and tannic acid inhibit biofilm formation by E. coli. J. Med. Microbiol. 2010, 59, 496–498. [Google Scholar] [CrossRef]
- Vikram, A.; Jayaprakasha, G.K.; Jesudhasan, P.R.; Pillai, S.D.; Patil, B.S. Suppression of bacterial cell-cell signaling, biofilm formation and type III secretion system by citrus flavonoids. J. Appl. Microbiol. 2010, 109, 515–527. [Google Scholar] [PubMed]
- Ferreira, I.; Barros, L.; Abreu, R. Antioxidants in wild mushroomns. Curr. Med. Chem. 2009, 12, 1543–1560. [Google Scholar] [CrossRef]
- Tunney, M.M.; Jones, D.S.; Gorman, S.P. Biofilm and biofilm-related encrustations of urinary tract devices. Methods Enzymol. 1999, 310, 558–566. [Google Scholar] [PubMed]
- Huber, B.; Eberl, L.; Feucht, W.; Polster, J. Influence of polyphenols on bacterial biofilm formation and quorumsensing. Z. Naturforschung 2003, 58, 874–884. [Google Scholar]
- Simoes, M.; Bennett, R.N.; Rosa, E.A. Understanding antimicrobial activities of phytochemicals against multidrug resistant bacteria and biofilm. Nat. Prod. Rep. 2009, 26, 746–757. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.N.; Singh, B.R.; Singh, R.L.; Prakash, D.; Dhakarec, R.; Upadhyay, G.; Singh, H.B. Oxidative DNA damage protective activity, antioxidant and anti-quorum sensing potentials of Moringa oleifera. Food Chem. Toxicol. 2009, 47, 1109–1116. [Google Scholar] [CrossRef] [PubMed]
- Barros, L.; Venturini, B.; Baptista, P.; Estevinho, L.; Ferreira, I.C.F.R. Chemical composition and biological properties of Portuguese wild mushrooms: A comprehensive study. J. Agric. Food Chem. 2008, 56, 3856–3862. [Google Scholar] [CrossRef] [PubMed]
- Barros, L.; Dueñas, M.; Ferreira, I.C.F.R.; Baptista, P.; Santos-Buelga, C. Phenolic acids determination by HPLC-DAD-ESI/MS in sixteen different Portuguese wild mushrooms species. Food Chem. Toxicol. 2009, 47, 1076–1079. [Google Scholar] [CrossRef] [PubMed]
- Heleno, S.A.; Barros, L.; Sousa, M.J.; Martins, A.; Ferreira, I.C.F.R. Tocopherols composition of Portuguese wild mushrooms with antioxidant capacity. Food Chem. 2010, 119, 1443–1450. [Google Scholar] [CrossRef]
- Yaltirak, T.; Aslim, B.; Ozturk, S.; Alli, H. Antimicrobial and antioxidant activities of Russula delica. Food Chem. Toxicol. 2009, 47, 2052–2056. [Google Scholar] [CrossRef] [PubMed]
- Performance Standards for Antimicrobial Susceptibilitty Testing; CLSI Document M100-S18; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2008.
- Comité de L’Antibiogramme De La Société Française de Microbiologie; Communiqué 2008 (Edition de Janvier 2008); Société Française de Microbiologie: Paris, France, 2008. (In French)
- Stepanovic, S.; Vukovic, D.; Dakic, I.; Savic, B.; Švabic-Vlahovic, M. A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar]
- Abreu, R.M.V.; Ferreira, I.C.F.R.; Calhelha, R.C.; Lima, R.T.; Vasconcelos, M.H.; Adega, F.; Chaves, R.; Queiroz, M.J.R.P. Anti-hepatocellular carcinoma activity using human HepG2 cells and hepatotoxicity of 6-substituted methyl 3 aminothieno[3,2-b]pyridine-2-carboxylate derivatives: In vitro evaluation, cell cycle analysisand QSAR studies. Eur. J. Med. Chem. 2011, 46, 5800–5806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Alves, M.J.; Ferreira, I.C.F.R.; Lourenço, I.; Costa, E.; Martins, A.; Pintado, M. Wild Mushroom Extracts as Inhibitors of Bacterial Biofilm Formation. Pathogens 2014, 3, 667-679. https://doi.org/10.3390/pathogens3030667
Alves MJ, Ferreira ICFR, Lourenço I, Costa E, Martins A, Pintado M. Wild Mushroom Extracts as Inhibitors of Bacterial Biofilm Formation. Pathogens. 2014; 3(3):667-679. https://doi.org/10.3390/pathogens3030667
Chicago/Turabian StyleAlves, Maria José, Isabel C. F. R. Ferreira, Inês Lourenço, Eduardo Costa, Anabela Martins, and Manuela Pintado. 2014. "Wild Mushroom Extracts as Inhibitors of Bacterial Biofilm Formation" Pathogens 3, no. 3: 667-679. https://doi.org/10.3390/pathogens3030667