
Asymtomatic Bacteriuria as a Model to Study the Coevolution of Hosts and Bacteria


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Urinary Tract Infection (UTI)
3. Asymptomatic Bacteriuria (ABU) Is Protective

4. Results and Discussion
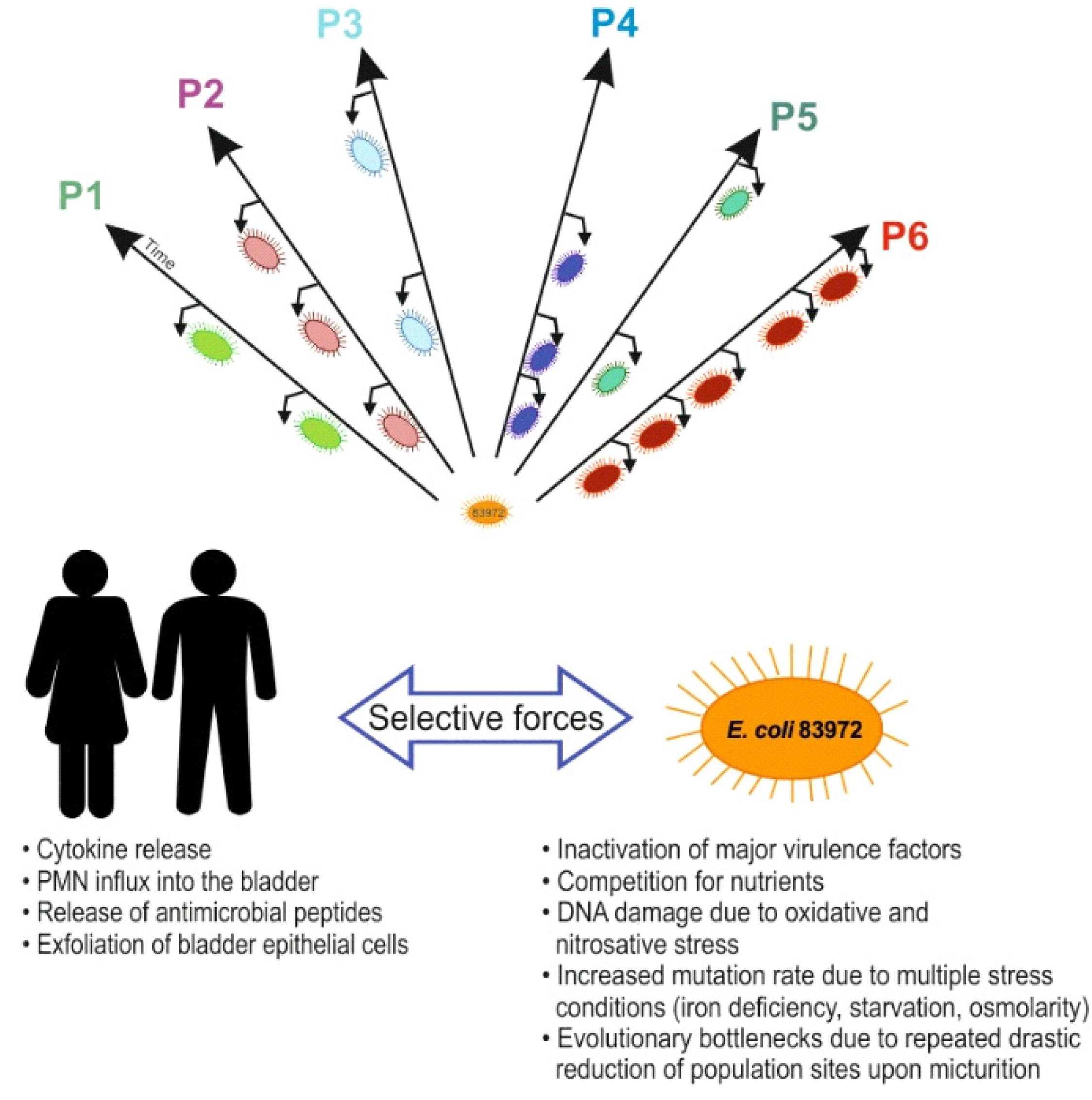
4.1. Bacterial Evolution towards Commensalism rather than Virulence
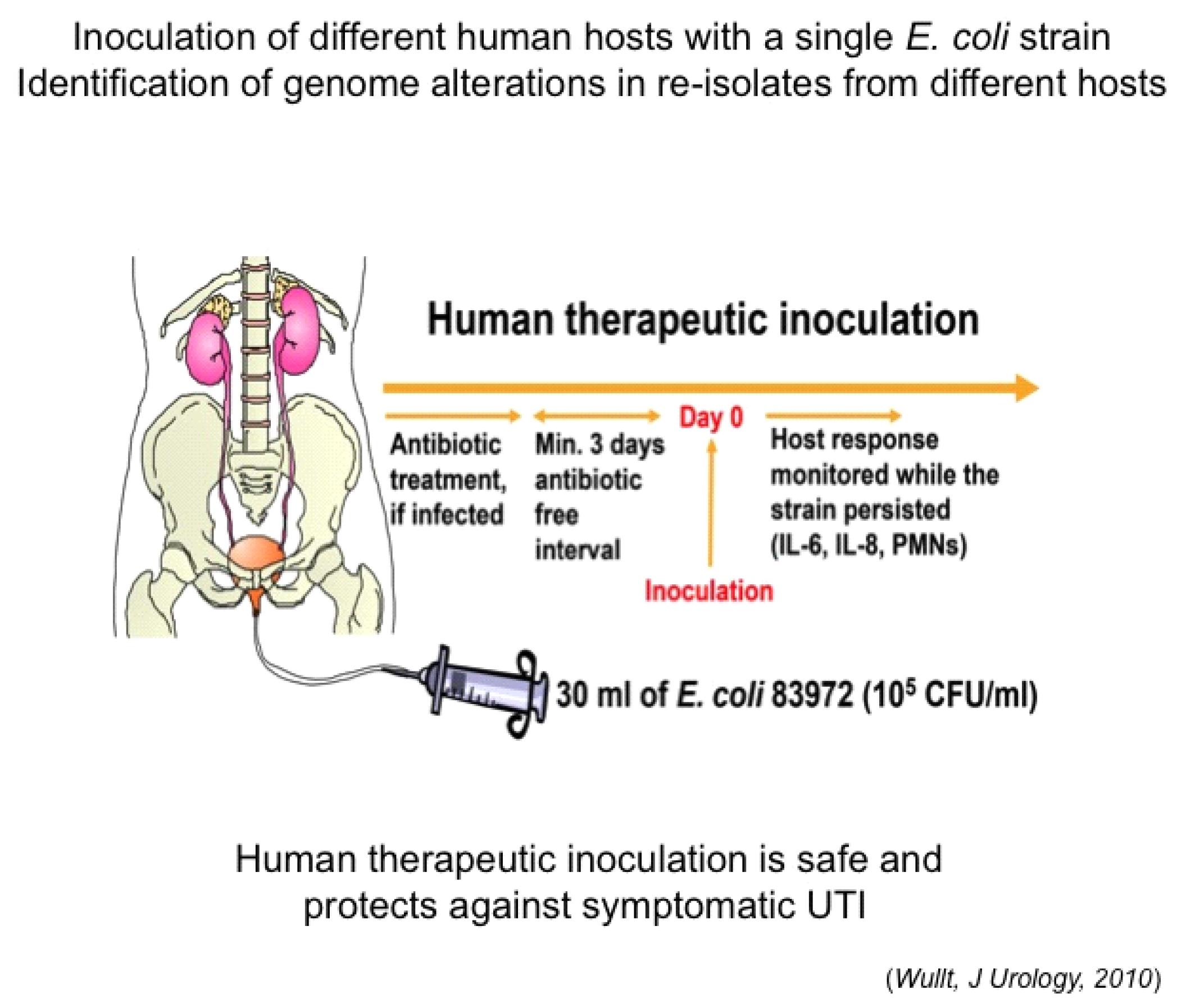
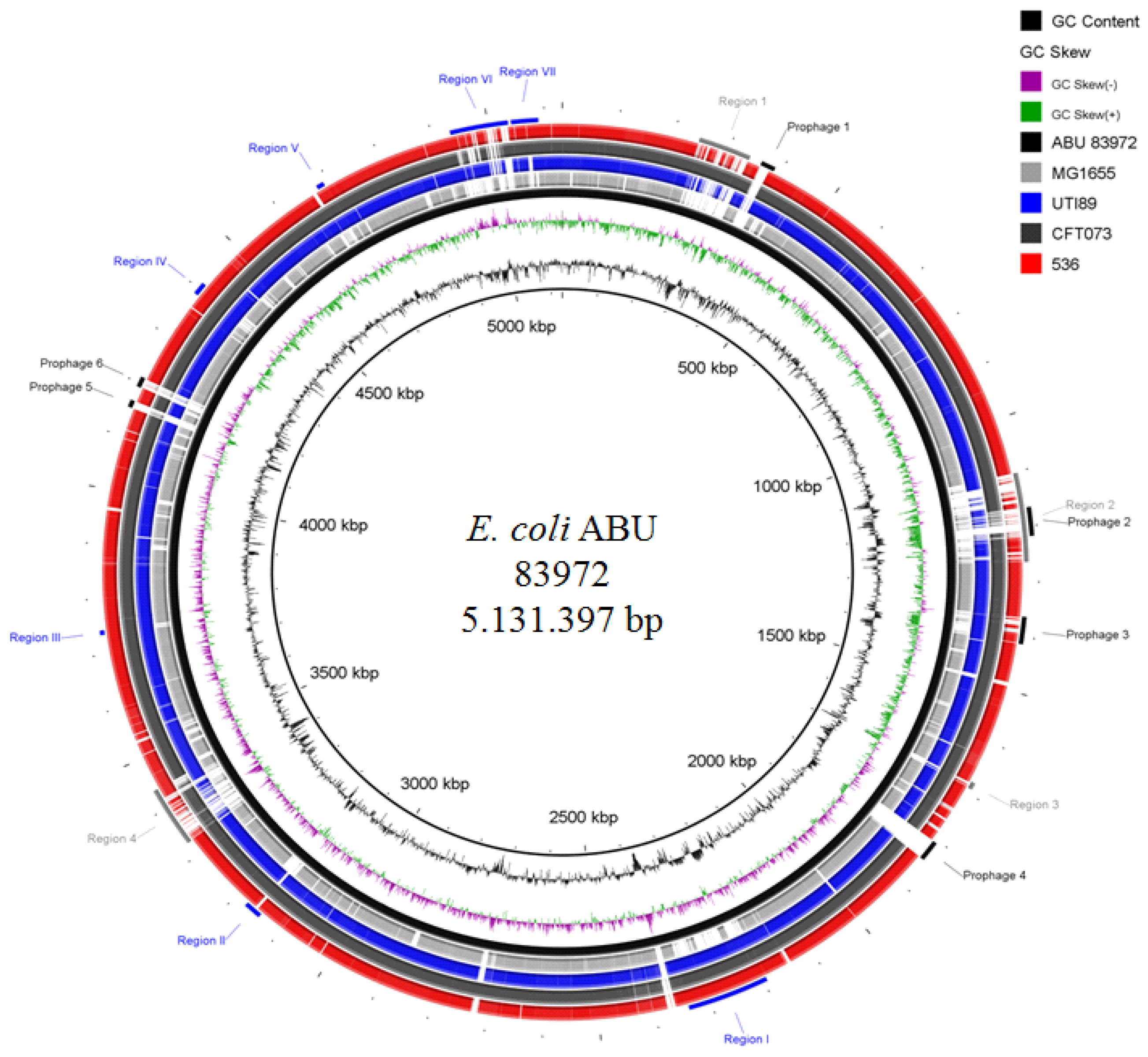
4.2. Host-Specific Genome Alterations in Inoculated Hosts


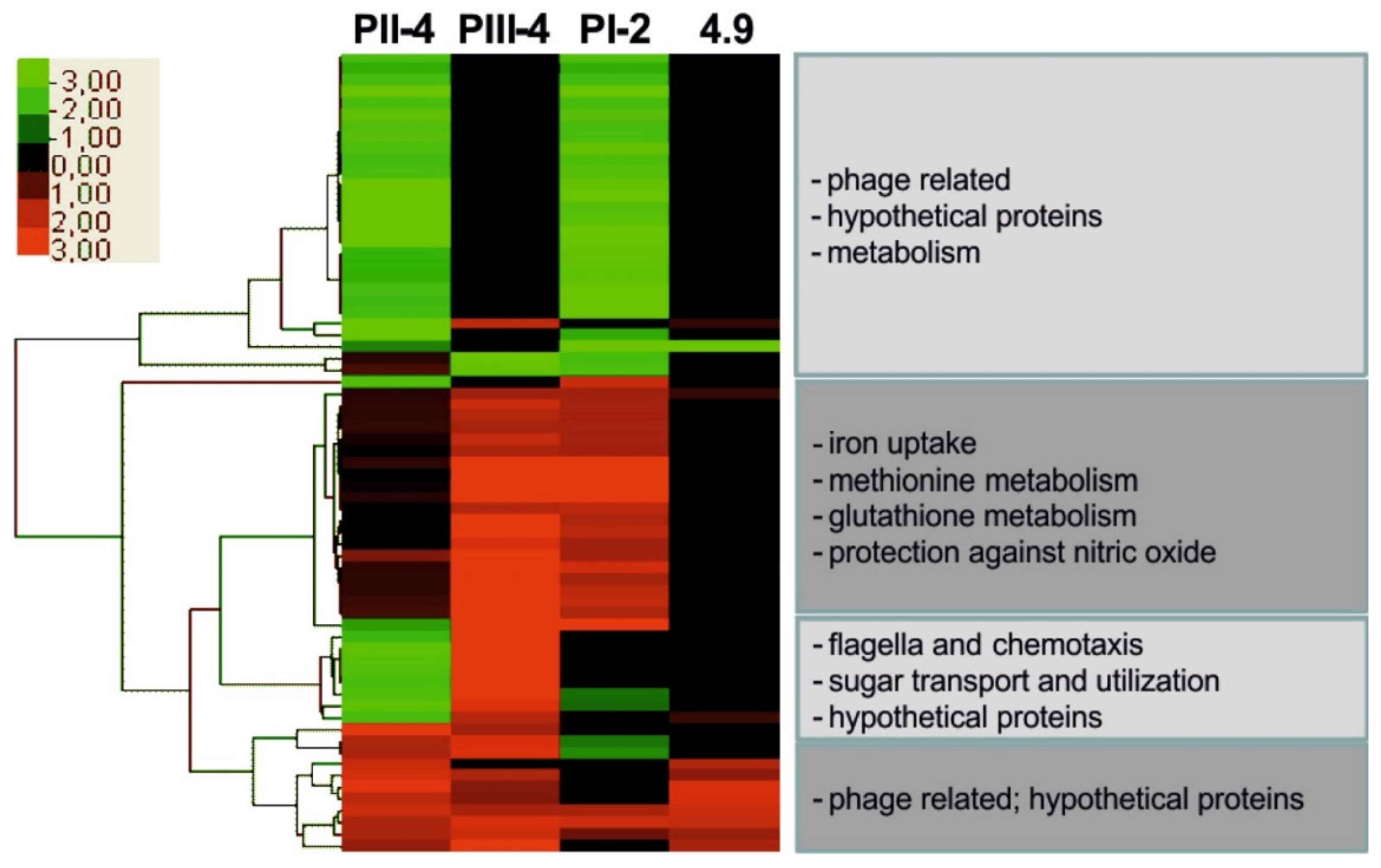
4.3. Host-Specific Gene Expression Levels and Phenotypic Variation in Inoculated Hosts

Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lutay, N.; Ambite, I.; Grönberg Hernandez, J.; Rydstrom, G.; Ragnarsdottir, B.; Puthia, M.; Nadeem, A.; Zhang, J.; Storm, P.; Dobrindt, U.; et al. Bacterial control of host gene expression through RNA polymerase II. J. Clin. Investig. 2013, 123, 2366–2379. [Google Scholar] [CrossRef] [PubMed]
- Ragnarsdottir, B.; Lutay, N.; Grönberg-Hernandez, J.; Köves, B.; Svanborg, C. Genetics of innate immunity and UTI susceptibility. Nat. Rev. Urol. 2011, 8, 449–468. [Google Scholar] [CrossRef] [PubMed]
- Fischer, H.; Lutay, N.; Ragnarsdottir, B.; Yadav, M.; Jonsson, K.; Urbano, A.; Al Hadad, A.; Ramisch, S.; Storm, P.; Dobrindt, U.; et al. Pathogen specific, IRF3-dependent signaling and innate resistance to human kidney infection. PLoS Pathog. 2010, 6, e1001109. [Google Scholar] [CrossRef] [PubMed]
- Jaillon, S.; Moalli, F.; Ragnarsdottir, B.; Bonavita, E.; Puthia, M.; Riva, F.; Barbati, E.; Nebuloni, M.; Cvetko Krajinovic, L.; Markotic, A.; et al. The humoral pattern recognition molecule PTX3 is a key component of innate immunity against urinary tract infection. Immunity 2014, 40, 621–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ipe, D.S.; Sundac, L.; Benjamin, W.H., Jr.; Moore, K.H.; Ulett, G.C. Asymptomatic bacteriuria: Prevalence rates of causal microorganisms, etiology of infection in different patient populations, and recent advances in molecular detection. FEMS Microbiol. Lett. 2013, 346, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Lindberg, U.; Claesson, I.; Hanson, L.A.; Jodal, U. Asymptomatic bacteriuria in schoolgirls. VIII Clinical course during a 3-year follow-up. J. Pediatr. 1978, 92, 194–199. [Google Scholar] [CrossRef]
- Sundén, F.; Hakansson, L.; Ljunggren, E.; Wullt, B. Escherichia coli 83972 bacteriuria protects against recurrent lower urinary tract infections in patients with incomplete bladder emptying. J. Urol. 2010, 184, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Andersson, P.; Engberg, I.; Lidin-Janson, G.; Lincoln, K.; Hull, R.; Hull, S.; Svanborg, C. Persistence of Escherichia coli bacteriuria is not determined by bacterial adherence. Infect. Immun. 1991, 59, 2915–2921. [Google Scholar] [PubMed]
- Wullt, B.; Bergsten, G.; Connell, H.; Rollano, P.; Gebretsadik, N.; Hull, R.; Svanborg, C. P fimbriae enhance the early establishment of Escherichia coli in the human urinary tract. Mol. Microbiol. 2000, 38, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Zdziarski, J.; Brzuszkiewicz, E.; Wullt, B.; Liesegang, H.; Biran, D.; Voigt, B.; Grönberg-Hernandez, J.; Ragnarsdottir, B.; Hecker, M.; Ron, E.Z.; et al. Host imprints on bacterial genomes—rapid, divergent evolution in individual patients. PLoS Pathog. 2010, 6, e1001078. [Google Scholar] [CrossRef] [PubMed]
- Zdziarski, J.; Svanborg, C.; Wullt, B.; Hacker, J.; Dobrindt, U. Molecular basis of commensalism in the urinary tract: Low virulence or virulence attenuation? Infect. Immun. 2008, 76, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Beatson, S.A.; Ben Zakour, N.L.; Totsika, M.; Forde, B.M.; Watts, R.E.; Mabbett, A.N.; Szubert, J.M.; Sarkar, S.; Phan, M.D.; Peters, K.M.; et al. Molecular analysis of asymptomatic bacteriuria Escherichia coli strain VR50 reveals adaptation to the urinary tract by gene acquisition. Infect. Immun. 2015, 83, 1749–1764. [Google Scholar] [CrossRef] [PubMed]
- Bergsten, G.; Wullt, B.; Svanborg, C. Escherichia coli, fimbriae, bacterial persistence and host response induction in the human urinary tract. Int. J. Med. Microbiol. 2005, 295, 487–502. [Google Scholar] [CrossRef] [PubMed]
- Eden, C.S.; Hanson, L.A.; Jodal, U.; Lindberg, U.; Akerlund, A.S. Variable adherence to normal human urinary-tract epithelial cells of Escherichia coli strains associated with various forms of urinary-tract infection. Lancet 1976, 1, 490–492. [Google Scholar] [PubMed]
- Plos, K.; Connell, H.; Jodal, U.; Marklund, B.I.; Marild, S.; Wettergren, B.; Svanborg, C. Intestinal carriage of P fimbriated Escherichia coli and the susceptibility to urinary tract infection in young children. J. Infect. Dis. 1995, 171, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Klemm, P.; Roos, V.; Ulett, G.C.; Svanborg, C.; Schembri, M.A. Molecular characterization of the Escherichia coli asymptomatic bacteriuria strain 83972: The taming of a pathogen. Infect. Immun. 2006, 74, 781–785. [Google Scholar] [CrossRef] [PubMed]
- Salvador, E.; Wagenlehner, F.; Köhler, C.D.; Mellmann, A.; Hacker, J.; Svanborg, C.; Dobrindt, U. Comparison of asymptomatic bacteriuria Escherichia coli isolates from healthy individuals versus those from hospital patients shows that long-term bladder colonization selects for attenuated virulence phenotypes. Infect. Immun. 2012, 80, 668–678. [Google Scholar] [CrossRef] [PubMed]
- Grönberg-Hernández, J.; Sundén, F.; Connolly, J.; Svanborg, C.; Wullt, B. Genetic Control of the Variable Innate Immune Response to Asymptomatic Bacteriuria. PLoS ONE 2011, 6, e28289. [Google Scholar]
- Köves, B.; Salvador, E.; Grönberg-Hernandez, J.; Zdziarski, J.; Wullt, B.; Svanborg, C.; Dobrindt, U. Rare emergence of symptoms during long-term asymptomatic Escherichia coli 83972 carriage without an altered virulence factor repertoire. J. Urol. 2014, 191, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Ipe, D.S.; Horton, E.; Ulett, G.C. The basics of bacteriuria: Strategies of microbes for persistence in urine. Front. Cell. Infect. Microbiol. 2016. [Google Scholar] [CrossRef]
- Smits, W.K.; Kuipers, O.P.; Veening, J.W. Phenotypic variation in bacteria: The role of feedback regulation. Nat. Rev. Microbiol. 2006, 4, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Chai, Y.; Chu, F.; Kolter, R.; Losick, R. Bistability and biofilm formation in Bacillus subtilis. Mol. Microbiol. 2008, 67, 254–263. [Google Scholar] [CrossRef] [PubMed]
- Cameron, A.; Frirdich, E.; Huynh, S.; Parker, C.T.; Gaynor, E.C. Hyperosmotic stress response of Campylobacter jejuni. J. Bacteriol. 2012, 194, 6116–6130. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dobrindt, U.; Wullt, B.; Svanborg, C. Asymtomatic Bacteriuria as a Model to Study the Coevolution of Hosts and Bacteria. Pathogens 2016, 5, 21. https://doi.org/10.3390/pathogens5010021
Dobrindt U, Wullt B, Svanborg C. Asymtomatic Bacteriuria as a Model to Study the Coevolution of Hosts and Bacteria. Pathogens. 2016; 5(1):21. https://doi.org/10.3390/pathogens5010021
Chicago/Turabian StyleDobrindt, Ulrich, Björn Wullt, and Catharina Svanborg. 2016. "Asymtomatic Bacteriuria as a Model to Study the Coevolution of Hosts and Bacteria" Pathogens 5, no. 1: 21. https://doi.org/10.3390/pathogens5010021