HPV16 E6 and E7 Upregulate Interferon-Induced Antiviral Response Genes ISG15 and IFIT1 in Human Trophoblast Cells

Abstract
:1. Introduction
2. Results
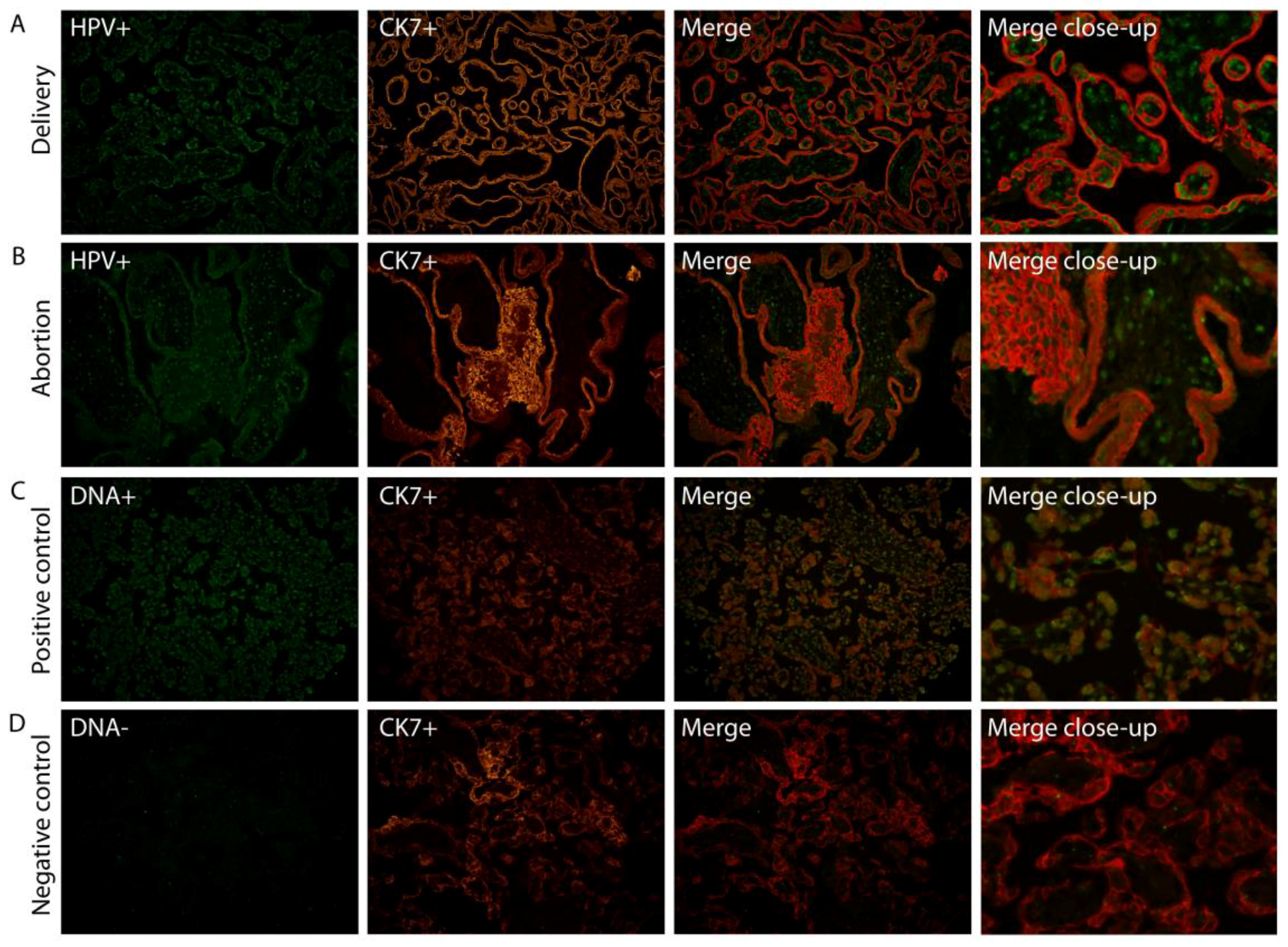
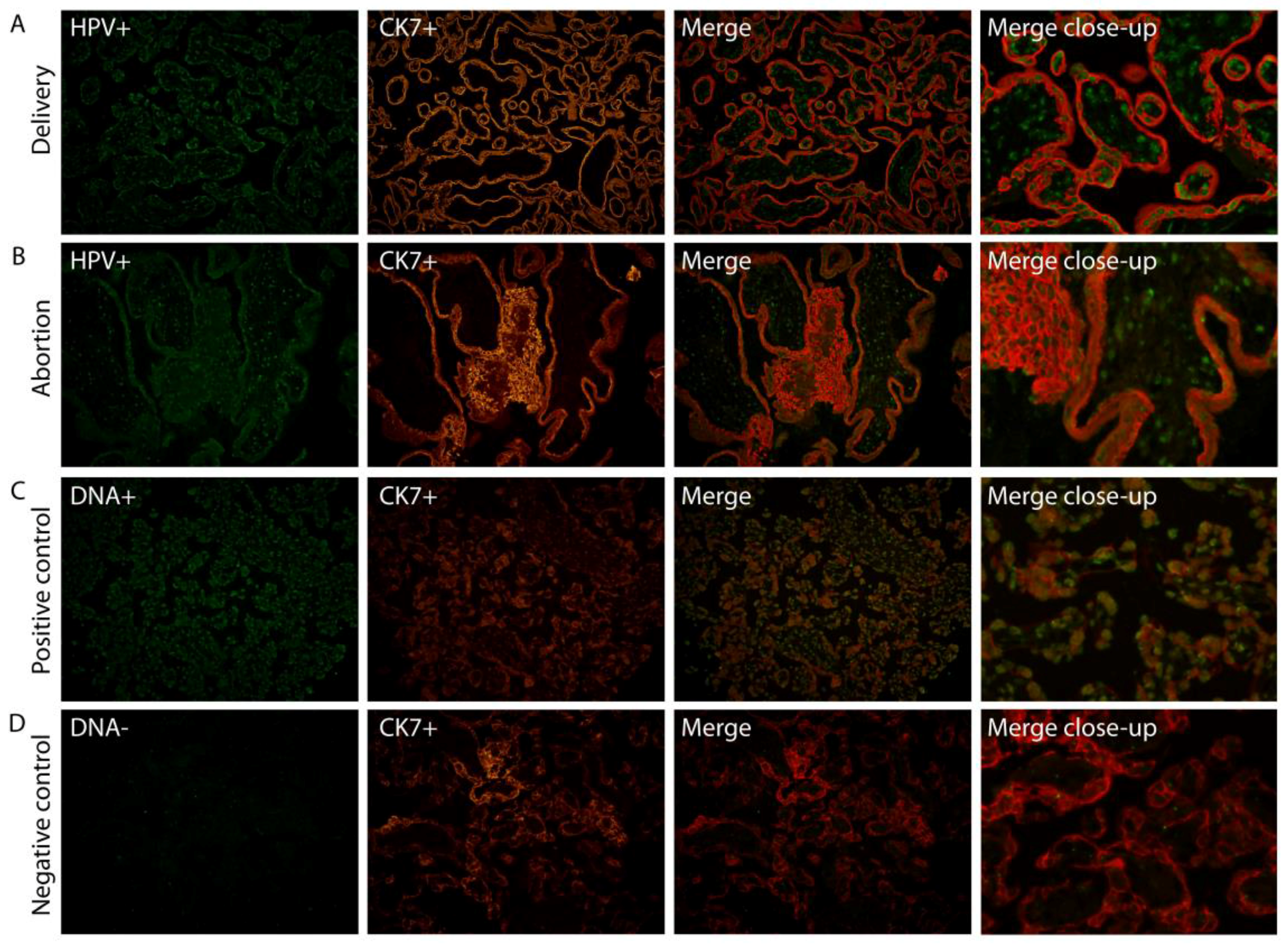
2.1. HPV Localizes in Syncytiotrophoblast Cells of the Placenta
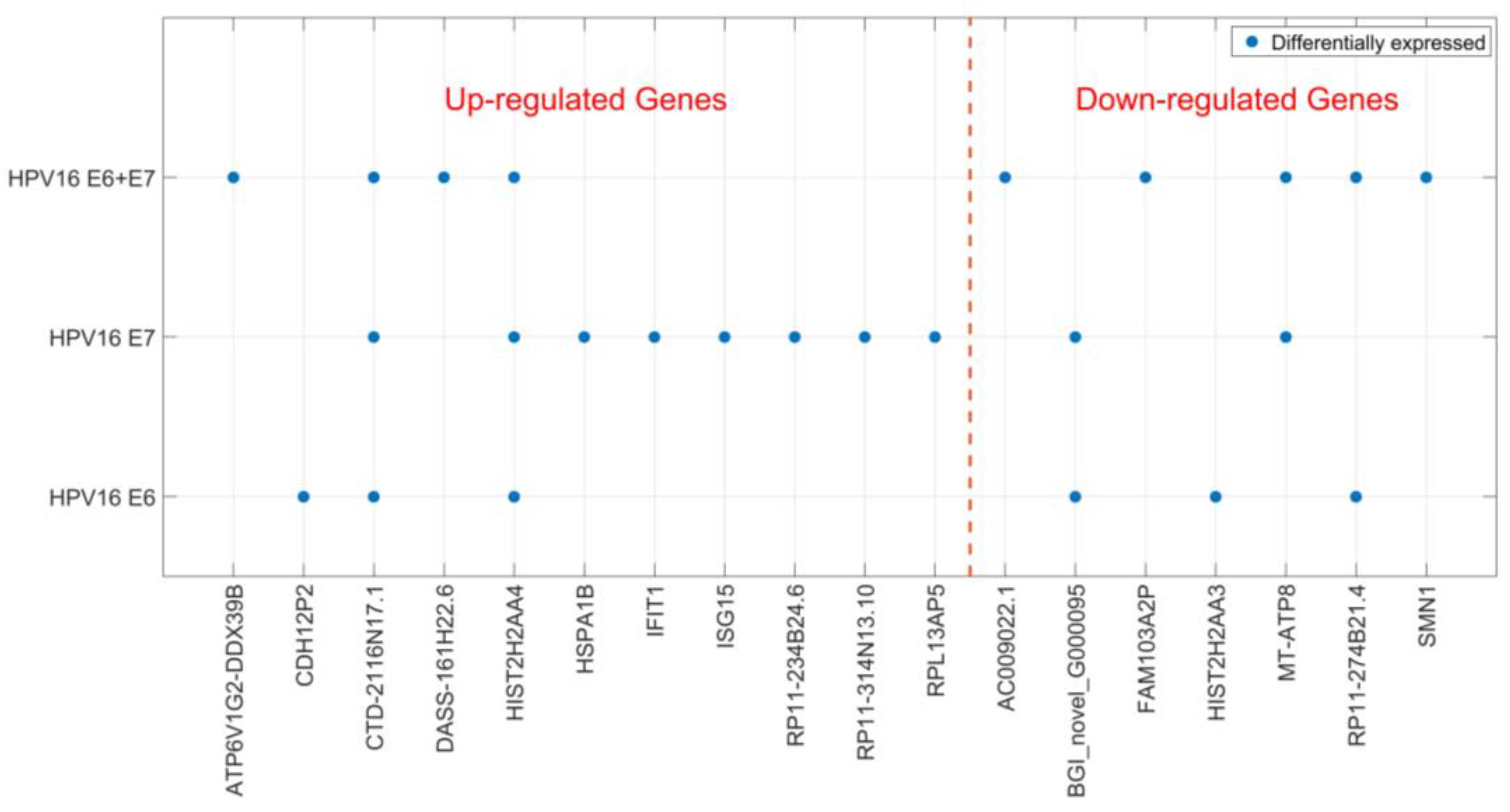
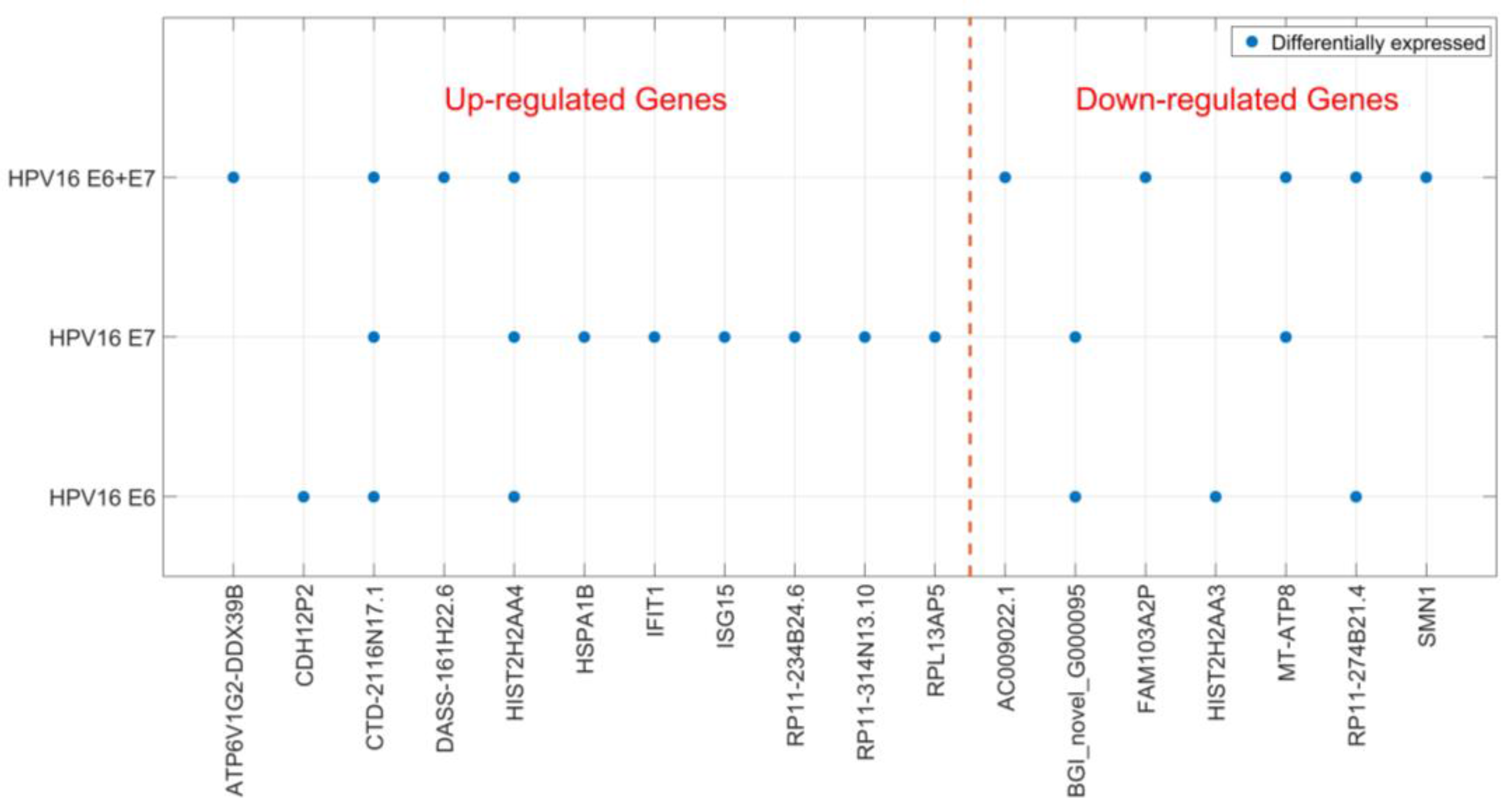
2.2. Gene Expression of JAR Cells is Affected by HPV16 E6 and E7 Expression
2.3. HPV16 E6 and E7 are not Transcribed in HPV16-Positive Placenta
3. Discussion
4. Materials and Methods
4.1. Placental Material and HPV Screening
4.2. Fluorescence in Situ Hybridization (FISH)/Fluorescence Immunohistochemistry (FIHC)
4.3. Cell Culture, Transfection and RNA Extraction
4.4. Quantitative Reverse Transcription PCR
- GAPDH, 5′-GACAGTCAGCCGCATCTTCT-3′, 5′-TTAAAAGCAGCCCTGGTGAC-3′;
- HPV16E6, 5′-CTGCAATGTTTCAGGACCCAC-3′, 5′-GTTGTTTGCAGCTCTGTGCAT-3′;
- HPV16E7, 5′-AGAACCGGACAGAGCCCATTA-3′, 5′-CGCACAACCGAAGCGTAGA-3′
4.5. RNA Sequencing and DEG Analyses
4.6. Ethical Approval
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zur Hausen, H. Papillomaviruses and cancer: From basic studies to clinical application. Nat. Rev. Cancer. 2002, 2, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Pfister, H. Biology and biochemistry of papillomaviruses. In Reviews of Physiology, Biochemistry and Pharmacology; Reviews of Physiology, Biochemistry and Pharmacology; Springer: Berlin/Heidelberg, Germany, 1984; Volume 99, pp. 111–181. [Google Scholar]
- Hermonat, P.L.; Han, L.; Wendel, P.J.; Quirk, J.G.; Stern, S.; Lowery, C.L.; Rechtin, T.M. Human papillomavirus is more prevalent in first trimester spontaneously aborted products of conception compared to elective specimens. Virus Gen. 1997, 14, 13–17. [Google Scholar]
- Gomez, L.M.; Ma, Y.; Ho, C.; McGrath, C.M.; Nelson, D.B.; Parry, S. Placental infection with human papillomavirus is associated with spontaneous preterm delivery. Hum. Reprod. 2008, 23, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Worda, C.; Huber, A.; Hudelist, G. Prevalence of cervical and intrauterine human papillomavirus infected in the third trimester in asymptomatic women. J. Soc. Gynecol. Investig. 2005, 12, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Armbruster-Moraes, E.; Ioshimoto, L.M.; Leao, E.; Zugaib, M. Detection of human papillomavirus deoxyribonucleic acid sequences in amniotic fluid during different periods of pregnancy. Am. J. Obstet. Gynecol. 1993, 169, 1074. [Google Scholar] [CrossRef]
- Burguete, T.; Fontanges-Darriet, M.; Roset, E.; Hager, H.; Ko, A.; Bischof, P. Evidence for infection of the human embryo with adeno-associated virus in pregnancy. Hum. Reprod. 1999, 14, 2396–2401. [Google Scholar] [CrossRef] [PubMed]
- Tseng, C.J.; Lin, C.Y.; Wang, R.L.; Chen, L.J.; Chang, Y.L.; Hsieh, T.T.; Pao, C.C. Possible transplacental transmission of human papillomaviruses. Am. J. Obstet. Gynecol. 1992, 166, 35–40. [Google Scholar] [CrossRef]
- Wang, X.; Zhu, Q.; Rao, H. Maternal-fetal transmission of human papillomavirus. Chin. Med. J. (Engl.) 1998, 111, 726–727. [Google Scholar] [PubMed]
- Hermonat, P.L.; Kechelava, S.; Lowery, C.L.; Korourian, S. Trophoblasts are the preferential target for human papilloma virus infection in spontaneously aborted products of conception. Hum. Pathol. 1998, 29, 170–174. [Google Scholar] [CrossRef]
- Liu, Y.; You, H.; Chiriva-Internati, M.; Korourian, S.; Lowery, C.L.; Carey, M.J.; Smith, C.V.; Hermonat, P.L. Display of complete life cycle of human papillomavirus type 16 in cultured placental trophoblasts. Virology 2001, 290, 99–105. [Google Scholar] [CrossRef] [PubMed]
- You, H.; Liu, Y.; Agrawal, N.; Prasad, C.K.; Chiriva-Internati, M.; Lowery, C.L.; Kay, H.H.; Hermonat, P.L. Infection, replication, and cytopathology of human papillomavirus type 31 in trophoblasts. Virology 2003, 316, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Stanley, M.A. Epithelial cell responses to infection with human papillomavirus. Clin. Microbiol. Rev. 2012, 25, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Ghittoni, R.; Accardi, R.; Hasan, U.; Gheit, T.; Sylla, B.; Tommasino, M. The biological properties of E6 and E7 oncoproteins from human papillomaviruses. Virus Genes. 2010, 40, 1–13. [Google Scholar] [CrossRef] [PubMed]
- You, H.; Liu, Y.; Carey, M.J.; Lowery, C.L.; Hermonat, P.L. Defective 3A trophoblast-endometrial cell adhesion and altered 3A growth and survival by human papillomavirus type 16 oncogenes. Mol. Cancer Res. 2002, 1, 25–31. [Google Scholar] [PubMed]
- Ambühl, L.M.M.; Baandrup, U.; Dybkær, K.; Blaakær, J.; Uldbjerg, N.; Sørensen, S. Human Papillomavirus Infection as a Possible Cause of Spontaneous Abortion and Spontaneous Preterm Delivery. Infect. Dis. Obstet. Gynecol. 2016, 2016, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Z.; Goel, S.; Carter, J.E. Association of cervical cytology and HPV DNA status during pregnancy with placental abnormalities and preterm birth. Am. J. Clin. Pathol. 2011, 136, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Sifakis, S.; Ergazaki, M.; Sourvinos, G.; Koffa, M.; Koumantakis, E.; Spandidos, D. Evaluation of Parvo B19, CMV and HPV viruses in human aborted material using the polymerase chain reaction technique. Eur. J. Obstet. Gynecol. Reprod. Biol. 1998, 76, 169–173. [Google Scholar] [CrossRef]
- Ambühl, L.M.M.; Leonhard, A.K.; Zakhary, C.W.; Jørgensen, A.; Blaakær, J.; Dybkær, K.; Baandrup, U.; Uldbjerg, N.; Sørensen, S. Human Papillomavirus infects placental trophoblast and Hofbauer cells, but appears not to play a causal role in miscarriage and preterm labor. Acta Obstet. Gynecol. Scand. 2017, in press. [Google Scholar]
- Tarazona, S.; Garcia-Alcalde, F.; Dopazo, J.; Ferrer, A.; Conesa, A. Differential expression in RNA-seq: A matter of depth. Genome Res. 2011, 21, 2213–2223. [Google Scholar] [CrossRef] [PubMed]
- Terenzi, F.; Saikia, P.; Sen, G.C. Interferon-inducible protein, P56, inhibits HPV DNA replication by binding to the viral protein E1. EMBO J. 2008, 27, 3311–3321. [Google Scholar] [CrossRef] [PubMed]
- Jeon, Y.J.; Yoo, H.M.; Chung, C.H. ISG15 and immune diseases. Biochim. Biophys. Acta 2010, 1802, 485–496. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.E.; Laimins, L.A. Microarray analysis identifies interferon-inducible genes and Stat-1 as major transcriptional targets of human papillomavirus type 31. J. Virol. 2000, 74, 4174–4182. [Google Scholar] [CrossRef] [PubMed]
- Karstensen, B.; Poppelreuther, S.; Bonin, M.; Walter, M.; Iftner, T.; Stubenrauch, F. Gene expression profiles reveal an upregulation of E2F and downregulation of interferon targets by HPV18 but no changes between keratinocytes with integrated or episomal viral genomes. Virology 2006, 353, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Nees, M.; Geoghegan, J.M.; Hyman, T.; Frank, S.; Miller, L.; Woodworth, C.D. Papillomavirus type 16 oncogenes downregulate expression of interferon-responsive genes and upregulate proliferation-associated and NF-kappaB-responsive genes in cervical keratinocytes. J. Virol. 2001, 75, 4283–4296. [Google Scholar] [CrossRef] [PubMed]
- Sarkola, M.E.; Grénman, S.E.; Rintala, M.M.; Syrjänen, K.J.; Syrjänen, S.M. Human papillomavirus in the placenta and umbilical cord blood. Acta Obstet. Gynecol. Scand. 2008, 87, 1181–1188. [Google Scholar] [CrossRef] [PubMed]
- Slatter, T.L.; Hung, N.G.; Clow, W.M.; Royds, J.A.; Devenish, C.J.; Hung, N.A. A clinicopathological study of episomal papillomavirus infection of the human placenta and pregnancy complications. Mod. Pathol. 2015, 28, 1369–1382. [Google Scholar] [CrossRef] [PubMed]
- Malhomme, O.; Dutheil, N.; Rabreau, M.; Armbruster-Moraes, E.; Schlehofer, R.; Dupressoir, T. Human genital tissues containing DNA of adeno-associated virus lack DNA sequences of the helper viruses adenovirus, herpes simplex virus or cytomegalovirus but frequently contain human papillomavirus DNA. J. Gen. Virol. 1997, 78, 1957–1962. [Google Scholar] [CrossRef] [PubMed]
- Fensterl, V.; Sen, G.C. Interferons and viral infections. BioFactors 2009, 35, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.; Struckhoff, J.J.; Schneider, J.; Martinez-Sobrido, L.; Wolff, T.; García-Sastre, A.; Zhang, D.E.; Lenschow, D.J. Mice lacking the ISG15 E1 enzyme UbE1L demonstrate increased susceptibility to both mouse-adapted and non-mouse-adapted influenza B virus infection. J. Virol. 2009, 83, 1147–1151. [Google Scholar] [CrossRef] [PubMed]
- Lenschow, D.J.; Lai, C.; Frias-Staheli, N.; Giannakopoulos N, V.; Lutz, A.; Wolff, T.; Osiak, A.; Levine, B.; Schmidt, R.E.; García-Sastre, A.; et al. IFN-stimulated gene 15 functions as a critical antiviral molecule against influenza, herpes, and Sindbis viruses. Proc. Natl. Acad. Sci. USA 2007, 104, 1371–1376. [Google Scholar] [CrossRef] [PubMed]
- Cross, J.C.; Werb, Z.; Fisher, S.J. Implantation and the placenta: Key pieces of the development puzzle. Science 1994, 266, 1508–1518. [Google Scholar] [CrossRef] [PubMed]
- Morales, D.J.; Lenschow, D.J. The Antiviral Activities of ISG15. J. Mol. Biol. 2013, 425, 4995–5008. [Google Scholar] [CrossRef] [PubMed]
- Manos, M.M.; Ting, Y.; Wright, D.K.; Lewis, A.J.; Broker, T.R.; Wolinsky, S.M. Use of polymerase chain reaction amplification for the detection of genital human papillomavirus. In Molecular Diagnostics of Human Cancer (Cancer Cells); Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989; Volume 7, pp. 209–214. [Google Scholar]
- De Roda Husman, A.M.; Walboomers, J.M.M.; Van den Brule, A.J.C.; Meijer, C.J.L.M.; Snijders, P.J.F. The use of general primers GP5 and GP6 elongated at their 3’ ends with adjacent highly conserved sequences improves human papillomavirus detection by PCR. J. Gen. Virol. 1995, 76, 1057–1062. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Transfection | Gene ID | Log 2 Fold Change | Probability | Up-/Down-Regulation |
|---|---|---|---|---|
| HPV16 E6 | ||||
| CDH12P2 | 8.2 | 0.885014 | Up | |
| CTD-2116N17.1 | 1.6 | 0.840587 | Up | |
| BGI_novel_G000095 | −1.3 | 0.833708 | Down | |
| HIST2H2AA3 | −4.8 | 0.953578 | Down | |
| RP11-274B21.4 | −7.5 | 0.810089 | Down | |
| HIST2H2AA4 | 2.7 | 0.882313 | Up | |
| HPV16 E7 | ||||
| CTD-2116N17.1 | 2.3 | 0.874182 | Up | |
| RPL13AP5 | 1.2 | 0.808273 | Up | |
| RP11-314N13.10 | 1.1 | 0.809181 | Up | |
| RP11-234B24.6 | 7.6 | 0.832868 | Up | |
| MT-ATP8 | −1.6 | 0.851965 | Down | |
| BGI_novel_G000095 | −1.4 | 0.838509 | Down | |
| IFIT1 | 6.0 | 0.817424 | Up | |
| HIST2H2AA4 | 2.0 | 0.836634 | Up | |
| ISG15 | 1.3 | 0.837189 | Up | |
| HSPA1B | 1.3 | 0.833963 | Up | |
| HPV16 E6 + E7 | ||||
| CTD-2116N17.1 | 2.0 | 0.859646 | Up | |
| MT-ATP8 | −3.2 | 0.916022 | Down | |
| ATP6V1G2-DDX39B | 2.1 | 0.814116 | Up | |
| RP11-274B21.4 | −7.5 | 0.815107 | Down | |
| FAM103A2P | −1.6 | 0.807606 | Down | |
| AC009022.1 | −7.5 | 0.817934 | Down | |
| DASS-161H22.6 | 3.8 | 0.874002 | Up | |
| HIST2H2AA4 | 2.0 | 0.837189 | Up | |
| SMN1 | −1.1 | 0.811964 | Down |
| Gene ID | Transfection | ∆∆Ct | Fold-Change (2−∆∆Ct) | Validation |
|---|---|---|---|---|
| IFIT1 | HPV16 E6 | −3.38 | 10.41 | Up-regulation confirmed |
| HPV16 E7 | −3.72 | 13.18 | ||
| HPV16 E6 + E7 | −2.17 | 4.5 | ||
| ISG15 | HPV16 E6 | −0.75 | 1.68 | Up-regulation confirmed |
| HPV16 E7 | −0.97 | 1.96 | ||
| HPV16 E6 + E7 | −0.2 | 1.15 | ||
| HSPA1B | HPV16 E6 | −0.48 | 1.39 | Up-regulation confirmed |
| HPV16 E7 | −1.21 | 2.31 | ||
| HPV16 E6 + E7 | −0.58 | 1.49 | ||
| SMN1 | HPV16 E6 | 0.21 | 0.86 | Down-regulation confirmed |
| HPV16 E7 | 0.48 | 0.72 | ||
| HPV16 E6 + E7 | 0.4 | 0.76 | ||
| HIST2H2AA4 | HPV16 E6 | −0.09 | 1.06 | Up-regulation NOT confirmed |
| HPV16 E7 | −0.06 | 1.04 | ||
| HPV16 E6 + E7 | 0.08 | 0.95 |
| Gene ID | ∆∆Ct | Fold Change (2−∆∆Ct) | Number of Samples Tested/Included |
|---|---|---|---|
| IFIT1 | 0.065 | 0.96 | 32/16 HPV positives |
| 32/14 HPV negatives | |||
| ISG15 | 0.143 | 0.91 | 32/18 HPV positives |
| 32/16 HPV negatives |
| Sample | Ct GAPDH | ∆Ct HPV16 E6 | ∆Ct HPV16 E7 |
|---|---|---|---|
| SiHa | 13.83 | 5.32 | 4.84 |
| HPV+ patient 1 | 16.2 | - | - |
| HPV+ patient 2 | 15.36 | - | - |
| HPV+ patient 3 | 14.92 | - | - |
| HPV+ patient 4 | 17.39 | - | - |
| HPV+ patient 5 | 15.26 | - | - |
| HPV− patient 1 | 16.31 | - | - |
| HPV− patient 2 | 16.61 | - | - |
| HPV− patient 3 | 14.2 | - | - |
| HPV− patient 4 | 14.7 | - | - |
| HPV− patient 5 | 14.86 | - | - |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ambühl, L.M.M.; Villadsen, A.B.; Baandrup, U.; Dybkær, K.; Sørensen, S. HPV16 E6 and E7 Upregulate Interferon-Induced Antiviral Response Genes ISG15 and IFIT1 in Human Trophoblast Cells. Pathogens 2017, 6, 40. https://doi.org/10.3390/pathogens6030040
Ambühl LMM, Villadsen AB, Baandrup U, Dybkær K, Sørensen S. HPV16 E6 and E7 Upregulate Interferon-Induced Antiviral Response Genes ISG15 and IFIT1 in Human Trophoblast Cells. Pathogens. 2017; 6(3):40. https://doi.org/10.3390/pathogens6030040
Chicago/Turabian StyleAmbühl, Lea M.M., Annemarie B. Villadsen, Ulrik Baandrup, Karen Dybkær, and Suzette Sørensen. 2017. "HPV16 E6 and E7 Upregulate Interferon-Induced Antiviral Response Genes ISG15 and IFIT1 in Human Trophoblast Cells" Pathogens 6, no. 3: 40. https://doi.org/10.3390/pathogens6030040