Characterization of Three Ocular Clinical Isolates of P. aeruginosa: Viability, Biofilm Formation, Adherence, Infectivity, and Effects of Glycyrrhizin

Abstract
:1. Introduction
2. Results
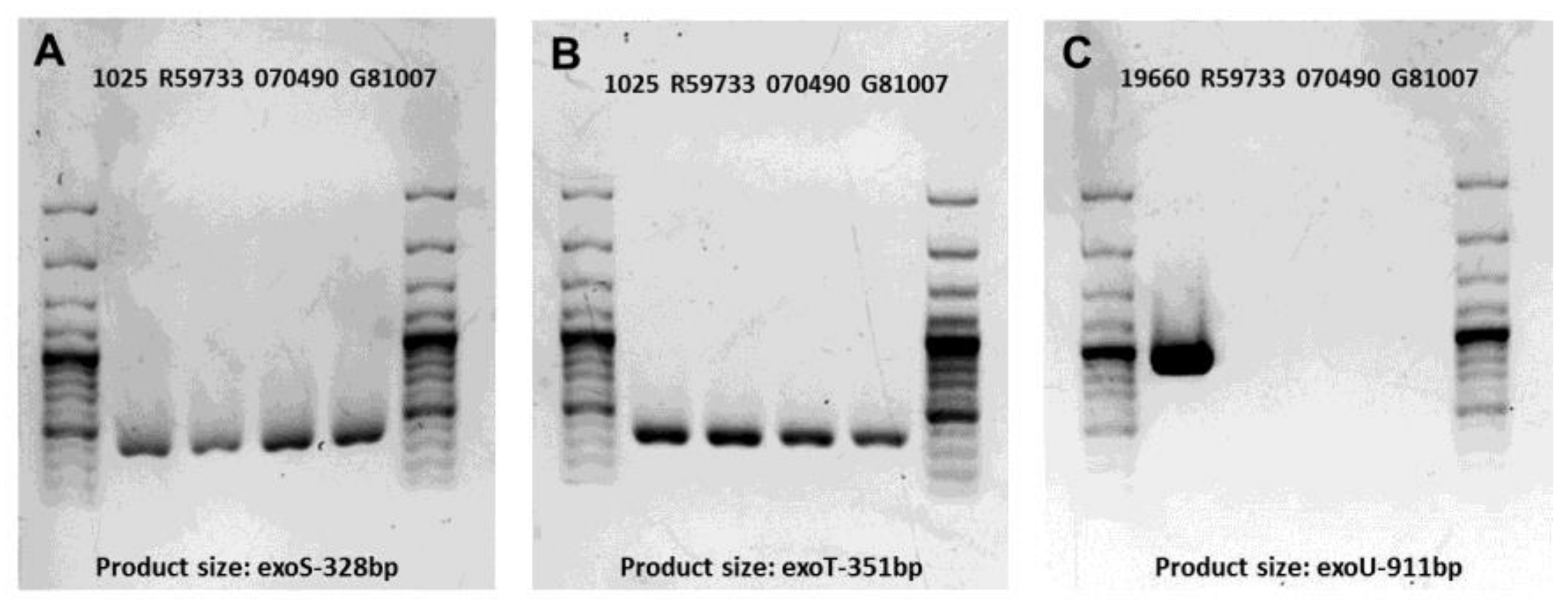
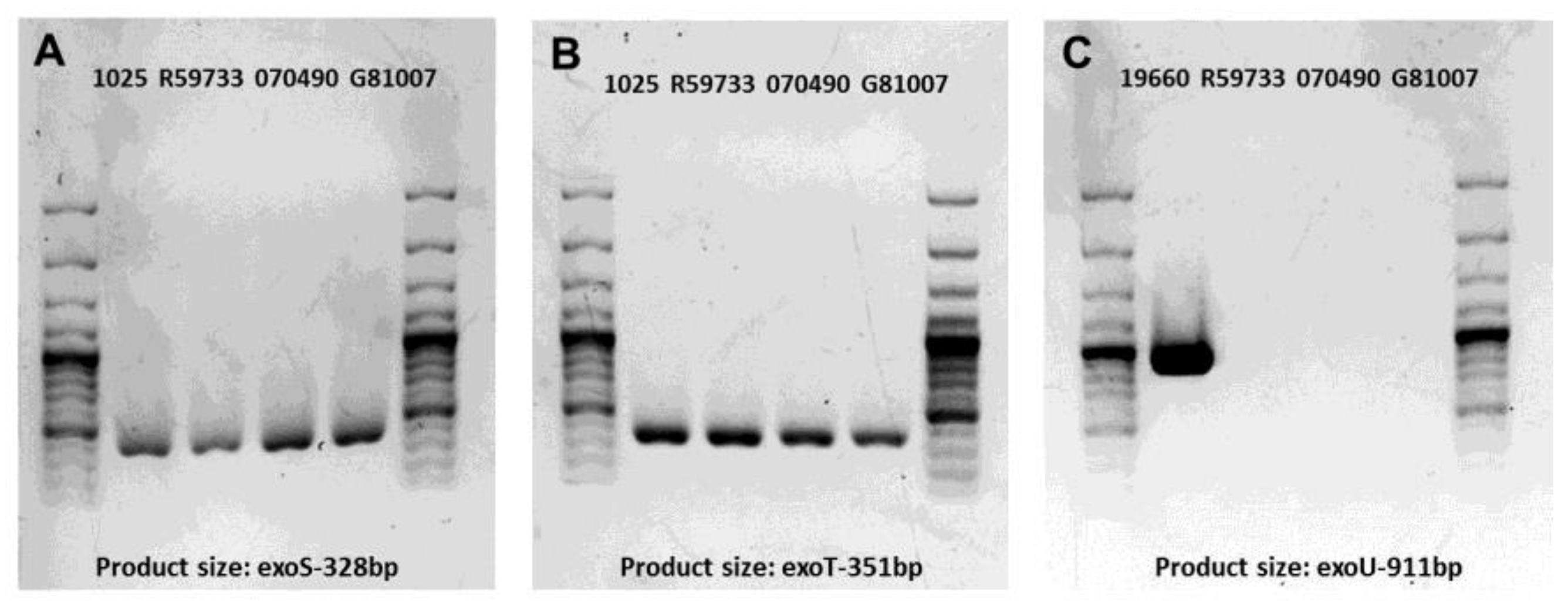
2.1. Gel Electrophoresis of PCR Products for Clinical Isolates
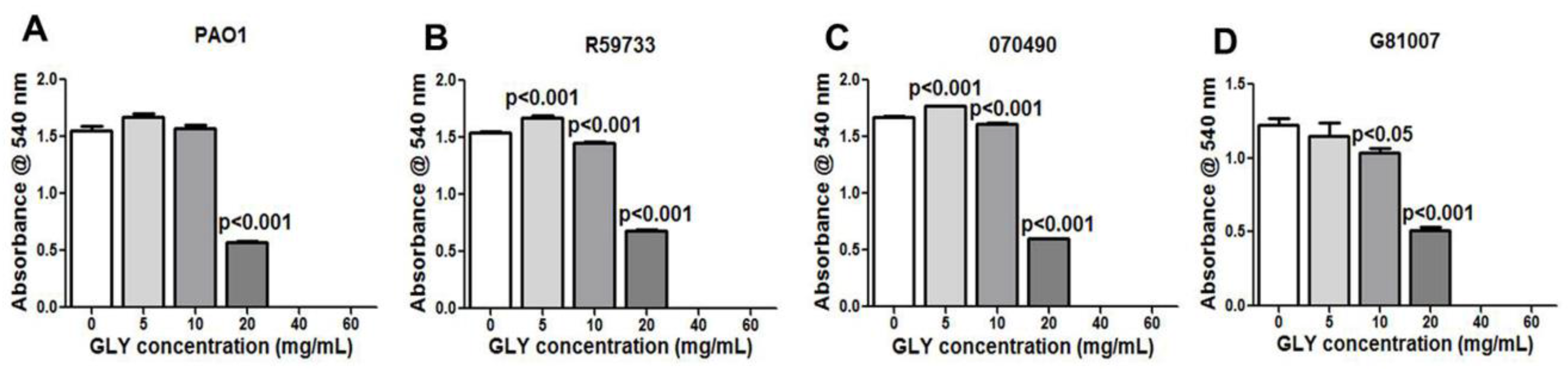
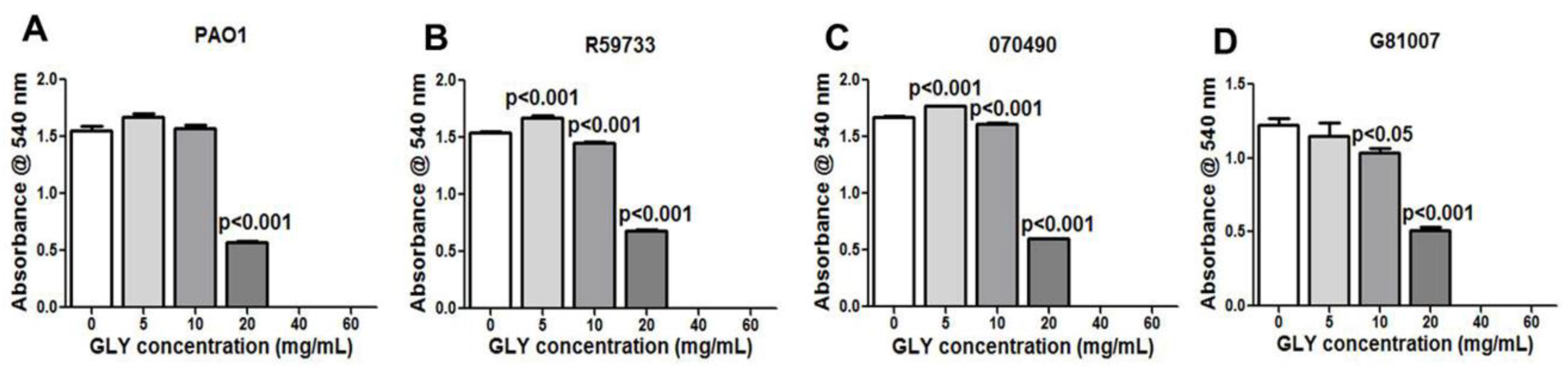
2.2. Minimum Inhibitory Concentration (MIC): Effect of GLY
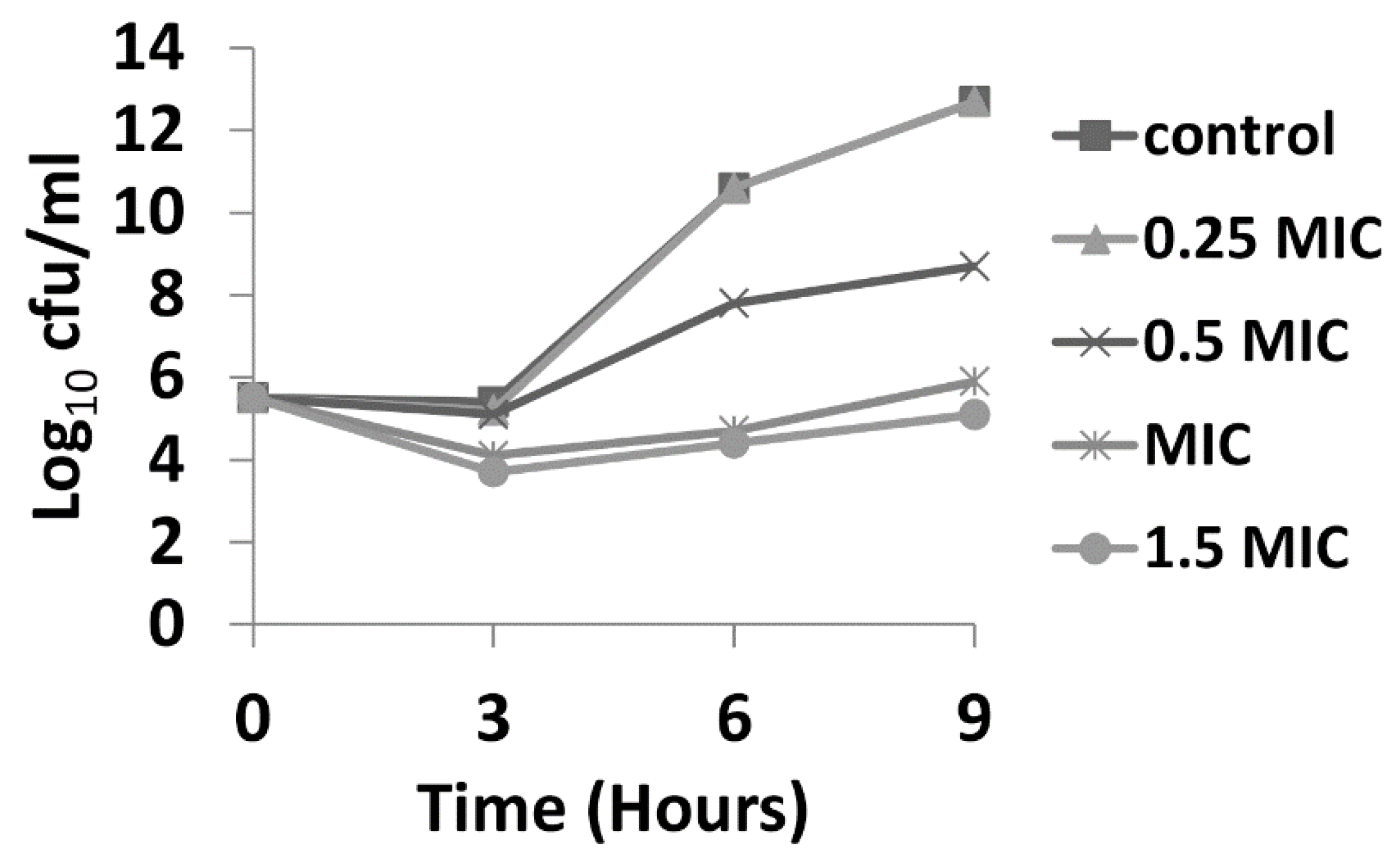
2.3. Time Kill Assay
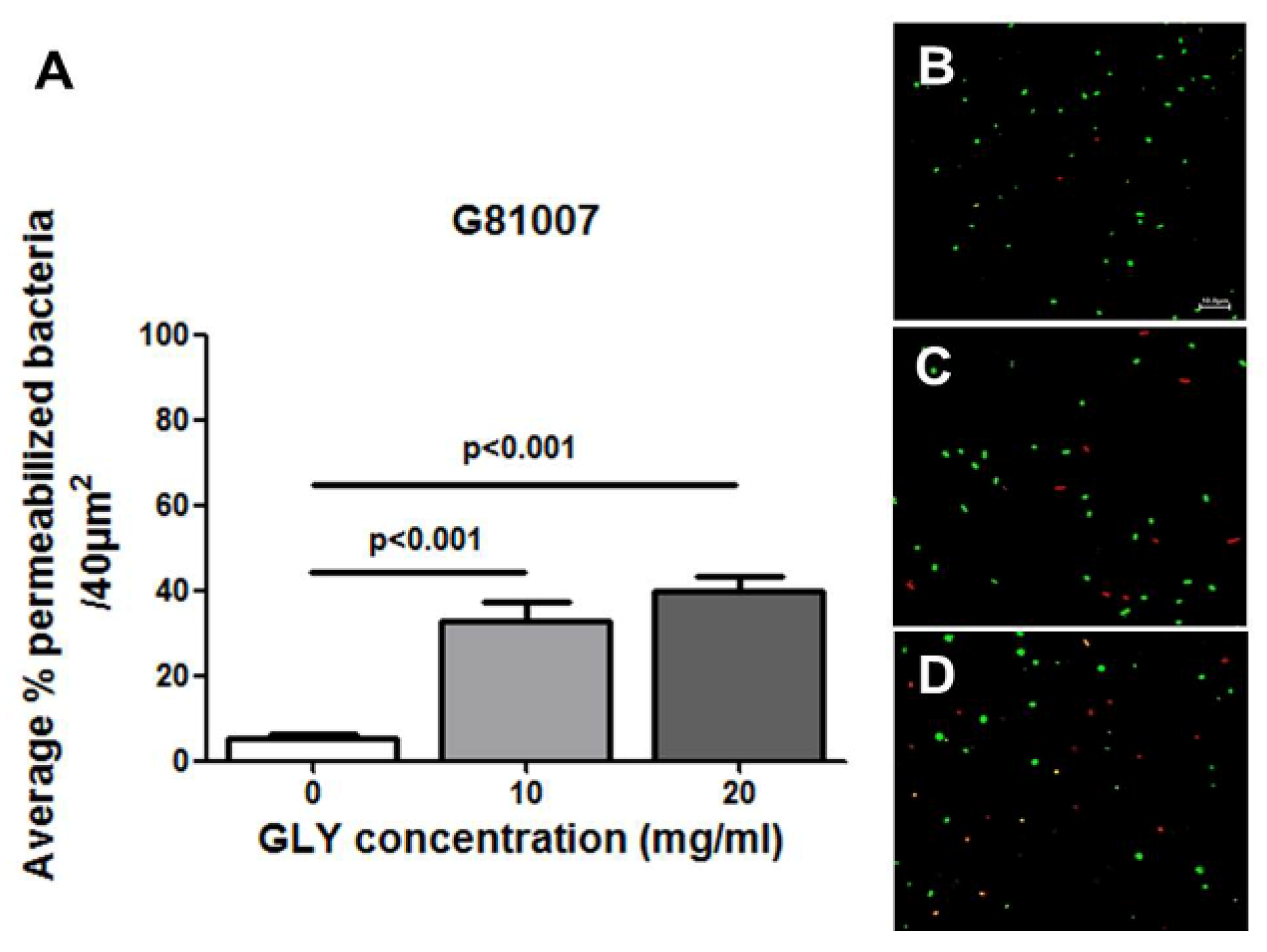
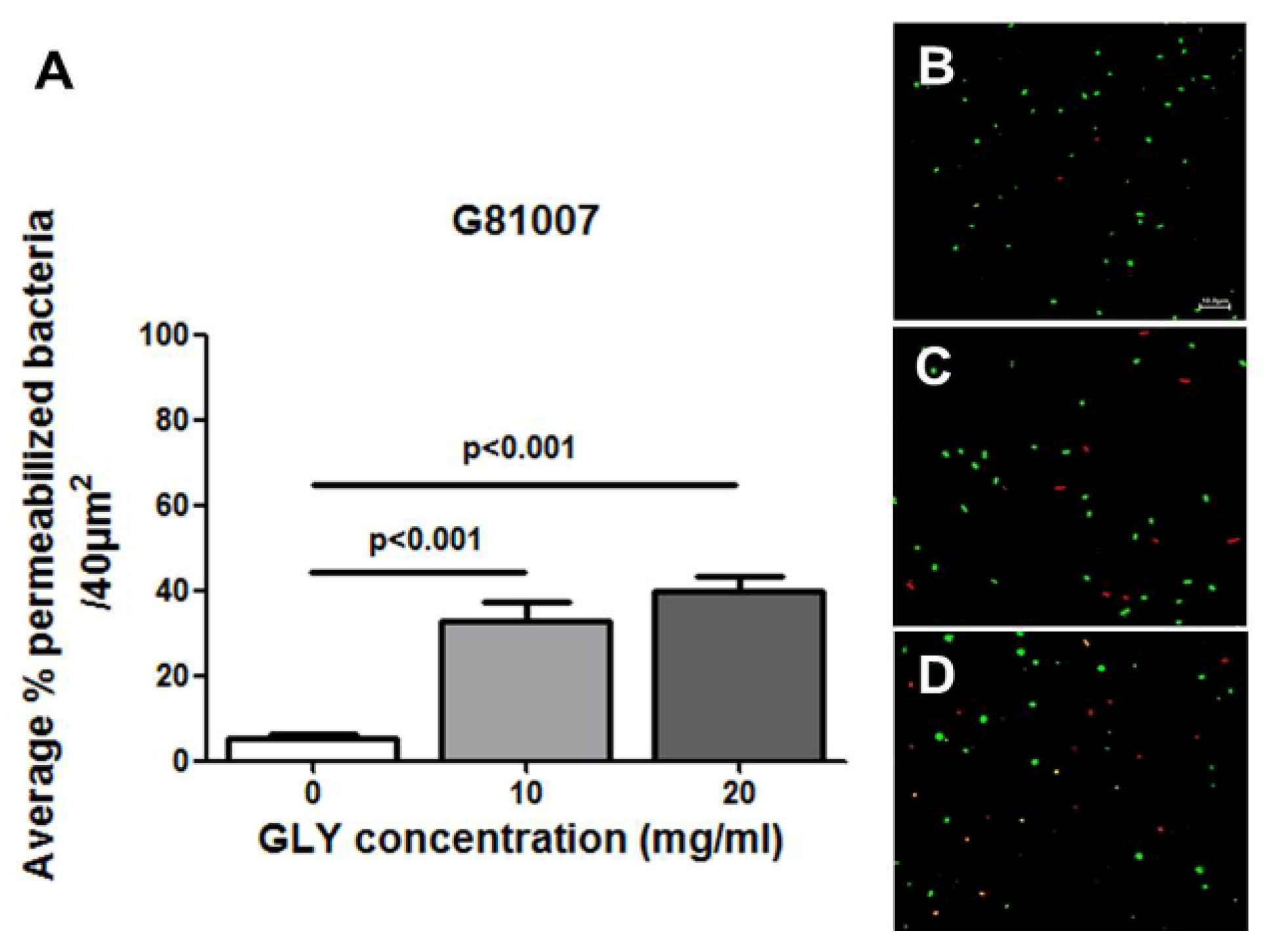
2.4. Live/Dead Baclight Assay
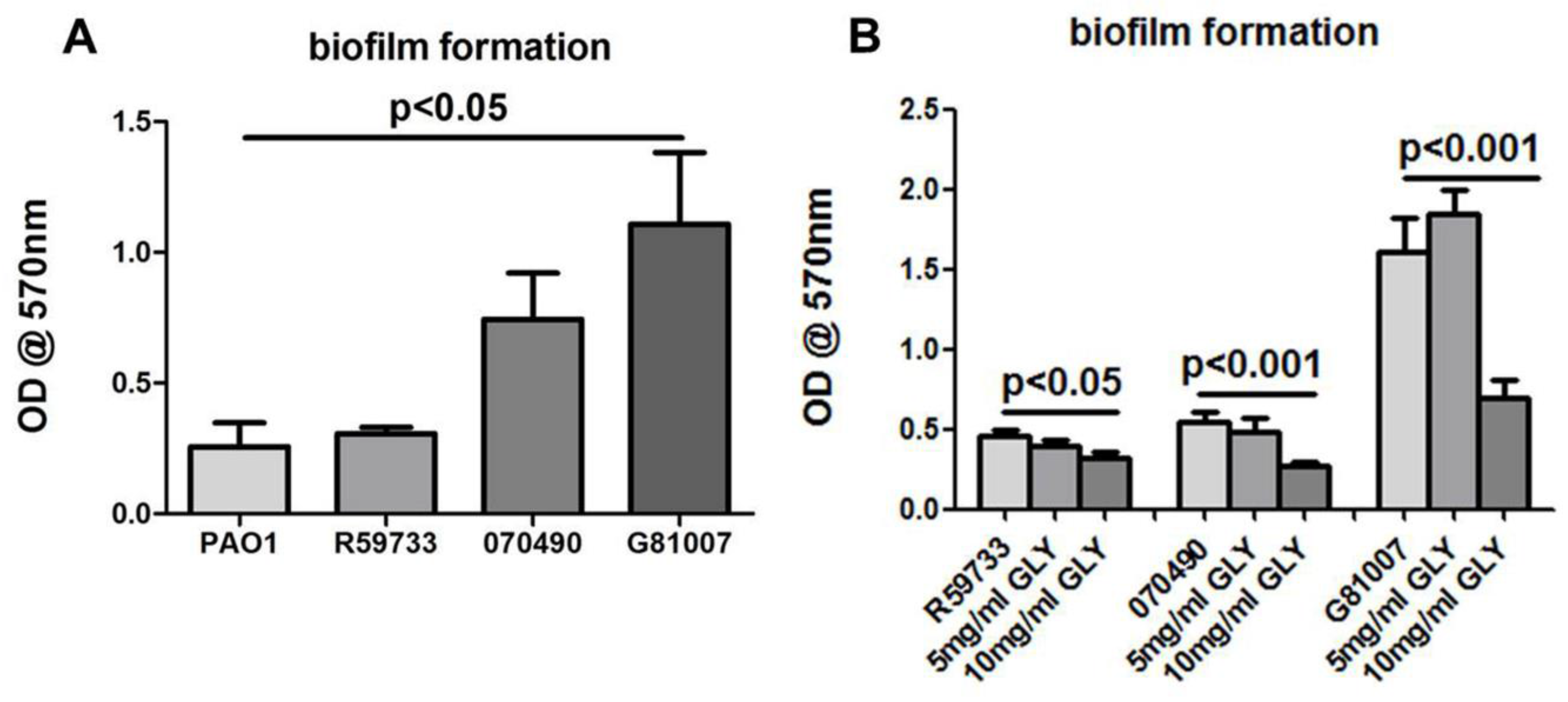
2.5. Biofilm Formation and Exposure to GLY
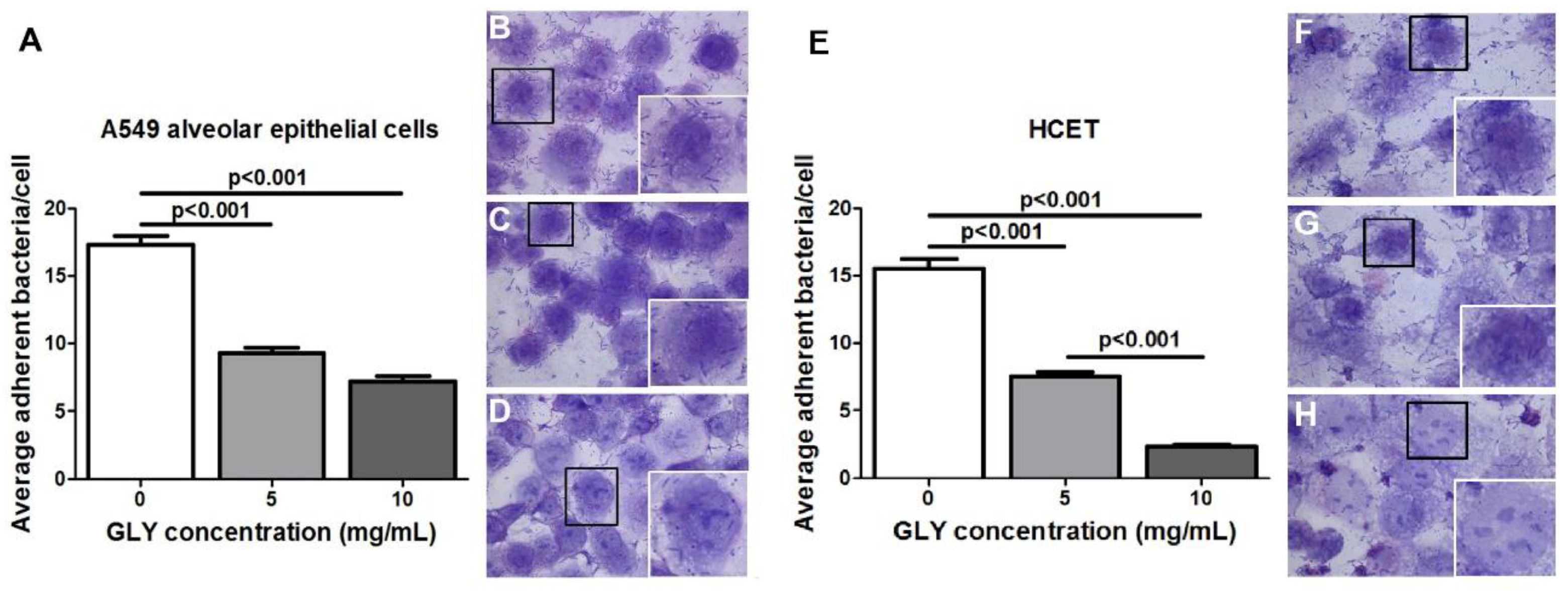
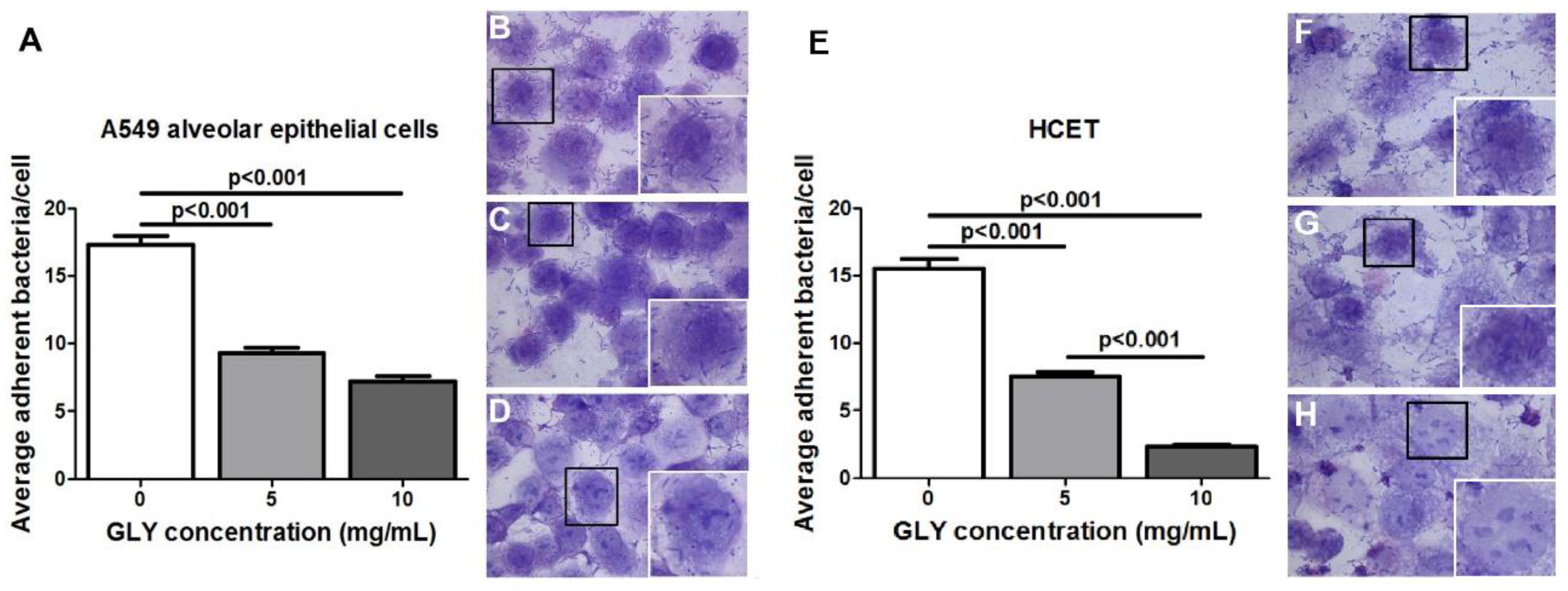
2.6. Bacterial Adherence Assay
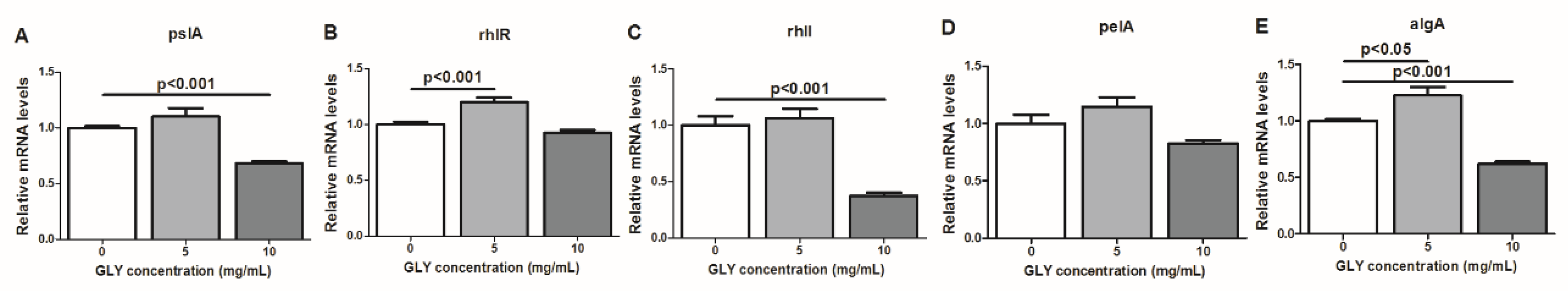
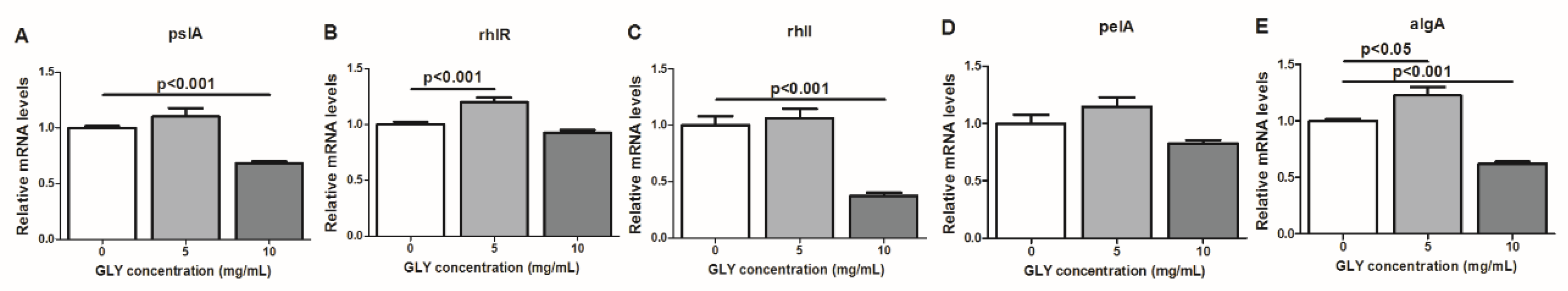
2.7. RT-PCR: mRNA Expression of Bacterial Factors Implicated in the Regulation of P. aeruginosa Biofilm Formation After GLY Treatment
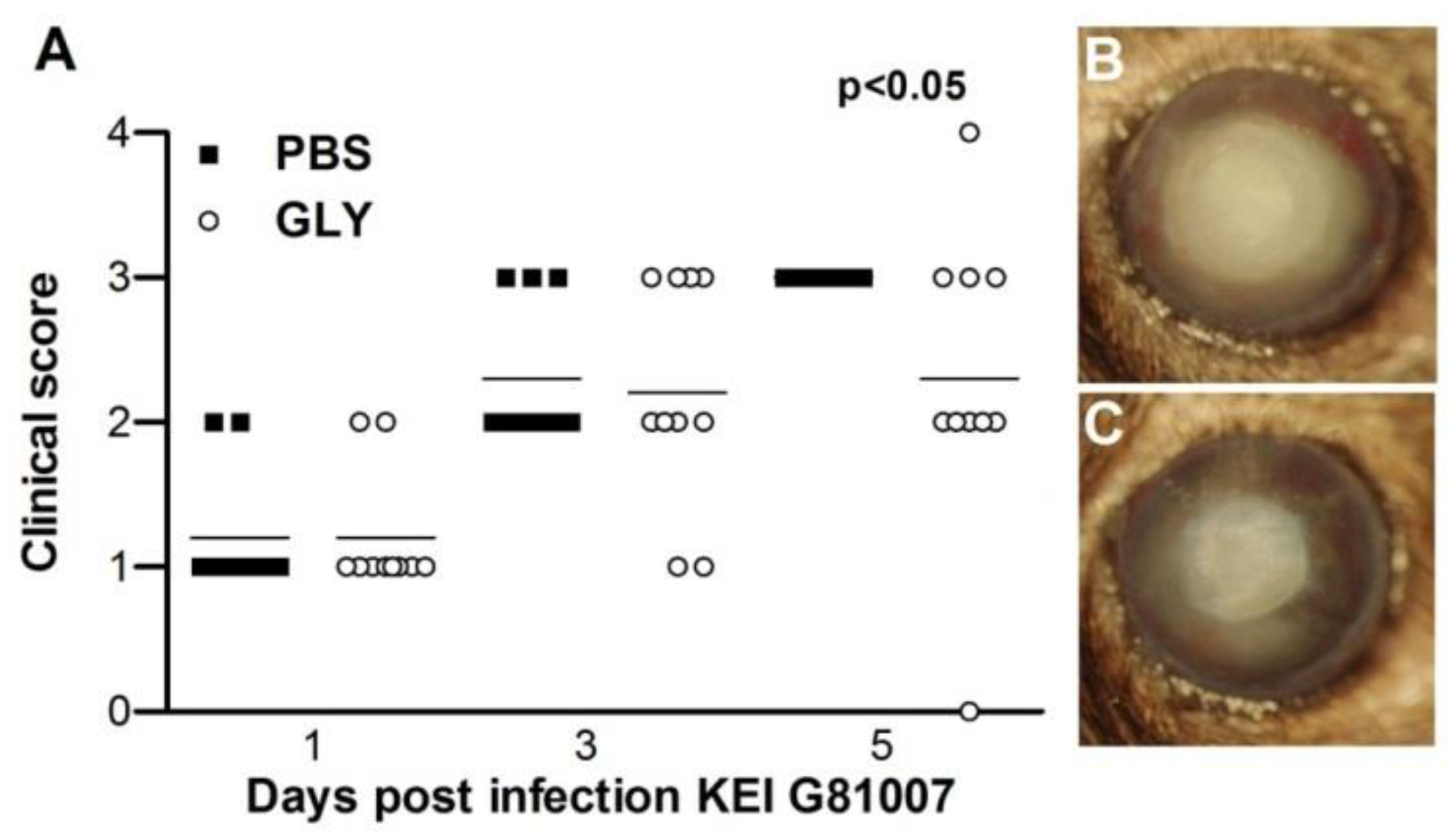
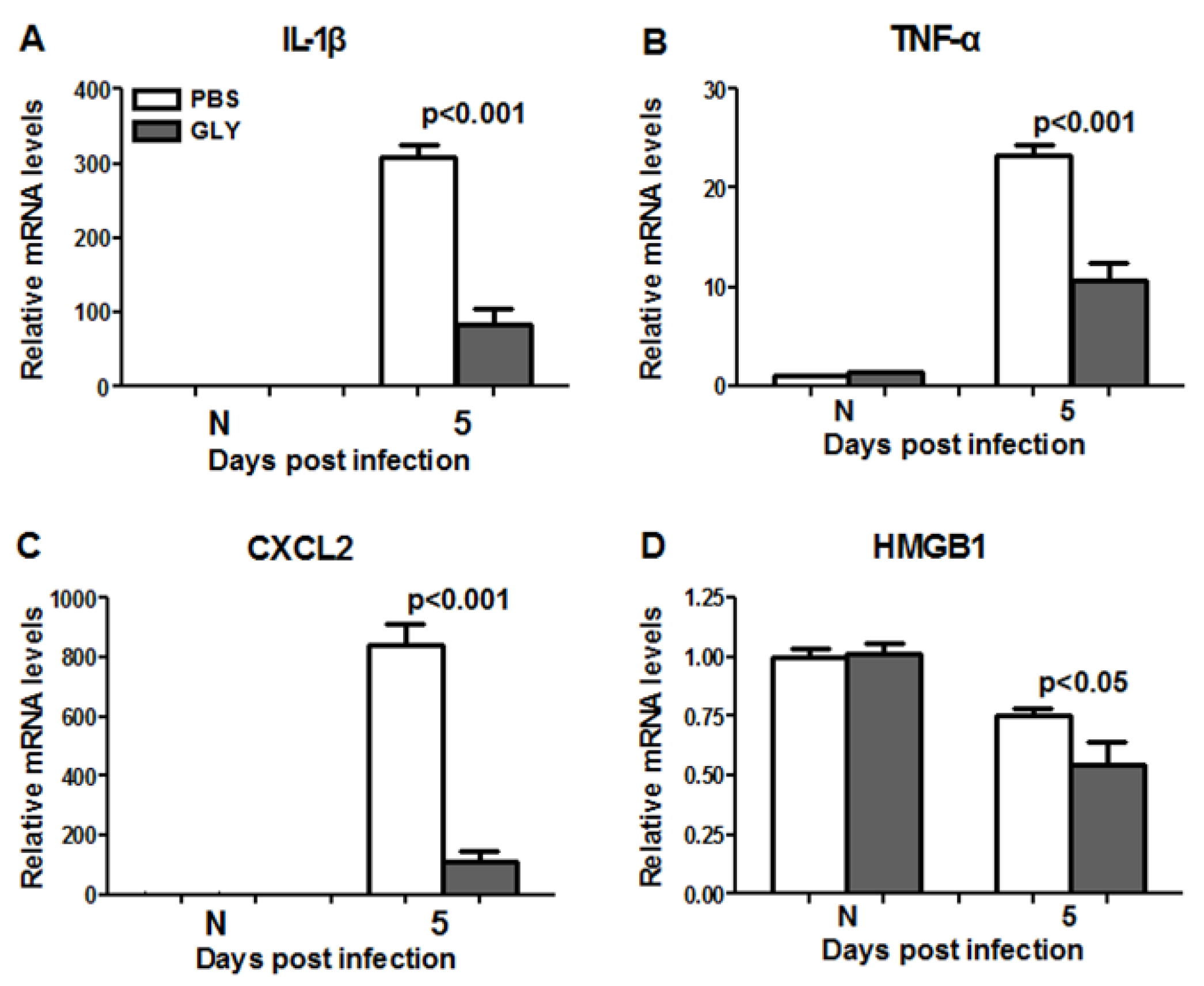
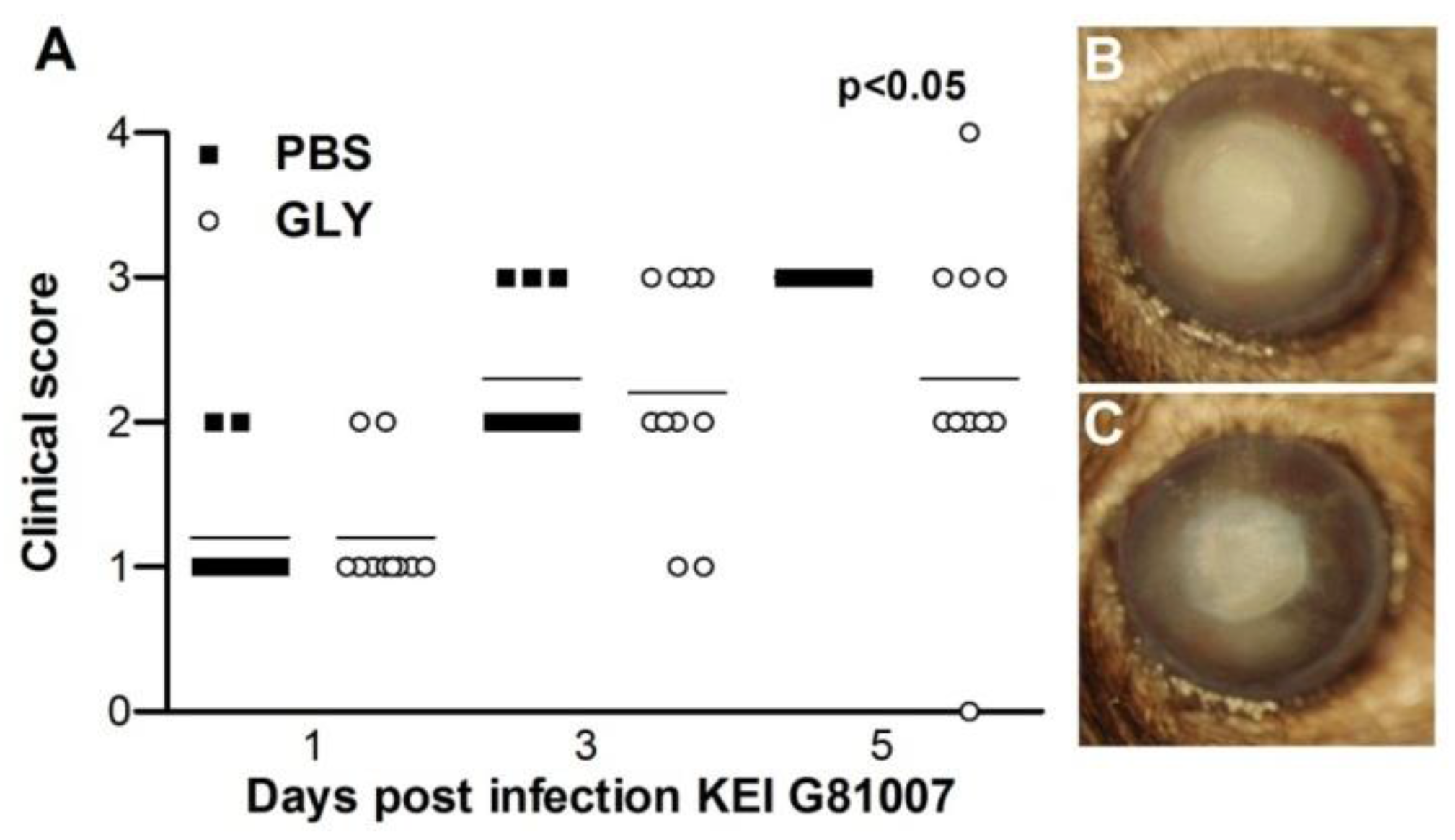
2.8. Infectivity of Isolate G81007 and GLY Treatment
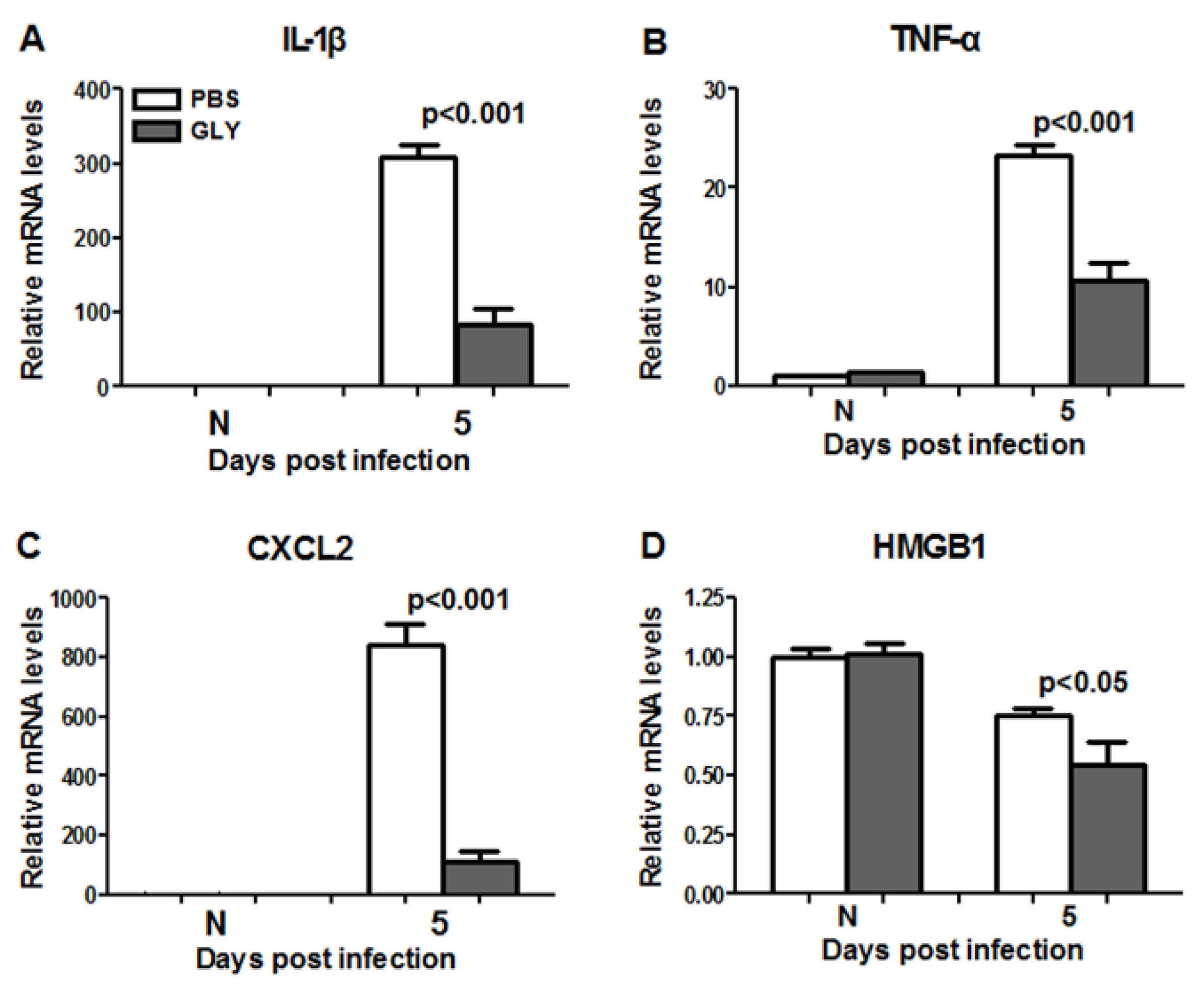
2.9. RT-PCR: GLY Treatment after G81007 Infection
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Bacterial Culture and Infection
4.3. Ocular Response to Bacterial Infection
4.4. GLY Treatment
4.5. Amplification of Virulence Genes Encoded by Type III Secretion System
4.6. Real Time RT-PCR
4.7. Minimum Inhibitory Concentration (MIC)
4.8. Time Kill Assay
4.9. Bacterial Adherence Assay
4.10. Live/Dead BacLight Bacterial Viability
4.11. Biofilm Formation and Exposure to Glycyrrhizin
4.12. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hazlett, L.D. Corneal response to Pseudomonas aeruginosa infection. Prog. Retin. Eye Res. 2004, 23, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Mondino, B.J.; Weissman, B.A.; Farb, M.D.; Pettit, T.H. Corneal ulcers associated with daily-wear and extended-wear contact lenses. Am. J. Ophthalmol. 1986, 102, 58–65. [Google Scholar] [CrossRef]
- Yildiz, E.H.; Airiani, S.; Hammersmith, K.M.; Rapuano, C.J.; Laibson, P.R.; Virdi, A.S.; Hongyok, T.; Cohen, E.J. Trends in contact lens-related corneal ulcers at a tertiary referral center. Cornea 2012, 31, 1097–1102. [Google Scholar] [CrossRef] [PubMed]
- Gellatly, S.L.; Hancock, R.E. Pseudomonas aeruginosa: New insights into pathogenesis and host defenses. Pathog. Dis. 2013, 67, 159–173. [Google Scholar] [CrossRef] [PubMed]
- Filloux, A. Protein Secretion Systems in Pseudomonas aeruginosa: An essay on diversity, evolution, and function. Front. Microbiol. 2011, 2, 155. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.H.; Huang, S.P.; Teng, H.C.; Ji, D.D.; Chen, Y.S.; Chen, Y.L. Presence of the exoU gene of Pseudomonas aeruginosa is correlated with cytotoxicity in MDCK cells but not with colonization in BALB/c mice. J. Clin. Microbiol. 2006, 44, 4596–4597. [Google Scholar] [CrossRef] [PubMed]
- Lee, V.T.; Smith, R.S.; Tummler, B.; Lory, S. Activities of Pseudomonas aeruginosa effectors secreted by the Type III secretion system in vitro and during infection. Infect. Immun. 2005, 73, 1695–1705. [Google Scholar] [CrossRef] [PubMed]
- Wong-Beringer, A.; Wiener-Kronish, J.; Lynch, S.; Flanagan, J. Comparison of type III secretion system virulence among fluoroquinolone-susceptible and -resistant clinical isolates of Pseudomonas aeruginosa. Clin. Microbiol. Infect. 2008, 14, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Hauser, A.R. The type III secretion system of Pseudomonas aeruginosa: Infection by injection. Nat. Rev. Microbiol. 2009, 7, 654–665. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Yoon, S.S. Pseudomonas aeruginosa biofilm, a programmed bacterial life for fitness. J. Microbiol. Biotechnol. 2017, 27, 1053–1064. [Google Scholar] [CrossRef] [PubMed]
- Tolker-Nielsen, T. Pseudomonas aeruginosa biofilm infections: From molecular biofilm biology to new treatment possibilities. APMIS Suppl. 2014, 122, 1–51. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.K.; Yeung, A.T.; Hancock, R.E. Antibiotic resistance in Pseudomonas aeruginosa biofilms: Towards the development of novel anti-biofilm therapies. J. Biotechnol. 2014, 191, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Lister, P.D.; Wolter, D.J.; Hanson, N.D. Antibacterial-resistant Pseudomonas aeruginosa: Clinical impact and complex regulation of chromosomally encoded resistance mechanisms. Clin. Microbiol. Rev. 2009, 22, 582–610. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.L.; Gangwani, R.A.; Yu, L.W.; Lai, J.S. New treatments for bacterial keratitis. J. Ophthalmol. 2012, 2012, 831502. [Google Scholar] [CrossRef] [PubMed]
- Andersson, U.; Tracey, K.J. HMGB1 is a therapeutic target for sterile inflammation and infection. Annu. Rev. Immunol. 2011, 29, 139–162. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Bloom, O.; Zhang, M.; Vishnubhakat, J.M.; Ombrellino, M.; Che, J.; Frazier, A.; Yang, H.; Ivanova, S.; Borovikova, L.; et al. HMG-1 as a late mediator of endotoxin lethality in mice. Science 1999, 285, 248–251. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Zeng, J.; Mo, B.; Wang, C.; Huang, J.; Sun, Y.; Yu, Y.; Liu, S. High mobility group box 1: A novel mediator of Th2-type response-induced airway inflammation of acute allergic asthma. J. Thorac. Dis. 2015, 7, 1732–1741. [Google Scholar] [CrossRef] [PubMed]
- Wahamaa, H.; Schierbeck, H.; Hreggvidsdottir, H.S.; Palmblad, K.; Aveberger, A.C.; Andersson, U.; Harris, H.E. High mobility group box protein 1 in complex with lipopolysaccharide or IL-1 promotes an increased inflammatory phenotype in synovial fibroblasts. Arthrit Res. Ther. 2011, 13, R136. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yang, H.; Tracey, K.J. Extracellular role of HMGB1 in inflammation and sepsis. J. Inter. Med. 2004, 255, 320–331. [Google Scholar] [CrossRef]
- Cavone, L.; Muzzi, M.; Mencucci, R.; Sparatore, B.; Pedrazzi, M.; Moroni, F.; Chiarugi, A. 18beta-glycyrrhetic acid inhibits immune activation triggered by HMGB1, a pro-inflammatory protein found in the tear fluid during conjunctivitis and blepharitis. Ocul. Immunol. Inflamm. 2011, 19, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Ekanayaka, S.A.; McClellan, S.A.; Barrett, R.P.; Kharotia, S.; Hazlett, L.D. Glycyrrhizin reduces HMGB1 and bacterial load in Pseudomonas aeruginosa keratitis. Investig. Ophthalmol. Vis. Sci. 2016, 57, 5799–5809. [Google Scholar] [CrossRef] [PubMed]
- Krausse, R.; Bielenberg, J.; Blaschek, W.; Ullmann, U. In vitro anti-Helicobacter pylori activity of Extractum liquiritiae, glycyrrhizin and its metabolites. J. Antimicrob. Chemother. 2004, 54, 243–246. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.H.; He, L.; Long, W.; Zhou, Q.; Zhu, S.; Wang, P.; Fan, S.; Wang, H. Novel Mechanisms of Herbal Therapies for Inhibiting HMGB1 Secretion or Action. Evid.-Based Complement. Altern. Med. 2015, 2015, 456305. [Google Scholar] [CrossRef] [PubMed]
- Mollica, L.; De Marchis, F.; Spitaleri, A.; Dallacosta, C.; Pennacchini, D.; Zamai, M.; Agresti, A.; Trisciuoglio, L.; Musco, G.; Bianchi, M.E. Glycyrrhizin binds to high-mobility group box 1 protein and inhibits its cytokine activities. Chem. Biol. 2007, 14, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.; Lee, J.O.; Nakamura, K.; Suzuki, S.; Hendon, D.N.; Kobayashi, M.; Suzuki, F. Effect of glycyrrhizin on pseudomonal skin infections in human-mouse chimeras. PLoS ONE 2014, 9, e83747. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.F.; Kuai, J.K.; Lu, Z.F.; Yang, G.D.; Fu, H.Y.; Wang, J.; Tian, F.; Yan, X.L.; Zhao, Y.C.; Wang, Y.J. Glycyrrhizin treatment is associated with attenuation of lipopolysaccharide-induced acute lung injury by inhibiting cyclooxygenase-2 and inducible nitric oxide synthase expression. J. Surg. Res. 2011, 165, e29–e35. [Google Scholar] [CrossRef] [PubMed]
- Li, K.L.; Zhang, J.G.; Wang, H.M.; Chen, J. The effects of glycyrrhizin-18alpha on the expression of NF-kappaB in the renal interstitium of rats with obstructive nephropathy. Chin. J. Cell. Mol. Immunol. 2004, 20, 31–33. [Google Scholar]
- Arase, Y.; Ikeda, K.; Murashima, N.; Chayama, K.; Tsubota, A.; Koida, I.; Suzuki, Y.; Saitoh, S.; Kobayashi, M.; Kumada, H. The long term efficacy of glycyrrhizin in chronic hepatitis C patients. Cancer 1997, 79, 1494–1500. [Google Scholar] [CrossRef]
- Pachigolla, G.; Blomquist, P.; Cavanagh, H.D. Microbial keratitis pathogens and antibiotic susceptibilities: A 5-year review of cases at an urban county hospital in north Texas. Eye Contact Lens 2007, 33, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Stapleton, F.; Carnt, N. Contact lens-related microbial keratitis: How have epidemiology and genetics helped us with pathogenesis and prophylaxis. Eye 2012, 26, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Asl, M.N.; Hosseinzadeh, H. Review of pharmacological effects of Glycyrrhiza sp. and its bioactive compounds. Phytother. Res. 2008, 22, 709–724. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xiang, J.; Liu, M.; Wang, S.; Lee, R.J.; Ding, H. Protective effects of glycyrrhizic acid by rectal treatment on a TNBS-induced rat colitis model. J. Pharm. Pharmacol. 2011, 63, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhao, F.; Fang, Y.; Li, X.; Shen, L.; Cao, T.; Zhu, H. Glycyrrhizin protects against porcine endotoxemia through modulation of systemic inflammatory response. Crit. Care 2013, 17, R44. [Google Scholar] [CrossRef] [PubMed]
- Gong, G.; Xiang, L.; Yuan, L.; Hu, L.; Wu, W.; Cai, L.; Yin, L.; Dong, H. Protective effect of glycyrrhizin, a direct HMGB1 inhibitor, on focal cerebral ischemia/reperfusion-induced inflammation, oxidative stress, and apoptosis in rats. PLoS ONE 2014, 9, e89450. [Google Scholar] [CrossRef] [PubMed]
- Davis, E.A.; Morris, D.J. Medicinal uses of licorice through the millennia: The good and plenty of it. Mol. Cell. Endocrinol. 1991, 78, 1–6. [Google Scholar] [CrossRef]
- McClellan, S.; Jiang, X.; Barrett, R.; Hazlett, L.D. High-mobility group box 1: A novel target for treatment of Pseudomonas aeruginosa keratitis. J. Immunol. 2015, 194, 1776–1787. [Google Scholar] [CrossRef] [PubMed]
- Musumeci, D.; Roviello, G.N.; Montesarchio, D. An overview on HMGB1 inhibitors as potential therapeutic agents in HMGB1-related pathologies. Pharmacol. Ther. 2014, 141, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Carion, T.W.; McWhirter, C.R.; Grewal, D.K.; Berger, E.A. Efficacy of VIP as Treatment for Bacteria-Induced Keratitis Against Multiple Pseudomonas aeruginosa Strains. Investig. Ophthalmol. Vis. Sci. 2015, 56, 6932–6940. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Wang, Q.; de la Fuente-Nunez, C.; Sun, F.J.; Xia, J.G.; Xia, P.Y.; Hancock, R.E. Increased IL-8 production in human bronchial epithelial cells after exposure to azithromycin-pretreated Pseudomonas aeruginosa in vitro. FEMS Microbiol. Lett. 2014, 355, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Berthelot, P.; Attree, I.; Plesiat, P.; Chabert, J.; de Bentzmann, S.; Pozzetto, B.; Grattard, F. Genotypic and phenotypic analysis of type III secretion system in a cohort of Pseudomonas aeruginosa bacteremia isolates: Evidence for a possible association between O serotypes and exo genes. J. Infect. Dis. 2003, 188, 512–518. [Google Scholar] [CrossRef] [PubMed]
- Wareham, D.W.; Curtis, M.A. A genotypic and phenotypic comparison of type III secretion profiles of Pseudomonas aeruginosa cystic fibrosis and bacteremia isolates. Internat. J. Med. Microbiol. 2007, 297, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Choy, M.H.; Stapleton, F.; Willcox, M.D.; Zhu, H. Comparison of virulence factors in Pseudomonas aeruginosa strains isolated from contact lens- and non-contact lens-related keratitis. J. Med. Microbiol. 2008, 57, 1539–1546. [Google Scholar] [CrossRef] [PubMed]
- Irani, M.; Sarmadi, M.; Bernard, F.; Ebrahimi Pour, G.H.; Shaker Bazarnov, H. Leaves Antimicrobial activity of Glycyrrhiza glabra. Iran. J. Pharm. Res. 2010, 9, 425–428. [Google Scholar] [PubMed]
- Yu, G.; Wang, X.; Dou, Y.; Wang, S.; Tian, M.; Qi, J.; Li, T.; Ding, C.; Wu, Y.; Yu, S. Riemerella anatipestifer M949_1360 Gene functions on the lipopolysaccharide biosynthesis and bacterial virulence. PLoS ONE 2016, 11, e0160708. [Google Scholar] [CrossRef] [PubMed]
- Bukhary, S.; Balto, H. Antibacterial efficacy of octenisept, alexidine, chlorhexidine, and sodium hypochlorite against Enterococcus faecalis biofilms. J. Endodont. 2017, 43, 643–647. [Google Scholar] [CrossRef] [PubMed]
- Berney, M.; Hammes, F.; Bosshard, F.; Weilenmann, H.U.; Egli, T. Assessment and interpretation of bacterial viability by using the LIVE/DEAD BacLight Kit in combination with flow cytometry. Appl. Environ. Microbiol. 2007, 73, 3283–3290. [Google Scholar] [CrossRef] [PubMed]
- Barbesti, S.; Citterio, S.; Labra, M.; Baroni, M.D.; Neri, M.G.; Sgorbati, S. Two and three-color fluorescence flow cytometric analysis of immunoidentified viable bacteria. Cytometry 2000, 40, 214–218. [Google Scholar] [CrossRef]
- Berney, M.; Weilenmann, H.U.; Egli, T. Flow-cytometric study of vital cellular functions in Escherichia coli during solar disinfection (SODIS). Microbiology 2006, 152, 1719–1729. [Google Scholar] [CrossRef] [PubMed]
- Kline, K.A.; Falker, S.; Dahlberg, S.; Normark, S.; Henriques-Normark, B. Bacterial adhesins in host-microbe interactions. Cell Host Microbe 2009, 5, 580–592. [Google Scholar] [CrossRef] [PubMed]
- Hahn, H.P. The type-4 pilus is the major virulence-associated adhesin of Pseudomonas aeruginosa—A Review. Gene 1997, 192, 99–108. [Google Scholar] [CrossRef]
- Zoutman, D.E.; Hulbert, W.C.; Pasloske, B.L.; Joffe, A.M.; Volpel, K.; Trebilcock, M.K.; Paranchych, W. The role of polar pili in the adherence of Pseudomonas aeruginosa to injured canine tracheal cells: A semiquantitative morphologic study. Scanning Microsc. 1991, 5, 109–124; discussion 124–126. [Google Scholar] [PubMed]
- Chi, E.; Mehl, T.; Nunn, D.; Lory, S. Interaction of Pseudomonas aeruginosa with A549 pneumocyte cells. Infect. Immun. 1991, 59, 822–828. [Google Scholar] [PubMed]
- Farinha, M.A.; Conway, B.D.; Glasier, L.M.; Ellert, N.W.; Irvin, R.T.; Sherburne, R.; Paranchych, W. Alteration of the pilin adhesin of Pseudomonas aeruginosa PAO results in normal pilus biogenesis but a loss of adherence to human pneumocyte cells and decreased virulence in mice. Infect. Immun. 1994, 62, 4118–4123. [Google Scholar] [PubMed]
- Kostakioti, M.; Hadjifrangiskou, M.; Hultgren, S.J. Bacterial biofilms: Development, dispersal, and therapeutic strategies in the dawn of the postantibiotic era. CSH Perspect. Med. 2013, 3, a010306. [Google Scholar] [CrossRef] [PubMed]
- Parsek, M.R.; Singh, P.K. Bacterial biofilms: An emerging link to disease pathogenesis. Annu. Rev. Microbiol. 2003, 7, 677–701. [Google Scholar] [CrossRef] [PubMed]
- Ryder, C.; Byrd, M.; Wozniak, D.J. Role of polysaccharides in Pseudomonas aeruginosa biofilm development. Curr. Opin. Microbiol. 2007, 10, 644–648. [Google Scholar] [CrossRef] [PubMed]
- Ghafoor, A.; Hay, I.D.; Rehm, B.H. Role of exopolysaccharides in Pseudomonas aeruginosa biofilm formation and architecture. Appl. Environ. Microbiol. 2011, 77, 5238–5246. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, I. Biofilm exopolysaccharides: A strong and sticky framework. Microbiolgy 2001, 147, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Colvin, K.M.; Gordon, V.D.; Murakami, K.; Borlee, B.R.; Wozniak, D.J.; Wong, G.C.; Parsek, M.R. The pel polysaccharide can serve a structural and protective role in the biofilm matrix of Pseudomonas aeruginosa. PLoS Pat. 2011, 7, e1001264. [Google Scholar] [CrossRef] [PubMed]
- Vasseur, P.; Vallet-Gely, I.; Soscia, C.; Genin, S.; Filloux, A. The pel genes of the Pseudomonas aeruginosa PAK strain are involved at early and late stages of biofilm formation. Microbiology 2005, 151, 985–997. [Google Scholar] [CrossRef] [PubMed]
- De Kievit, T.R. Quorum sensing in Pseudomonas aeruginosa biofilms. Environ. Microbiol. 2009, 11, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Parsek, M.R.; Greenberg, E.P. Sociomicrobiology: The connections between quorum sensing and biofilms. Trends Microbiol. 2005, 13, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, P.N.; Koch, G.; Thompson, J.A.; Xavier, K.B.; Cool, R.H.; Quax, W.J. The multiple signaling systems regulating virulence in Pseudomonas aeruginosa. Microbiol. Mol. Biol. Rev. 2012, 76, 46–65. [Google Scholar] [CrossRef] [PubMed]
- Sakuragi, Y.; Kolter, R. Quorum-sensing regulation of the biofilm matrix genes (pel) of Pseudomonas aeruginosa. J. Bacteriol. 2007, 189, 5383–5386. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, L.R.; D’Argenio, D.A.; MacCoss, M.J.; Zhang, Z.; Jones, R.A.; Miller, S.I. Aminoglycoside antibioitics induce bacterial biofilm formation. Nat. Lett. 2005, 436, 1171–1175. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Fang, Y.; Deng, S.; Li, X.; Zhou, Y.; Gong, Y.; Zhu, H.; Wang, W. Glycyrrhizin protects rats from sepsis by blocking HMGB1 signaling. BioMed Res. Int. 2017, 2017, 9719647. [Google Scholar] [CrossRef] [PubMed]
- Hazlett, L.D.; Moon, M.M.; Strejc, M.; Berk, R.S. Evidence for N-acetylmannosamine as an ocular receptor for P. aeruginosa adherence to scarified cornea. Investig. Ophthalmol. Vis. Sci. 1987, 28, 1978–1985. [Google Scholar]
- Mencucci, R.; Favuzza, E.; Menchini, U. Assessment of the tolerability profile of an ophthalmic solution of 5% glycyrrhizin and copolymer PEG/PPG on healthy volunteers and evaluation of its efficacy in the treatment of moderate to severe blepharitis. Clin. Ophthalmol. 2013, 7, 1403–1410. [Google Scholar] [CrossRef] [PubMed]
- Azimi, S.; Kafil, H.S.; Baghi, H.B.; Shokrian, S.; Najaf, K.; Asgharzadeh, M.; Yousefi, M.; Shahrivar, F.; Aghazadeh, M. Presence of exoY, exoS, exoU and exoT genes, antibiotic resistance and biofilm production among Pseudomonas aeruginosa isolates in Northwest Iran. GMS Hyg. Infect. Ctrl 2016, 11, Doc04. [Google Scholar] [CrossRef]
- Jabalameli, F.; Mirsalehian, A.; Khoramian, B.; Aligholi, M.; Khoramrooz, S.S.; Asadollahi, P.; Taherikalani, M.; Emaneini, M. Evaluation of biofilm production and characterization of genes encoding type III secretion system among Pseudomonas aeruginosa isolated from burn patients. J. Int. Soc. Burn Inj. 2012, 38, 1192–1197. [Google Scholar] [CrossRef] [PubMed]
- Lesic, B.; Starkey, M.; He, J.; Hazan, R.; Rahme, L.G. Quorum sensing differetially regultes Pseudomonas aeruginosa type VI secreation locus I and homologus loci II and III, which are required for pathogenesis. Microbiology 2009, 155, 2845–2855. [Google Scholar] [CrossRef] [PubMed]
- Franzblau, S.G.; DeGroote, M.A.; Cho, S.H.; Andries, K.; Nuermberger, E.; Orme, I.M.; Mdluli, K.; Angulo-Barturen, I.; Dick, T.; Dartois, V.; et al. Comprehensive analysis of methods used for the evaluation of compounds against Mycobacterium tuberculosis. Tuberculosis 2012, 92, 453–488. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, A.J.; Chopra, I. Preclinical evaluation of novel antibacterial agents by microbiological and molecular techniques. Expert Opin. Investig. Drugs 2004, 13, 1045–1063. [Google Scholar] [CrossRef] [PubMed]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef]
- Barry, A.L.; Craig, W.A.; Nadler, H.; Reller, L.B.; Sanders, C.C.; Swenson, J.M. Methods for determining bactericidal activity of antimicrobial agents: Approved guideline. In NCCLS Document M26-A; 1999; Volume 19. [Google Scholar]
- Senoo, T.; Joyce, N.C. Cell cycle kinetics in corneal endothelium from old and young donors. Investig. Ophthalmol. Vis. Sci. 2000, 41, 660–667. [Google Scholar]
- Stepanovic, S.; Vukovic, D.; Hola, V.; Di Bonaventura, G.; Djukic, S.; Cirkovic, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. APMIS 2007, 115, 891–899. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cytotoxin Gene | Nucleotide Sequence (5′-3′) | Product Size (bp) | Reference |
|---|---|---|---|
| exoS | F-ATC CTC AGG CGT ACA TCC | 328 | [69] |
| R-ACG ACG GCT ATC TCT CCA C | |||
| exoT | F-AGA ACC CGT CTT TCG TGG CTG AGT T | 351 | [69] |
| R-CAG CTC GCT CGC CTT GCC AAG T | |||
| exoU | F-CCT TAG CCA TCT CAA CGG TAG TC | 911 | [70] |
| R-GAG GGC GAA GCT GGG GAG GTA |
| Gene | Nucleotide Sequence | Primer | GenBank |
|---|---|---|---|
| β-actin | 5′-GAT TAC TGC TCT GGC TCC TAG C-3′ | F | NM_007393.3 |
| 5′-GAC TCA TCG TAC TCC TGC TTG C-3′ | R | ||
| HMGB1 | 5′-TGG CAA AGG CTG ACA AGG CTC-3′ | F | NM_010439.3 |
| 5′-GGA TGC TCG CCT TTG ATT TTG G-3′ | R | ||
| IL-1β | 5′-CGC AGC AGC ACA TCA ACA AGA GC-3′ | F | NM_008361.3 |
| 5′-TGT CCT CAT CCT GGA AGG TCC ACG-3′ | R | ||
| CXCL2 | 5′-TGT CAA TGC CTG AAG ACC CTG CC-3′ | F | NM_009140.2 |
| 5′-AAC TTT TTG ACC GCC CTT GAG AGT GG-3′ | R | ||
| TNF-α | 5′-ACC CTC ACA CTC AGA TCA TCT T-3′ | F | NM_013693.2 |
| 5′-GGT TGT CTT TGA GAT CCA TGC-3′ | R |
| Gene | Nucleotide Sequence | Primer | Gen ID |
|---|---|---|---|
| rpoD | 5′-CAT CCG CAT GAT CAA CGA CA-3′ | F | 882038 |
| 5′-GAT CGA TAT AGC CGC TGA GG-3′ | R | ||
| pslA | 5′-AAG ATC AAG AAA CGC GTG GAA T-3′ | F | 879717 |
| 5′-TGT AGA GGT CGA ACC ACA CCG-3′ | R | ||
| rhlR | 5′-CTG GGC TTC GAT TAC TAC GC-3′ | F | 878968 |
| 5′-CCC GTA GTT CTG CAT CTG GT-3′ | R | ||
| rhlI | 5′-GGA GCG CTA TTT CGT TCG-3′ | F | 878967 |
| 5′-GTA GGC CGG GAA GCT GAT-3′ | R | ||
| pelA | 5′-CCT TCA GCC ATC CGT TCT TCT-3′ | F | 878833 |
| 5′-TCG CGT ACG AAG TCG ACC TT-3′ | R | ||
| algA | 5′-AGA ACC AGT CGA CCT ACA TC-3′ | F | 879142 |
| 5′-ACT GCA CCT CGA TGA TCT C-3′ | R |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, X.; Ekanayaka, S.A.; McClellan, S.A.; Barrett, R.P.; Vistisen, K.; Hazlett, L.D. Characterization of Three Ocular Clinical Isolates of P. aeruginosa: Viability, Biofilm Formation, Adherence, Infectivity, and Effects of Glycyrrhizin. Pathogens 2017, 6, 52. https://doi.org/10.3390/pathogens6040052
Peng X, Ekanayaka SA, McClellan SA, Barrett RP, Vistisen K, Hazlett LD. Characterization of Three Ocular Clinical Isolates of P. aeruginosa: Viability, Biofilm Formation, Adherence, Infectivity, and Effects of Glycyrrhizin. Pathogens. 2017; 6(4):52. https://doi.org/10.3390/pathogens6040052
Chicago/Turabian StylePeng, Xudong, Sandamali A. Ekanayaka, Sharon A. McClellan, Ronald P. Barrett, Kerry Vistisen, and Linda D. Hazlett. 2017. "Characterization of Three Ocular Clinical Isolates of P. aeruginosa: Viability, Biofilm Formation, Adherence, Infectivity, and Effects of Glycyrrhizin" Pathogens 6, no. 4: 52. https://doi.org/10.3390/pathogens6040052