Variations in the Peritrophic Matrix Composition of Heparan Sulphate from the Tsetse Fly, Glossina morsitans morsitans

 , and
, and
Abstract
:1. Introduction
1.1. The Peritrophic Matrix of Glossina morsitans morsitans
1.2. The Composition of the Peritrophic Matrix May Influence Infection and Tissue Tropism
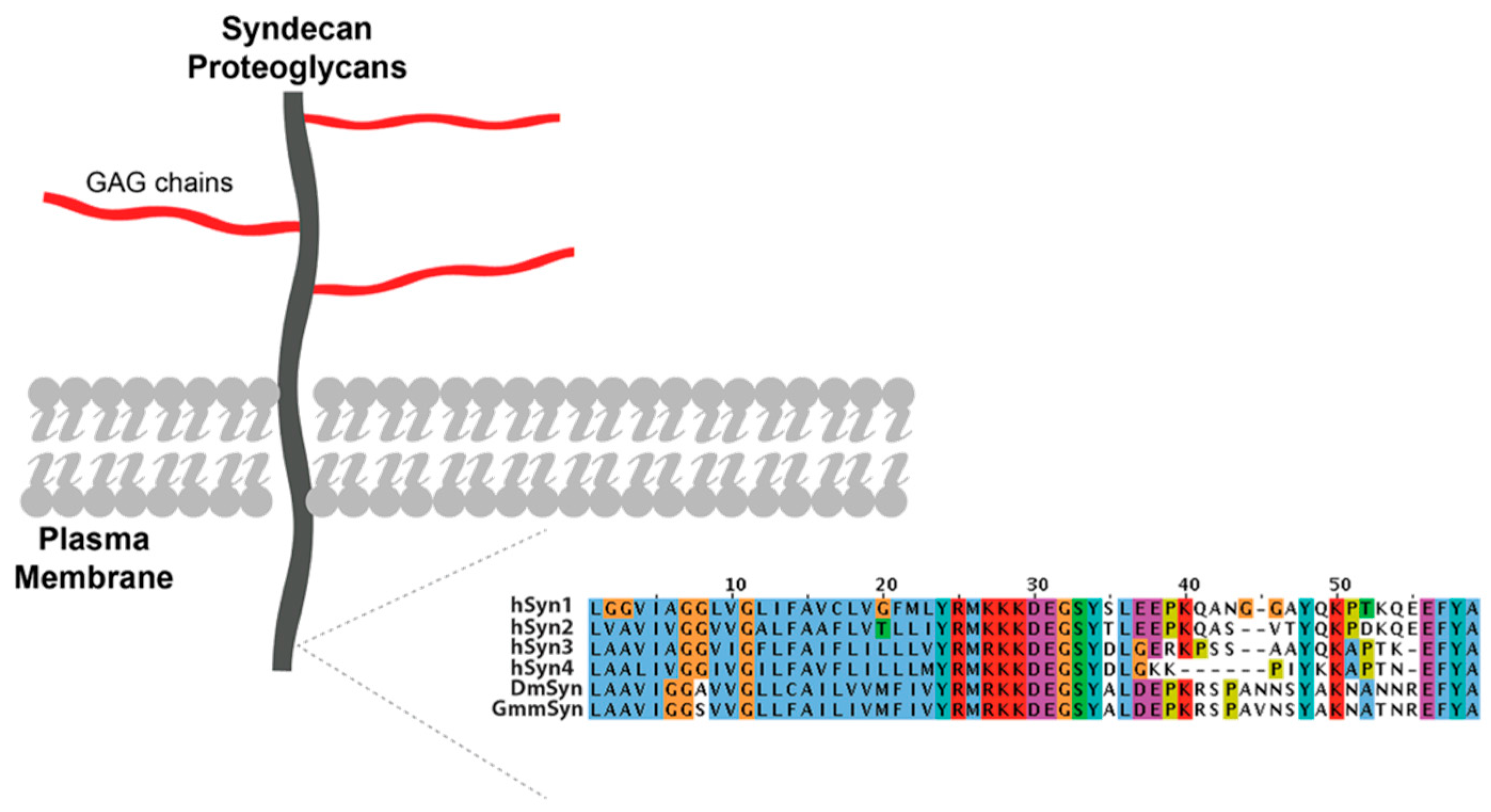
1.3. Proteoglycans in Other Members of the Diptera
1.4. The Overall Structure of Heparan Sulphate Polysaccharides
2. Results
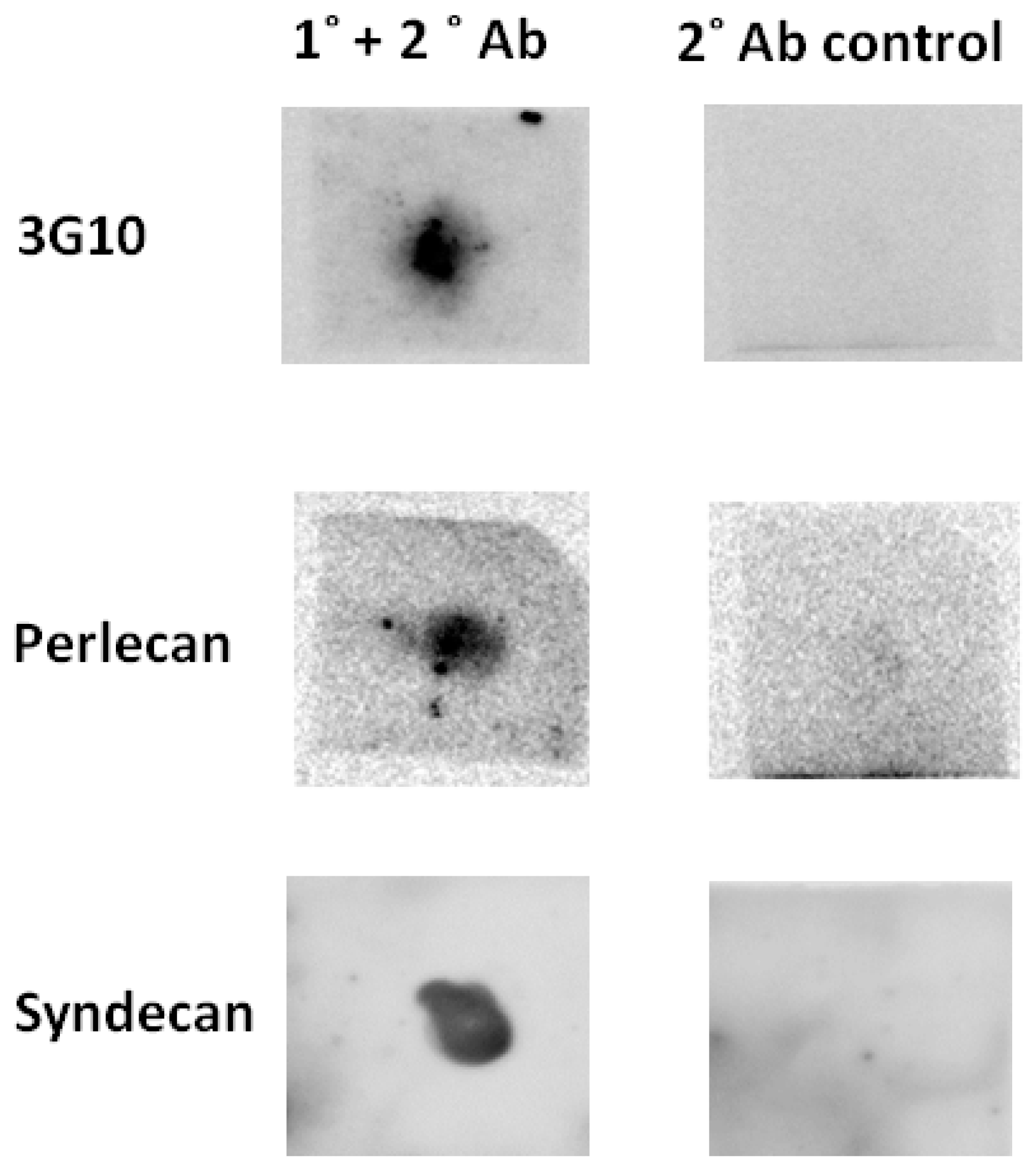
2.1. Heparan Sulphate Proteoglycans Exist in the Peritrophic Matrix of Glossina morsitans morsitans
2.2. It Is Likely That Only One Heparan Sulphate Proteoglycan Exists in Glossina morsitans morsitans
2.3. The Peritrophic Matrix of Glossina morsitans morsitans Contains Heparan Sulphate
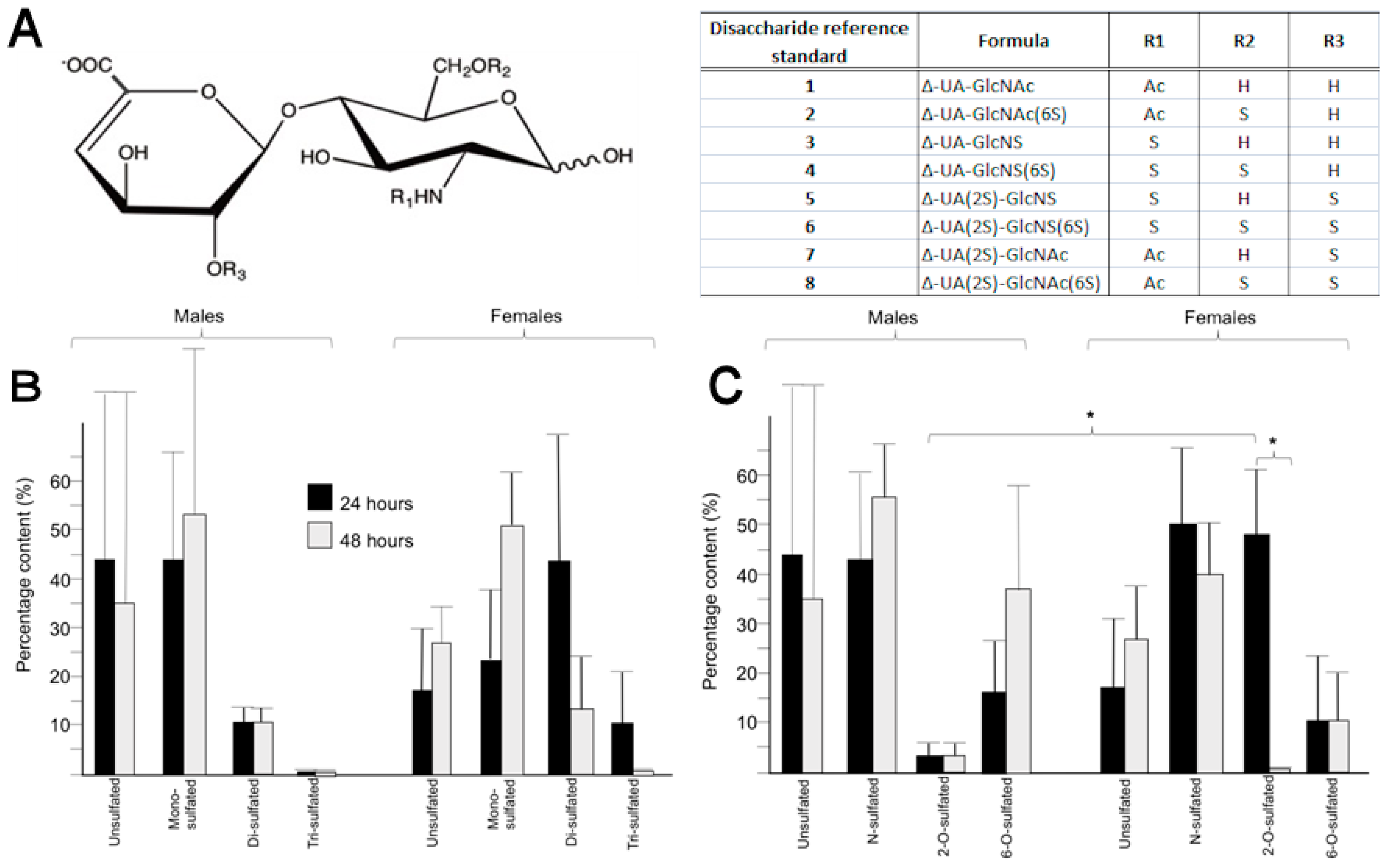
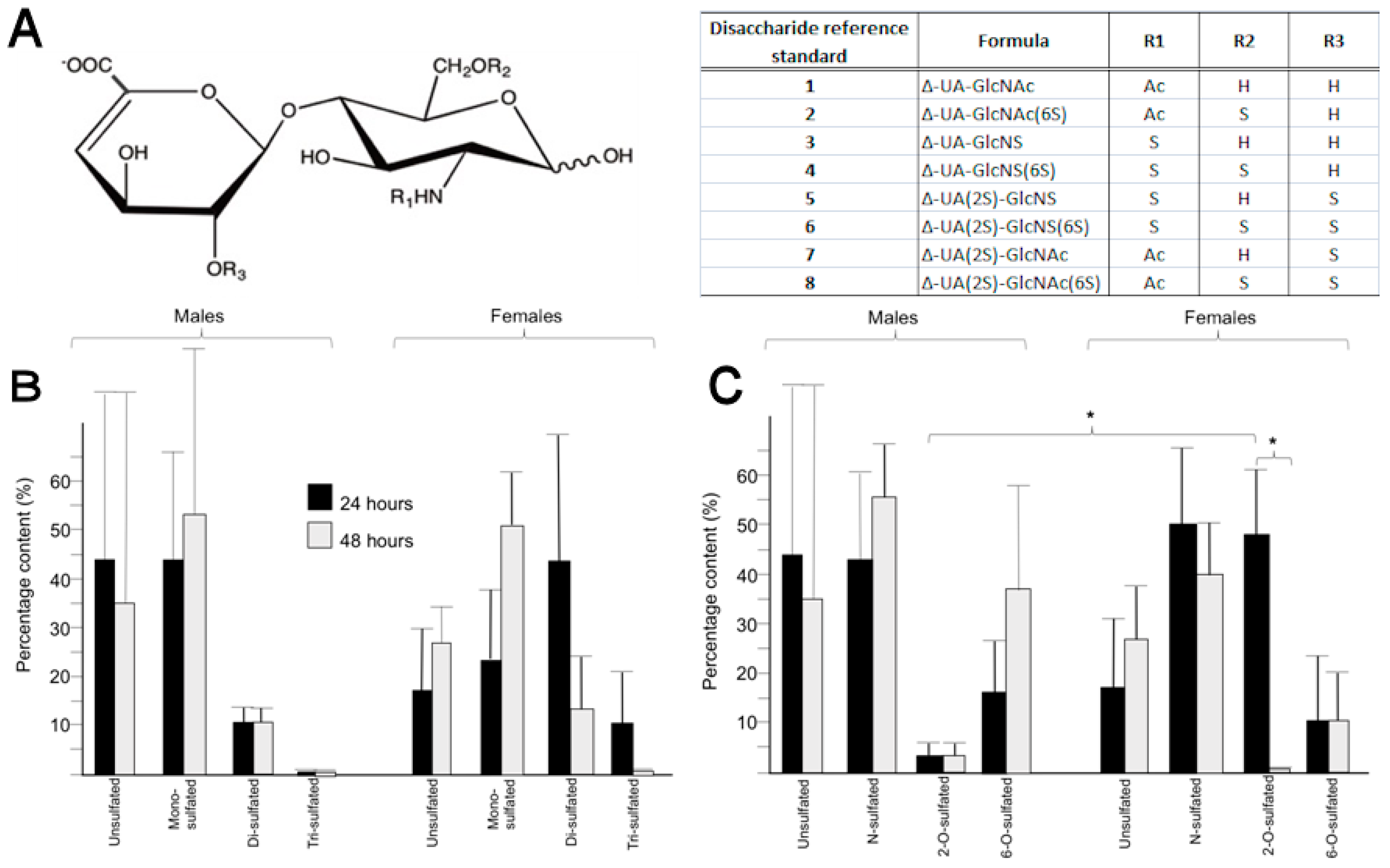
2.4. Heparan Sulphate from the Peritrophic Matrix Is Characterised by Moderate Levels of Sulphation
2.5. Differences in the Position of Sulphate Group Substitution, Particularly 2-O-Sulphation, Occur between Sexes and over Time
3. Discussion
4. Materials and Methods
4.1. Cultivation of Tsetse Fly (Glossina morsitans morsitans)
4.2. Isolation of Peritrophic Matrices
4.3. Heparinase Treatment of Peritrophic Matrices
4.4. Dot Blots of Heparinase-Digested Peritrophic Matrices
4.5. In Silico Sequence Analysis
4.6. Fluorescent Labeling
4.7. Thin Layer Chromatography
4.8. High Performance Anion Exchange Chromatography
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Service, M. Medical Entomology, 5th ed.; Cambridge University Press: Cambridge, UK, 2012. [Google Scholar]
- Peacock, L.; Ferris, V.; Sharma, V.; Sunter, J.; Bailey, M.; Carrington, M.; Gibson, W. Identification of the meiotic life cycle stage of Trypanosoma brucei in the tsetse fly. Proc. Natl. Acad. Sci. USA 2011, 108, 3671–3676. [Google Scholar] [CrossRef] [PubMed]
- Miller, N.; Lehane, M.J. Ionic environment and the permeability properties of the peritrophic membrane of Glossina morsitans morsitans. J. Insect Physiol. 1993, 39, 139–144. [Google Scholar] [CrossRef]
- Rose, C.; Belomonte, R.; Armstrong, S.D.; Molyneux, G.; Haines, L.R.; Lehane, M.J.; Wastling, J.; Acosta-Serrano, A. An Investigation into the Protein Composition of the Teneral Glossina morsitans morsitans Peritrophic Matrix. PLoS Negl. Trop. Dis. 2014, 8, e2691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehane, M.J.; Allingham, P.G.; Weglicki, P. Composition of the peritrophic matrix of the tsetse fly, Glossina morsitans morsitans. Cell Tissue Res. 1996, 283, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Abubakar, L.U.; Bulimo, W.D.; Mulaa, F.J.; Osir, E.O. Molecular characterization of a tsetse fly mid-gut proteolytic lectin that mediates differentiation of African trypanosomes. Insect Biochem. Mol. Biol. 2006, 36, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Hao, Z.; Kasumba, I.; Lehane, M.J.; Gibson, W.C.; Kwon, J.; Aksoy, S. tsetse immune responses and trypanosome transmission: Implications for the development of tsetse-based strategies to reduce trypa-nosomiasis. Proc. Natl. Acad. Sci. USA 2001, 98, 12648–12653. [Google Scholar] [CrossRef] [PubMed]
- Weiss, B.L.; Wang, J.; Maltz, M.A.; Wu, Y.; Aksoy, S. Trypanosome infection establishment in the tsetse fly gut is influenced by microbiome-regulated host immune barriers. PLoS Pathog. 2013, 9, e1003318. [Google Scholar] [CrossRef] [PubMed]
- Weiss, B.L.; Savage, A.F.; Griffith, B.C.; Wu, Y.; Aksoy, S. The peritrophic matrix mediates differential infection outcomes in the tsetse fly gut following challenge with commensal, pathogenic, and parasitic microbes. J. Immunol. 2014, 193, 773–782. [Google Scholar] [CrossRef] [PubMed]
- Aksoy, E.; Vigneron, A.; Bing, X.; Zhao, X.; O’Neill, M.; Wu, Y.N.; Bangs, J.D.; Weiss, B.L.; Aksoy, S. Mammalian African trypanosome VSG coat enhances tsetse’s vector competence. Proc. Natl. Acad. Sci. USA 2016, 113, 6961–6966. [Google Scholar] [CrossRef] [PubMed]
- International Glossina Genome Initiative. Genome sequence of the tsetse fly (Glossina morsitans): Vector of African trypanosomiasis. Science 2014, 344, 380–386. [Google Scholar]
- Calvet, C.M.; Toma, L.; De Souza, F.R.; Meirelles, M.D.E.; Pereira, M.C. Heparan sulphate proteoglycans mediate the invasion of cardiomyocytes by Trypanosoma cruzi. J. Eukaryot. Microbiol. 2003, 50, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Rostand, K.S.; Esko, J.D. Microbial adherence to and invasion through proteoglycans. Infect. Immun. 1997, 65, 1–8. [Google Scholar] [PubMed]
- Dale, C.; Maudlin, I. Sodalis gen. nov. and Sodalis glossinidius sp. nov. a microaerophilic secondary endosymbiont of the tsetse fly Glossina morsitans morsitans. Int. J. Syst. Bacteriol. 1999, 49, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Haines, L.R.; Lehane, S.M.; Pearson, T.W.; Lehane, M.J. Tsetse EP protein protects the fly midgut from trypanosome establishment. PLoS Pathog. 2010, 6, e1000793. [Google Scholar] [CrossRef] [PubMed]
- Kamimura, K.; Maeda, N.; Nakato, H. In vivo manipulation of heparan sulphate structure and its effect on Drosophila development. Glycobiology 2011, 21, 607–618. [Google Scholar] [CrossRef] [PubMed]
- Bax, X.; Van der Schueren, B.; Cassiman, J.J.; Van der Berghe, H.; David, G. Differential expression of multiple cell-surface heparan sulphate proteoglycans during embryonic tooth development. J. Histochem. Cytochem. 1994, 42, 1043–1054. [Google Scholar]
- David, G.; Bai, X.M.; Van der Schueren, B.; Cassiman, J.J.; Van den Berghe, H. Developmental chang-es in heparan sulphate expression: In situ detection with mAbs. J. Cell Biol. 1992, 119, 961–975. [Google Scholar] [CrossRef] [PubMed]
- Desai, U.R.; Wang, H.-M.; Linhardt, R.J. Specificity studies on the heparin lyases from Flavobacterium heparinium. Biochemistry 1993, 32, 8140–8145. [Google Scholar] [CrossRef] [PubMed]
- Skidmore, M.A.; Guimond, S.E.; Dumax-Vorzet, A.F.; Atrih, A.; Yates, E.A.; Turnbull, J.E. High sensitivity separation and detection of heparan sulphate disaccharides. J. Chromatogr. A 2006, 1135, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Powell, A.K.; Yates, E.A.; Fernig, D.G.; Turnbull, J.E. Interactions of heparin/heparan sulphate with pro-teins: Appraisal of structural factors and experimental approaches. Glycobiology 2004, 14, 17R–30R. [Google Scholar] [CrossRef] [PubMed]
- Perrimon, N.; Bernfield, M. Specificities of HS proteoglycans in developmental processes. Nature 2000, 404, 725–728. [Google Scholar] [CrossRef] [PubMed]
- Kusche-Gullberg, M.; Nybakken, K.; Perrimon, N.; Lindahl, U. Drosophila heparan sulphate, a novel design. J. Biol. Chem. 2012, 287, 21950–21956. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, S.; Li, C.; Zhao, S. Phylogeny of genus Glossina (Diptera: Glossinidae) according to ITS2 sequences. Sci. China C 1999, 42, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Ford, J. The Geographical Distribution of Glossina; Allen & Unwin: London, UK, 1970. [Google Scholar]
- Bai, X.; Bame, K.J.; Habuchi, H.; Kimata, K.; Esko, J.D. Turnover of heparan sulphate depends on 2-O-sulphation of uronic acids. J. Biol. Chem. 1997, 272, 23172–23179. [Google Scholar] [CrossRef] [PubMed]
- Dinglasan, R.R.; Alaganan, A.; Ghosh, A.K.; Saito, A.; van Kuppevelt, T.H.; Jacobs-Lorena, M. Plasmodium falciparum ookinetes require mosquito midgut chondroitin sulphate proteoglycans for cell invasion. Proc. Natl. Acad. Sci. USA 2007, 104, 15882–15887. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Giraldo-Calderón, G.I.; Emrich, S.J.; MacCallum, J.M.; Maslen, G.; Dialynas, E.; Topalis, P.; Ho, N.; Gesing, S.; Collins, F.H.; Lawson, D.; et al. VectorBase: An updated bioinformatics resource for invertebrate vectors and other organisms related with human diseases. Nucleic Acids Res. 2015, 43, D707–D713. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Protein (Uniprot Accession Number) | EST Entry | E-Value | Identity (%) |
|---|---|---|---|
| Syndecan-1 (P18827) | EZ422907.1 | 5 × 10−9 | 52.2 |
| EXT1 (Q16394) | FM983148.1 | 4 × 10−48 | 46 |
| NDST-1 (P52848) | FM959941.1 | 2 × 10−17 | 29.2 |
| 2OST-1 (Q7LGA3) | DV604335.1 | 2 × 10−73 | 55.7 |
| 6OST-1 (O60243) | FM985074.1 | 1 × 10−36 | 67.4 |
| 3OST-1 (O14792) | FM959941.1 | 1 × 10−54 | 40.1 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rogerson, E.; Pelletier, J.; Acosta-Serrano, A.; Rose, C.; Taylor, S.; Guimond, S.; Lima, M.; Skidmore, M.; Yates, E. Variations in the Peritrophic Matrix Composition of Heparan Sulphate from the Tsetse Fly, Glossina morsitans morsitans. Pathogens 2018, 7, 32. https://doi.org/10.3390/pathogens7010032
Rogerson E, Pelletier J, Acosta-Serrano A, Rose C, Taylor S, Guimond S, Lima M, Skidmore M, Yates E. Variations in the Peritrophic Matrix Composition of Heparan Sulphate from the Tsetse Fly, Glossina morsitans morsitans. Pathogens. 2018; 7(1):32. https://doi.org/10.3390/pathogens7010032
Chicago/Turabian StyleRogerson, Evelyn, Julien Pelletier, Alvaro Acosta-Serrano, Clair Rose, Sarah Taylor, Scott Guimond, Marcelo Lima, Mark Skidmore, and Edwin Yates. 2018. "Variations in the Peritrophic Matrix Composition of Heparan Sulphate from the Tsetse Fly, Glossina morsitans morsitans" Pathogens 7, no. 1: 32. https://doi.org/10.3390/pathogens7010032