
Interactions of Methylotrophs with Plants and Other Heterotrophic Bacteria

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methanol-Utilizing Methylotrophs on Plants
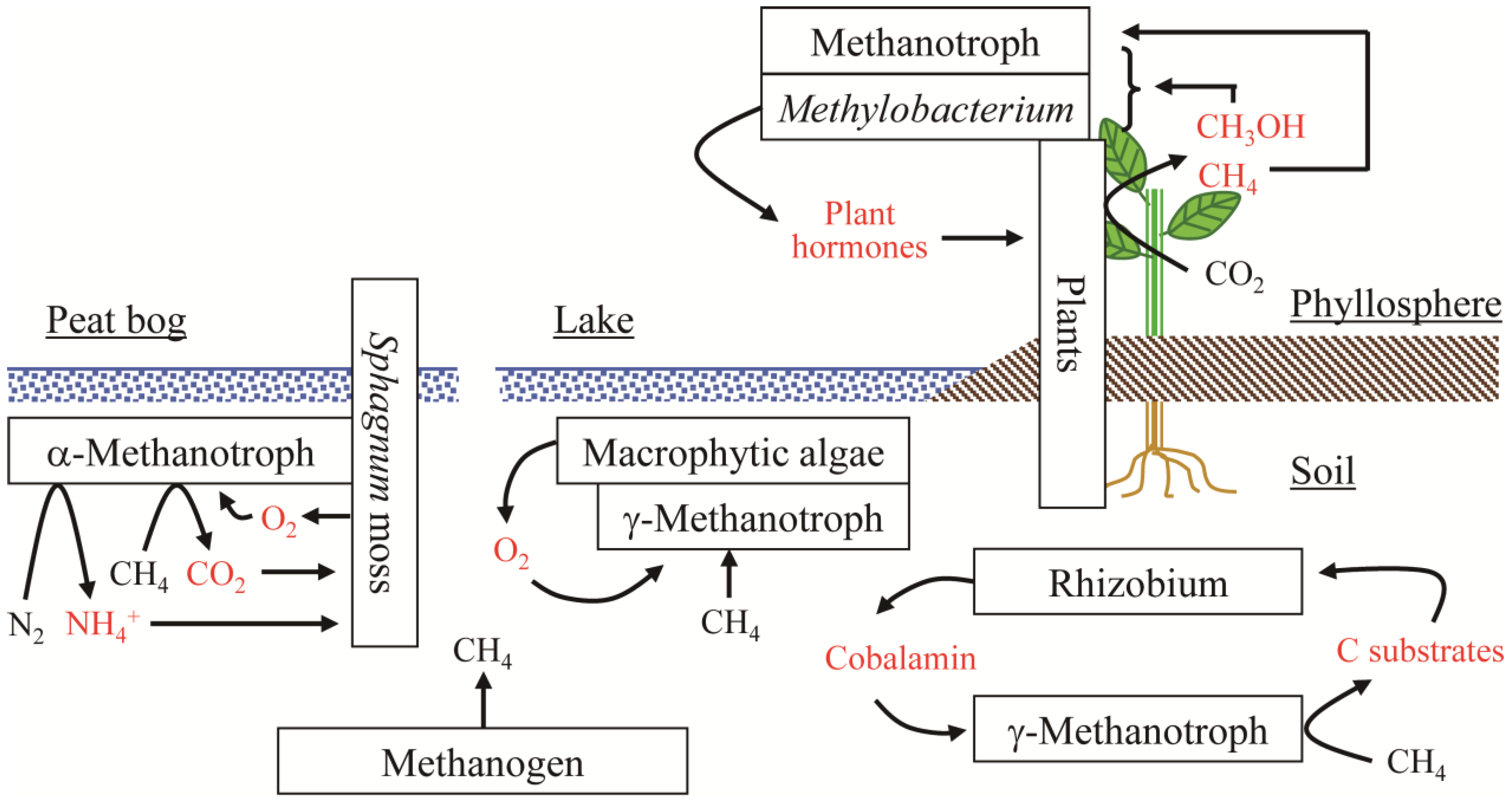
2.1. Positive Effects of Methylobacterium Species on Plant Growth
2.2. Methylobacterium Community in the Phyllosphere
3. Plant-Associated Methanotrophs
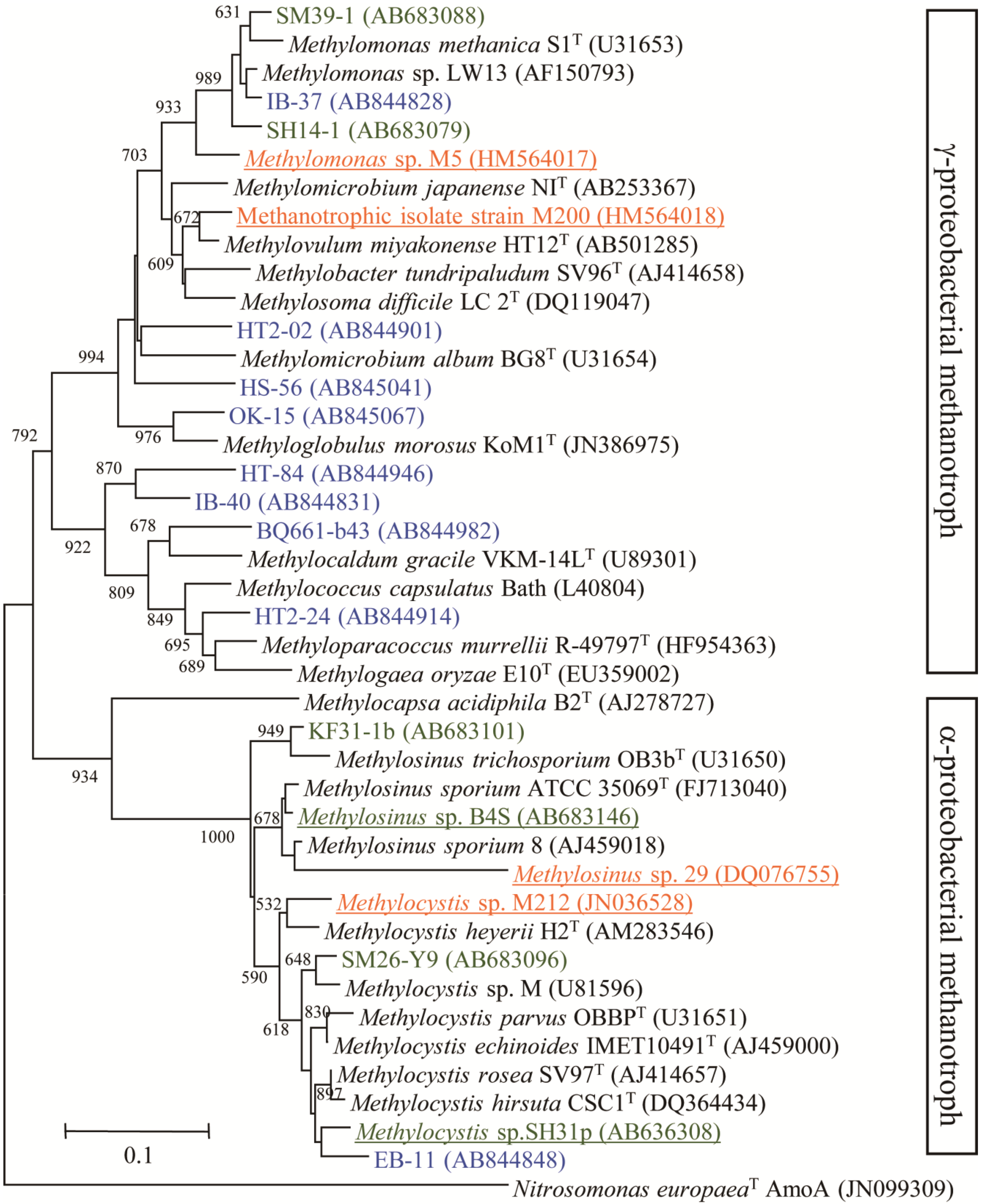
3.1. Characteristics of Methanotrophs Inhabiting the Phyllosphere

3.2. High Methane Oxidation by Methanotrophs on Aquatic Plants
4. Stimulation of Methanotrophic Activity by Heterotrophic Bacteria
5. Conclusions and Perspective

Acknowledgments
Conflicts of Interest
References
- IPCC. Climate Change 2007: The Physical Science Basis. Summary for Policymakers; Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Keppler, F.; Hamilton, J.T.G.; Brass, M.; Rockmann, T. Methane emissions from terrestrial plants under aerobic conditions. Nature 2006, 439, 187–191. [Google Scholar]
- Keppler, F.; Hamilton, J.T.G.; McRoberts, W.C.; Vigano, I.; Brass, M.; Rockmann, T. Methoxyl groups of plant pectin as a precursor of atmospheric methane: Evidence from deuterium labelling studies. New Phytol. 2008, 178, 808–814. [Google Scholar]
- Bruhn, D.; Moller, I.M.; Mikkelsen, T.N.; Ambus, P. Terrestrial plant methane production and emission. Physiol. Plant 2012, 144, 201–209. [Google Scholar]
- Galbally, I.E.; Kirstine, W. The production of methanol by flowering plants and the global cycle of methanol. J. Atmos. Chem. 2002, 43, 195–229. [Google Scholar]
- Nemecekmarshall, M.; Macdonald, R.C.; Franzen, F.J.; Wojciechowski, C.L.; Fall, R. Methanol emission from leaves: Enzymatic detection of gas-phase methanol and relation of methanol fluxes to stomatal conductance and leaf development. Plant Physiol. 1995, 108, 1359–1368. [Google Scholar]
- Macdonald, R.C.; Fall, R. Detection of substantial emissions of methanol from plants to the atmosphere. Atmos. Environ. 1993, 27, 1709–1713. [Google Scholar]
- Nazaries, L.; Murrell, J.C.; Millard, P.; Baggs, L.; Singh, B.K. Methane, microbes and models: Fundamental understanding of the soil methane cycle for future predictions. Environ. Microbiol. 2013, 15, 2395–2417. [Google Scholar]
- Ettwig, K.F.; Butler, M.K.; Le Paslier, D.; Pelletier, E.; Mangenot, S.; Kuypers, M.M.; Schreiber, F.; Dutilh, B.E.; Zedelius, J.; de Beer, D.; et al. Nitrite-driven anaerobic methane oxidation by oxygenic bacteria. Nature 2010, 464, 543–548. [Google Scholar]
- Semrau, J.D.; DiSpirito, A.A.; Vuilleumier, S. Facultative methanotrophy: False leads, true results, and suggestions for future research. FEMS Microbiol. Lett. 2011, 323, 1–12. [Google Scholar]
- Semrau, J.D.; DiSpirito, A.A.; Yoon, S. Methanotrophs and copper. FEMS Microbiol. Rev. 2010, 34, 496–531. [Google Scholar]
- Dedysh, S.N.; Knief, C.; Dunfield, P.F. Methylocella species are facultatively methanotrophic. J. Bacteriol. 2005, 187, 4665–4670. [Google Scholar]
- Dunfield, P.F.; Belova, S.E.; Vorob’ev, A.V.; Cornish, S.L.; Dedysh, S.N. Methylocapsa aurea sp. nov., a facultative methanotroph possessing a particulate methane monooxygenase, and emended description of the genus Methylocapsa. Int. J. Syst. Evol. Microbiol. 2010, 60, 2659–2664. [Google Scholar]
- Belova, S.E.; Baani, M.; Suzina, N.E.; Bodelier, P.L.E.; Liesack, W.; Dedysh, S.N. Acetate utilization as a survival strategy of peat-inhabiting Methylocystis spp. Environ. Microbiol. Rep. 2010, 3, 36–46. [Google Scholar]
- Kolb, S. Aerobic methanol-oxidizing bacteria in soil. FEMS Microbiol. Lett. 2009, 300, 1–10. [Google Scholar]
- Guenther, A. The contribution of reactive carbon emissions from vegetation to the carbon balance of terrestrial ecosystems. Chemosphere 2002, 49, 837–844. [Google Scholar]
- Aronson, E.L.; Allison, S.D.; Helliker, B.R. Environmental impacts on the diversity of methane-cycling microbes and their resultant function. Front. Microbiol. 2013, 4, 225. [Google Scholar]
- Murase, J.; Frenzel, P. A methane-driven microbial food web in a wetland rice soil. Environ. Microbiol. 2007, 9, 3025–3034. [Google Scholar]
- Petersen, J.M.; Dubilier, N. Methanotrophic symbioses in marine invertebrates. Environ. Microbiol. Rep. 2009, 1, 319–335. [Google Scholar]
- Qiu, Q.; Conrad, R.; Lu, Y. Cross-feeding of methane carbon among bacteria on rice roots revealed by DNA-stable isotope probing. Environ. Microbiol. Rep. 2009, 1, 355–361. [Google Scholar]
- Radajewski, S.; Webster, G.; Reay, D.S.; Morris, S.A.; Ineson, P.; Nedwell, D.B.; Prosser, J.I.; Murrell, J.C. Identification of active methylotroph populations in an acidic forest soil by stable-isotope probing. Microbiology 2002, 148, 2331–2342. [Google Scholar]
- He, R.; Wooller, M.J.; Pohlman, J.W.; Quensen, J.; Tiedje, J.M.; Leigh, M.B. Diversity of active aerobic methanotrophs along depth profiles of arctic and subarctic lake water column and sediments. ISME J. 2012, 6, 1937–1948. [Google Scholar]
- Raghoebarsing, A.A.; Smolders, A.J.; Schmid, M.C.; Rijpstra, W.I.; Wolters-Arts, M.; Derksen, J.; Jetten, M.S.; Schouten, S.; Sinninghe Damste, J.S.; Lamers, L.P.; et al. Methanotrophic symbionts provide carbon for photosynthesis in peat bogs. Nature 2005, 436, 1153–1156. [Google Scholar]
- Corpe, W.A.; Basile, D.V. Methanol-utilizing bacteria associated with green plants. Dev. Ind. Microbiol. 1982, 23, 483–493. [Google Scholar]
- Lopez-Velasco, G.; Welbaum, G.E.; Boyer, R.R.; Mane, S.P.; Ponder, M.A. Changes in spinach phylloepiphytic bacteria communities following minimal processing and refrigerated storage described using pyrosequencing of 16S rRNA amplicons. J. Appl. Microbiol. 2011, 110, 1203–1214. [Google Scholar]
- Reisberg, E.E.; Hildebrandt, U.; Riederer, M.; Hentschel, U. Distinct phyllosphere bacterial communities on Arabidopsis wax mutant leaves. PLoS ONE 2013, 8, e78613. [Google Scholar]
- Reisberg, E.E.; Hildebrandt, U.; Riederer, M.; Hentschel, U. Phyllosphere bacterial communities of trichome-bearing and trichomeless Arabidopsis thaliana leaves. Anton. Leeuw. 2012, 101, 551–560. [Google Scholar]
- Vorholt, J.A. Microbial life in the phyllosphere. Nat. Rev. Microbiol. 2012, 10, 828–840. [Google Scholar]
- Fedorov, D.N.; Doronina, N.V.; Trotsenko, Y.A. Phytosymbiosis of aerobic methylobacteria: New facts and views. Microbiology 2011, 80, 443–454. [Google Scholar]
- Trotsenko, Y.A.; Ivanova, E.G.; Doronina, N.V. Aerobic methylotrophic bacteria as phytosymbionts. Microbiology 2001, 70, 623–632. [Google Scholar]
- Madhaiyan, M.; Poonguzhali, S.; Senthilkumar, M.; Seshadri, S.; Chung, H.Y.; Yang, J.C.; Sundaram, S.; Sa, T.M. Growth promotion and induction of systemic resistance in rice cultivar Co-47 (Oryza sativa L.) by Methylobacterium spp. Bot. Bull. Acad. Sinica. 2004, 45, 315–324. [Google Scholar]
- Tani, A.; Takai, Y.; Suzukawa, I.; Akita, M.; Murase, H.; Kimbara, K. Practical application of methanol-mediated mutualistic symbiosis between Methylobacterium species and a roof greening moss, Racomitrium japonicum. PLoS ONE 2012, 7, e33800. [Google Scholar]
- Meena, K.K.; Kumar, M.; Kalyuzhnaya, M.G.; Yandigeri, M.S.; Singh, D.P.; Saxena, A.K.; Arora, D.K. Epiphytic pink-pigmented methylotrophic bacteria enhance germination and seedling growth of wheat (Triticum aestivum) by producing phytohormone. Anton. Leeuw. 2012, 101, 777–786. [Google Scholar]
- Madhaiyan, M.; Poonguzhali, S.; Kang, B.G.; Lee, Y.J.; Chung, J.B.; Sa, T.M. Effect of co-inoculation of methylotrophic Methylobacterium oryzae with Azospirillum brasilense and Burkholderia pyrrocinia on the growth and nutrient uptake of tomato, red pepper and rice. Plant Soil 2010, 328, 71–82. [Google Scholar]
- Verginer, M.; Siegmund, B.; Cardinale, M.; Muller, H.; Choi, Y.; Miguez, C.B.; Leitner, E.; Berg, G. Monitoring the plant epiphyte Methylobacterium extorquens DSM 21961 by real-time PCR and its influence on the strawberry flavor. FEMS Microbiol. Ecol. 2010, 74, 136–145. [Google Scholar]
- Kwak, M.J.; Jeong, H.; Madhaiyan, M.; Lee, Y.; Sa, T.M.; Oh, T.K.; Kim, J.F. Genome information of Methylobacterium oryzae, a plant-probiotic methylotroph in the phyllosphere. PLoS ONE 2014, 9, e106704. [Google Scholar]
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; Ver Loren van Themaat, E.; Schulze-Lefert, P. Structure and functions of the bacterial microbiota of plants. Annu. Rev. Plant Biol. 2013, 64, 807–838. [Google Scholar]
- Morgan, J.V.; Tukey, H.B. Characterization of leachate from plant foliage. Plant Physiol. 1964, 39, 590–593. [Google Scholar]
- Fiala, V.; Glad, C.; Martin, M.; Jolivet, E.; Derridj, S. Occurrence of soluble carbohydrates on the phylloplane of maize (Zea Mays L.): Variations in relation to leaf heterogeneity and position on the plant. New Phytol. 1990, 115, 609–615. [Google Scholar]
- Kawaguchi, K.; Yurimoto, H.; Oku, M.; Sakai, Y. Yeast methylotrophy and autophagy in a methanol-oscillating environment on growing Arabidopsis thaliana leaves. PLoS ONE 2011, 6, e25257. [Google Scholar]
- Sy, A.; Timmers, A.C.; Knief, C.; Vorholt, J.A. Methylotrophic metabolism is advantageous for Methylobacterium extorquens during colonization of Medicago truncatula under competitive conditions. Appl. Environ. Microbiol. 2005, 71, 7245–7252. [Google Scholar]
- Finkel, O.M.; Burch, A.Y.; Elad, T.; Huse, S.M.; Lindow, S.E.; Post, A.F.; Belkin, S. Distance-decay relationships partially determine diversity patterns of phyllosphere bacteria on Tamarix trees across the Sonoran Desert. Appl. Environ. Microbiol. 2012, 78, 6187–6193. [Google Scholar]
- Siefert, A.; Fridley, J.D.; Ritchie, M.E. Community functional responses to soil and climate at multiple spatial scales: When does intraspecific variation matter? PLoS ONE 2014, 9, e111189. [Google Scholar]
- Redford, A.J.; Bowers, R.M.; Knight, R.; Linhart, Y.; Fierer, N. The ecology of the phyllosphere: Geographic and phylogenetic variability in the distribution of bacteria on tree leaves. Environ. Microbiol. 2010, 12, 2885–2893. [Google Scholar]
- Knief, C.; Ramette, A.; Frances, L.; Alonso-Blanco, C.; Vorholt, J.A. Site and plant species are important determinants of the Methylobacterium community composition in the plant phyllosphere. ISME J. 2010, 4, 719–728. [Google Scholar]
- Mizuno, M.; Yurimoto, H.; Yoshida, N.; Iguchi, H.; Sakai, Y. Distribution of pink-pigmented facultative methylotrophs on leaves of vegetables. Biosci. Biotechnol. Biochem. 2012, 76, 578–580. [Google Scholar]
- Knief, C.; Frances, L.; Vorholt, J.A. Competitiveness of diverse Methylobacterium strains in the phyllosphere of Arabidopsis thaliana and identification of representative models, including M. extorquens PA1. Microb. Ecol. 2010, 60, 440–452. [Google Scholar]
- Mizuno, M.; Yurimoto, H.; Iguchi, H.; Tani, A.; Sakai, Y. Dominant colonization and inheritance of Methylobacterium sp. strain OR01 on perilla plants. Biosci. Biotechnol. Biochem. 2013, 77, 1533–1538. [Google Scholar]
- Lindow, S.E.; Brandl, M.T. Microbiology of the phyllosphere. Appl. Environ. Microbiol. 2003, 69, 1875–1883. [Google Scholar]
- Iguchi, H.; Sato, I.; Sakakibara, M.; Yurimoto, H.; Sakai, Y. Distribution of methanotrophs in the phyllosphere. Biosci. Biotechnol. Biochem. 2012, 76, 1580–1583. [Google Scholar]
- Doronina, N.V.; Ivanova, E.G.; Suzina, N.E.; Trotsenko, Y.A. Methanotrophs and methylobacteria are found in woody plant tissues within the winter period. Microbiology 2004, 73, 702–709. [Google Scholar]
- Ikeda, S.; Anda, M.; Inaba, S.; Eda, S.; Sato, S.; Sasaki, K.; Tabata, S.; Mitsui, H.; Sato, T.; Shinano, T.; et al. Autoregulation of nodulation interferes with impacts of nitrogen fertilization levels on the leaf-associated bacterial community in soybeans. Appl. Environ. Microbiol. 2011, 77, 1973–1980. [Google Scholar]
- Finkel, O.M.; Burch, A.Y.; Lindow, S.E.; Post, A.F.; Belkin, S. Geographical location determines the population structure in phyllosphere microbial communities of a salt-excreting desert tree. Appl. Environ. Microbiol. 2011, 77, 7647–7655. [Google Scholar]
- Knief, C.; Delmotte, N.; Chaffron, S.; Stark, M.; Innerebner, G.; Wassmann, R.; von Mering, C.; Vorholt, J.A. Metaproteogenomic analysis of microbial communities in the phyllosphere and rhizosphere of rice. ISME J. 2012, 6, 1378–1390. [Google Scholar]
- Hanson, R.S.; Hanson, T.E. Methanotrophic bacteria. Microbiol. Rev. 1996, 60, 439–471. [Google Scholar]
- Yoshida, N.; Iguchi, H.; Yurimoto, H.; Murakami, A.; Sakai, Y. Aquatic plant surface as a niche for methanotrophs. Front. Microbiol. 2014, 5, 30. [Google Scholar]
- Kip, N.; Ouyang, W.; van Winden, J.; Raghoebarsing, A.; van Niftrik, L.; Pol, A.; Pan, Y.; Bodrossy, L.; van Donselaar, E.G.; Reichart, G.J.; et al. Detection, isolation, and characterization of acidophilic methanotrophs from Sphagnum mosses. Appl. Environ. Microbiol. 2011, 77, 5643–5654. [Google Scholar]
- Iguchi, H.; Sato, I.; Yurimoto, H.; Sakai, Y. Stress resistance and C1 metabolism involved in plant colonization of a methanotroph Methylosinus sp. B4S. Arch. Microbiol. 2013, 195, 717–726. [Google Scholar]
- Lieberman, R.L.; Rosenzweig, A.C. Biological methane oxidation: Regulation, biochemistry, and active site structure of particulate methane monooxygenase. Crit. Rev. Biochem. Mol. Biol. 2004, 39, 147–164. [Google Scholar]
- Gourion, B.; Rossignol, M.; Vorholt, J.A. A proteomic study of Methylobacterium extorquens reveals a response regulator essential for epiphytic growth. Proc. Natl. Acad. Sci. USA 2006, 103, 13186–13191. [Google Scholar]
- Gourion, B.; Francez-Charlot, A.; Vorholt, J.A. PhyR is involved in the general stress response of Methylobacterium extorquens AM1. J. Bacteriol. 2008, 190, 1027–1035. [Google Scholar]
- Kaczmarczyk, A.; Campagne, S.; Danza, F.; Metzger, L.C.; Vorholt, J.A.; Francez-Charlot, A. Role of Sphingomonas sp. strain Fr1 PhyR-NepR-sEcfG cascade in general stress response and identification of a negative regulator of PhyR. J. Bacteriol. 2011, 193, 6629–6638. [Google Scholar]
- Staron, A.; Mascher, T. General stress response in a-proteobacteria: PhyR and beyond. Mol. Microbiol. 2010, 78, 271–277. [Google Scholar]
- Borrel, G.; Jezequel, D.; Biderre-Petit, C.; Morel-Desrosiers, N.; Morel, J.P.; Peyret, P.; Fonty, G.; Lehours, A.C. Production and consumption of methane in freshwater lake ecosystems. Res. Microbiol. 2011, 162, 832–847. [Google Scholar]
- Van Bodegom, P.; Stams, F.; Mollema, L.; Boeke, S.; Leffelaar, P. Methane oxidation and the competition for oxygen in the rice rhizosphere. Appl. Environ. Microbiol. 2001, 67, 3586–3597. [Google Scholar]
- Wu, L.; Ma, K.; Lu, Y. Rice roots select for type I methanotrophs in rice field soil. Syst. Appl. Microbiol. 2009, 32, 421–428. [Google Scholar]
- Eller, G.; Frenzel, P. Changes in activity and community structure of methane-oxidizing bacteria over the growth period of rice. Appl. Environ. Microbiol. 2001, 67, 2395–2403. [Google Scholar]
- Heilman, M.A.; Carlton, R.G. Methane oxidation associated with submersed vascular macrophytes and its impact on plant diffusive methane flux. Biogeochemistry 2001, 52, 207–224. [Google Scholar]
- King, G.M. Associations of methanotrophs with the roots and rhizomes of aquatic vegetation. Appl. Environ. Microbiol. 1994, 60, 3220–3227. [Google Scholar]
- Calhoun, A.; King, G.M. Regulation of root-associated methanotrophy by oxygen availability in the rhizosphere of two aquatic macrophytes. Appl. Environ. Microbiol. 1997, 63, 3051–3058. [Google Scholar]
- Sorrell, B.K.; Downes, M.T.; Stanger, C.L. Methanotrophic bacteria and their activity on submerged aquatic macrophytes. Aquat. Bot. 2002, 72, 107–119. [Google Scholar]
- Larmola, T.; Leppanen, S.M.; Tuittila, E.S.; Aarva, M.; Merila, P.; Fritze, H.; Tiirola, M. Methanotrophy induces nitrogen fixation during peatland development. Proc. Natl. Acad. Sci. USA 2014, 111, 734–739. [Google Scholar]
- Kip, N.; van Winden, J.F.; Pan, Y.; Bodrossy, L.; Reichart, G.J.; Smolders, A.J.P.; Jetten, M.S.M.; Damste, J.S.S.; Op den Camp, H.J.M. Global prevalence of methane oxidation by symbiotic bacteria in peat-moss ecosystems. Nat. Geosci. 2010, 3, 617–621. [Google Scholar]
- Liebner, S.; Zeyer, J.; Wagner, D.; Schubert, C.; Pfeiffer, E.M.; Knoblauch, C. Methane oxidation associated with submerged brown mosses reduces methane emissions from Siberian polygonal tundra. J. Ecol. 2011, 99, 914–922. [Google Scholar]
- Putkinen, A.; Larmola, T.; Tuomivirta, T.; Siljanen, H.M.P.; Bodrossy, L.; Tuittila, E.S.; Fritze, H. Water dispersal of methanotrophic bacteria maintains functional methane oxidation in Sphagnum mosses. Front. Microbiol. 2012, 3, 15. [Google Scholar]
- Rahalkar, M.; Deutzmann, J.; Schink, B.; Bussmann, I. Abundance and activity of methanotrophic bacteria in littoral and profundal sediments of lake constance (Germany). Appl. Environ. Microbiol. 2009, 75, 119–126. [Google Scholar]
- Tsutsumi, M.; Kojima, H.; Fukui, M. Vertical profiles of abundance and potential activity of methane-oxidizing bacteria in sediment of Lake Biwa, Japan. Microb. Environ. 2012, 27, 67–71. [Google Scholar]
- Chen, Y.; Dumont, M.G.; McNamara, N.P.; Chamberlain, P.M.; Bodrossy, L.; Stralis-Pavese, N.; Murrell, J.C. Diversity of the active methanotrophic community in acidic peatlands as assessed by mRNA and SIP-PLFA analyses. Environ. Microbiol. 2008, 10, 446–459. [Google Scholar]
- Iguchi, H.; Yurimoto, H.; Sakai, Y. Stimulation of methanotrophic growth in co-cultures by cobalamin excreted by rhizobia. Appl. Environ. Microbiol. 2011, 77, 8509–8515. [Google Scholar]
- Bowman, J. The methanotrophs—The families Methylococcaceae and Mzethylocystaceae. In The Prokaryotes; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; pp. 266–289. [Google Scholar]
- Wilkinson, T.G.; Topiwala, H.H.; Hamer, G. Interactions in a mixed bacterial population growing on methane in continuous culture. Biotechnol. Bioeng. 1974, 16, 41–59. [Google Scholar]
- Lamb, S.C.; Garver, J.C. Batch- and continuous-culture studies of a methane-utilizing mixed culture. Biotechnol. Bioeng. 1980, 22, 2097–2118. [Google Scholar]
- Iguchi, H.; Yurimoto, H.; Sakai, Y.; Kyoto University, Kyoto, Japan. Unpublished work. 2012.
- Colby, J.; Stirling, D.I.; Dalton, H. The soluble methane mono-oxygenase of Methylococcus capsulatus (Bath). Its ability to oxygenate n-alkanes, n-alkenes, ethers, and alicyclic, aromatic and heterocyclic compounds. Biochem. J. 1977, 165, 395–402. [Google Scholar]
- Hoefman, S.; van der Ha, D.; Iguchi, H.; Yurimoto, H.; Sakai, Y.; Boon, N.; Vandamme, P.; Heylen, K.; de Vos, P. Methyloparacoccus murrellii gen. nov., sp. nov., a methanotroph isolated from pond water. Int. J. Syst. Evol. Microbiol. 2014, 64, 2100–2107. [Google Scholar]
- Roth, J.R.; Lawrence, J.G.; Bobik, T.A. Cobalamin (coenzyme B12): Synthesis and biological significance. Annu. Rev. Microbiol. 1996, 50, 137–181. [Google Scholar]
- Sanudo-Wilhelmy, S.A.; Gomez-Consarnau, L.; Suffridge, C.; Webb, E.A. The role of B vitamins in marine biogeochemistry. Annu. Rev. Mar. Sci. 2014, 6, 339–367. [Google Scholar]
- Stock, M.; Hoefman, S.; Kerckhof, F.M.; Boon, N.; de Vos, P.; de Baets, B.; Heylen, K.; Waegeman, W. Exploration and prediction of interactions between methanotrophs and heterotrophs. Res. Microbiol. 2013, 164, 1045–1054. [Google Scholar]
- Ho, A.; de Roy, K.; Thas, O.; de Neve, J.; Hoefman, S.; Vandamme, P.; Heylen, K.; Boon, N. The more, the merrier: Heterotroph richness stimulates methanotrophic activity. ISME J. 2014, 8, 1945–1948. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iguchi, H.; Yurimoto, H.; Sakai, Y. Interactions of Methylotrophs with Plants and Other Heterotrophic Bacteria. Microorganisms 2015, 3, 137-151. https://doi.org/10.3390/microorganisms3020137
Iguchi H, Yurimoto H, Sakai Y. Interactions of Methylotrophs with Plants and Other Heterotrophic Bacteria. Microorganisms. 2015; 3(2):137-151. https://doi.org/10.3390/microorganisms3020137
Chicago/Turabian StyleIguchi, Hiroyuki, Hiroya Yurimoto, and Yasuyoshi Sakai. 2015. "Interactions of Methylotrophs with Plants and Other Heterotrophic Bacteria" Microorganisms 3, no. 2: 137-151. https://doi.org/10.3390/microorganisms3020137