
Lactic Fermentation as an Efficient Tool to Enhance the Antioxidant Activity of Tropical Fruit Juices and Teas

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain and Culture Media
2.2. EPS Production
2.3. Kinetics of Acidification
2.4. Growth in Apple Juice
2.5. Preparation of Substrate
2.6. Fermentation
2.7. Qualitative Sensory Analysis and Determination of Fermentation Optimal Time
2.8. Shelf-Life Determination
2.9. Sugar Content Determination
2.10. Sensory Analysis
2.11. Determination of Total Phenolic Content (TPC)
2.12. 1,1-Diphenyl-2-Picrylhydrazyl (DPPH) Free-Radical Scavenging Activity (RSA)
2.13. Red Blood Cell Haemolysis Assay
2.14. Low Density Lipoprotein (LDL) Oxidation
2.15. Statistics
3. Results and Discussion
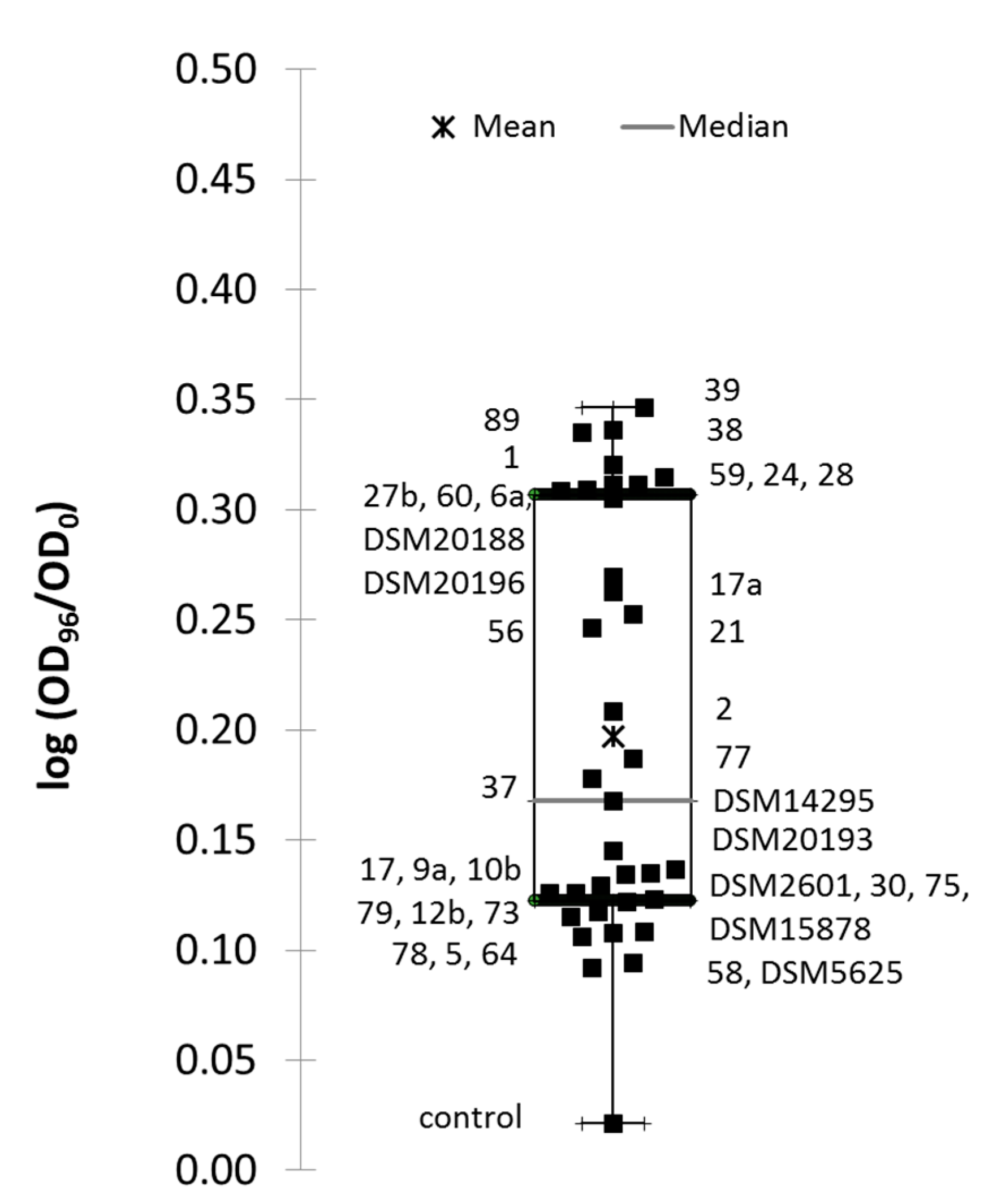
3.1. EPS Formation and Acidification Kinetics
3.2. Growth in Apple Juice
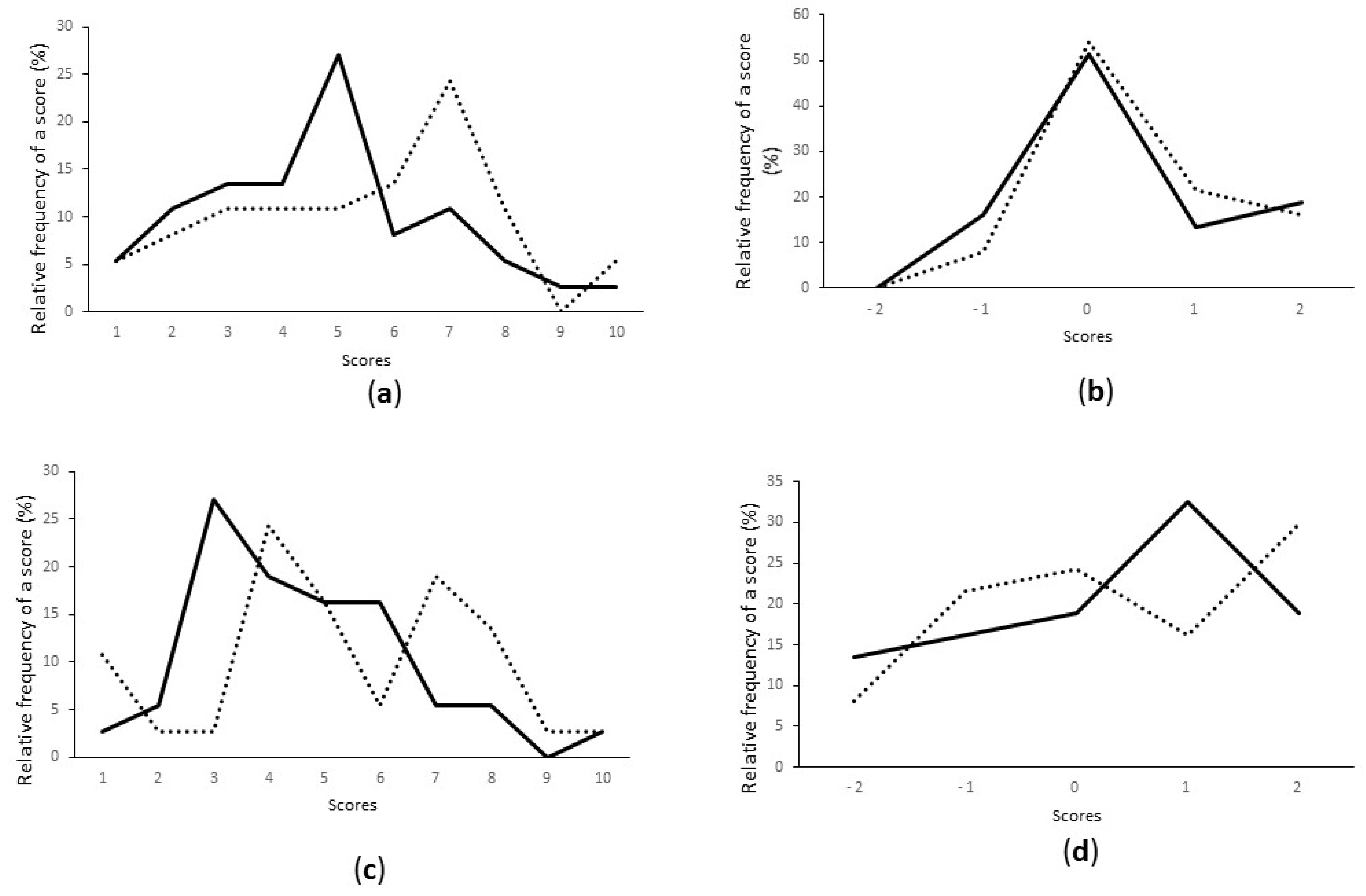
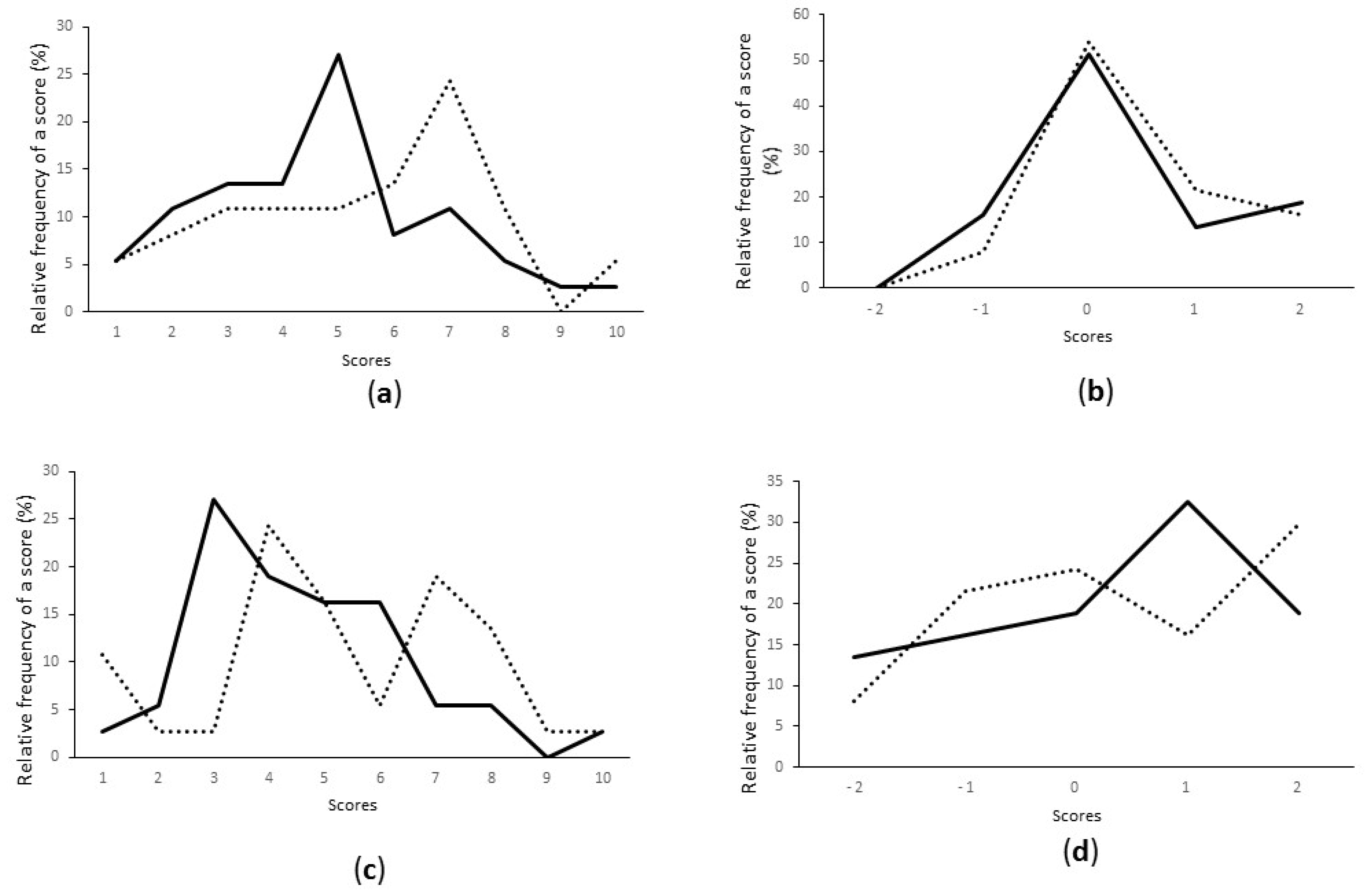
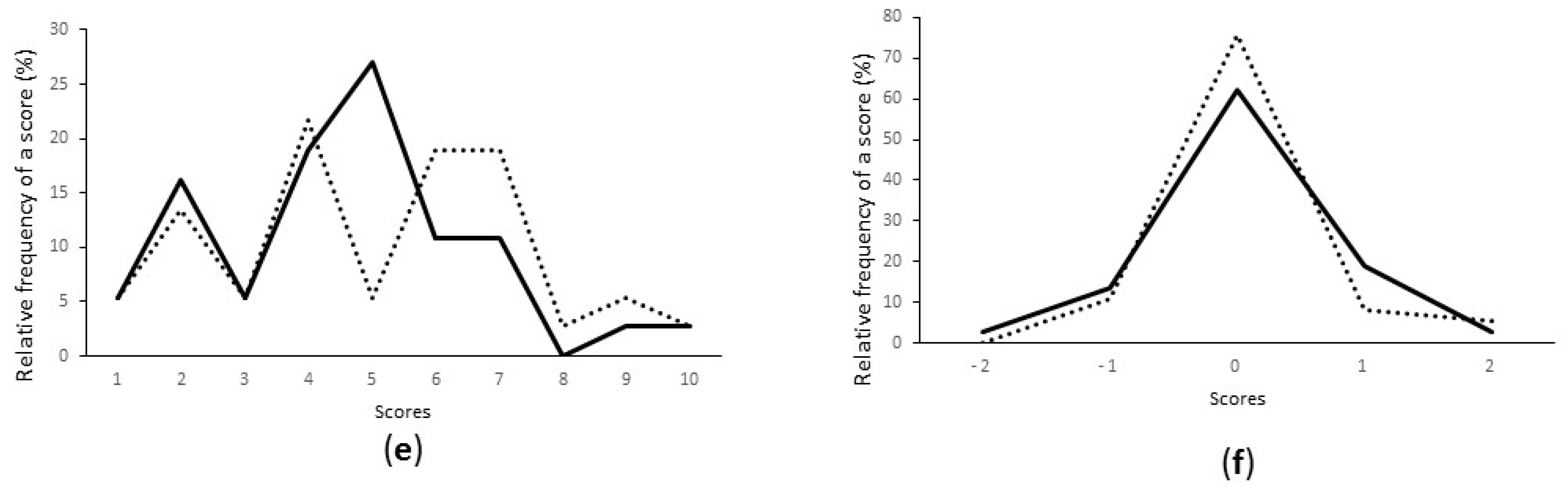
3.3. Starter Selection Based on Hedonic Evaluation
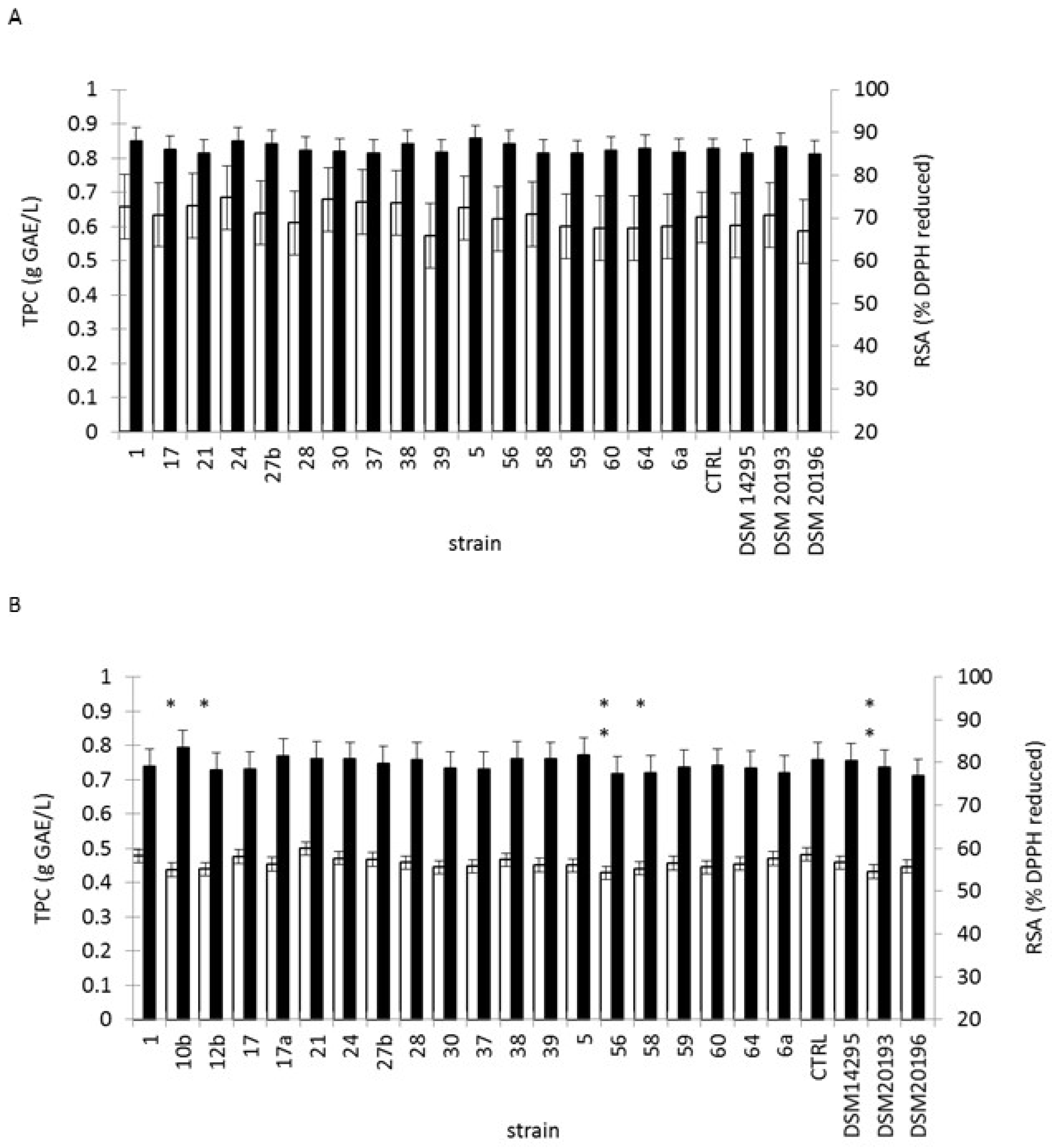
3.4. Lactic Fermentation of Black or Green Tea Infusion
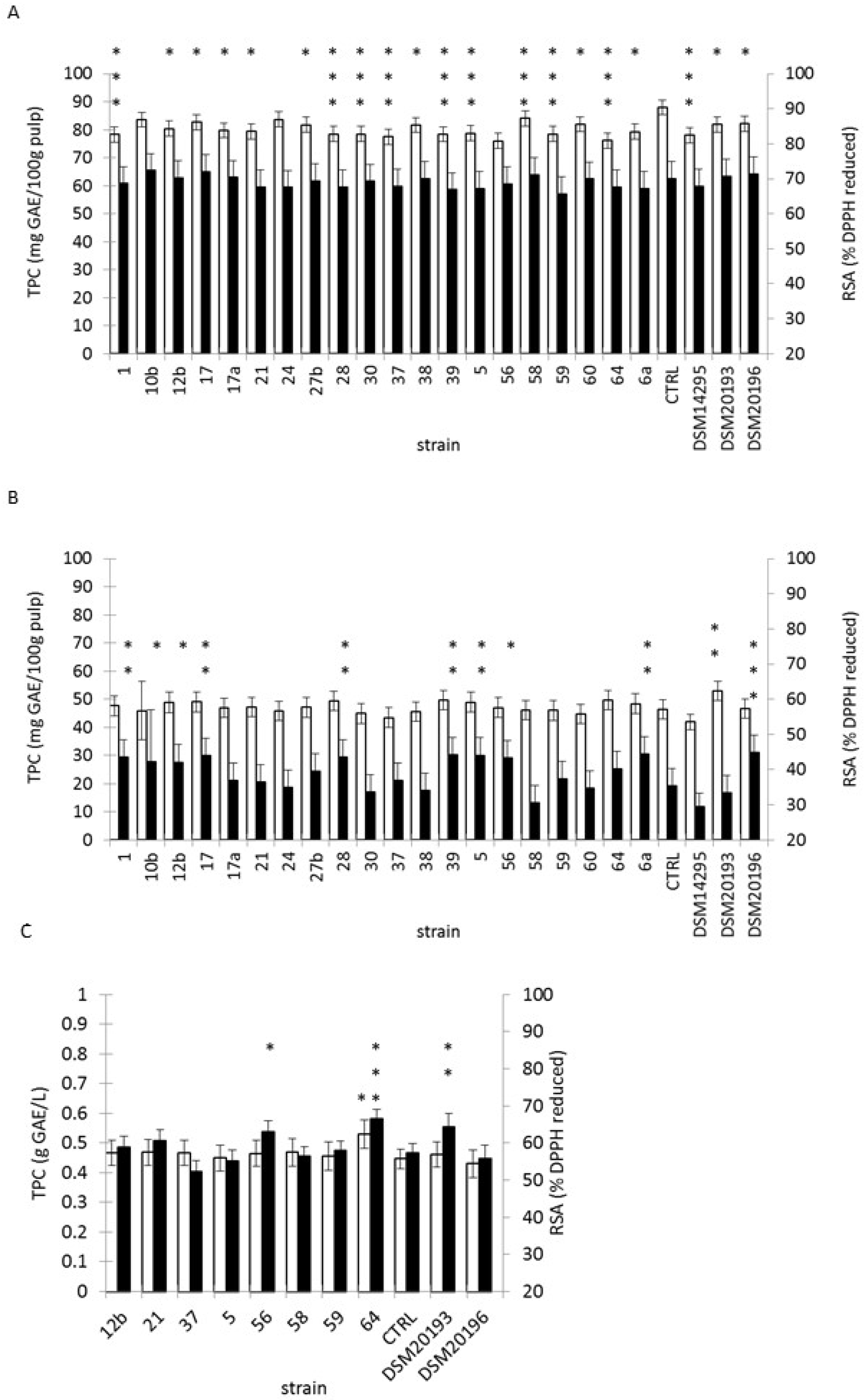
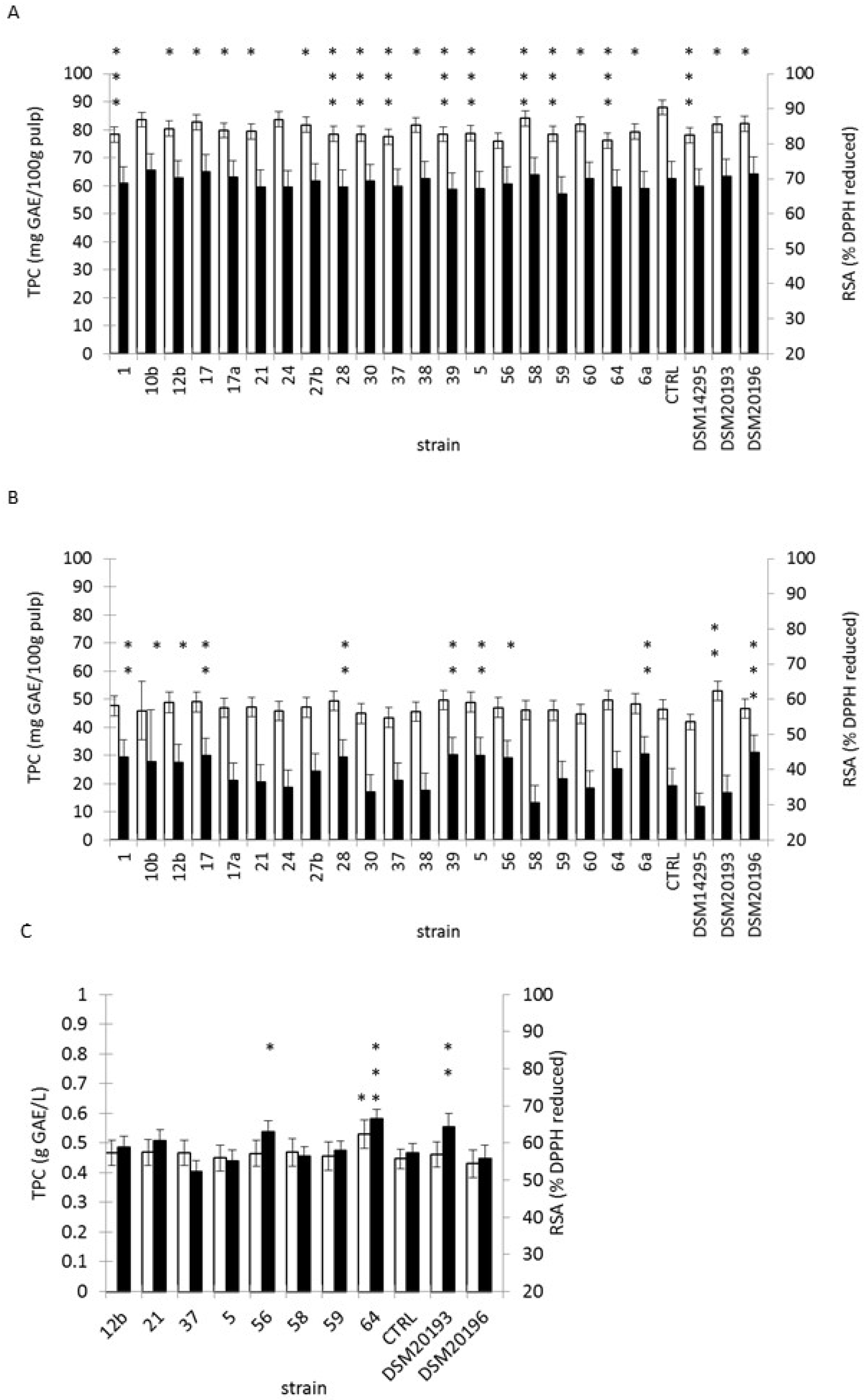
3.5. Fermentation of Tropical Fruit Preparations
3.6. Changes in Microbial Counts, pH, Sugar and RSA Over-Fermentation and Refrigerated Storage
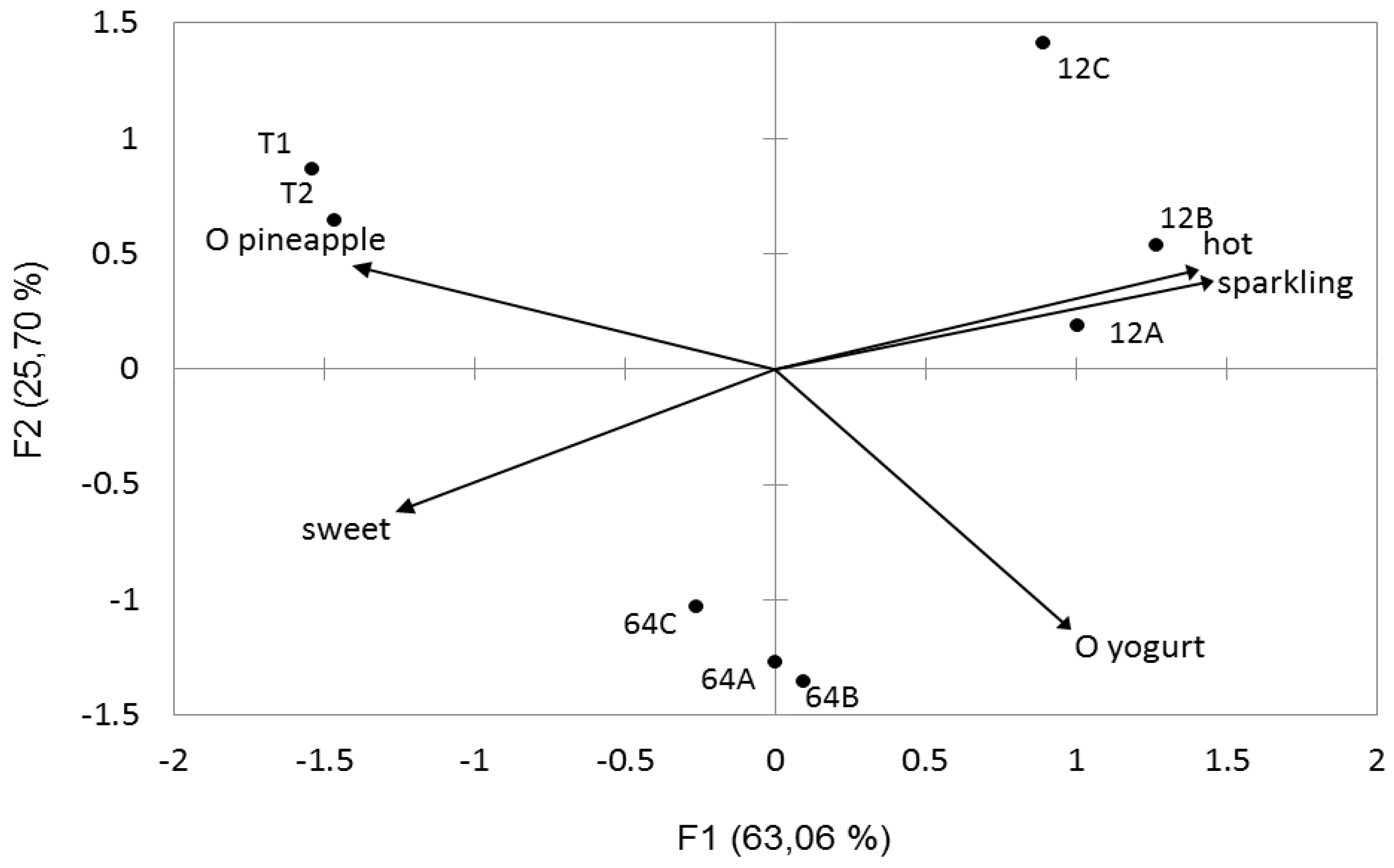
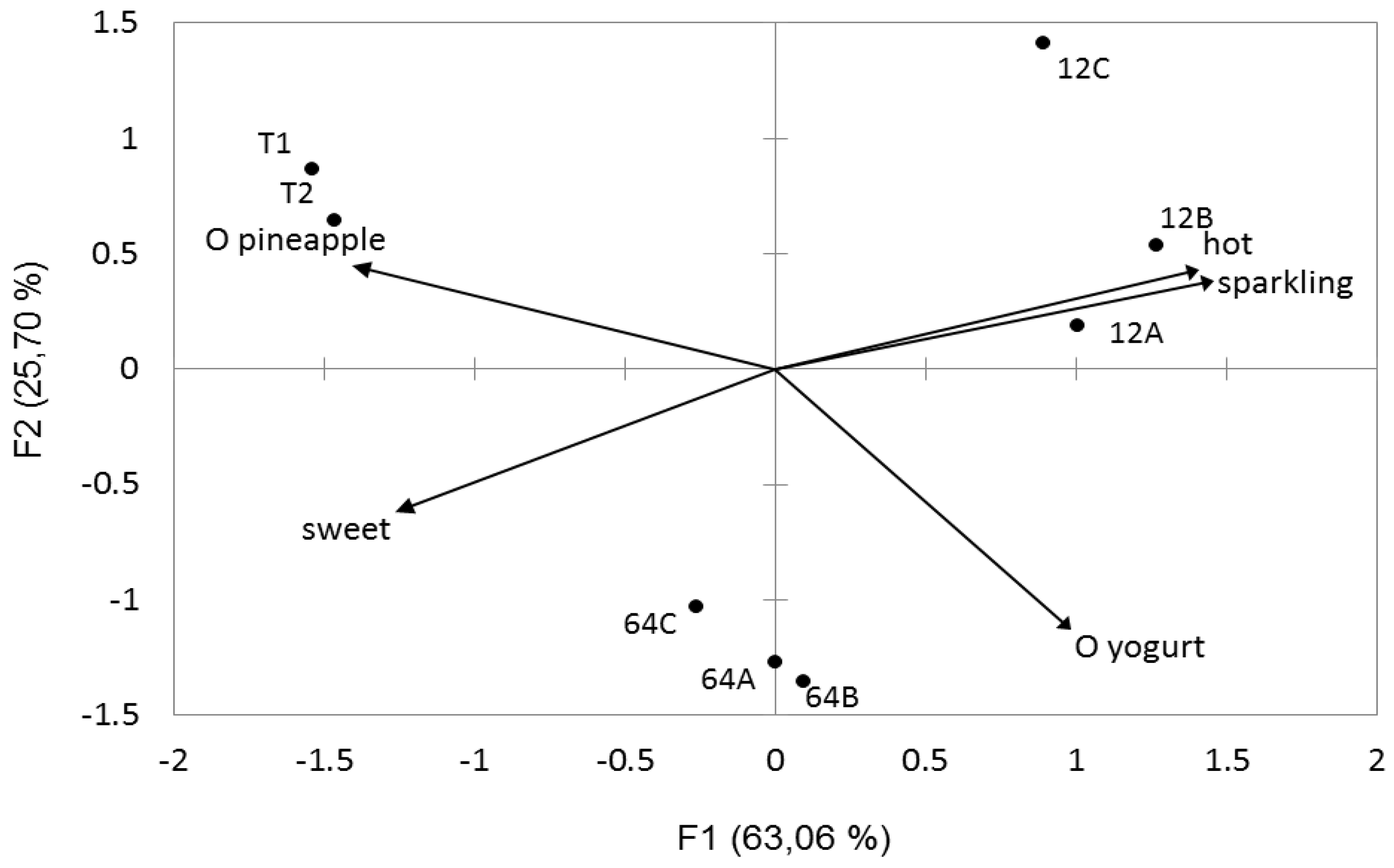
3.7. Sensory Profile and Fermented Juice Quality Evaluation
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Arts, I.C.W.; Hollman, P.C.H. Polyphenols and disease risk in epidemiologic studies 1–4. Am. J. Clin. Nutr. 2005, 81, 3175–3255. [Google Scholar]
- McKay, D.L.; Blumberg, J.B. The role of tea in human health: An update. J. Am. Coll. Nutr. 2002, 21, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Di Cagno, R.; Coda, R.; de Angelis, M.; Gobbetti, M. Exploitation of vegetables and fruits through lactic acid fermentation. Food Microbiol. 2013, 33, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Nicoli, M.; Anese, M.; Parpinel, M. Influence of processing on the antioxidant properties of fruit and vegetables. Trends Food Sci. Technol. 1999, 10, 94–100. [Google Scholar] [CrossRef]
- Ioannou, I.; Hafsa, I.; Hamdi, S.; Charbonnel, C.; Ghoul, M. Review of the effects of food processing and formulation on flavonol and anthocyanin behaviour. J. Food Eng. 2012, 111, 208–217. [Google Scholar] [CrossRef]
- Oboh, G. Effect of blanching on the antioxidant properties of some tropical green leafy vegetables. LWT Food Sci. Technol. 2005, 38, 513–517. [Google Scholar] [CrossRef]
- Beena Divya, J.; Kulangara Varsha, K.; Madhavan Nampoothiri, K.; Ismail, B.; Pandey, A. Probiotic fermented foods for health benefits. Eng. Life Sci. 2012, 12, 377–390. [Google Scholar] [CrossRef]
- Dufresne, C.; Farnworth, E. Tea, Kombucha, and health: a review. Food Res. Int. 2000, 33, 409–421. [Google Scholar] [CrossRef]
- Park, K.Y.; Jeong, J.K.; Lee, Y.E.; Daily, J.W. Health benefits of Kimchi (Korean fermented vegetables) as a probiotic food. J. Med. Food 2014, 17, 6–20. [Google Scholar] [CrossRef] [PubMed]
- Di Cagno, R.; Minervini, G.; Rizzello, C.G.; de Angelis, M.; Gobbetti, M. Effect of lactic acid fermentation on antioxidant, texture, color and sensory properties of red and green smoothies. Food Microbiol. 2011, 28, 1062–1071. [Google Scholar] [CrossRef] [PubMed]
- Filannino, P.; Azzi, L.; Cavoski, I.; Vincentini, O.; Rizzello, C.G.; Gobbetti, M.; di Cagno, R. Exploitation of the health-promoting and sensory properties of organic pomegranate (Punica granatum L.) juice through lactic acid fermentation. Int. J. Food Microbiol. 2013, 163, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Hur, S.J.; Lee, S.Y.; Kim, Y.C.; Choi, I.; Kim, G.B. Effect of fermentation on the antioxidant activity in plant-based foods. Food Chem. 2014, 160, 346–356. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.C.; Wang, C.Y.; Wang, Y.P.; Tzeng, W.S.; Shyu, Y.T. Lactic acid bacterial fermentation on the production of functional antioxidant herbal Anoectochilus formosanus Hayata. J. Biosci. Bioeng. 2011, 111, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Di Cagno, R.; Surico, R.F.; Paradiso, A.; de Angelis, M.; Salmon, J.C.; Buchin, S.; de Gara, L.; Gobbetti, M. Effect of autochthonous lactic acid bacteria starters on health-promoting and sensory properties of tomato juices. Int. J. Food Microbiol. 2009, 128, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Di Cagno, R.; Surico, R.F.; Siragusa, S.; de Angelis, M.; Paradiso, A.; Minervini, F.; de Gara, L.; Gobbetti, M. Selection and use of autochthonous mixed starter for lactic acid fermentation of carrots, French beans or marrows. Int. J. Food Microbiol. 2008, 127, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Dueñas, M.; Fernández, D.; Hernández, T.; Estrella, I.; Muñoz, R. Bioactive phenolic compounds of cowpeas (Vigna sinensis L). Modifications by fermentation with natural microflora and with Lactobacillus plantarum ATCC 14917. J. Sci. Food Agric. 2005, 85, 297–304. [Google Scholar]
- Marazza, J.A.; Nazareno, M.A.; Savoy de Giori, G.; Garro, M.S. Enhancement of the antioxidant capacity of soymilk by fermentation with Lactobacillus rhamnosus. J. Funct. Foods 2012, 4, 594–601. [Google Scholar] [CrossRef]
- Pyo, Y.H.; Lee, T.C.; Lee, Y.C. Enrichment of bioactive isoflavones in soymilk fermented with β-glucosidase-producing lactic acid bacteria. Food Res. Int. 2005, 38, 551–559. [Google Scholar] [CrossRef]
- Zhao, D.; Shah, N.P. Lactic acid bacterial fermentation modified phenolic composition in tea extracts and enhanced their antioxidant activity and cellular uptake of phenolic compounds following in vitro digestion. J. Funct. Foods 2016, 20, 182–194. [Google Scholar] [CrossRef]
- Fessard, A.; Bourdon, E.; Payet, B.; Remize, F. Identification, stress tolerance, and antioxidant activity of lactic acid bacteria isolated from tropically grown fruits and leaves. Can. J. Microbiol. 2016, 62, 550–561. [Google Scholar] [CrossRef] [PubMed]
- Naser, S.M.; Thompson, F.L.; Hoste, B.; Gevers, D.; Dawyndt, P.; Vancanneyt, M.; Swings, J. Application of multilocus sequence analysis (MLSA) for rapid identification of Enterococcus species based on rpoA and pheS genes. Microbiology 2005, 151, 2141–2150. [Google Scholar] [CrossRef] [PubMed]
- Rubio, R.; Jofré, A.; Martín, B.; Aymerich, T.; Garriga, M. Characterization of lactic acid bacteria isolated from infant faeces as potential probiotic starter cultures for fermented sausages. Food Microbiol. 2014, 38, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Torriani, S.; Felis, G.E.; Dellaglio, F. Differentiation of Lactobacillus plantarum, L. pentosus, and L. paraplantarum by recA Gene Sequence Analysis and Multiplex PCR Assay with recA Gene-Derived Primers. Appl. Environ. Microbiol. 2001, 67, 3450–3454. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Septembre-Malaterre, A.; Stanislas, G.; Douraguia, E.; Gonthier, M.P.P. Evaluation of nutritional and antioxidant properties of the tropical fruits banana, litchi, mango, papaya, passion fruit and pineapple cultivated in Reunion French Island. Food Chem. 2016, 212, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Vos, P.; Garrity, G.; Jones, D.; Krieg, N.R.; Ludwig, W.; Rainey, F.A.; Schleifer, K.H.; Whitman, W. Bergey’s Manual of Systematic Bacteriology; Springer: New York, NY, USA, 2009. [Google Scholar]
- Espirito-Santo, A.P.; Carlin, F.; Renard, C.M.G.C. Apple, grape or orange juice: Which one offers the best substrate for lactobacilli growth?—A screening study on bacteria viability, superoxide dismutase activity, folates production and hedonic characteristics. Food Res. Int. 2015, 78, 352–360. [Google Scholar] [CrossRef] [PubMed]
- Almajano, M.P.; Carbó, R.; Jiménez, J.A.L.; Gordon, M.H. Antioxidant and antimicrobial activities of tea infusions. Food Chem. 2007, 108, 55–63. [Google Scholar] [CrossRef]
- Atoui, A. Tea and herbal infusions: Their antioxidant activity and phenolic profile. Food Chem. 2005, 89, 27–36. [Google Scholar] [CrossRef]
- Karori, S.M.; Wachira, F.N.; Wanyoko, J.K.; Ngure, R.M. Antioxidant capacity of different types of tea products. Afr. J. Biotechnol. 2007, 6, 2287–2296. [Google Scholar] [CrossRef]
- Malbaša, R.V.; Lončar, E.S.; Vitas, J.S.; Čanadanović-Brunet, J.M. Influence of starter cultures on the antioxidant activity of kombucha beverage. Food Chem. 2011, 127, 1727–1731. [Google Scholar] [CrossRef]
- Peterson, J.; Dwyer, J.; Bhagwat, S.; Haytowitz, D.; Holden, J.; Eldridge, A.L.; Beecher, G.; Aladesanmi, J. Major flavonoids in dry tea. J. Food Compos. Anal. 2005, 18, 487–501. [Google Scholar] [CrossRef]
- Mukhtar, H.; Ahmad, N. Tea polyphenols: Prevention of cancer and optimizing health. Am. J. Clin. Nutr. 2000, 71, 1698–1702. [Google Scholar]
- Sang, S.; Lambert, J.D.; Ho, C.T.; Yang, C.S. The chemistry and biotransformation of tea constituents. Pharmacol. Res. 2011, 64, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.L.; Chen, S.G.; Zhao, Y.Y.; Luo, C.X.; Li, J.; Gao, Y.Q. Total phenolic contents of 33 fruits and their antioxidant capacities before and after in vitro digestion. Ind. Crop. Prod. 2014, 57, 150–157. [Google Scholar] [CrossRef]
- Palafox-Carlos, H.; Yahia, E.M.; González-Aguilar, G.A. Identification and quantification of major phenolic compounds from mango (Mangifera indica, cv. Ataulfo) fruit by HPLC–DAD–MS/MS-ESI and their individual contribution to the antioxidant activity during ripening. Food Chem. 2012, 135, 105–111. [Google Scholar] [CrossRef]
- Nguyen, T.T.T.; Shaw, P.N.; Parat, M.O.; Hewavitharana, A.K. Anticancer activity of Carica papaya: A review. Mol. Nutr. Food Res. 2013, 57, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Pastrana, D.M.; Yahia, E.M.; González-Aguilar, G.A. Phenolic and carotenoid profiles of papaya fruit (Carica papaya L.) and their contents under low temperature storage. J. Sci. Food Agric. 2010, 90, 2358–2365. [Google Scholar] [PubMed]
- Rodriguez, H.; Curiel, J.A.; Landete, J.M.; de las Rivas, B.; Lopez de Felipe, F.; Gomez-Cordoves, C.; Mancheno, J.M.; Munoz, R. Food phenolics and lactic acid bacteria. Int. J. Food Microbiol. 2009, 132, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, H.; Landete, J.M.; de las Rivas, B.; Muñoz, R. Metabolism of food phenolic acids by Lactobacillus plantarum CECT 748T. Food Chem. 2008, 107, 1393–1398. [Google Scholar] [CrossRef]
- Kostinek, M.; Specht, I.; Edward, V.A.; Pinto, C.; Egounlety, M.; Sossa, C.; Mbugua, S.; Dortu, C.; Thonart, P.; Taljaard, L.; et al. Characterisation and biochemical properties of predominant lactic acid bacteria from fermenting cassava for selection as starter cultures. Int. J. Food Microbiol. 2007, 114, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Ramos-de-la-Peña, A.M.; Contreras-Esquivel, J.C. Methods and substrates for feruloyl esterase activity detection, a review. J. Mol. Catal. B Enzym. 2016, 130, 74–87. [Google Scholar] [CrossRef]
- Niki, E.; Noguchi, N. Evaluation of Antioxidant Capacity. What Capacity is Being Measured by Which Method? IUBMB Life 2001, 50, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Bartolomé, A.P.; Rupérez, P.; Fúster, C. Pineapple fruit: Morphological characteristics, chemical composition and sensory analysis of red Spanish and Smooth Cayenne cultivars. Food Chem. 1995, 53, 75–79. [Google Scholar] [CrossRef]
- Cárnara, M.; Díez, C.; Torija, E. Chemical characterization of pineapple juices and nectars. Principal components analysis. Food Chem. 1995, 54, 93–100. [Google Scholar] [CrossRef]
- Corona, O.; Alfonzo, A.; Ventimiglia, G.; Nasca, A.; Francesca, N.; Martorana, A.; Moschetti, G.; Settanni, L. Industrial application of selected lactic acid bacteria isolated from local semolinas for typical sourdough bread production. Food Microbiol. 2016, 59, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Pogačić, T.; Maillard, M.B.; Leclerc, A.; Hervé, C.; Chuat, V.; Valence, F.; Thierry, A. Lactobacillus and Leuconostoc volatilomes in cheese conditions. Appl. Microbiol. Biotechnol. 2016, 100, 2335–2346. [Google Scholar] [CrossRef] [PubMed]
- Altay, F.; Karbancıoglu-Güler, F.; Daskaya-Dikmen, C. A review on traditional Turkish fermented non-alcoholic beverages: Microbiota, fermentation process and quality characteristics. Int. J. Food Microbiol. 2013, 167, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.Y.; Lee, S.H.; Lee, H.J.; Seo, H.Y.; Park, W.S.; Jeon, C.O. Effects of Leuconostoc mesenteroides starter cultures on microbial communities and metabolites during kimchi fermentation. Int. J. Food Microbiol. 2012, 153, 378–387. [Google Scholar] [CrossRef] [PubMed]
- Park, E.J.; Chun, J.; Cha, C.J.; Park, W.S.; Jeon, C.O.; Bae, J.W. Bacterial community analysis during fermentation of ten representative kinds of kimchi with barcoded pyrosequencing. Food Microbiol. 2012, 30, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Abriouel, H.; Lerma, L.L.; Casado Muñoz, M.d.C.; Montoro, B.P.; Kabisch, J.; Pichner, R.; Cho, G.S.; Neve, H.; Fusco, V.; Franz, C.M.A.P.; et al. The controversial nature of the Weissella genus: Technological and functional aspects versus whole genome analysis-based pathogenic potential for their application in food and health. Front. Microbiol. 2015, 6, 1197. [Google Scholar] [CrossRef] [PubMed]
- Jeong, D.W.; Lee, J.H. Antibiotic resistance, hemolysis and biogenic amine production assessments of Leuconostoc and Weissella isolates for kimchi starter development. LWT Food Sci. Technol. 2015, 64, 1078–1084. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Species | Origin | Production EPS | Aspect |
|---|---|---|---|---|
| 1 | Lc. mesenteroides | Papaya | + | Liquid |
| 2 | Lc. citreum | Sliced cabbage | + | Creamy |
| 5 | Lc. mesenteroides | Sliced cabbage | + | Liquid |
| 17 | W. confusa | Sliced cabbage | ND * | ND * |
| 21 | W. cibaria | Sliced cabbage | + | Creamy |
| 24 | Lc. lactis | Sliced cabbage | + | Creamy |
| 28 | Lc. mesenteroides | Sliced cabbage | + | Creamy |
| 30 | W. cibaria | Sliced cabbage | + | Liquid |
| 37 | W. paramesenteroides | Papaya | - | - |
| 38 | W. confusa | Papaya | + | Creamy |
| 39 | Lc. pseudomesenteroides | Papaya | + | Liquid |
| 56 | Lc. pseudomesenteroides | Sliced cabbage | + | Creamy |
| 58 | W. soli | Sliced cabbage | − | - |
| 59 | W. confusa | Sliced cabbage | + | Liquid |
| 60 | Lc. pseudomesenteroides | Sliced cabbage | + | Creamy |
| 64 | W. cibaria | Sliced cabbage | + | Creamy |
| 73 | Lb. paraplantarum | Sliced cabbage | − | - |
| 75 | Lb. plantarum | Sliced cabbage | − | - |
| 77 | Fb. tropaeoli | Papaya | − | - |
| 78 | Lc. pseudomesenteroides | Papaya | + | Liquid |
| 79 | Lc. pseudomesenteroides | Papaya | + | Liquid |
| 89 | Lc. pseudomesenteroides | Sliced cabbage | + | Creamy |
| 10b | W. cibaria | Tomato | + | Creamy |
| 12b | Lc. pseudomesenteroides | Tomato | + | Liquid |
| 17a | Lb. plantarum | Tomato | − | - |
| 27b | Lc. pseudomesenteroides | Papaya | + | Liquid |
| 6a | Lc. mesenteroides | Papaya | + | Creamy |
| 9a | Lc. citreum | Tomato | + | Creamy |
| DSM14295 | W. cibaria | Kimchi | + | Creamy |
| DSM20188 | Lc. citreum | ND * | + | Creamy |
| DSM20193 | Lc. pseudomesenteroides | Sugar cane juice | + | Liquid |
| DSM20196 | W. confusa | Cane sugar | + | Creamy |
| DSM2601 | Lb. plantarum | Pickled cabbage | − | - |
| DSM5625 | Lc. pseudomesenteroides | Commercial starter | + | Creamy |
| Strain | Species | Latency (h) | pH min | Vm (mUpH·h−1) | Tm (h) | pHm |
|---|---|---|---|---|---|---|
| 1 | Lc. mesenteroides | 25.3 ± 7.3 | 3.9 ± 0.4 | 205 ± 91 | 4.5 ± 0.2 | 5 ± 0.1 |
| 2 | Lc. citreum | 12.0 ± 0.8 | 4.3 ± 0.2 | 154 ± 2 | 4.1 ± 0.5 | 5.1 ± 0.1 |
| 5 | Lc. mesenteroides | 12.3 ± 5.1 | 3.7 ± 0.2 | 218 ± 47 | 5.4 ± 1.2 | 4.9 ± 0.2 |
| 17 | W. confusa | 2.5 ± 0.0 | 4.1 ± 0 | 211 ± 33 | 3.6 ± 0.3 | 5.1 ± 0.2 |
| 21 | W. cibaria | 9.0 ± 3.6 | 4 ± 0.1 | 180 ± 38 | 4.1 ± 0.9 | 5 ± 0.1 |
| 24 | Lc. lactis | 6.6 ± 3.3 | 4.1 ± 0.1 | 170 ± 36 | 3.3 ± 0.7 | 5.3 ± 0.2 |
| 28 | Lc. mesenteroides | 17.3 ± 5.3 | 3.9 ± 0.2 | 212 ± 68 | 4.7 ± 0.5 | 5 ± 0.2 |
| 30 | W. cibaria | 8.4 ± 0.2 | 4.1 ± 0.1 | 180 ± 72 | 4.3 ± 0.1 | 5 ± 0.2 |
| 37 | W. paramesenteroides | 12.0 ± 3.7 | 3.9 ± 0.1 | 84 ± 9 | 4.8 ± 0.3 | 5 ± 0 |
| 38 | W. confusa | 16.2 ± 2.0 | 4.1 ± 0.1 | 144 ± 25 | 4.3 ± 0.9 | 5 ± 0.1 |
| 39 | Lc. pseudomesenteroides | 7.5 ± 3.1 | 4 ± 0.2 | 196 ± 63 | 4.9 ± 1.1 | 5 ± 0.1 |
| 56 | Lc. pseudomesenteroides | 12.2 ± 4.6 | 4 ± 0.2 | 258 ± 44 | 5 ± 0 | 5 ± 0 |
| 58 | W. soli | 18.2 ± 4.1 | 4.3 ± 0.2 | 128 ± 20 | 8.6 ± 0.4 | 5 ± 0.5 |
| 59 | W. confusa | 16.1 ± 2.6 | 4.3 ± 0.1 | 126 ± 51 | 4.8 ± 0.5 | 5.2 ± 0.1 |
| 60 | Lc. pseudomesenteroides | 14.1 ± 0.6 | 4 ± 0.1 | 257 ± 55 | 3.2 ± 0.6 | 5 ± 0 |
| 64 | W. cibaria | 10.8 ± 2.0 | 4 ± 0.2 | 192 ± 11 | 3.3 ± 0.2 | 4.9 ± 0.2 |
| 73 | Lb. paraplantarum | 11.2 ± 1.4 | 3.8 ± 0.1 | 234 ± 5 | 5 ± 0.4 | 5 ± 0 |
| 75 | Lb. plantarum | 8.8 ± 3.4 | 3.7 ± 0.1 | 225 ± 83 | 4.2 ± 0.5 | 5.1 ± 0.1 |
| 77 | Fb. tropaeoli | 13.3 ± 6.9 | 3.7 ± 0.1 | 263 ± 28 | 4.9 ± 0.3 | 5 ± 0 |
| 78 | Lc. pseudomesenteroides | 14.7 ± 0.6 | 4 ± 0.2 | 206 ± 48 | 4.3 ± 0.4 | 4.9 ± 0.1 |
| 79 | Lc. pseudomesenteroides | 17.8 ± 7.1 | 4.5 ± 0.4 | 133 ± 12 | 4.6 ± 0.6 | 5.1 ± 0.2 |
| 89 | Lc. pseudomesenteroides | 26.1 ± 7.1 | 3.8 ± 0.3 | 165 ± 23 | 6.1 ± 1.5 | 4.8 ± 0.1 |
| 10b | W. cibaria | 9 ± ND * | 4.1 ± ND | 100 ± ND | 6.3 ± ND | 5 ± ND |
| 12b | Lc. pseudomesenteroides | 8.6 ± 2.7 | 4 ± 0.3 | 138 ± 19 | 8.8 ± 2.8 | 5 ± 0.2 |
| 17a | Lb. plantarum | 7.3 ± 4.7 | 3.6 ± 0.2 | 237 ± 35 | 4.6 ± 1.1 | 5 ± 0.3 |
| 27b | Lc. pseudomesenteroides | 10 ± 3.9 | 3.9 ± 0.2 | 149 ± 13 | 8.1 ± 1.5 | 4.9 ± 0.3 |
| 6a | Lc. mesenteroides | 10.6 ± 5.2 | 4 ± 0 | 184 ± 15 | 4.1 ± 0.8 | 5.1 ± 0.2 |
| 9a | Lc. citreum | 16.5 ± 0.7 | 3.8 ± 0.1 | 243 ± 40 | 4.1 ± 0.2 | 5 ± 0 |
| DSM14295 | W. cibaria | 10.5 ± 1.4 | 4 ± 0.1 | 200 ± 6 | 3.7 ± 0.6 | 5.1 ± 0.1 |
| DSM20188 | Lc. citreum | 10 ± 1.4 | 4 ± 0.1 | 184 ± 45 | 8.1 ± 1.6 | 5.2 ± 0.1 |
| DSM20193 | Lc. pseudomesenteroides | 13.1 ± 3 | 4.1 ± 0.6 | 221 ± 73 | 4.1 ± 0.5 | 5.1 ± 0.1 |
| DSM20196 | W. confusa | 14.7 ± 3 | 4 ± 0.1 | 377 ± 31 | 2.8 ± 0.5 | 5 ± 0.1 |
| DSM2601 | Lb. plantarum | 7.2 ± 1.6 | 3.6 ± 0 | 193 ± 52 | 5.6 ± 0.6 | 4.8 ± 0 |
| DSM5625 | Lc. pseudomesenteroides | 10.5 ± 1.9 | 3.9 ± 0.1 | 159 ± 36 | 5.9 ± 0 | 4.9 ± 0.1 |
| Sensory Properties of Pineapple Juice After Fermentation | |||
|---|---|---|---|
| Strain | Species | 2 Days | 4 Days |
| 5 | Lc. mesenteroides | + | + |
| 17 | W. confusa | ++ | ++ |
| 21 | W. cibaria | + | + |
| 37 | W. paramesenteroides | ++ | ++ |
| 56 | Lc. pseudomesenteroides | + | + |
| 58 | W. soli | + | ++ |
| 59 | W. confusa | ++ | ++ |
| 12b | Lc. pseudomesenteroides | + | + |
| 1 | Lc. mesenteroides | + | - |
| 24 | Lc. lactis | ++ | - |
| 28 | Lc. mesenteroides | + | - |
| 30 | W. cibaria | − | ++ |
| 38 | W. confusa | − | ++ |
| 39 | Lc. pseudomesenteroides | − | + |
| 60 | Lc. pseudomesenteroides | − | + |
| 10b | W. cibaria | + | - |
| 17a | Lb. plantarum | − | + |
| 27b | Lc. pseudomesenteroides | − | ++ |
| 6a | Lc. mesenteroides | ++ | - |
| 64 | W. cibaria | + | + |
| DSM14295 | W. cibaria | − | + |
| DSM20193 | Lc. pseudomesenteroides | − | ++ |
| DSM20196 | W. confusa | + | + |
| DSM2601 | Lb. plantarum | − | - |
| 2 | Lc. citreum | − | - |
| 73 | Lb. paraplantarum | − | - |
| 75 | Lb. plantarum | − | - |
| 77 | Fb. tropaeoli | − | - |
| 78 | Lc. pseudomesenteroides | − | - |
| 79 | Lc. pseudomesenteroides | − | - |
| 89 | Lc. pseudomesenteroides | − | - |
| 9a | Lc. citreum | − | - |
| DSM20188 | Lc. citreum | − | - |
| Condition | Half-Time for Haemolysis (Min) | Half-Time for Ldl Oxidation (Min) | ||
|---|---|---|---|---|
| CONTROL CELLS | 269 ± 20 | 28.1 ± 4.2 | B | |
| PAPAYA PULP | 302 ± 24 | 45.1 ± 5.1 | AB | |
| PAPAYA/STRAIN 1 | 340 ± 27 | * vs control cells | ||
| PAPAYA/STRAIN 10B | 257 ± 94 | * vs pulp | ||
| PAPAYA/STRAIN 12B | 264 ± 70 | 52.3 ± 23.0 | AB | |
| PAPAYA/STRAIN 28 | 280 ± 40 | |||
| PAPAYA/STRAIN DSM20193 | 319 ± 20 | |||
| PAPAYA/STRAIN 56 | 259 ± 46 | 52.0 ± 15.0 | AB | |
| PAPAYA/STRAIN 64 | 279 ± 12 | 54.2 ± 28.2 | AB | |
| PAPAYA/STRAIN 17 | 278 ± 7 | |||
| PINEAPPLE JUICE | 282 ± 24 | 71.7 ± 9.4 | A | |
| PINEAPPLE/STRAIN 12B | 208 ± 72 | * vs juice | 75.9 ± 12.7 | A |
| PINEAPPLE/STRAIN DSM20193 | 273 ± 73 | |||
| PINEAPPLE/STRAIN 56 | 234 ± 79 | 75.7 ± 12.4 | A | |
| PINEAPPLE/STRAIN 64 | 289 ± 47 | 54.2 ± 28.2 | A | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fessard, A.; Kapoor, A.; Patche, J.; Assemat, S.; Hoarau, M.; Bourdon, E.; Bahorun, T.; Remize, F. Lactic Fermentation as an Efficient Tool to Enhance the Antioxidant Activity of Tropical Fruit Juices and Teas. Microorganisms 2017, 5, 23. https://doi.org/10.3390/microorganisms5020023
Fessard A, Kapoor A, Patche J, Assemat S, Hoarau M, Bourdon E, Bahorun T, Remize F. Lactic Fermentation as an Efficient Tool to Enhance the Antioxidant Activity of Tropical Fruit Juices and Teas. Microorganisms. 2017; 5(2):23. https://doi.org/10.3390/microorganisms5020023
Chicago/Turabian StyleFessard, Amandine, Ashish Kapoor, Jessica Patche, Sophie Assemat, Mathilde Hoarau, Emmanuel Bourdon, Theeshan Bahorun, and Fabienne Remize. 2017. "Lactic Fermentation as an Efficient Tool to Enhance the Antioxidant Activity of Tropical Fruit Juices and Teas" Microorganisms 5, no. 2: 23. https://doi.org/10.3390/microorganisms5020023