Recombinant Inga Laurina Trypsin Inhibitor (ILTI) Production in Komagataella Phaffii Confirms Its Potential Anti-Biofilm Effect and Reveals an Anti-Tumoral Activity

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain Constructions and Culture Conditions
2.2. Heterologous Expression of Inga laurina Trypsin Inhibitor (ILTI) in K. Phaffii
2.3. High Cell-Density Fermentation of GSrILTI
2.4. Determination of Dry Cell Weight (DCW) and Substrate Concentrations
2.5. Matrix-Assisted Laser Desorption Ionization–Time-of-Flight (MALDI-TOF) Analysis
2.6. Trypsin Inhibitory Activity in Fermented Broth and Protein Quantification
2.7. Bacterial Strain Used to Biofilm Assay
2.8. Biofilm Formation Assay and Determination of Minimal Biofilm Inhibitory Concentration (MBIC)
2.9. Biofilm Detachment Assay and Determination of Minimal Biofilm Eradication Concentration (MBEC)
2.10. Ethics Statement
2.11. Anti-Tumor Activity Evaluation
2.12. Statistical Analysis
3. Results and Discussion
3.1. Expression of Recombinant ILTI (rILTI)
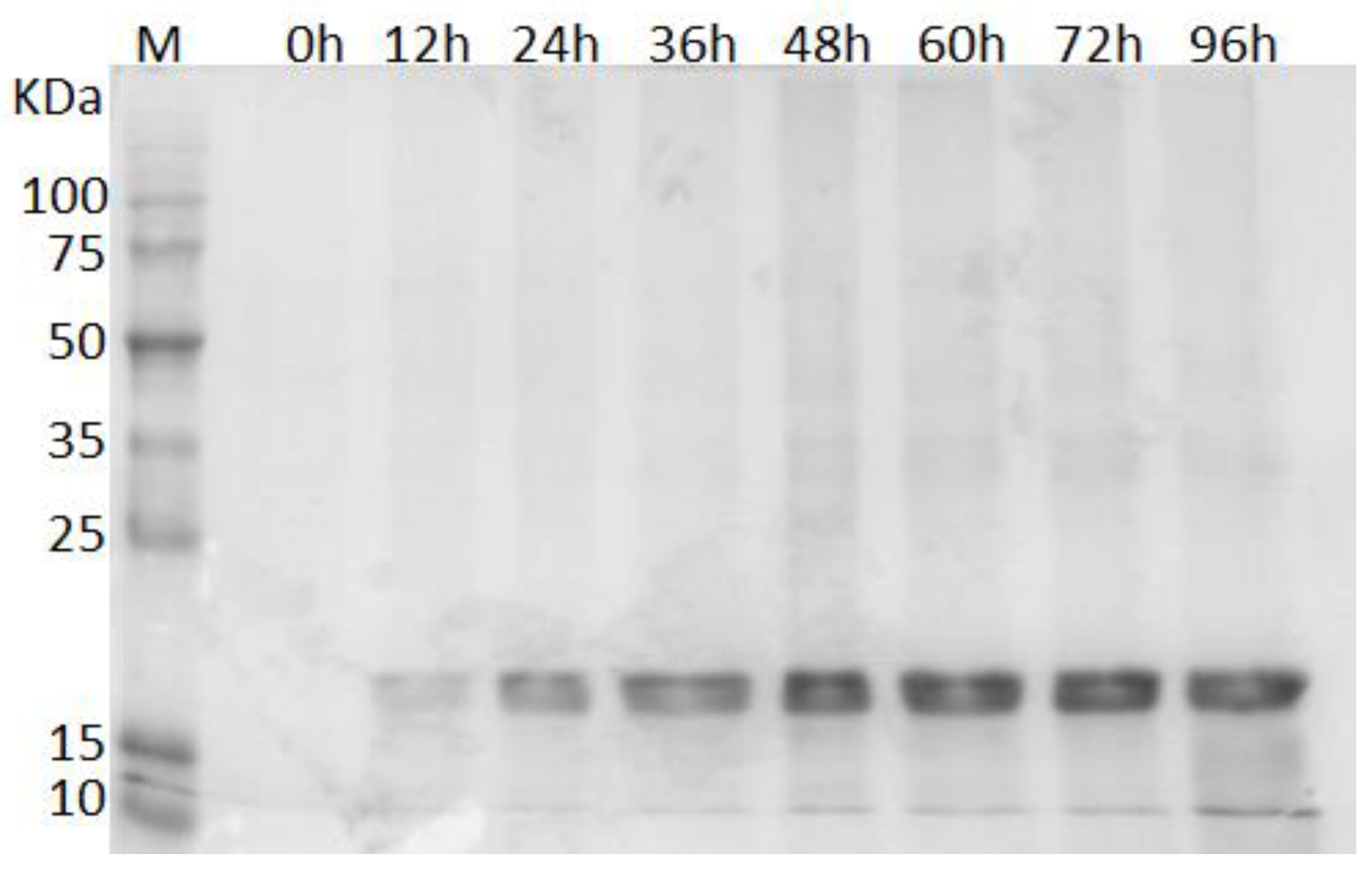
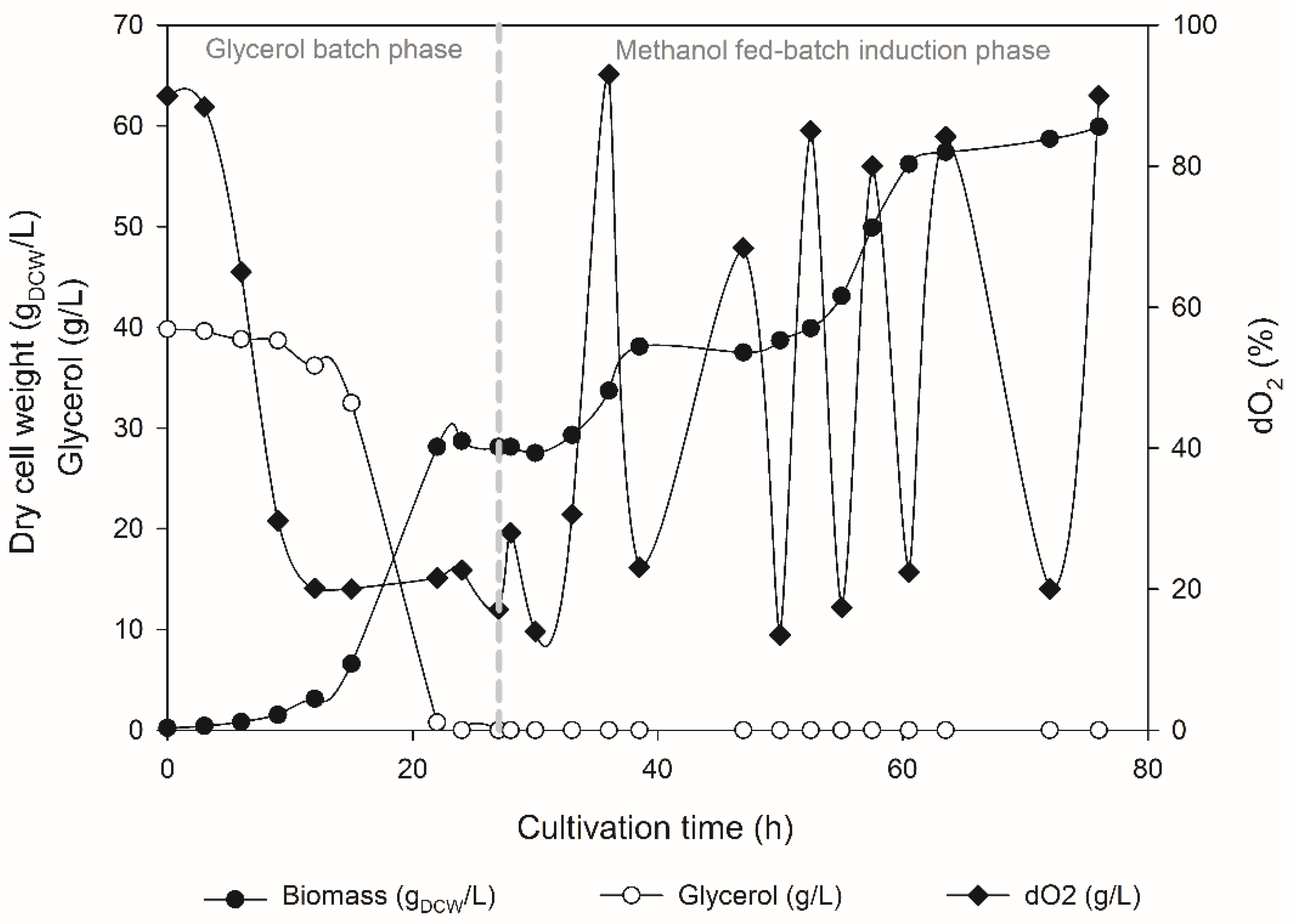
3.2. Production of rILTI in the Bioreactor
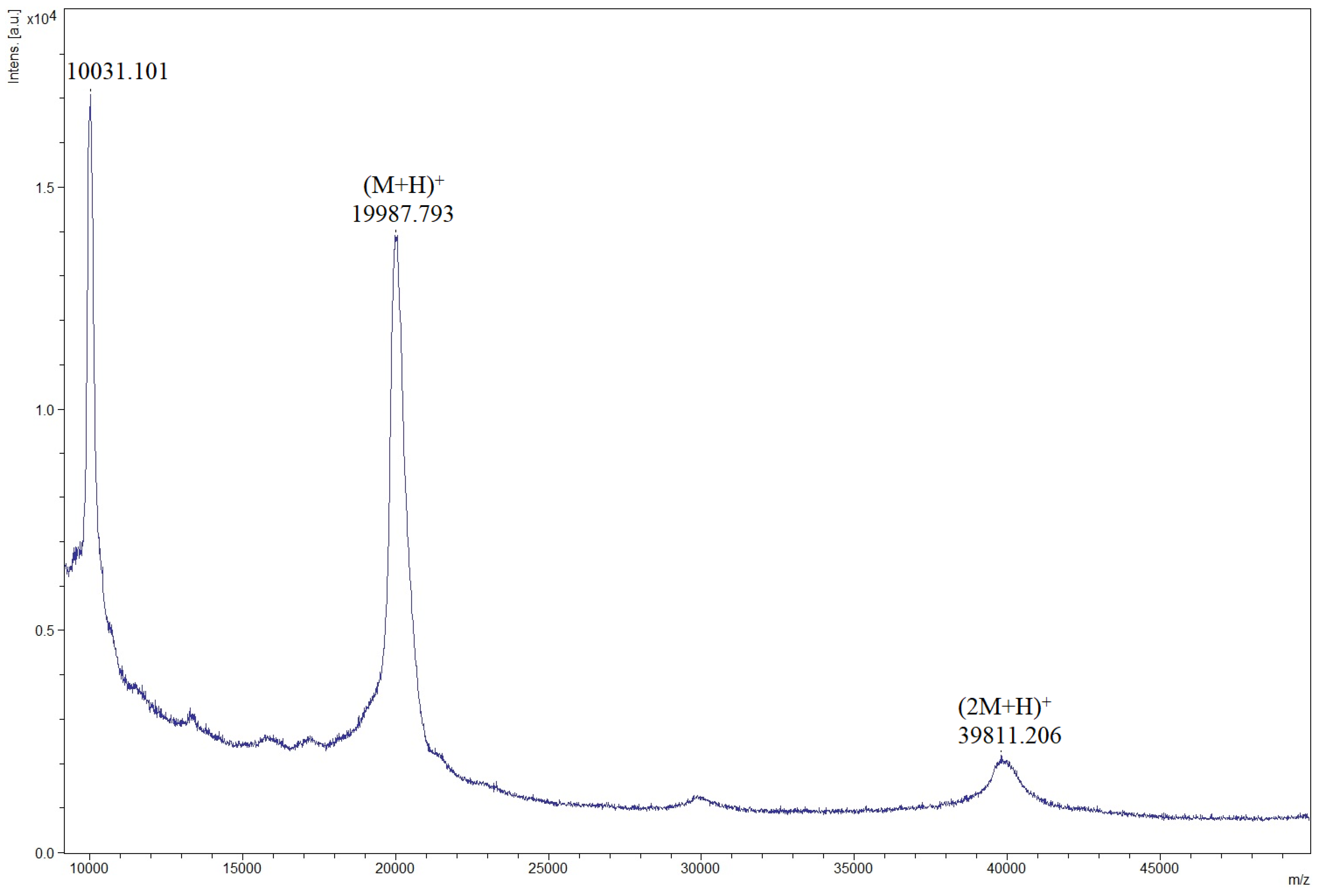
3.3. Confirmation of rILTI Production by MALDI–TOF Analysis
3.4. Determination of rILTI Activity
3.5. Anti-Biofilm Activity of rILTI
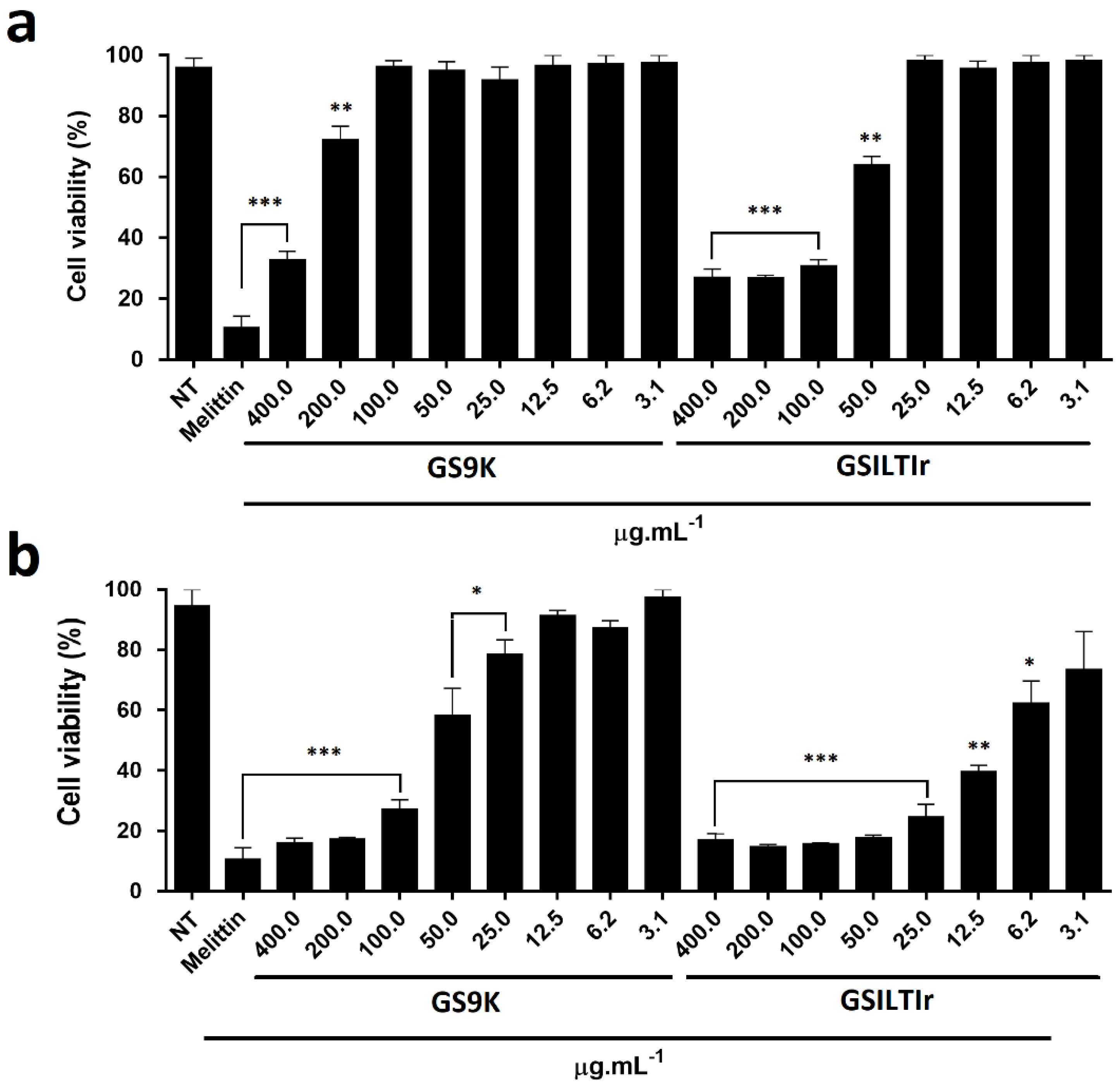
3.6. Evaluation of Anti-Tumoral Activity
4. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Shamsi, T.N.; Parveen, R.; Fatima, S. Characterization, biomedical and agricultural applications of protease inhibitors: A review. Int. J. Biol. Macromol. 2016, 91, 1120–1133. [Google Scholar] [CrossRef] [PubMed]
- Deeks, S.G.; Smith, M.; Holodniy, M.; Kahn, J.O. HIV-1 Protease Inhibitors. JAMA J. Am. Med. Assoc. 1997, 277, 145–153. [Google Scholar] [CrossRef]
- Kim, J.Y.; Park, S.C.; Kim, M.H.; Lim, H.T.; Park, Y.; Hahm, K.S. Antimicrobial activity studies on a trypsin-chymotrypsin protease inhibitor obtained from potato. Biochem. Biophys. Res. Commun. 2005, 330, 921–927. [Google Scholar] [CrossRef] [PubMed]
- Zhu-Salzman, K.; Bi, J.-L.; Liu, T.-X. Molecular strategies of plant defense and insect counter-defense. Insect Sci. 2005, 12, 3–15. [Google Scholar] [CrossRef]
- Dewan, S.S. Global Markets for Enzyme Inhibitors; BIO057C; BCC Research LLC: Wellesley, MA, USA, Jan 2018. [Google Scholar]
- Transparency Market Research (TMR). Biopesticides Market—Major Industry Insights and Forecast 2017–2025; TMR: Pune, India, Sep 2017. [Google Scholar]
- Bijina, B.; Chellappan, S.; Basheer, S.M.; Elyas, K.K.; Bahkali, A.H.; Chandrasekaran, M. Protease inhibitor from Moringa oleifera leaves: Isolation, purification, and characterization. Process Biochem. 2011, 46, 2291–2300. [Google Scholar] [CrossRef]
- Macedo, M.L.R.; Freire, M.D.G.M.; Franco, O.L.; Migliolo, L.; de Oliveira, C.F.R. Practical and theoretical characterization of Inga laurina Kunitz inhibitor on the control of Homalinotus coriaceus. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2011, 158, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Richardson, M. The proteinase inhibitors of plants and micro-organisms. Phytochemistry 1977, 16, 159–169. [Google Scholar] [CrossRef]
- Mello, M.O.; Tanaka, A.S.; Silva-Filho, M.C. Molecular evolution of Bowman-Birk type proteinase inhibitors in flowering plants. Mol. Phylogenet. Evol. 2003, 27, 103–112. [Google Scholar] [CrossRef]
- Ryan, C.A. Protease Inhibitors in Plants: Genes for Improving Defenses Against Insects and Pathogens. Annu. Rev. Phytopathol. 1990, 28, 425–449. [Google Scholar] [CrossRef]
- Ranasinghe, S.; McManus, D.P. Structure and function of invertebrate Kunitz serine protease inhibitors. Dev. Comp. Immunol. 2013, 39, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Garcia, F.C.P.; Fernandes, J.M. Lista de Espécies da Flora do Brasil. Available online: http://floradobrasil.jbrj.gov.br/jabot/floradobrasil/FB23039 (accessed on 20 March 2018).
- Macedo, M.L.R.; Garcia, V.A.; Freire, M.; Richardson, M. Characterization of a Kunitz trypsin inhibitor with a single disulfide bridge from seeds of Inga laurina (SW.) Willd. Phytochemistry 2007, 68, 1104–1111. [Google Scholar] [CrossRef] [PubMed]
- Lomate, P.R.; Bonning, B.C. Proteases and nucleases involved in the biphasic digestion process of the brown marmorated stink bug, Halyomorpha halys (Hemiptera: Pentatomidae). Arch. Insect Biochem. Physiol. 2018, e21459. [Google Scholar] [CrossRef] [PubMed]
- Parde, V.D.; Sharma, H.C.; Kachole, M.S. In vivo inhibition of Helicoverpa armigera gut pro-proteinase activation by non-host plant protease inhibitors. J. Insect Physiol. 2010, 56, 1315–1324. [Google Scholar] [CrossRef] [PubMed]
- Ramos, V.D.S.; Cabrera, O.G.; Camargo, E.L.O.; Ambrósio, A.B.; Vidal, R.O.; da Silva, D.S.; Guimarães, L.C.; Marangoni, S.; Parra, J.R.P.; Pereira, G.A.G.; et al. Molecular cloning and insecticidal effect of Inga laurina trypsin inhibitor on Diatraea saccharalis and Heliothis virescens. Comp. Biochem. Physiol. C. Toxicol. Pharmacol. 2012, 156, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Kurtzman, C.P. Description of Komagataella phaffii sp. nov. and the transfer of Pichia pseudopastoris to the methylotrophic yeast genus Komagataella. Int. J. Syst. Evol. Microbiol. 2005, 973–976. [Google Scholar] [CrossRef] [PubMed]
- Kurtzman, C.P. Biotechnological strains of Komagataella (Pichia) pastoris are Komagataella phaffii as determined from multigene sequence analysis. J. Ind. Microbiol. Biotechnol. 2009, 36, 1435–1438. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Matsuda, M.; Maeda, K.; Mikata, K. The Phylogenetic Relationships of Methanol-assimilating Yeasts Based on the Partial Sequences of 18S and 26S Ribosomal RNAs: The Proposal of Komagataella Gen. Nov. (Saccharomycetaceae). Biosci. Biotechnol. Biochem. 1995, 59, 439–444. [Google Scholar] [CrossRef] [PubMed]
- Mattanovich, D.; Branduardi, P.; Dato, L.; Gasser, B.; Sauer, M.; Porro, D. Recombinant protein production in yeasts. In Methods in Molecular Biology; Springer: Clifton, NJ, USA, 2012; Volume 824, pp. 329–358. ISBN 978-1-61779-432-2. [Google Scholar]
- Cregg, J.M.; Tolstorukov, I.; Kusari, A.; Sunga, J.; Madden, K.; Chappell, T. Expression in the yeast Pichia pastoris. Methods Enzym. 2009, 463, 169–189. [Google Scholar] [CrossRef]
- Kobayashi, K.; Kuwae, S.; Ohya, T.; Ohda, T.; Ohyama, M.; Tomomitsu, K. High level secretion of recombinant human serum albumin by fed-batch fermentation of the methylotrophic yeast, Pichia pastoris, based on optimal methanol feeding strategy. J. Biosci. Bioeng. 2000, 90, 280–288. [Google Scholar] [CrossRef]
- Zepeda, A.B.; Figueroa, C.A.; Abdalla, D.S.P.; Maranhão, A.Q.; Ulloa, P.H.; Pessoa, A.; Farías, J.G. HSF-1, HIF-1 and HSP90 expression on recombinant Pichia pastoris under fed-batch fermentation. Braz. J. Microbiol. 2014, 45, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.-S.; Fan, W.-M.; Zhang, Y.-X. [Effects of different methanol feeding strategy on hirudin production in high-density fermentation by recombinant Pichia pastoris]. Sheng Wu Gong Cheng Xue Bao 2002, 18, 348–351. [Google Scholar] [PubMed]
- Arnau, C.; Ramon, R.; Casas, C.; Valero, F. Optimization of the heterologous production of a Rhizopus oryzae lipase in Pichia pastoris system using mixed substrates on controlled fed-batch bioprocess. Enzyme Microb. Technol. 2010, 46, 494–500. [Google Scholar] [CrossRef] [PubMed]
- Cabrera-Muñoz, A.; Rojas, L.; Gil, D.F.; González-González, Y.; Mansur, M.; Camejo, A.; Pires, J.R.; Alonso-del-Rivero Antigua, M. Heterologous expression of Cenchritis muricatus protease inhibitor II (CmPI-II) in Pichia pastoris system: Purification, isotopic labeling and preliminary characterization. Protein Expr. Purif. 2016, 126, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Gil, D.F.; García-Fernández, R.; Alonso-del-Rivero, M.; Lamazares, E.; Pérez, M.; Varas, L.; Díaz, J.; Chávez, M.A.; González-González, Y.; Mansur, M. Recombinant expression of ShPI-1A, a non-specific BPTI-Kunitz-type inhibitor, and its protection effect on proteolytic degradation of recombinant human miniproinsulin expressed in Pichia pastoris. FEMS Yeast Res. 2011, 11, 575–586. [Google Scholar] [CrossRef] [PubMed]
- Cregg, J.M.; Cereghino, J.L.; Shi, J.; Higgins, D.R. Recombinant protein expression in Pichia pastoris. Mol. Biotechnol. 2000, 16, 23–52. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D.W. Preparation and Transformation of Competent E. coli Using Calcium Chloride. CSH Protoc. 2006, 2006. [Google Scholar] [CrossRef]
- Maurer, M.; Kühleitner, M.; Gasser, B.; Mattanovich, D. Versatile modeling and optimization of fed batch processes for the production of secreted heterologous proteins with Pichia pastoris. Microb. Cell Fact. 2006, 5, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunelle, J.L.; Green, R. One-dimensional SDS-polyacrylamide gel electrophoresis (1D SDS-PAGE). Methods Enzymol. 2014, 541, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Erlanger, B.F.; Kokowsky, N.; Cohen, W. The preparation and properties of two new chromogenic substrates of trypsin. Arch. Biochem. Biophys. 1961, 95, 271–278. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Stepanovic, S.; Vukovic, D.; Hola, V.; Di Bonaventura, G.; Djukic, S.; Cirkovic, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. APMIS 2007, 115, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Ceri, H.; Olson, M.E.; Stremick, C.; Read, R.R.; Morck, D.; Buret, A. The Calgary Biofilm Device: New technology for rapid determination of antibiotic susceptibilities of bacterial biofilms. J. Clin. Microbiol. 1999, 37, 1771–1776. [Google Scholar] [PubMed]
- Patra, S.; Muthuraman, M.S. Gracilaria edulis extract induces apoptosis and inhibits tumor in Ehrlich Ascites tumor cells in vivo. BMC Complement. Altern. Med. 2013, 13, 331. [Google Scholar] [CrossRef] [PubMed]
- Justo, G.Z.; Durán, N.; Queiroz, M.L. Myelopoietic response in tumour-bearing mice by an aggregated polymer isolated from Aspergillus oryzae. Eur. J. Pharmacol. 2000, 388, 219–226. [Google Scholar] [CrossRef]
- Lu, H.; Huang, J.; Li, G.; Ge, K.; Wu, H.; Huang, Q. Expression, purification and characterization of recombinant human serine proteinase inhibitor Kazal-type 6 (SPINK6) in Pichia pastoris. Protein Expr. Purif. 2012, 82, 144–149. [Google Scholar] [CrossRef] [PubMed]
- García-Fernández, R.; Ziegelmüller, P.; González, L.; Mansur, M.; Machado, Y.; Redecke, L.; Hahn, U.; Betzel, C.; de los Chávez, M.Á. Two variants of the major serine protease inhibitor from the sea anemone Stichodactyla helianthus, expressed in Pichia pastoris. Protein Expr. Purif. 2016, 123, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Moy, A.; Gomez, S.R.; Franz, A.H.; Lin-Cereghino, J.; Lin-Cereghino, G.P. An improved method for enhanced production and biological activity of human secretory leukocyte protease inhibitor (SLPI) in Pichia pastoris. Biochem. Biophys. Res. Commun. 2010, 402, 519–524. [Google Scholar] [CrossRef] [PubMed]
- Looser, V.; Brühlmann, B.; Bumbak, F.; Stenger, C.; Costa, M.; Camattari, A.; Fotiadis, D.; Kovar, K. Cultivation strategies to enhance productivity of Pichia pastoris: A review. Biotechnol. Adv. 2015, 33, 1177–1193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tschopp, J.F.; Brust, P.F.; Cregg, J.M.; Stillman, C.A.; Gingeras, T.R. Expression of the lacZ gene from two methanol-regulated promoters in Pichia pastoris. Nucleic Acids Res. 1987, 15, 3859–3876. [Google Scholar] [CrossRef] [PubMed]
- Dietzsch, C.; Spadiut, O.; Herwig, C. A dynamic method based on the specific substrate uptake rate to set up a feeding strategy for Pichia pastoris. Microb. Cell Fact. 2011, 10, 14. [Google Scholar] [CrossRef] [PubMed]
- Major, I.T.; Constabel, C.P. Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against Herbivores. Plant Physiol. 2008, 146, 888–903. [Google Scholar] [CrossRef] [PubMed]
- Terada, S.; Fujimura, S.; Kino, S.; Kimoto, E. Purification and characterization of three proteinase inhibitors from canavalia lineata seeds. Biosci. Biotechnol. Biochem. 1994, 58, 371–375. [Google Scholar] [CrossRef] [PubMed]
- Macedo, M.L.R.; Ribeiro, S.F.F.; Taveira, G.B.; Gomes, V.M.; de Barros, K.M.C.A.; Maria-Neto, S. Antimicrobial Activity of ILTI, a Kunitz-Type Trypsin Inhibitor from Inga laurina (SW.) Willd. Curr. Microbiol. 2016, 72, 538–544. [Google Scholar] [CrossRef] [PubMed]
- Pletzer, D.; Hancock, R.E.W. Antibiofilm peptides: Potential as broadspectrum agents. J. Bacteriol. 2016, 198, 2572–2578. [Google Scholar] [CrossRef] [PubMed]
- Le, K.Y.; Dastgheyb, S.; Ho, T.V.; Otto, M. Molecular determinants of staphylococcal biofilm dispersal and structuring. Front. Cell. Infect. Microbiol. 2014, 4, 167. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Bezerra, C.; De Oliveira, C.F.R.; Machado, O.L.T.; De Mello, G.S.V.; Da Rocha Pitta, M.G.; De Melo Rêgo, M.J.B.; Napoleão, T.H.; Paiva, P.M.G.; De Fátima Ferreira Ribeiro, S.; Gomes, V.M.; et al. Exploiting the biological roles of the trypsin inhibitor from Inga vera seeds: A multifunctional Kunitz inhibitor. Process Biochem. 2016, 51, 792–803. [Google Scholar] [CrossRef]
- Fang, E.F.; Wong, J.H.; Ng, T.B. Thermostable Kunitz trypsin inhibitor with cytokine inducing, antitumor and HIV-1 reverse transcriptase inhibitory activities from Korean large black soybeans. J. Biosci. Bioeng. 2010, 109, 211–217. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Protein (µg/mL) | Inhibitory Activity (U) | Specific Activity (U/mg) |
|---|---|---|---|
| GS9K | 64,571 | 30,53 | 0,18 |
| GSrILTI | 146,984 | 351,27 | 2,07 |
| Strains | MBIC (µg/mL) | MBEC (µg/mL) Range | ||||
|---|---|---|---|---|---|---|
| ILTI | rILTI | Vancomycin | ILTI | rILTI | Vancomycin | |
| S. epidermidis(ATCC 35984) | 25 | 25 | 15 | 150–200 | 120–220 | 80–100 |
| S. epidermidis(ATCC 12228) | NA | NA | NA | NA | NA | NA |
| S. aureusMSSA ATCC80958 | 20 | 20 | 25 | ⁻ | ⁻ | 80–120 |
| S. aureusMRSA (ATCC33591) | 25 | 30 | 30 | ⁻ | ⁻ | 100–250 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carneiro, F.C.; Weber, S.S.; Silva, O.N.; Jacobowski, A.C.; Ramada, M.H.S.; Macedo, M.L.R.; Franco, O.L.; Parachin, N.S. Recombinant Inga Laurina Trypsin Inhibitor (ILTI) Production in Komagataella Phaffii Confirms Its Potential Anti-Biofilm Effect and Reveals an Anti-Tumoral Activity. Microorganisms 2018, 6, 37. https://doi.org/10.3390/microorganisms6020037
Carneiro FC, Weber SS, Silva ON, Jacobowski AC, Ramada MHS, Macedo MLR, Franco OL, Parachin NS. Recombinant Inga Laurina Trypsin Inhibitor (ILTI) Production in Komagataella Phaffii Confirms Its Potential Anti-Biofilm Effect and Reveals an Anti-Tumoral Activity. Microorganisms. 2018; 6(2):37. https://doi.org/10.3390/microorganisms6020037
Chicago/Turabian StyleCarneiro, Fábio C., Simone S. Weber, Osmar N. Silva, Ana Cristina Jacobowski, Marcelo H. S. Ramada, Maria L. R. Macedo, Octávio L. Franco, and Nádia S. Parachin. 2018. "Recombinant Inga Laurina Trypsin Inhibitor (ILTI) Production in Komagataella Phaffii Confirms Its Potential Anti-Biofilm Effect and Reveals an Anti-Tumoral Activity" Microorganisms 6, no. 2: 37. https://doi.org/10.3390/microorganisms6020037