Cosmopolitanism and Biogeography of the Genus Manganonema (Nematoda: Monhysterida) in the Deep Sea



Abstract
:Simple Summary
The deep sea comprises more than 60% of the Earth surface, and likely represents the largest reservoir of as yet undiscovered biodiversity. Nematodes are the most abundant taxon on Earth and are particularly abundant and diverse in the deep sea. Nevertheless, knowledge of their biogeography especially in the deep sea is still at its infancy. This article explores the distribution of the genus Manganonema in the deep Atlantic Ocean and Mediterranean Sea providing new insights about this apparently rare deep-sea genus.
Abstract
Spatial patterns of species diversity provide information about the mechanisms that regulate biodiversity and are important for setting conservation priorities. Present knowledge of the biogeography of meiofauna in the deep sea is scarce. This investigation focuses on the distribution of the deep-sea nematode genus Manganonema, which is typically extremely rare in deep-sea sediment samples. Forty-four specimens of eight different species of this genus were recorded from different Atlantic and Mediterranean regions. Four out of the eight species encountered are new to science. We report here that this genus is widespread both in the Atlantic and in the Mediterranean Sea. These new findings together with literature information indicate that Manganonema is a cosmopolitan genus, inhabiting a variety of deep-sea habitats and oceans. Manganonema shows the highest diversity at water depths >4,000 m. Our data, therefore, indicate that this is preferentially an abyssal genus that is able, at the same time, to colonize specific habitats at depths shallower than 1,000 m. The analysis of the distribution of the genus Manganonema indicates the presence of large differences in dispersal strategies among different species, ranging from locally endemic to cosmopolitan. Lacking meroplanktonic larvae and having limited dispersal ability due to their small size, it has been hypothesized that nematodes have limited dispersal potential. However, the investigated deep-sea nematodes were present across different oceans covering macro-scale distances. Among the possible explanations (hydrological conditions, geographical and geological pathways, long-term processes, specific historical events), their apparent preference of colonizing highly hydrodynamic systems, could suggest that these infaunal organisms are transported by means of deep-sea benthic storms and turbidity currents over long distances.
1. Introduction
Nematodes are the most abundant metazoans on Earth [1], they are ubiquitous in marine habitats [2] and their numerical dominance increases in the deep sea (up to >90%) with increasing water depth [3-6]. This phylum is characterized by a very high species number: more than 20,000 have been described [7], and, among them, about 4,000-5,000 are marine species. Meanwhile, the number of marine nematode species is evaluated from 10,000 to 20,000 species according to the most conservative estimation [8]. In particular, deep-sea nematode assemblages are characterized by a very high α-diversity and evenness, which is comparable with that of the tropical sub-littoral zone [9]. Even if the deep sea comprises about 91% of the ocean seafloor, investigations focused on the study of nematode diversity have been so far performed on a cumulative area of only 60–70 m2 [10]. Moreover, because of the small size, the taxonomy of deep-sea nematodes is difficult [11] and information on their distribution remains extremely scant [12].
Spatial patterns of species diversity provide information about the mechanisms that regulate biodiversity [13,14] and are important for setting conservation priorities [15]. Nevertheless, the present knowledge of the biogeography of deep-sea meiofauna is limited [16,17]. Cosmopolitan species are known among shallow-water nematodes [18]. Cosmopolitanism is known also for some deep-sea nematode genera, whilst only a few genera have been reported to inhabit a single habitat [19]. The cosmopolitanism of most genera does not necessarily apply to all species of the genus; and several studies suggest the presence of large differences in species distribution at both local and regional scale [10,12,19-22]. Studies on deep-sea nematode species distribution over large areas demonstrate that the distribution of some species is widespread [7,12,23]. For example, among the nine species of the genus Dichromadora described from the Weddell Sea, seven are found at more than one location and can cover distances in excess of 2,500 km [12].
This study focuses on the distribution of the deep-sea nematode genus Manganonema (Bussau 1993). This genus belong to the family Monhysteridae, recognized as one of the most under-investigated deep sea taxa [10]. Manganonema has been recorded in very low abundances (typically <2%), at water depths >600 m in many oceans [24-35]. As previously reported for the genera Bathyeurystomina and Bathychaetosoma, the genus Manganonema is apparently restricted to the deep sea. At present, the genus Manganonema contains six described species [29]. Different Manganonema species are typically reported from different sites, suggesting a potentially restricted distribution [29]. The aim of the present study is to investigate large-scale biogeography of the genus Manganonema in the deep sea. Forty-four specimens of eight different species were recorded from different Atlantic and Mediterranean regions. Four of the encountered species are new to science. The geographical and bathymetrical distributions of Manganonema genus and species were analyzed in order to test the cosmopolitan character of this genus also at species level.
2. Experimental Section
Specimens were collected from 20 distinct deep-sea sites from the Atlantic Ocean and the Mediterranean Sea at water depth ranging from 567 to 4,987 (Table 1). Sediment samples were collected from several oceanographic cruises in the North Atlantic at Rockall Through (R/V Pelagia, 2006), and off the Portugal coast (R/V Pelagia, 2006), in the Gulf of Guinea (R/V Pourquoi pas?, 2008), in the Eastern Atlantic and in the Western and in the Central Mediterranean Sea (R/V Urania, 2007, 2008 and 2009) and in the Eastern Mediterranean Sea (R/Vs Aegaeo and Universitatis, 2006 and 2007 respectively). Sediment samples were collected by multiple-, box- and interface-corers and preserved in buffered 4% formalin solution and stained with Rose Bengal.
Sediment samples were pre-screened through a 1,000-μm mesh net, and the organisms retained on a 20-μm mesh net. The fraction remaining on the latter sieve was re-suspended and centrifuged three times with Ludox HS40 (density 1.31 g cm3) and nematodes were mounted on slides after formalin-ethanol-glycerol treatment following the detailed protocols reported by Danovaro [36].
Drawings and photos were made on Leica DMLS microscope. Cobb formula, showing the distance of each character from anterior end was calculated for each specimen. Ratio a represents the body length divided by the body maximum width, ratio b represents the body length divided by the pharynx length, and ratio c represents body length divided by the tail length.
3. Results and Discussion
3.1. Distribution of the Genus Manganonema
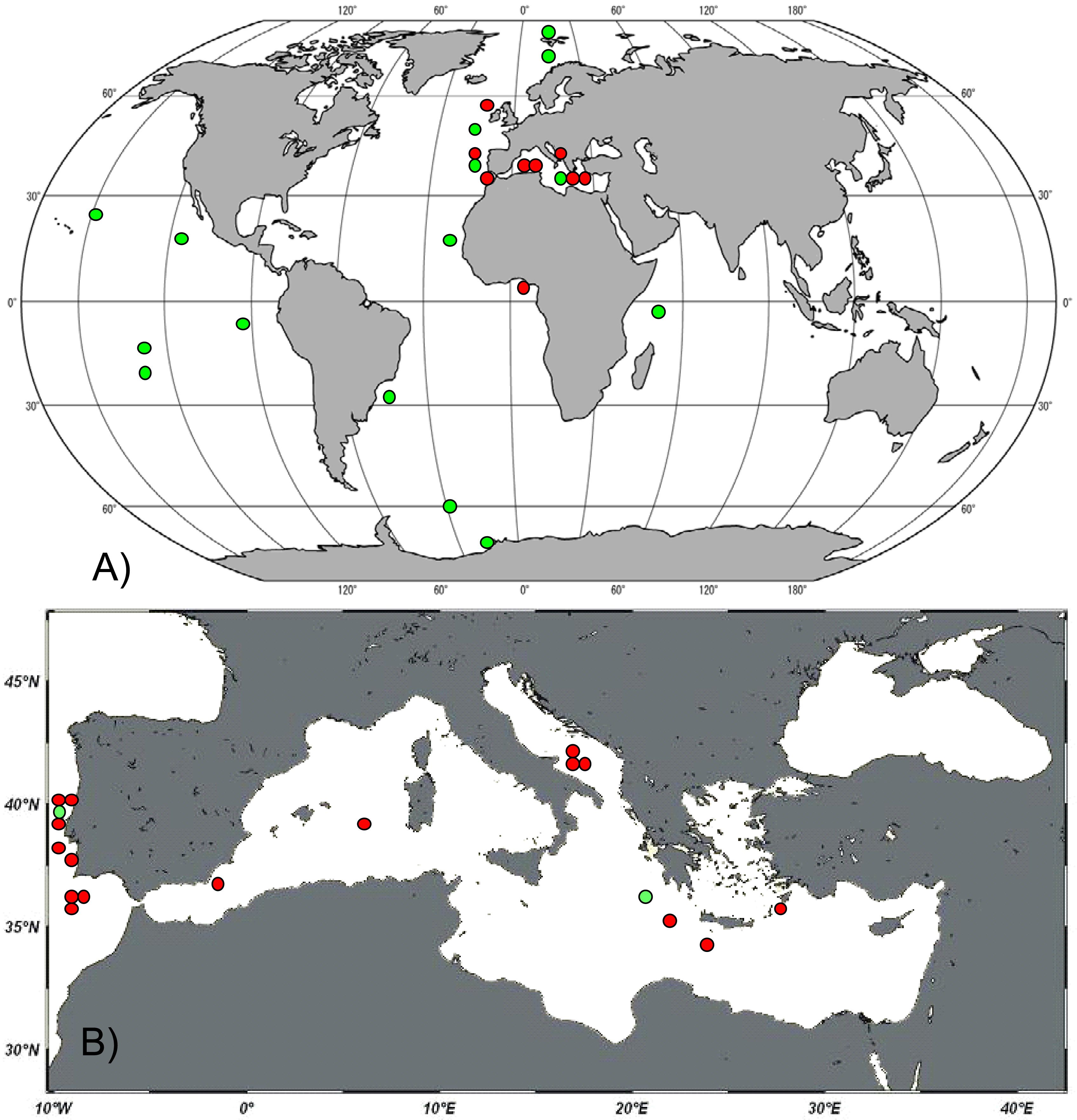
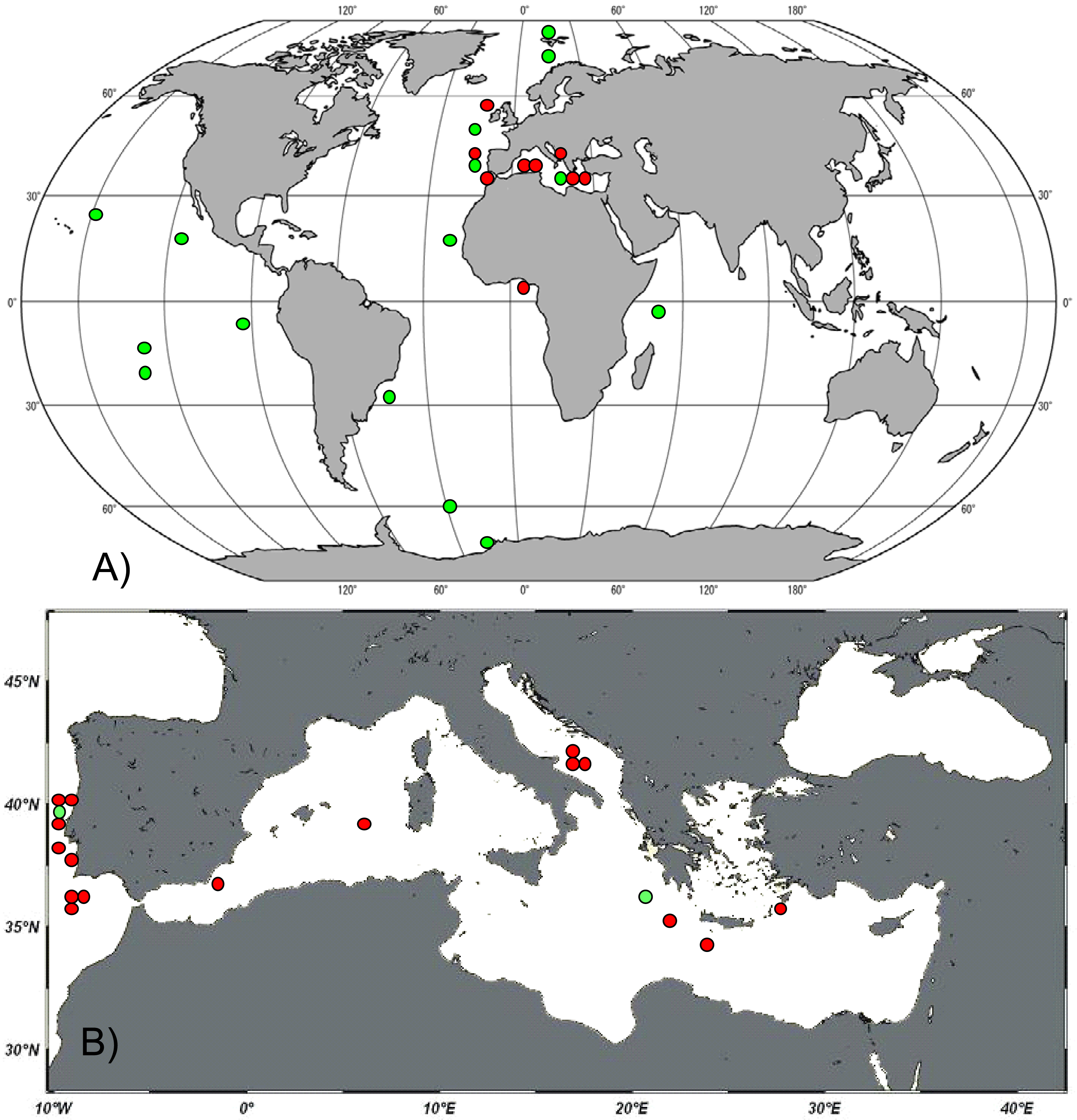
We encountered the genus Manganonema in 20 different deep-sea sites from the Atlantic Ocean and the Mediterranean Sea (Table 1). The presence of the genus Manganonema was recorded in the Atlantic Ocean (Rockall Through, Portuguese Margin, Gulf of Cadiz and Gulf of Guinea) and in the Mediterranean Sea (Western, Central and Eastern). Figure 1(A) shows the distribution of Manganonema at the global scale, including the records available in literature. The new findings of Manganonema ssp. specimens together with literature data confirm the cosmopolitan character of this genus, which can be found in all oceans (Atlantic, Indian, Pacific, Arctic, and Antarctic) and at latitudes spanning from 70°N to 60°S. We discovered Manganonema in 16 specimens from the Mediterranean Sea (Figure 1(B)). Only one study previously documented the presence of this genus in the Mediterranean Sea [31]. Manganonema was recorded in three sites of the Eastern Mediterranean Sea, in three sites of the Southern Adriatic Sea and in two sites in the Western Mediterranean Sea. This demonstrates that the genus Manganonema is widespread in the deep-sea sediments of the Mediterranean Sea, from the Eastern part of the basin to the Atlantic Ocean. In addition half of the Manganonema species identified in the present study were reported in the Mediterranean Sea, including one species new to science. This supports the hypothesis that the deep Mediterranean basin provides a variety of environmental conditions that might favor the colonization of several rare species [22].
3.2. Bathymetric Distribution of the Genus Manganonema
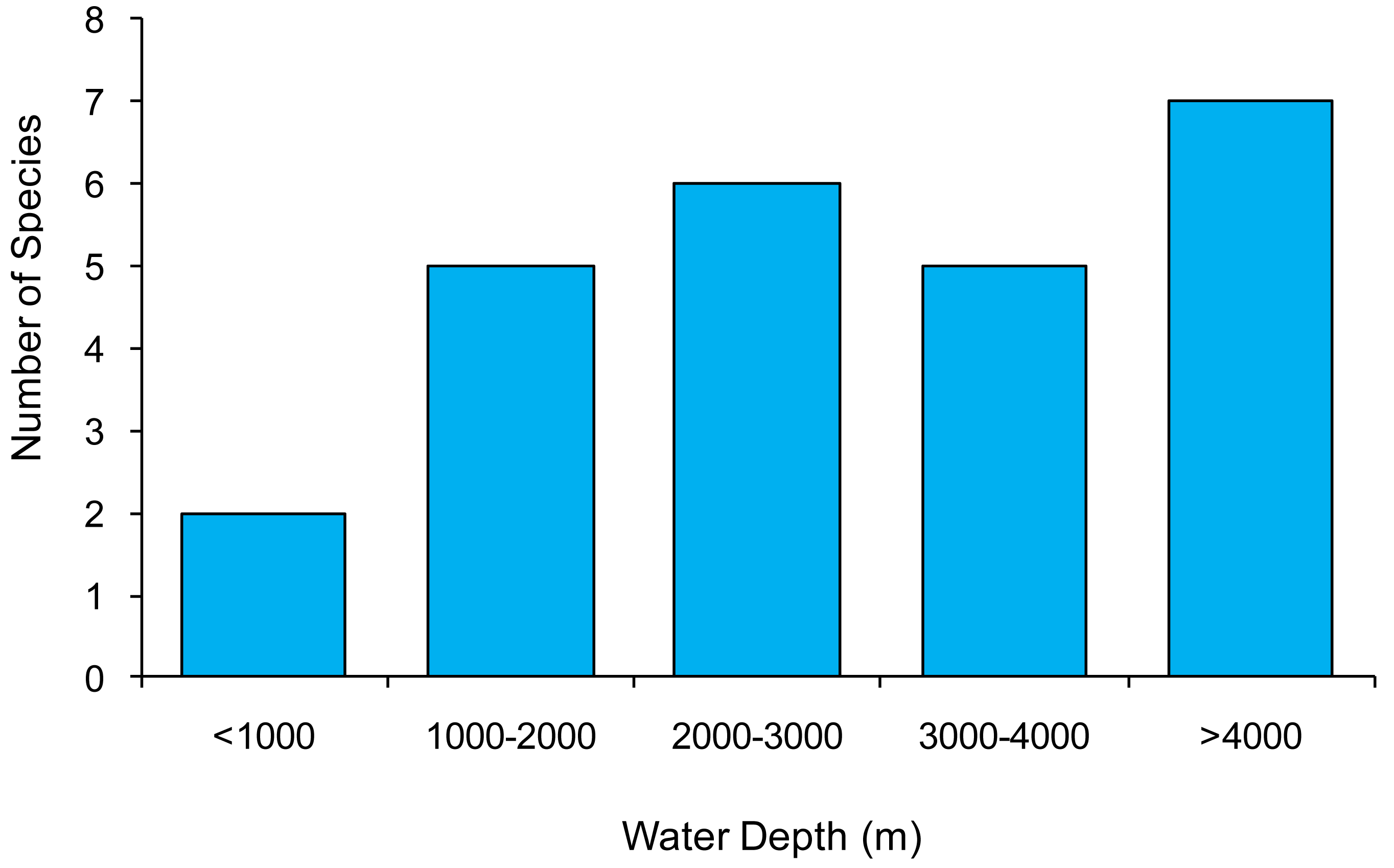
Our data show that Manganonema is present at depths ranging from 567 to 4,997 m. The highest diversity (7 species) was found at a water depth >4,000 m (Figure 2). Manganonema is considered a deep-sea genus as for Bathyeurystomina and Bathychaetosoma [29]. However, two species encountered at water depths <1,000 m in the North-east Atlantic Ocean (Rockall Trough, 567 m) and in the Central Mediterranean Sea (Southern Adriatic Sea, between 590–824 m) indicate that this genus can be piezotolerant. The only other study that described the presence of Manganonema at shallow depths (600 m) was in the Weddell Sea [29], this was explained by the fact that the peculiar conditions of this marine system could be compared with those of the deepest ocean, and thus offer the opportunity to this genus to colonize shallow sediments. Accordingly, here we discovered Manganonema in shallow sites that are characterized by habitats such as cold-water corals, seamounts, erosional features and different physical conditions (e.g., bottom temperature of the Mediterranean Sea and Atlantic Ocean), which share the presence of high hydrodynamics (i.e., strong bottom currents).
3.3. Habitats Inhabited by Manganonema
The genus Manganonema was previously reported from diverse deep-sea habitats, from the Arctic to the Antarctic Ocean, in reduced ecosystems (hydrothermal vents and mud volcanoes), in submarine canyons and in polymetallic nodule deposits [24,26-29,32-35]. Here we discovered Manganonema in cold-water coral habitats and in seamounts. Furthermore, we recorded Manganonema also in different morphologies and sedimentary features related to submarine landslides, unstable slopes or seafloor erosion (e.g., mud waves and furrows). This study reinforces the idea that the genus Manganonema is able to colonize a very different variety of deep-sea habitats.
3.4. Distribution of the Manganonema Species
Some meiofaunal species (including nematodes) display a wide spatial distribution [37] and a trans-oceanic dispersal of free-living marine nematodes is not rare [18]. Most dominant deep-sea nematode genera are considered to be cosmopolitan, but at the species level little is known about the geographic distribution of their species.
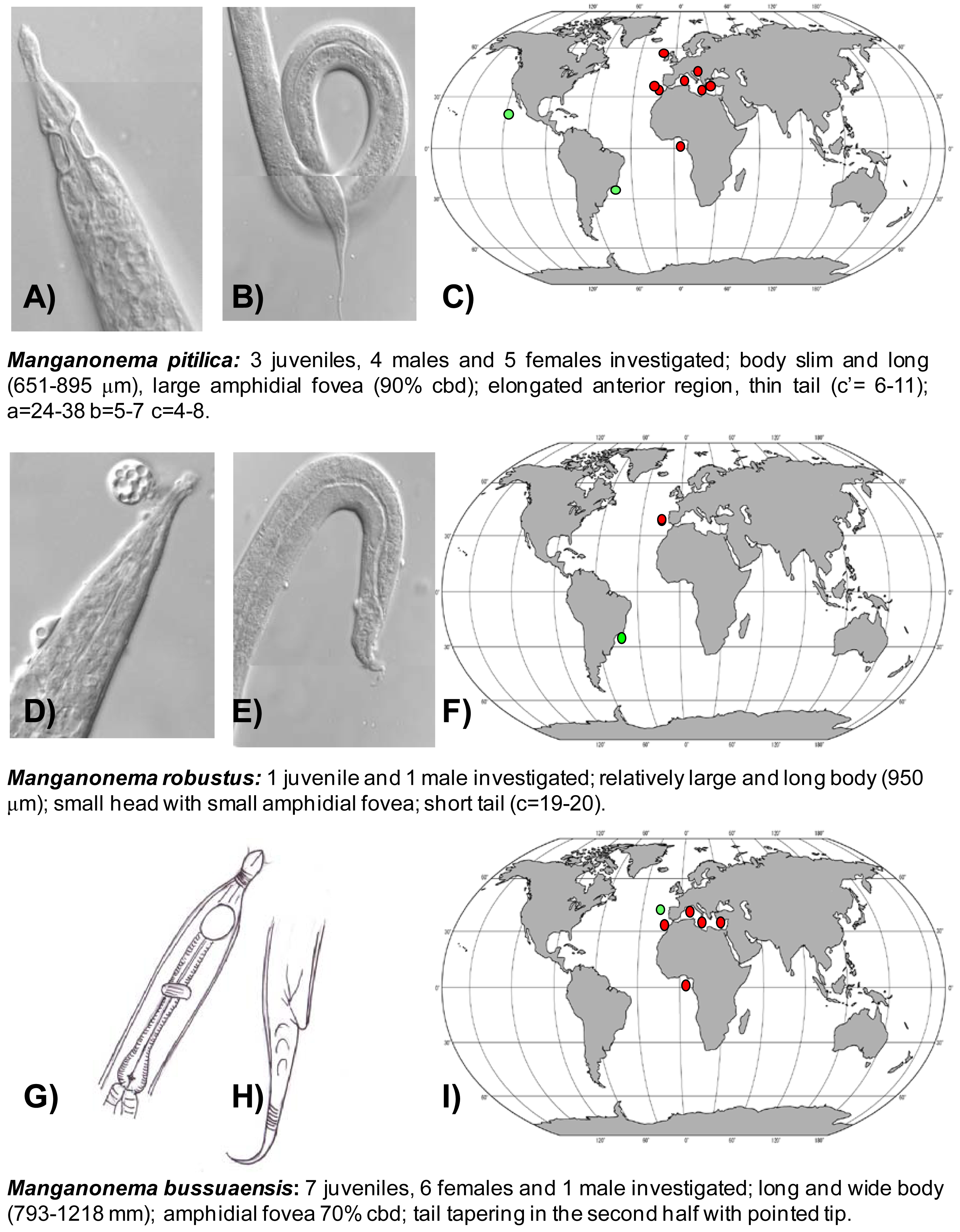
The first exhaustive analysis of the Manganonema species hypothesised a restricted distribution of this genus [29]. However, subsequent studies revealed that some species of Manganonema are not endemic [35]. In the present study 44 specimens of Manganonema belonging of eight species were analysed. Although a description of the new Manganonema species is beyond the scope of this work, a pictorial key is provided in Figure 3, Figure 4 and Figure 5. Species represented only by females and juveniles were considered as putative species (Manganonema sp.2 and Manganonema sp.3).
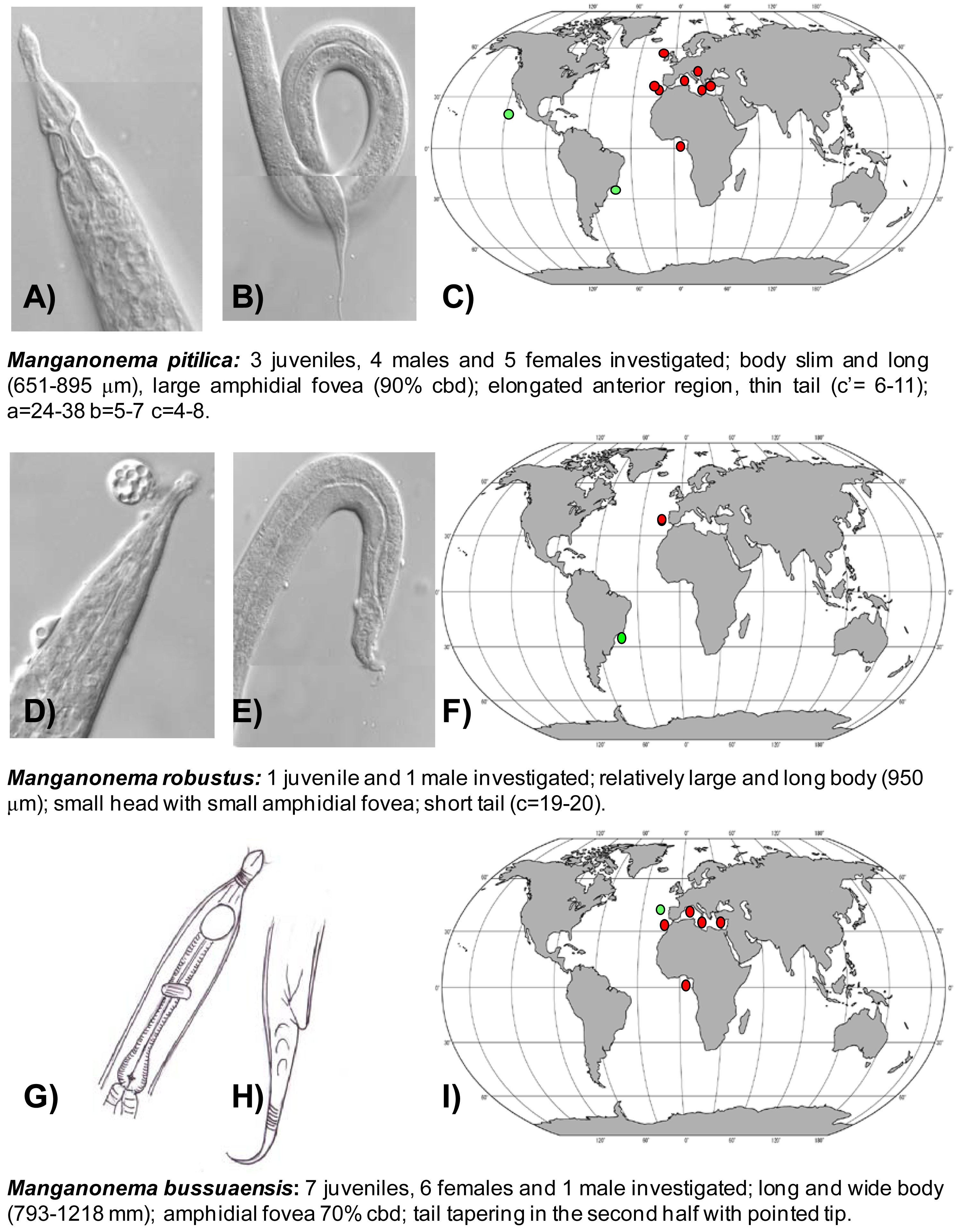
Manganonema pitilica (Fonseca, Decraemer, Vanreusel, 2006) was previously found in the Pacific Ocean and in the South-west Atlantic Ocean (Figure 3(A,B,C)) [29,35]. We found this species in the North-east Atlantic (Rockall Trough, Portuguese Margin, Gulf of Cadiz, and in Gulf of Guinea) and in the Western, Central and Eastern Mediterranean Sea at all investigated depths. M. pitilica is widely distributed and able to colonize different habitats from manganese nodule areas [35] to seamounts and cold-water coral ecosystems.
Manganonema robustus (Fonseca, Decraemer, Vanreusel, 2006) was discovered in the North-east Atlantic Ocean (Portuguese Margin) at water depths between 2,000 and 5,000 m. This species was previously reported only in the Southwest Atlantic, off the Brazilian coasts (Figure 3(D,E,F)) [29].
Manganonema bussuaensis (Fonseca, Decraemer, Vanreusel, 2006) was found in the North-east Atlantic Ocean (Gulf of Cadiz and Gulf of Guinea) and in Western, Central and Eastern Mediterranean Sea from 714 to 4,400 m water depth. This species was previously reported only in the North Atlantic at 2,000 m water depth (Figure 3(G,H,I)) [29].
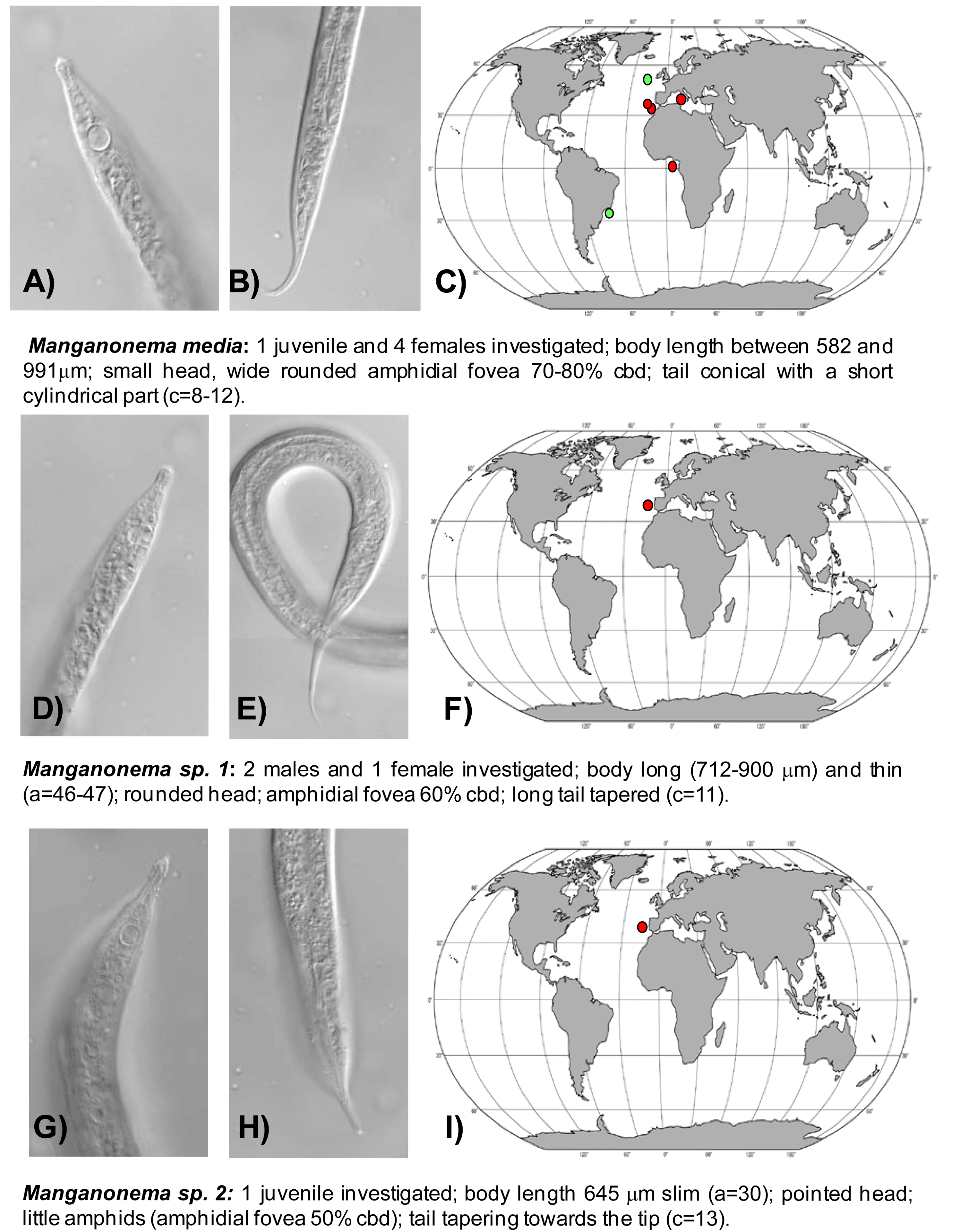
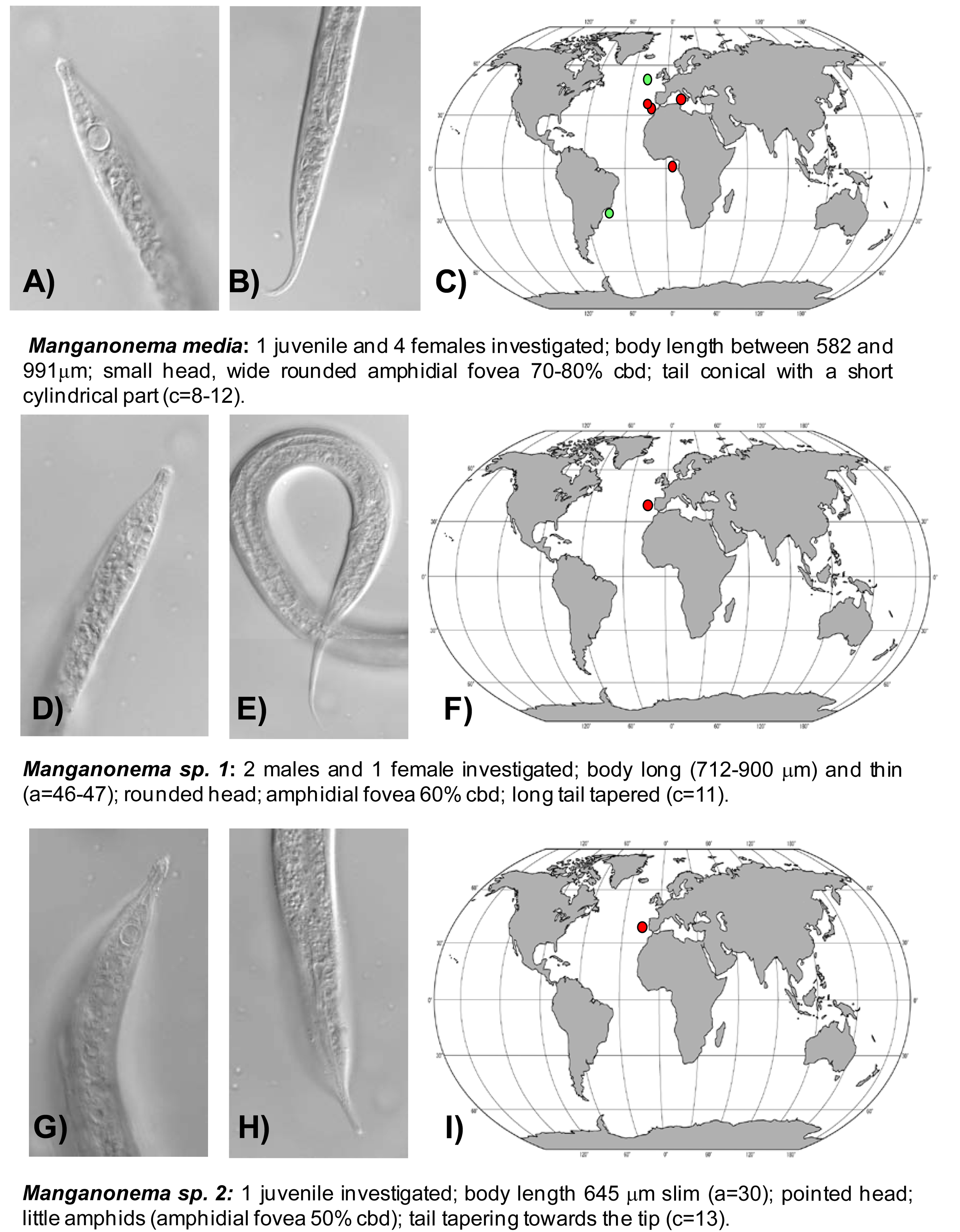
Manganonema media (Fonseca, Decraemer, Vanreusel, 2006) was encountered in the North-east Atlantic Ocean (Portuguese Margin, Gulf of Cadiz and Gulf of Guinea) and in Western Mediterranean Sea at water depth between 2000 and 5000 m. This species was previously reported only in the South-west Atlantic, off the Brazilian coasts and in the North-east Atlantic, at Goban Spur (Figure 4(A,B,C)) [29].
Manganonema sp. 1 was found in the North-east Atlantic Ocean (Portuguese Margin) at 2,000 and 4,000 m water depth (Figure 4(D,E,F)).
Manganonema sp. 2 (putative species) was encountered in the North-east Atlantic Ocean (Portuguese Margin) at 3475 m water depth (Figure 4(G,H,I)).
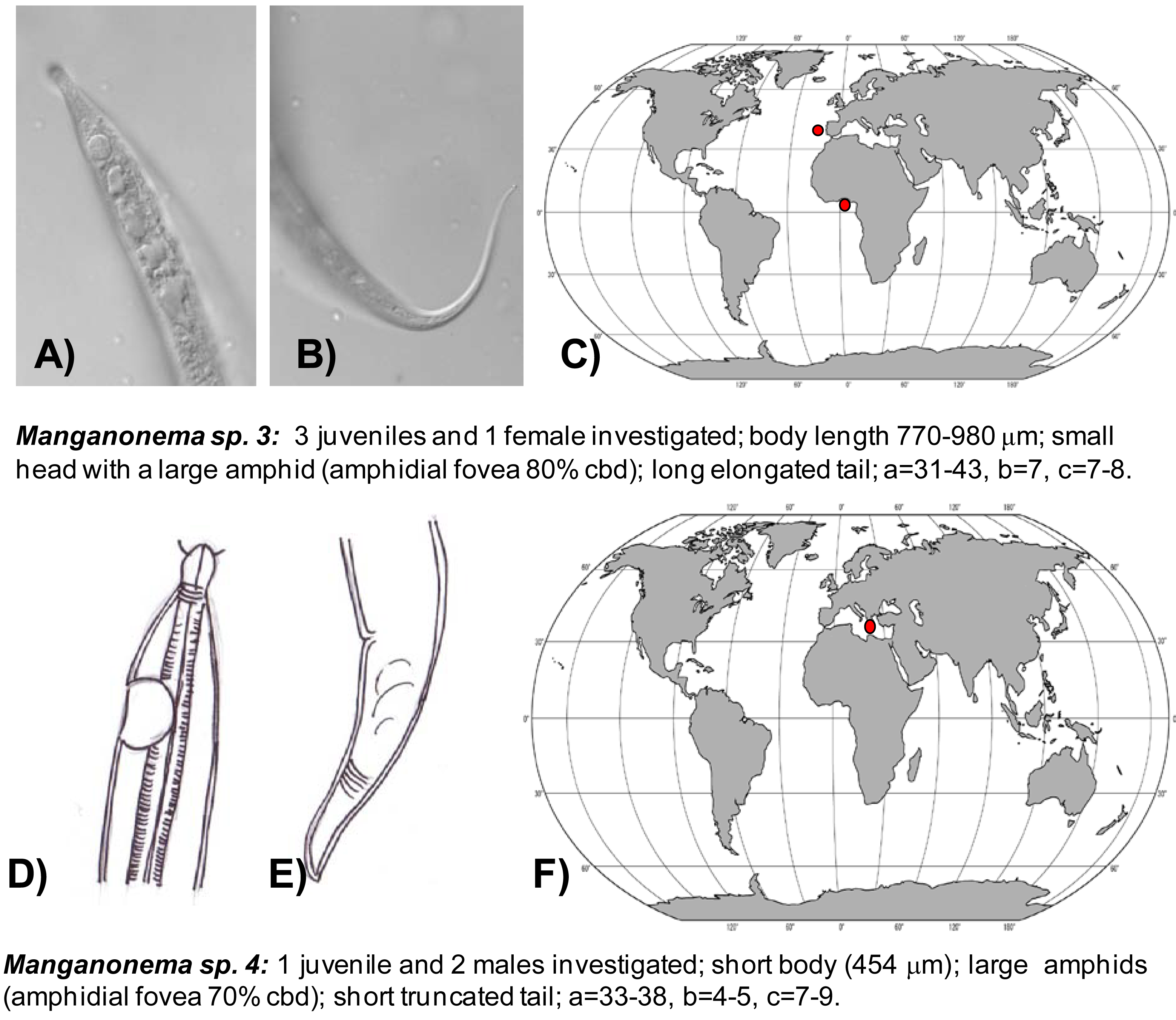
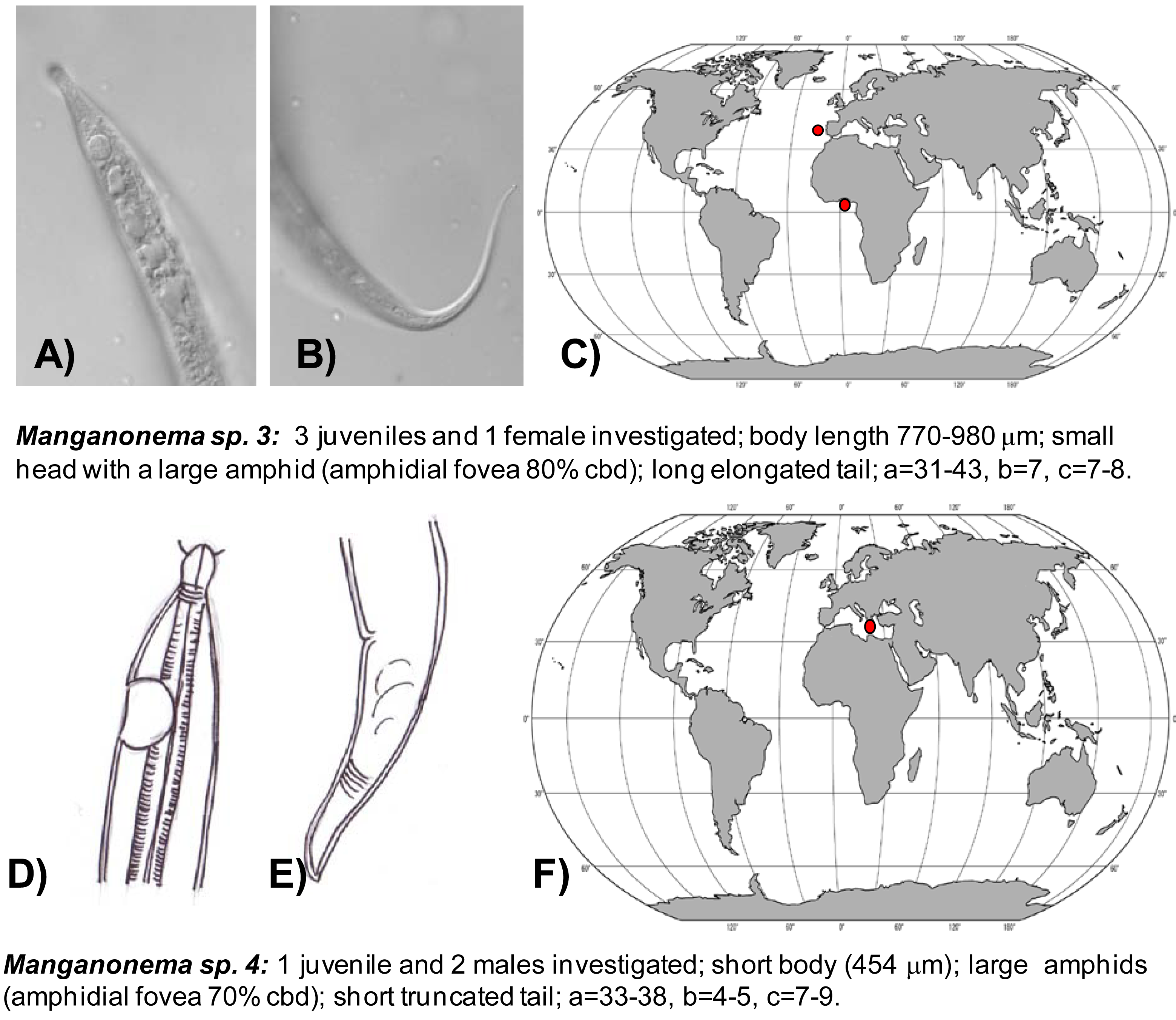
Manganonema sp. 3 (putative species) was reported in the North-east Atlantic Ocean (Portuguese Margin and Gulf of Guinea) at water depth between 1,700 and 5,000 m (Figure 5(A,B,C)).
Manganonema sp. 4 was encountered in the Eastern Mediterranean Sea at 3000 m water depth (Figure 5(D,E,F)).
Finally, in order to provide a complete review on biogeography and distribution of the Manganonema genus, the distribution of the remaining described species (not encountered in this study) are presented.
Manganonema antarctica was reported in two different sites of the Southern Ocean: the Weddell Sea and the South Sandwich Trench at water depth between 600 and 4,000 m [29].
Manganonema microchepalum was reported in the Peru Basin, Central Pacific Ocean at 4,000 m water depth [26,29].
The results of this study suggest that Manganonema species are not isolated as previously supposed [29]. Based on distribution patterns of single species we observed that Manganonema species can be either apparently restricted or widespread and one Manganonema species (M. pitilica) is a possible cosmopolitan species. Furthermore, at some sites, up to 3 different species were found to coexist, supporting the hypothesis of a trophic specialization and resulting in an important contribution to the local species richness [29].
3.5. Dispersal Mechanisms of Deep-Sea Nematodes
Some Manganonema species are apparently confined to specific areas, while others have a much wider spatial distribution. These contrasting dispersal strategies may be linked to their behaviour or life cycle. Adult nematodes are expected to have limited dispersal capabilities and hence a reduced gene flow, making restricted species distributions more plausible [38]. However some deep-sea nematodes have been demonstrated to have important dispersal capabilities [7]. The continental and cosmopolitan distributions of many meiofaunal taxa appear to result from a variety of dispersive mechanisms. Several geological, geographical and hydrodynamic pathways (i.e., passive erosive suspension, active emergence, seamounts, sediment transports) and natural/anthropogenic rafting should be considered [2]. Adult benthic organisms can be transported also within moving sediment, for example benthic storms, submarine landslides and turbidity currents can carry large amounts of sediment over long distances [39,40], with an additional effect of defaunating large areas that are then free for re-colonisation. In addition to the adult macrofaunal organisms, larvae and buoyant eggs can stay in the water column for a long time, extending their potential dispersal [41]. The lack of meroplanktonic larvae or buoyant eggs does not allow nematodes to use these strategies for their dispersal, but sediment resuspension due to bottom currents or benthic storms can certainly contribute to the dispersal of the nematodes. This could apply particularly to the genus Manganonema that apparently prefers habitats characterized by high hydrodynamic conditions.
4. Conclusions
Manganonema is a cosmopolitan genus, inhabiting all oceans (Atlantic, Indian, Pacific, Arctic, and Antarctic) and spanning across a large variety of deep-sea habitats. This genus can be found at latitudes ranging from 70°N to 60°S. We report here that the genus Manganonema is widespread also in both basins of the Mediterranean Sea. Fifty percent of the species encountered were present also in the Mediterranean Sea, and one of these is new to science. Manganonema is a deep-sea genus characterized by the highest diversity at depths >4,000 m. However, this genus is able to colonize also sediments at a water depth <1,000 m, where peculiar environmental conditions exist, for example the presence of strong near bottom currents.
Finally, the results of this study show that Manganonema showed remarkable differences among species, spanning the entire range between endemic to cosmopolitan. This genus shows the co-presence of several species, indicating a potential local diversity. Moreover, half of the species encountered are new to science, revealing the potential and hidden biodiversity of deep-sea nematodes. In spite of the evidence that adult nematodes have limited dispersal capabilities, the deep-sea nematodes revealed a high dispersion. A possible explanation is that infaunal organisms, and thus also nematodes, can be transported along with surface sediments by turbidity currents and benthic storms over long distances.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Station | Latitude | Longitude | Water depth (m) | Region | Habitat | Manganonema specimens |
|---|---|---|---|---|---|---|
| 1 | 55°29.71′N | 15°48.56′W | 567 | North-east Atlantic Ocean (Rockall Through) | Cold-water corals | 1 |
| 2 | 41°52.54′N | 17°00.47′E | 590 | Central Mediterranean Sea (Southern Adriatic Sea) | Erosional structure (mud wave) | 2 |
| 3 | 41°47.34′N | 17°01.85′E | 714 | Central Mediterranean Sea (Southern Adriatic Sea) | Erosional structure (furrow) | 1 |
| 4 | 41°31.88′N | 17°25.15′E | 824 | Central Mediterranean Sea (Southern Adriatic Sea) | Dauno seamount | 1 |
| 5 | 02°57.26′N | 06°49.51′E | 1,671 | North-east Atlantic Ocean (Gulf of Guinea) | Open continental slope | 3 |
| 6 | 02°56.87′N | 06°49.44′E | 1,701 | North-east Atlantic Ocean (Gulf of Guinea) | Submarine landslides | 3 |
| 7 | 34°40.26′N | 24°07.66′E | 1,998 | Eastern Mediterranean Sea | Unstable continental slope | 1 |
| 8 | 37°50.00′N | 09°45.00′W | 2,130 | North-east Atlantic Ocean (Portuguese Margin) | Open continental slope | 2 |
| 9 | 36°47.38′N | 00°29.06′W | 2,689 | Western Mediterranean Sea | Open continental slope | 2 |
| 10 | 35°00.34′N | 08°16.27′W | 2,788 | North-east Atlantic Ocean (Gulf of Cadiz) | Open continental slope | 6 |
| 11 | 39°18.80′N | 06°04.25′E | 2,855 | Western Mediterranean Sea | Open continental slope | 1 |
| 12 | 35°08.33′N | 20°50.88′E | 3,000 | Eastern Mediterranean Sea | Open continental slope | 6 |
| 13 | 35°57.64′N | 28°17.63′E | 3,009 | Eastern Mediterranean Sea | Open continental slope | 2 |
| 14 | 40°10.00′N | 09°59.99′W | 3,475 | North-east Atlantic Ocean (Portuguese Margin) | Open continental slope | 1 |
| 15 | 39°13.74′N | 10°59.00′W | 4,060 | North-east Atlantic Ocean (Portuguese Margin) | Unstable continental slope | 3 |
| 16 | 34°37.49′N | 09°17,00′W | 4,335 | North-east Atlantic Ocean (Gulf of Cadiz) | Open continental slope | 1 |
| 17 | 34°32.75′N | 09°45.38′W | 4,381 | North-east Atlantic Ocean (Gulf of Cadiz) | Open continental slope | 1 |
| 18 | 34°08.02′N | 09°33.28′W | 4,385 | North-east Atlantic Ocean (Gulf of Cadiz) | Open continental slope | 3 |
| 19 | 40°10.01′N | 10°59.99′W | 4,902 | North-east Atlantic Ocean (Portuguese Margin) | Open continental slope | 2 |
| 20 | 37°50.01′N | 11°00.01′W | 4,987 | North-east Atlantic Ocean (Portuguese Margin) | Open Continental Slope | 2 |
Acknowledgements
This research was supported by a research grant provided by “Fondazione Luigi e Francesca Brusarosco”, in the framework of the projects funded by the European Commission HERMES (Hot Spot Ecosystem Research on the Margins of European Seas FPVI, Contract No. GOCE-CT-2005-511234-1) and HERMIONE (Hotspot Ecosystem Research and Man's Impact on European Seas FP7-ENV-2008-1, Contract No. 226354), the project funded by ESF BIOFUN (BIOdiversity and Ecosystem FUNctioning Contrasting Southern European Deep-Sea Environments), and the Italian project OBAMA (COFIN-PRIN). The authors are indebted to Marianna Mea and Cristina Gambi (DISMAR, Italy) for providing specimens of Manganonema, to Michele Voisset and Antonio Cattaneo (cruise ERIG3D, IFREMER, France) for support in the sampling activities, and to the crews of the R/Vs Urania and Universitatis (Italy), Pelagia (The Netherlands), Aegeao (Greece) and Pourquoi pas? (France) for their valuable help during the sea-going activities.
Conflict of Interest
The authors declare no conflict of interest.
References
- Lambshead, P.J.D. Marine nematode biodiversity. In Nematode Morphology, Physiology and Ecology; Chen, Z.X., Chen, S.Y., Dickson, D.W., Eds.; University Press: Beijing, China, 2004; Volume 1, pp. 438–492. [Google Scholar]
- Giere, O. Meiobenthology. The Microscopic Motile Fauna of Aquatic Sediments; Giere, O., Ed.; Springer-Verlag: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- Thiel, H. The size structure of the deep-sea benthos. Int. Rev. Gesamten Hydrobiol. 1975, 60, 575–606. [Google Scholar]
- Heip, C.; Vincx, M.; Vranken, G. The ecology of marine nematodes. Oceanogr. Mar. Biol. Annu. Rev. 1985, 23, 399–489. [Google Scholar]
- Lambshead, P.J.D.; Schalk, P. Overview of marine invertebrate biodiversity. In Encyclopaedia of Biodiversity; Levin, S., Ed.; Academic Press: San Diego, CA, USA, 2001; Volume 1, pp. 543–559. [Google Scholar]
- Danovaro, R.; Gambi, C.; Della Croce, N. Meiofauna hotspot in the Atacama Trench (southern Pacific Ocean). Deep-Sea Res. I 2002, 49, 843–857. [Google Scholar]
- Lambshead, P.J.D.; Boucher, G. Marine nematode deep-sea biodiversity—Hyperdiverse or hype? J. Biogeogr. 2003, 30, 475–485. [Google Scholar]
- Mokievsky, V.; Azovsky, A. Re-evaluation of species diversity patterns of free-living marine nematodes. Mar. Ecol. Prog. Ser. 2002, 238, 101–108. [Google Scholar]
- Boucher, G.; Lambshead, P.J.D. Ecological biodiversity of marine nematodes in samples from temperate, tropical, and deep-sea regions. Conserv. Biol. 1995, 9, 1594–1604. [Google Scholar]
- Miljutin, M.D.; Gad, G.; Miljutina, M.M.; Mokievsky, V.O.; Fonseca-Genevois, V.; Esteves, A.M. The state of knowledge on deep-sea nematode taxonomy: How many valid species are known down there? Mar. Biodiv. 2010, 40, 143–159. [Google Scholar]
- Godfray, H.C.J. Challenges for taxonomy—The discipline will have to reinvent itself if it is to survive and flourish. Nature 2002, 417, 17–19. [Google Scholar]
- Vermeeren, H.; Vanreusel, A.; Vanhove, S. Species distribution within the free-living marine nematode genus Dichromadora in the Weddell Sea and adjacent areas. Deep-Sea Res. II 2004, 51, 1643–1664. [Google Scholar]
- Holyoak, M.; Leibold, M.A.; Holt, R.D. Metacommunities: Spatial Dynamics and Ecological Communities; Holyoak, M., Leibold, M.A., Holt, R.D., Eds.; Chicago University Press: Chicago, IL, USA, 2005. [Google Scholar]
- Gaston, K.; Blackburn, T. Pattern and Process in Macroecology; Blackwell Science: Oxford, UK, 2005. [Google Scholar]
- Ferrier, S.; Powell, G.V.N.; Richardson, K.S.; Manion, G.; Overtone, J.M.; Allnutt, T.F.; Cameron, S.E.; Mantle, K.; Burgess, N.D.; Faith, D.P.; Lamoreux, J.F.; Kier, G.; Hijmans, R.J.; Funk, V.A.; Cassis, G.A.; Fisher, B.L.; Flemons, P.; Lees, D.; Lovett, J.C.; Van Rompaey, R.S.A.R. Mapping more of terrestrial biodiversity for global conservation assessment. BioScience 2004, 54, 1101–1109. [Google Scholar]
- Fonseca, G.; Vanreusel, A.; Decraemer, W. Taxonomy and biogeography of Molgolaimus Ditlvsen, 1921 (Nematoda: Chromadoria) with reference to the origins of deep sea nematodes. Antarct. Sci. 2006, 18, 23–50. [Google Scholar]
- Gheerardyn, H.; Veit-Köhler, G. Diversity and large-scale biogeography of Paramesochridae (Copepoda, Harpacticoida) in South Atlantic Abyssal Plains and the deep Southern Ocean. Deep-Sea Res. I 2009, 56, 1804–1815. [Google Scholar]
- Decraemer, W.; Gourbault, N.; Helleouet, M.N. Cosmopolitanism among nematodes: Examples from Epsilonematidae. Vie Milieu 2001, 51, 11–19. [Google Scholar]
- Vanreusel, A.; Fonseca, G.; Danovaro, R.; DaSilva, M.; Esteves, A.; Ferrero, T.; Gad, G.; Galtsova, V.; Gambi, C.; da Fonseca Genevois, V.; Ingels, J.; Ingole, B.; Lampadariou, N.; Merckx, B.; Miljutin, D.; Miljutina, M.; Muthumbi, A.; Netto, S.; Portnova, D.; Radziejewska, T.; Raes, M.; Tchesunov, A.; Vanaverbeke, J.; Van Gaever, S.; Venekey, V.; Bezerra, T.N.; Flint, H.; Copley, J.; Pape, E.; Zeppilli, D.; Arbizu Martinez, P.; Galeron, J. The contribution of deep-sea macrohabitat heterogeneity to global nematode diversity. PSZN I: Mar. Ecol. 2010, 31, 6–20. [Google Scholar]
- Ingels, J.; Vanhove, S.; De Mesel, I.; Vanreusel, A. The biodiversity and biogeography of the free-living nematode genera Desmodora and Desmodorella (family Desmodoridae) at both sides of the Scotia Arc. Polar Biol. 2006, 29, 936–949. [Google Scholar]
- Fonseca, G.; Muthumbi, A.W.; Vanreusel, A. Species richness of the genus Molgolaimus (Nematoda) from local to ocean scale along continental slopes. PSZN I: Mar. Ecol. 2007, 28, 446–459. [Google Scholar]
- Danovaro, R.; Bianchelli, S.; Gambi, C.; Mea, M.; Zeppilli, D. α-, β-, γ-, δ- and ε-diversity of deep-sea nematodes in canyons and open slopes of Northeast Atlantic and Mediterranean margins. Mar. Ecol. Prog. Ser. 2009, 396, 197–209. [Google Scholar]
- De Mesel, I.; Lee, H.J.; Vanhove, S.; Vincx, M.; Vanreusel, A. Species diversity and distribution within the deep-sea nematode genus Acantholaimus on the continental shelf and slope in Antarctica. Polar Biol. 2006, 29, 860–871. [Google Scholar]
- Vanreusel, A.; Van den Bossche, I.; Thiermann, F. Free-living marine nematodes from hydrothermal sediments: similarities with communities from diverse reduced habitats. Mar. Ecol. Prog. Ser. 1997, 157, 207–219. [Google Scholar]
- Vanhove, S.; Arntz, W.; Vincx, M. Comparative study of the nematode communities on the southeastern Weddell Sea shelf and slope (Antarctica). Mar. Ecol. Prog. Ser. 1999, 181, 237–256. [Google Scholar]
- Vopel, K.; Thiel, H. Abyssal nematode assemblages in physically disturbed and adjacent sites of the eastern equatorial Pacific. Deep-Sea Res. II 2001, 48, 3795–3808. [Google Scholar]
- Lambshead, P.J.D.; Brown, C.J.; Ferrero, T.; Mitchell, N.; Hawkins, L.E.; Smith, C.R. Biodiversity of nematode assemblages from the Clarion-Clipperton Fracture Zone, an area of commercial mining interest. BMC Ecology 2003, 3, 1. [Google Scholar]
- Vanhove, S.; Vermeeren, H.; Vanreusel, A. Meiofauna towards the South Sandwich trench (750–6,300 m) focus on nematodes. Deep-Sea Res II 2004, 51, 1665–1687. [Google Scholar]
- Fonseca, G.; Decraemer, W.; Vanreusel, A. Taxonomy and species distribution of the genus Manganonema Bussau, 1993 (Nematoda: Monhysterida). Cah. Biol. Mar. 2006, 47, 189–203. [Google Scholar]
- Sebastian, S.; Raes, M.; De Mesel, I.; Vanreusel, A. Comparison of the nematode fauna from the Weddell Sea Abyssal Plain with two North Atlantic abyssal sites. Deep-Sea Res. II 2007, 54, 1727–1736. [Google Scholar]
- Danovaro, R.; Gambi, C.; Lampadariou, N.; Tselepides, A. Deep-sea nematode biodiversity in the Mediterranean basin: Testing for longitudinal, bathymetric and energetic gradients. Ecography 2008, 31, 231–244. [Google Scholar]
- Ingels, J.; Kiriakoulakis, K.; Wolff, G.A.; Vanreusel, A. Nematode diversity and its relation to the quantity and quality of sedimentary organic matter in the deep Nazaré Canyon, Western Iberian Margin. Deep-Sea Res. I 2009, 56, 1521–1539. [Google Scholar]
- Soltwedel, T.; Mokievsky, V.; Schewe, I.; Haseman, C. Yermak Plateau revisited: Spatial and temporal patterns of meiofaunal assemblages under permanent ice-coverage. Polar Biol. 2009, 32, 1159–1176. [Google Scholar]
- Van Gaever, S.; Olu, K.; Derycke, S.; Vanreusel, A. Metazoan meiofaunal communities at cold seeps along the Norwegian margin: Influence of habitat heterogeneity and evidence for connection with shallow-water habitats. Deep-Sea Res. I 2009, 56, 772–785. [Google Scholar]
- Miljutina, M.A.; Miljutin, D.M.; Mahatma, R.; Galéron, J. Deep-sea nematode assemblages of the Clarion-Clipperton Nodule Province (Tropical North-Eastern Pacific). Mar. Biodiv. 2010, 40, 1–15. [Google Scholar]
- Danovaro, R. Methods for the Study of Deep-Sea Sediments, Their Functioning and Biodiversity; Danovaro, R., Ed.; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar]
- Gerlach, S.A. Means of meiofaunal dispersal. Mikrofauna Meeresboden 1977, 61, 89–103. [Google Scholar]
- Brandt, A.; Gooday, A.J.; Branda, S.N.; Brix, S.; Brökeland, W.; Cedhagen, T.; Choudhury, M.; Cornelius, N.; Danis, B.; De Mesel, I.; Diaz, R.D.; Gillan, D.C.; Ebbe, B.; Howe, J.A.; Janussen, D.; Kaiser, S.; Linse, K.; Malyutina, M.; Pawlowski, J.; Raupach, M.; Vanreusel, A. First insights into the biodiversity and biogeography of the Southern Ocean deep sea. Nature 2007, 447, 307–312. [Google Scholar]
- Schüller, M.; Ebbe, B. Global distributional patterns of selected deep-sea Polychaeta (Annelida) from the Southern Ocean. Deep-Sea Res. II 2007, 54, 1737–1751. [Google Scholar]
- Scheltema, R.S. Adaptations for reproduction among deep-sea benthic molluscs: An appraisal for the existing evidence. In Reproduction, Larval Biology, and Recruitment of the Deep-Sea Benthos; Young, C.M., Eckelbarger, K.J., Eds.; Columbia University Press: New York, NY, USA, 1994; pp. 44–75. [Google Scholar]
- Pearse, J.S. Cold-water echinoderms break “Thorson's rule”. In Reproduction, Larval Biology, and Recruitment of the Deep-Sea Benthos; Young, C.M., Eckelbarger, K.J., Eds.; Columbia University Press: New York, NY, USA, 1994; pp. 26–43. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zeppilli, D.; Vanreusel, A.; Danovaro, R. Cosmopolitanism and Biogeography of the Genus Manganonema (Nematoda: Monhysterida) in the Deep Sea. Animals 2011, 1, 291-305. https://doi.org/10.3390/ani1030291
Zeppilli D, Vanreusel A, Danovaro R. Cosmopolitanism and Biogeography of the Genus Manganonema (Nematoda: Monhysterida) in the Deep Sea. Animals. 2011; 1(3):291-305. https://doi.org/10.3390/ani1030291
Chicago/Turabian StyleZeppilli, Daniela, Ann Vanreusel, and Roberto Danovaro. 2011. "Cosmopolitanism and Biogeography of the Genus Manganonema (Nematoda: Monhysterida) in the Deep Sea" Animals 1, no. 3: 291-305. https://doi.org/10.3390/ani1030291