Behavior and Characteristics of Sap-Feeding North Island kākā (Nestor meridionalis septentrionalis) in Wellington, New Zealand

Abstract
:Simple Summary
Abstract

1. Introduction
2. Methods
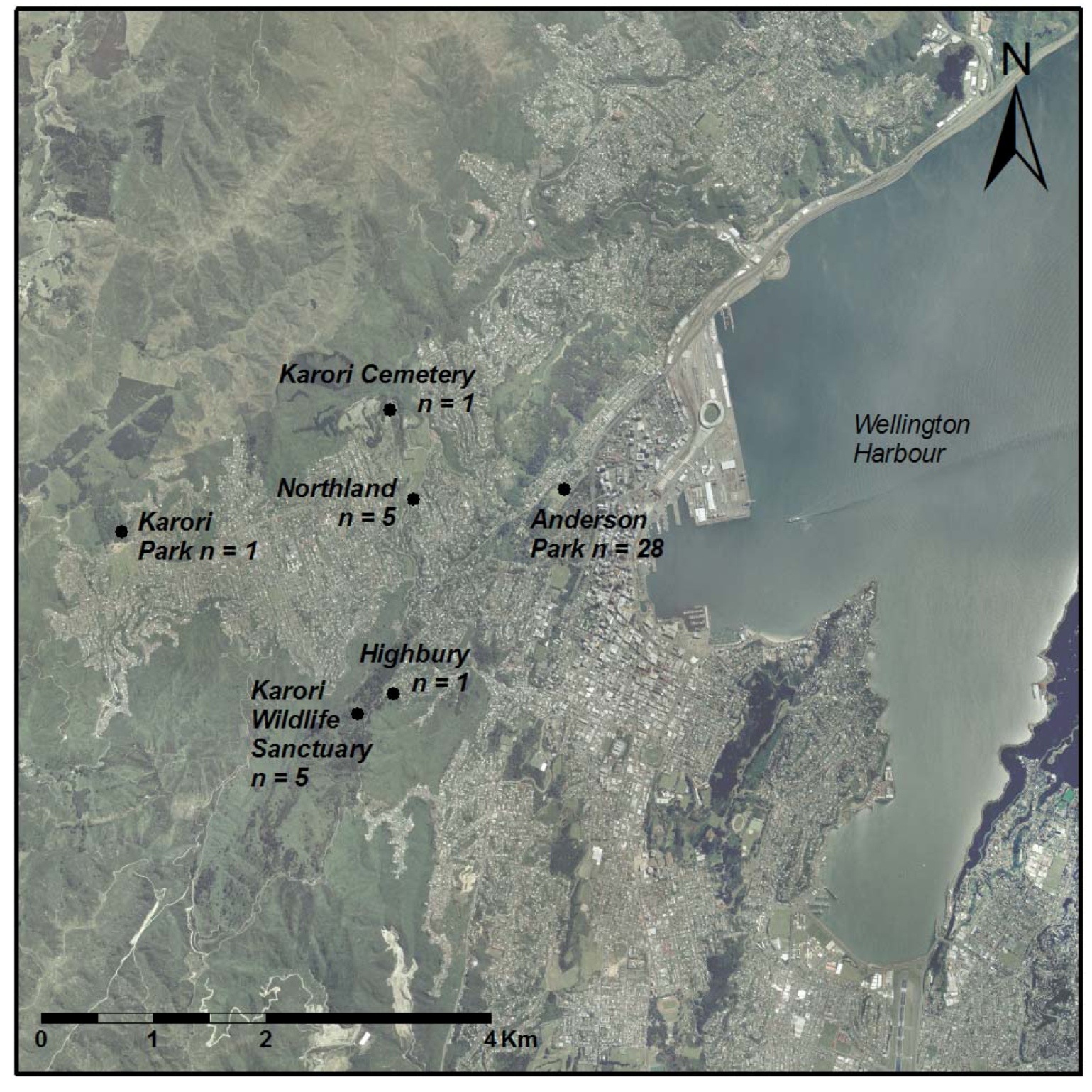
2.1. Study Population
2.2. Behavioral Observations

2.3. Analysis
3. Results
3.1. Sap Feeding Observations

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Leg band code 1 | Sap feeding observations | Season fledged | Sex | Method of sex determination 3 | ||
|---|---|---|---|---|---|---|
| No | Duration (mins) | Location 2 | ||||
| BG-V | 1 | 11 | WBG | 11/12 | M | Estimate from DFA |
| B-KB | 1 | 16 | BG, KWS | 08/09 | F | Breeding |
| BP-P | 1 | 8 | KWS | 02/03 | M | Breeding |
| GG-V | 6 | 122 | Northland, BG | 11/12 | F | Estimate from DFA |
| GW-V | 2 | 4 | WBG | 11/12 | M | Estimate from DFA |
| KO-G | 1 | 30 | KWS | 04/05 | F | Breeding |
| LW-O | 2 | 36 | WBG | 10/11 | M | Estimate from DFA |
| MG-V | 1 | 11 | WBG | 11/12 | F | Estimate from DFA |
| MP-V | 1 | 2 | WBG | 11/12 | F | Estimate from DFA |
| MR-O | 1 | 15 | WBG | 10/11 | F | Breeding |
| MW-O | 1 | 2 | WBG | 10/11 | F | Estimate from DFA |
| O-MY | 1 | 19 | WBG | 10/11 | M | Estimate from behavior |
| PB-V | 1 | 2 | WBG | 11/12 | M | Estimate from DFA |
| PL-O | 1 | 28 | KWS | 10/11 | F | Estimate from DFA |
| RP-V | 1 | 1 | WBG | 11/12 | M | Estimate from DFA |
| V-YW | 1 | 14 | WBG | 11/12 | F | Breeding |
| WB-W | 1 | 13 | KWS | 07/08 | F | Estimate from DFA |
| WK-O | 1 | 15 | Highbury | 10/11 | F | Breeding |
| W-OO | 1 | 52 | Karori Park | 08/09 | F? | Unknown |
| YK-O | 1 | 5 | WBG | 10/11 | M | Estimate from DFA |
| Y-RO | 2 | 53 | Northland | 08/09 | F | Breeding |
| Unbanded | 8 | 62 | WBG, Karori Cemetery | |||
| Unidentified | 4 | 34 | WBG | |||


3.2. Characteristics of Sap Feeding Individuals
4. Discussion
5. Conclusions
Acknowledgments
Conflict of Interest
References
- Organ, J.F.; Ellingwood, M.R. Wildlife stakeholder acceptance capacity for black bears, beavers, and other beasts in the east. Hum. Dimens. Wildl. 2000, 5, 63–75. [Google Scholar] [CrossRef]
- Urbanek, R.E.; Allen, K.R.; Nielsen, C.K. Urban and suburban deer management by state wildlife-conservation agencies. Wildlife Soc. B 2011, 35, 310–315. [Google Scholar] [CrossRef]
- Brockie, R.E.; Duncan, C. Long term trends in Wellington City bird counts: 1969–2006. Notornis 2012, 59, 1–6. [Google Scholar]
- Messmer, T.A. The emergence of human-wildlife conflict management: turning challenges into opportunities. Int. Biodeter. Biodegr. 2000, 45, 97–102. [Google Scholar] [CrossRef]
- Jonker, S.A.; Muth, R.M.; Organ, J.F.; Zwick, R.R.; Siemer, W.F. Experiences with beaver damage and attitudes of Massachusetts residents toward beaver. Wildlife Soc. B 2006, 34, 1009–1021. [Google Scholar] [CrossRef]
- Ditchkoff, S.S.; Saalfeld, S.T.; Gibson, C.J. Animal behavior in urban ecosystems: Modifications due to human-induced stress. Urban Ecosyst. 2006, 9, 5–12. [Google Scholar] [CrossRef]
- Francis, R.A.; Chadwick, M.A. What makes a species synurbic? Appl. Geogr. 2012, 32, 514–521. [Google Scholar] [CrossRef]
- Jones, D.N.; Thomas, L.K. Managing to live with Brisbane’s wildlife: magpies and the management of positive and negative interactions. Proc. R. Soc. Queensland 1998, 107, 45–49. [Google Scholar]
- Beaven, B.M. Sap feeding behaviour of North Island kaka (Nestor meridionalis septentrionalis, Lorenz 1896) in plantation forests. MSc Thesis, Waikato University, Hamilton, New Zealand, 1996. [Google Scholar]
- O’Donnell, C.F.J.; Dilks, P.J. Sap-feeding by the kaka (Nestor meridionalis) in South Westland New Zealand. Notornis 1989, 36, 65–71. [Google Scholar]
- Miskelly, C.; Empson, R.; Wright, K. Forest birds recolonising Wellington. Notornis 2005, 52, 21–26. [Google Scholar]
- Charles, K.E. Urban human-wildlife conflict: North Island kākā (Nestor meridionalis septentrionalis) in Wellington City. MSc Thesis, Victoria University of Wellington, Wellington, New Zealand, 2013. [Google Scholar]
- Charles, K.E. Tree damage in Wellington as a result of foraging for sap and bark-dwelling invertebrates by the North Island kaka (Nestor meridionalis septentrionalis). Notornis 2012, 59, 180–184. [Google Scholar]
- Berry, R. Reintroduction of kaka (Nestor meridionalis septentrionalis) to Mount Bruce Reserve, Wairarapa, New Zealand; Science for Conservation 89; Department of Conservation: Wellington, New Zealand, 1998. [Google Scholar]
- Moorhouse, R.J. The diet of the North Island kaka (Nestor meridionalis septentrionalis) on Kapiti Island. New Zeal. J. Ecol. 1997, 21, 141–152. [Google Scholar]
- Wilson, P.R.; Karl, B.J.; Toft, R.J.; Beggs, J.R.; Taylor, R.H. The role of introduced predators and competitors in the decline of kaka (Nestor meridionalis) populations in New Zealand. Biol. Conserv. 1998, 83, 175–185. [Google Scholar] [CrossRef]
- Innes, J. Kaka Damage to Exotic Plantation Trees at Whirinaki Forest—A Preliminary Report; Landcare Research Contract Report LC9495/36; Manaaki Whenua-Landcare Research: Hamilton, New Zealand, 1994. [Google Scholar]
- Karori Sanctuary Trust. Unpublished data, Wellington, New Zealand, 2013.
- Altmann, J. Observational study of behavior: Sampling methods. Behaviour 1974, 69, 227–267. [Google Scholar] [CrossRef]
- Miller, J.L. ; Karori Sanctuary Trust, Wellington, New Zealand. Personal communication, 2013.
- Greene, T.C.; Powlesland, R.G.; Dilks, P.J.; Moran, L. Research Summary and Options for Conservation of kaka (Nestor meridionalis); DOC Science Internal Series 178; Department of Conservation: Wellington, New Zealand, 2004. [Google Scholar]
- Chapman, A.; Bradford, M.G.; Hoskin, C.J. Sap suckers: A novel bird 'guild' in wet sclerophyll forests of tropical north Queensland. Emu 1999, 99, 69–72. [Google Scholar] [CrossRef]
- O’Donnell, C.F.J.; Dilks, P.J. Foods and foraging of forest birds in temperate rainforest, South Westland, New Zealand. New Zeal. J. Ecol. 1994, 18, 87–107. [Google Scholar]
- Temby, I.D. Urban wildlife issues in Australia. In Proceedings of the 4th International Symposium on Urban Wildlife Conservation, Tucson, AZ, USA, 1–5 May 2004; 2004; pp. 26–34. [Google Scholar]
- Temby, I.D. Managing Impacts of the Little Corella on the Fleurieu Peninsula. Unpublished Management Report, 2010. [Google Scholar]
- Temby, I.D. Reducing Cockatoo Damage to Trees, Fixtures, Houses, Sports Grounds and the Environment; Department of Sustainability and Environment: Melbourne, Australia, 2003. [Google Scholar]
- Conover, M. Resolving Human-Wildlife Conflicts. The Science of Wildlife Damage Management; CRC Press: Boca Raton, FL, USA, 2001. [Google Scholar]
- Dickman, A.J. Complexities of conflict: the importance of considering social factors for effectively resolving human-wildlife conflict. Anim. Conserv. 2010, 13, 458–466. [Google Scholar] [CrossRef]
- Patterson, M.E.; Montag, J.M.; Williams, D.R. The urbanization of wildlife management: Social science, conflict, and decision making. Urban For. Urban Green. 2003, 1, 171–183. [Google Scholar] [CrossRef]
- Bright, A.D.; Manfredo, M.J.; Fulton, D.C. Segmenting the public: an application of value orientations to wildlife planning in Colorado. Wildlife Soc. B 2000, 28, 218–226. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Charles, K.E.; Linklater, W.L. Behavior and Characteristics of Sap-Feeding North Island kākā (Nestor meridionalis septentrionalis) in Wellington, New Zealand. Animals 2013, 3, 830-842. https://doi.org/10.3390/ani3030830
Charles KE, Linklater WL. Behavior and Characteristics of Sap-Feeding North Island kākā (Nestor meridionalis septentrionalis) in Wellington, New Zealand. Animals. 2013; 3(3):830-842. https://doi.org/10.3390/ani3030830
Chicago/Turabian StyleCharles, Kerry E., and Wayne L. Linklater. 2013. "Behavior and Characteristics of Sap-Feeding North Island kākā (Nestor meridionalis septentrionalis) in Wellington, New Zealand" Animals 3, no. 3: 830-842. https://doi.org/10.3390/ani3030830