Single Nucleotide Polymorphisms in the HIRA Gene Affect Litter Size in Small Tail Han Sheep

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. WGS and Detection of Single Nucleotide Polymorphisms
2.2. Selection of Animals and DNA Preparation
2.3. Primer Design
2.4. Genotyping
2.5. RNA Extraction, cDNA Synthesis and qPCR
2.6. Statistics
3. Results
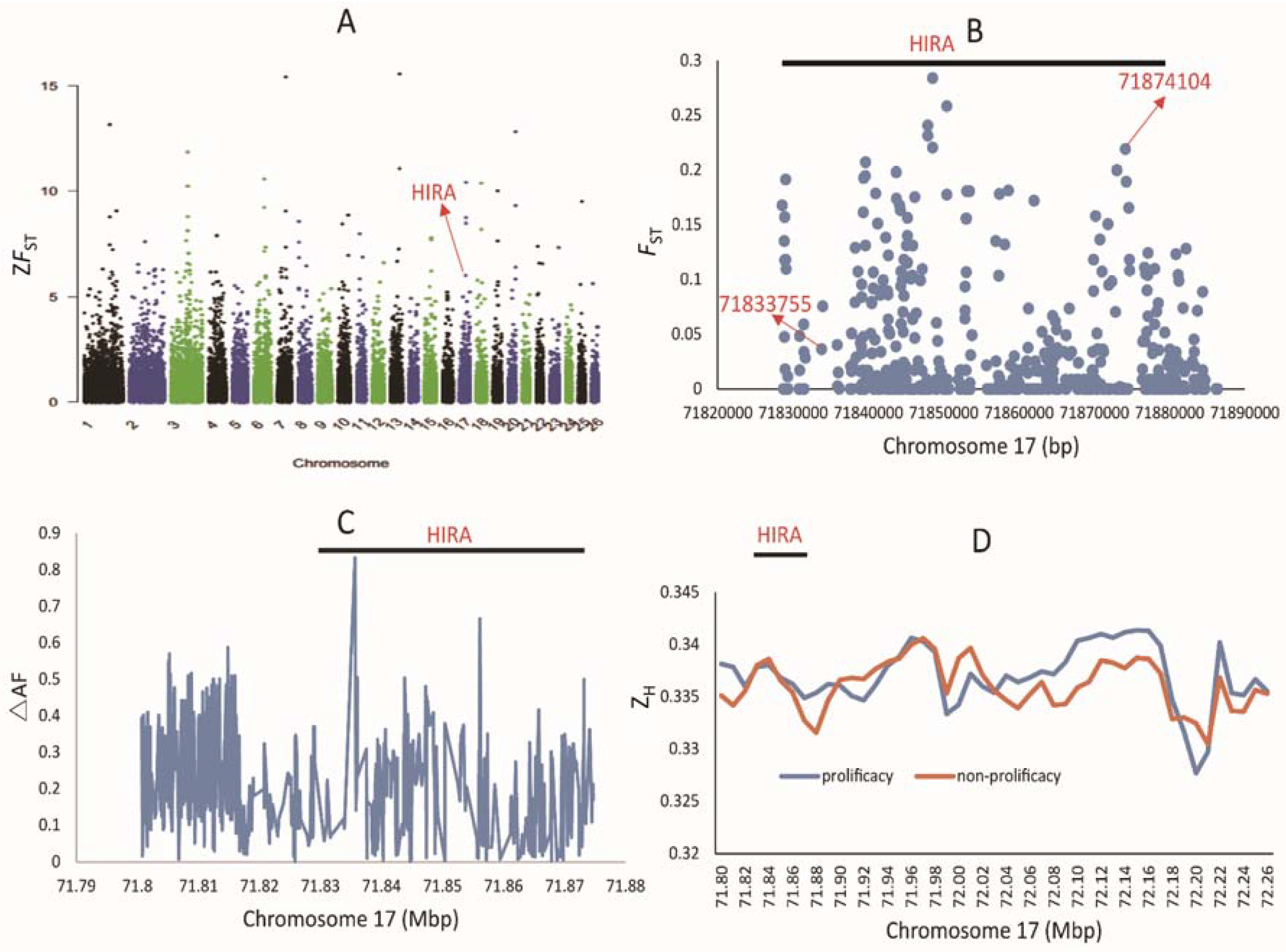
3.1. Genotype Analysis of Variants Located in the Significant Sweep Region
3.2. Population Genetic Analysis of Polymorphism in the HIRA Gene
3.3. Association Analysis of Two Polymorphisms with Litter Size in Small Tail Han Sheep
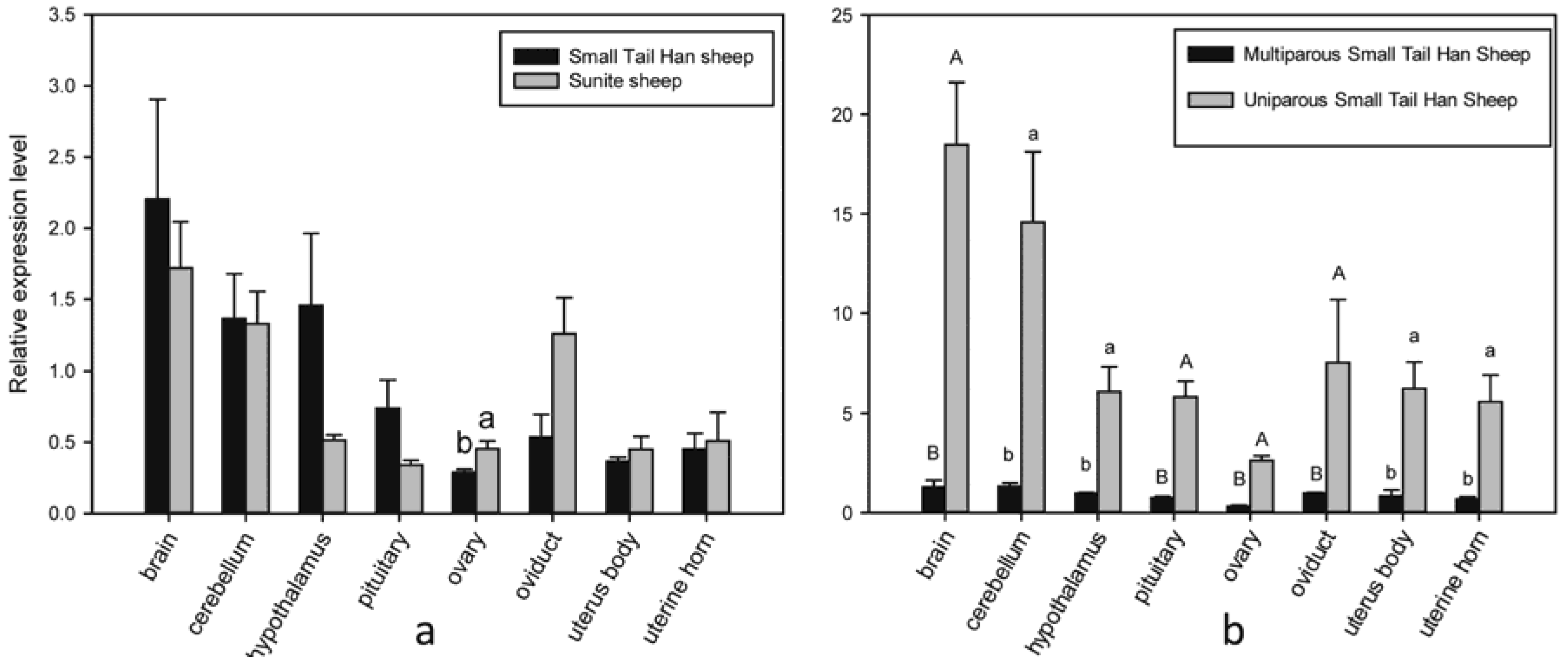
3.4. Expression of HIRA in Sheep with Different Fecundity and in Small Tail Han Sheep with Different Litter Size
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflict of Interest
References
- Bravo, S.; Larama, G.; Paz, E.; Inostroza, K.; Montaldo, H.H.; Sepúlveda, N. Polymorphism of the GDF9 gene associated with litter size in Araucana creole sheep. Anim. Genet. 2016, 47, 390–391. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, D.; Xiang, H.; Dun, W.; Brahi, D.O.; Yin, T.; Zhao, X. Mitochondrial DNA T7719G in tRNA-Lys gene affects litter size in Small-tailed Han sheep. J. Anim. Sci. Biotechnol. 2017, 8, 31. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.M.; Moore, S.S.; Stothard, P.; Liao, X.; Coltman, D.W. Harnessing cross-species alignment to discover SNPs and generate a draft genome sequence of a bighorn sheep (Ovis canadensis). BMC Genom. 2015, 16, 397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ai, H.; Fang, X.; Yang, B.; Huang, Z.; Chen, H.; Mao, L.; Zhang, F.; Zhang, L.; Cui, L.; He, W. Adaptation and possible ancient interspecies introgression in pigs identified by whole-genome sequencing. Nat. Genet. 2015, 47, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Malde, K.; Seliussen, B.B.; Quintela, M.; Dahle, G.; Besnier, F.; Skaug, H.J.; Øien, N.; Solvang, H.K.; Haug, T.; Skern-Mauritzen, R. Whole genome resequencing reveals diagnostic markers for investigating global migration and hybridization between minke whale species. BMC Genom. 2017, 18, 76. [Google Scholar] [CrossRef] [PubMed]
- Yano, K.; Yamamoto, E.; Aya, K.; Takeuchi, H.; Lo, P.; Li, H.; Yamasaki, M.; Yoshida, S.; Kitano, H.; Hirano, K. Genome-wide association study using whole-genome sequencing rapidly identifies new genes influencing agronomic traits in rice. Nat. Genet. 2017, 48, 927. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Wang, H.; Liu, G.; Wu, M.; Cao, J.; Liu, Z.; Liu, R.; Zhao, F.; Zhang, L.; Lu, J. Genome-wide analysis reveals population structure and selection in Chinese indigenous sheep breeds. BMC Genom. 2015, 16, 194. [Google Scholar] [CrossRef] [PubMed]
- Adam, S.; Polo, S.; Almouzni, G. Transcription Recovery after DNA Damage Requires Chromatin Priming by the H3.3 Histone Chaperone HIRA. Cell 2013, 155, 94–106. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.H.; Song, T.Y.; Chanhee, J.; Jinyoung, P.; Han-Young, L.; Song, I.; Hong, S.; Young, J.K.; Jaehoon, K.; Han, J.W. Differential regulation of the histone chaperone HIRA during muscle cell differentiation by a phosphorylation switch. Exp. Mol. Med. 2016, 48, e252. [Google Scholar] [CrossRef] [PubMed]
- Dilg, D.; Saleh, R.N.M.; Phelps, S.E.L.; Rose, Y.; Dupays, L.; Murphy, C.; Mohun, T.; Anderson, R.H.; Scambler, P.J.; Chapgier, A.L.A. HIRA is required for heart development and directly regulates Tnni2 and Tnnt3. PLoS ONE 2016, 11, e0161096. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.Y.; Guo, Q.H.; Du, X.Z.; Li, Z.; Qian, L.; Zeng, Q.H.; Wang, J.L.; Zhao, H.B.; Wang, Y.F. HIRA is essential for the development of gibel carp. Fish Physiol. Biochem. 2014, 40, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Raygallet, D.; Quivy, J.P.; Scamps, C.; Martini, E.M.; Lipinski, M.; Almouzni, G. HIRA is critical for a nucleosome assembly pathway independent of DNA synthesis. Mol. Cell 2002, 9, 1091–1100. [Google Scholar] [CrossRef]
- Gw, V.D.H.; Dieker, J.W.; Derijck, A.A.; Muller, S.; Berden, J.H.; Braat, D.D.; van, d.V.J.; De, B.P. Asymmetry in histone H3 variants and lysine methylation between paternal and maternal chromatin of the early mouse zygote. Mech. Dev. 2005, 122, 1008–1022. [Google Scholar] [CrossRef]
- Roberts, C.; Sutherland, H.F.; Farmer, H.; Kimber, W.; Halford, S.; Carey, A.; Brickman, J.M.; Wynshaw-Boris, A.; Scambler, P.J. Targeted Mutagenesis of the Hira Gene Results in Gastrulation Defects and Patterning Abnormalities of Mesoendodermal Derivatives Prior to Early Embryonic Lethality. Mol. Cell. Biol. 2002, 22, 2318–2328. [Google Scholar] [CrossRef] [PubMed]
- Nashun, B.; Hill, P.W.S.; Smallwood, S.A.; Festenstein, R.J.; Kelsey, G.; Hajkova, P. Continuous histone replacement by Hira is essential for normal transcriptional regulation and de novo DNA methylation during mouse oogenesis. Mol. Cell 2015, 60, 611–625. [Google Scholar] [CrossRef] [PubMed]
- Veselovska, L.; Smallwood, S.A.; Saadeh, H.; Stewart, K.R.; Krueger, F.; Maupetitméhouas, S.; Arnaud, P.; Tomizawa, S.; Andrews, S.; Kelsey, G. Deep sequencing and de novo assembly of the mouse oocyte transcriptome define the contribution of transcription to the DNA methylation landscape. Genome Biol. 2015, 16, 209. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.K.; Wang, Y.F. The function of histone chaperones during development. Hereditas 2010, 32, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Loppin, B.; Bonnefoy, E.; Anselme, C.; Laurençon, A.; Karr, T.L.; Couble, P. The histone H3.3 chaperone HIRA is essential for chromatin assembly in the male pronucleus. Nature 2005, 437, 1386–1390. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.H.; Zhao, Z.K.; Wang, Y.F.; Gui, J.F. Expression analysis of Hira mRNA and protein during oogenesis in gynogenetic and gonochoristic crucian carps. Acta Hydrobiol. Sin. 2010, 34, 611–617. [Google Scholar] [CrossRef]
- Paz, E.; Quiñones, J.; Bravo, S.; Montaldo, H.H.; Sepúlveda, N. Genotyping of BMPR1B, BMP15 and GDF9 genes in Chilean sheep breeds and association with prolificacy. Anim. Genet. 2015, 46, 98–99. [Google Scholar] [CrossRef] [PubMed]
- Ahlawat, S.; Sharma, R.; Roy, M.; Mandakmale, S.; Prakash, V.; Tantia, M.S. Genotyping of Novel SNPs in BMPR1B, BMP15, and GDF9 Genes for Association with Prolificacy in Seven Indian Goat Breeds. Anim. Biotechnol. 2016, 27, 199. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.Y.; Pan, Z.Y.; Wang, X.Y.; Hu, W.P.; Di, R.; Yao, Y.X.; Chu, M.X. Progress on major genes for high fecundity in ewes. Front. Agric. Sci. Eng. 2014, 1, 282. [Google Scholar] [CrossRef]
- Davis, G.H.; Balakrishnan, L.; Ross, I.K.; Wilson, T.; Galloway, S.M.; Lumsden, B.M.; Hanrahan, J.P.; Mullen, M.; Mao, X.Z.; Wang, G.L. Investigation of the Booroola (FecB) and Inverdale (FecX(I)) mutations in 21 prolific breeds and strains of sheep sampled in 13 countries. Anim. Reprod. Sci. 2006, 92, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Bodin, L.; Di, P.E.; Fabre, S.; Bontoux, M.; Monget, P.; Persani, L.; Mulsant, P. A novel mutation in the bone morphogenetic protein 15 gene causing defective protein secretion is associated with both increased ovulation rate and sterility in Lacaune sheep. Endocrinology 2007, 148, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Galloway, S.M.; Mcnatty, K.P.; Cambridge, L.M.; Laitinen, M.P.; Juengel, J.L.; Jokiranta, T.S.; Mclaren, R.J.; Luiro, K.; Dodds, K.G.; Montgomery, G.W. Mutations in an oocyte-derived growth factor gene (BMP15) cause increased ovulation rate and infertility in a dosage-sensitive manner. Nat. Genet. 2000, 25, 279–283. [Google Scholar] [CrossRef] [PubMed]
- Hanrahan, J.P.; Gregan, S.M.; Mulsant, P.; Mullen, M.; Davis, G.H.; Powell, R.; Galloway, S.M. Mutations in the genes for oocyte-derived growth factors GDF9 and BMP15 are associated with both increased ovulation rate and sterility in Cambridge and Belclare sheep (Ovis aries). Biol. Reprod. 2004, 70, 900–909. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Royo, A.; Jurado, J.J.; Smulders, J.P.; Martí, J.I.; Alabart, J.L.; Roche, A.; Fantova, E.; Bodin, L.; Mulsant, P.; Serrano, M. A deletion in the bone morphogenetic protein 15 gene causes sterility and increased prolificacy in Rasa Aragonesa sheep. Anim. Genet. 2008, 39, 294–297. [Google Scholar] [CrossRef] [PubMed]
- Monteagudo, L.V.; Ponz, R.; Tejedor, M.T.; Lavicla, A.; Sierra, I. A 17bp deletion in the Bone Morphogenetic Protein 15 (BMP15) gene is associated to increased prolificacy in the Rasa Aragonesa sheep breed. Anim. Reprod. Sci. 2009, 110, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Demars, J.; Fabre, S.; Sarry, J.; Rossetti, R.; Gilbert, H.; Persani, L.; Tosser-Klopp, G.; Mulsant, P.; Nowak, Z.; Drobik, W. Genome-Wide Association Studies Identify Two Novel BMP15 Mutations Responsible for an Atypical Hyperprolificacy Phenotype in Sheep. PLoS Genet. 2013, 9, e1003482. [Google Scholar] [CrossRef] [PubMed]
- Gorlov, I.F.; Kolosov, Y.A.; Shirokova, N.V.; Getmantseva, L.V.; Slozhenkina, M.I.; Mosolova, N.I.; Bakoev, N.F.; Leonova, M.A.; Kolosov, A.Y.; Zlobina, E.Y. GDF9 gene polymorphism and its association with litter size in two Russian sheep breeds. Rend. Lincei Sci. Fis. Nat. 2018, 29, 61–66. [Google Scholar] [CrossRef]
- Pan, Z.Y.; Li, S.D.; Liu, Q.Y.; Wang, Z.; Zhou, Z.K.; Di, R.; Miao, B.P.; Hu, W.P.; Wang, X.Y.; Hu, X.X.; Xu, Z. Whole-genome sequences of 89 Chinese sheep suggest role of RXFP2 in the development of unique horn phenotype as response to semi-feralization. Gigascience 2018. [Google Scholar] [CrossRef] [PubMed]
- Alachiotis, N.; Pavlidis, P. Scalable linkage-disequilibrium-based selective sweep detection: A performance guide. Gigascience 2016, 5, 7. [Google Scholar] [CrossRef] [PubMed]
- Munch, K.; Nam, K.; Schierup, M.H.; Mailund, T. Selective Sweeps across Twenty Millions Years of Primate Evolution. Mol. Biol. Evol. 2016, 33, 3065–3074. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Yuan, A.; Shriner, D.; Tekolaayele, F.; Zhou, J.; Bentley, A.R.; Zhou, Y.; Wang, C.; Newport, M.J.; Adeyemo, A. An Improved Fst Estimator. PLoS ONE 2015, 10, e0135368. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed]
- Chu, M.X.; Liu, Z.H.; Jiao, C.L.; He, Y.Q.; Fang, L.; Ye, S.C.; Chen, G.H.; Wang, J.Y. Mutations in BMPR-IB and BMP-15 genes are associated with litter size in Small Tailed Han sheep (Ovis aries). J. Anim. Sci. 2007, 85, 598–603. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Maniatis, T.E.; Fritsch, E.F. A laboratory Manual. Anal Biochem 1983, 186, 182–183. [Google Scholar]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed]
- Ortega, M.S.; Denicol, A.C.; Cole, J.B.; Null, D.J.; Hansen, P.J. Use of single nucleotide polymorphisms in candidate genes associated with daughter pregnancy rate for prediction of genetic merit for reproduction in Holstein cows. Anim. Genet. 2016, 47, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Szenker, E.; Raygallet, D.; Almouzni, G. The double face of the histone variant H3.3. Cell Res. 2011, 21, 421–434. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Tyler, J.K. Nucleosome disassembly during human non-homologous end joining followed by concerted HIRA- and CAF-1-dependent reassembly. Elife 2016, 5, e15129. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Zhang, Z. O-linked N-acetylglucosamine transferase (OGT) interacts with the histone chaperone HIRA complex and regulates nucleosome assembly and cellular senescence. Proc. Natl. Acad. Sci. USA 2016, 113, E3213. [Google Scholar] [CrossRef] [PubMed]
- Herndon, M.K.; Nilson, J.H. Maximal expression of Foxl2 in pituitary gonadotropes requires ovarian hormones. PLoS ONE 2015, 10, e0126527. [Google Scholar] [CrossRef] [PubMed]
- Camille, M.L.; Maguire, J. GABAergic regulation of the HPA and HPG axes and the impact of stress on reproductive function. J. Steroid Biochem. 2015, 160, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Pchelintsev, N.A.; Mcbryan, T.; Rai, T.S.; Tuyn, J.V.; Raygallet, D.; Almouzni, G.; Adams, P.D. Placing the HIRA histone chaperone complex in the chromatin landscape. Cell Rep. 2013, 3, 1012–1019. [Google Scholar] [CrossRef] [PubMed]
- Bonnefoy, E.; Orsi, G.A.; Couble, P.; Loppin, B. The Essential Role of Drosophila HIRA for De Novo Assembly of Paternal Chromatin at Fertilization. PLoS Genet. 2007, 3, 1991–2006. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.J.; Koh, F.M.; Wong, P.; Conti, M.; Ramalho-Santos, M. Hira-mediated H3.3 incorporation is required for DNA replication and ribosomal RNA transcription in the mouse zygote. Dev. Cell 2014, 30, 268–279. [Google Scholar] [CrossRef] [PubMed]
- Orsi, G.A.; Algazeery, A.; Meyer, R.E.; Capri, M.; Sapeytriomphe, L.M.; Horard, B.; Gruffat, H.; Couble, P.; Aïtahmed, O.; Loppin, B. Drosophila Yemanuclein and HIRA Cooperate for de novo Assembly of H3.3-Containing Nucleosomes in the Male Pronucleus. PLoS Genet. 2013, 9, e1003285. [Google Scholar] [CrossRef] [PubMed]
- Clift, D.; Schuh, M. Restarting life: Fertilization and the transition from meiosis to mitosis. Nat. Rev. Mol. Cell Biol. 2013, 14, 549–562. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Breed | Number | Type | District |
|---|---|---|---|
| Small Tail Han sheep | 380 | multiparous | Farmers home, Yuncheng, Shandong Province, China |
| Tan sheep | 80 | uniparous | Yanchi, Ningxia Hui Autonomous Region, China |
| Sunite sheep | 100 | uniparous | Wulatezhongqi, Bayannaoer, Inner Mongolia Autonomous Region, China |
| Suffolk sheep | 39 | uniparous | Beijing Aoxin Stud Farm Co. Ltd. located in Shunyi District, Beijing, China |
| Dorper sheep | 30 | uniparous | Beijing Aoxin Stud Farm Co. Ltd. located in Shunyi District, Beijing, China |
| Prairie Tibetan sheep | 131 | uniparous | Dangxiong, Tibet Autonomous Region, China |
| Primer Name | Primer Sequence | Product Size | Usage |
|---|---|---|---|
| HIRA-1-F | 5′-ACGTTGGATGAAACGAAACCAGAGCTCTCC-3′ | 133 bp | Polymerase Chain Reaction (PCR) for g.71874104G>A |
| HIRA-1-R | 5′-ACGTTGGATGTGTGCAGAGGGTCTGATAAC-3′ | ||
| HIRA-1-E | 5′-GGGCCCGGCAACCGAGTTAGTC-3′ | Extension reaction | |
| HIRA-2-F | 5′-ACGTTGGATGAGCCCGTGGGAAGACACTGT-3′ | 126 bp | PCR for g.71833755T>C |
| HIRA-2-R | 5′-ACGTTGGATGAAGGCTTTCTGACAGTCCTC-3′ | ||
| HIRA-2-E | 5′-CAGACCCGAGTAGAGCTCCCCT-3′ | Extension reaction | |
| HIRA-3-F | 5′-CTGAGCGAGGAGGAGAAGAG-3′ | 108 bp | qPCR |
| HIRA-3-R | 5′-CATCTCGGGGTTCTCGATGA-3′ | ||
| β-actin-F | 5′-GCTGTATTCCCCTCCATCGT-3′ | 97 bp | qPCR |
| β-actin-R | 5′-GGATACCTCTCTTGCTCTGG-3′ |
| Locus | Breed | Genotype Frequency | Allele Frequency | PIC | HE | NE | Chi-Square Test (p-Value) | |||
|---|---|---|---|---|---|---|---|---|---|---|
| g.71874104G>A | GG | GA | AA | G | A | |||||
| Small Tail Han sheep | 0.11 (42) | 0.45 (167) | 0.44 (166) | 0.33 | 0.67 | 0.35 | 0.45 | 1.80 | 1.00 | |
| Tan sheep | 0.16 (13) | 0.46 (37) | 0.38 (30) | 0.39 | 0.61 | 0.36 | 0.48 | 1.91 | 0.78 | |
| Sunite sheep | 0.06 (6) | 0.36 (36) | 0.58 (57) | 0.24 | 0.76 | 0.30 | 0.37 | 1.58 | 0.92 | |
| Suffolk sheep | 0.11 (4) | 0.34 (13) | 0.55 (21) | 0.28 | 0.72 | 0.32 | 0.40 | 1.67 | 0.37 | |
| Dorper sheep | 0.18 (5) | 0.32 (9) | 0.50 (14) | 0.34 | 0.66 | 0.35 | 0.45 | 1.81 | 0.13 | |
| Prairie Tibetan sheep | 0.10 (13) | 0.37 (48) | 0.53 (69) | 0.28 | 0.72 | 0.32 | 0.41 | 1.69 | 0.29 | |
| g.71833755T>C | TT | CT | CC | T | C | |||||
| Small Tail Han sheep | 0.58 (215) | 0.34 (126) | 0.09 (32) | 0.75 | 0.25 | 0.30 | 0.37 | 1.60 | 0.03 | |
| Tan sheep | 0.64 (51) | 0.28 (22) | 0.08 (7) | 0.78 | 0.22 | 0.29 | 0.35 | 1.54 | 0.06 | |
| Sunite sheep | 0.72 (69) | 0.22 (21) | 0.06 (6) | 0.83 | 0.17 | 0.24 | 0.28 | 1.40 | 0.02 | |
| Suffolk sheep | 0.92 (36) | 0.08 (3) | 0 (0) | 0.96 | 0.04 | 0.07 | 0.07 | 1.08 | 0.80 | |
| Dorper sheep | 1 (30) | 0 (0) | 0 (0) | 1.00 | 0.00 | 0.00 | 0.00 | 1.00 | ||
| Prairie Tibetan sheep | 0.74 (95) | 0.17 (22) | 0.09 (12) | 0.82 | 0.18 | 0.25 | 0.29 | 1.41 | 0.00 | |
| Locus | Genotype | Litter Size of the First Parity | Litter Size of the Second Parity | Litter Size of the Third Parity | Average Litter Size |
|---|---|---|---|---|---|
| g.71874104G>A | GG | 1.97 ± 0.13 (35) | 2.24 ± 0.13 (41) | 2.83 ± 0.29 b (7) | 2.25 ± 0.13 (83) |
| GA | 2.19 ± 0.06 (154) | 2.39 ± 0.07 (163) | 3.09 ± 0.13 a (64) | 2.51 ± 0.05 (381) | |
| AA | 2.11 ± 0.06 (141) | 2.11 ± 0.07 (159) | 2.71 ± 0.13 b (54) | 2.34 ± 0.05 (354) | |
| g.71833755T>C | TT | 2.16 ± 0.06 (193) | 2.13 ± 0.06 b (211) | 2.77 ± 0.11 b (78) | 2.35 ± 0.04 b (466) |
| CT | 2.13 ± 0.07 (115) | 2.45 ± 0.08 a (124) | 3.16 ± 0.17 a (37) | 2.56 ± 0.06 a (277) | |
| CC | 2.04 ± 0.15 (27) | 2.28 ± 0.15 ab (32) | 2.67 ± 0.29 b (12) | 2.29 ± 0.12 ab (68) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, M.; Pan, Z.; Cao, X.; Guo, X.; He, X.; Sun, Q.; Di, R.; Hu, W.; Wang, X.; Zhang, X.; et al. Single Nucleotide Polymorphisms in the HIRA Gene Affect Litter Size in Small Tail Han Sheep. Animals 2018, 8, 71. https://doi.org/10.3390/ani8050071
Zhou M, Pan Z, Cao X, Guo X, He X, Sun Q, Di R, Hu W, Wang X, Zhang X, et al. Single Nucleotide Polymorphisms in the HIRA Gene Affect Litter Size in Small Tail Han Sheep. Animals. 2018; 8(5):71. https://doi.org/10.3390/ani8050071
Chicago/Turabian StyleZhou, Mei, Zhangyuan Pan, Xiaohan Cao, Xiaofei Guo, Xiaoyun He, Qing Sun, Ran Di, Wenping Hu, Xiangyu Wang, Xiaosheng Zhang, and et al. 2018. "Single Nucleotide Polymorphisms in the HIRA Gene Affect Litter Size in Small Tail Han Sheep" Animals 8, no. 5: 71. https://doi.org/10.3390/ani8050071