Effect of Curcumin on Growth Performance, Inflammation, Insulin level, and Lipid Metabolism in Weaned Piglets with IUGR

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Curcumin Preparation
2.2. Animal Experiment Design
2.3. Sample Collection
2.4. Serum Tumor Necrosis α, Interleukin 1β, and Interleukin 6
2.5. Liver Aminotransferase Activities
2.6. Serum Insulin and Glucose
2.7. Serum and Liver Biochemistry Parameters
2.8. Hepatic Gene Expression Assays
2.9. Statistical Analysis
3. Results
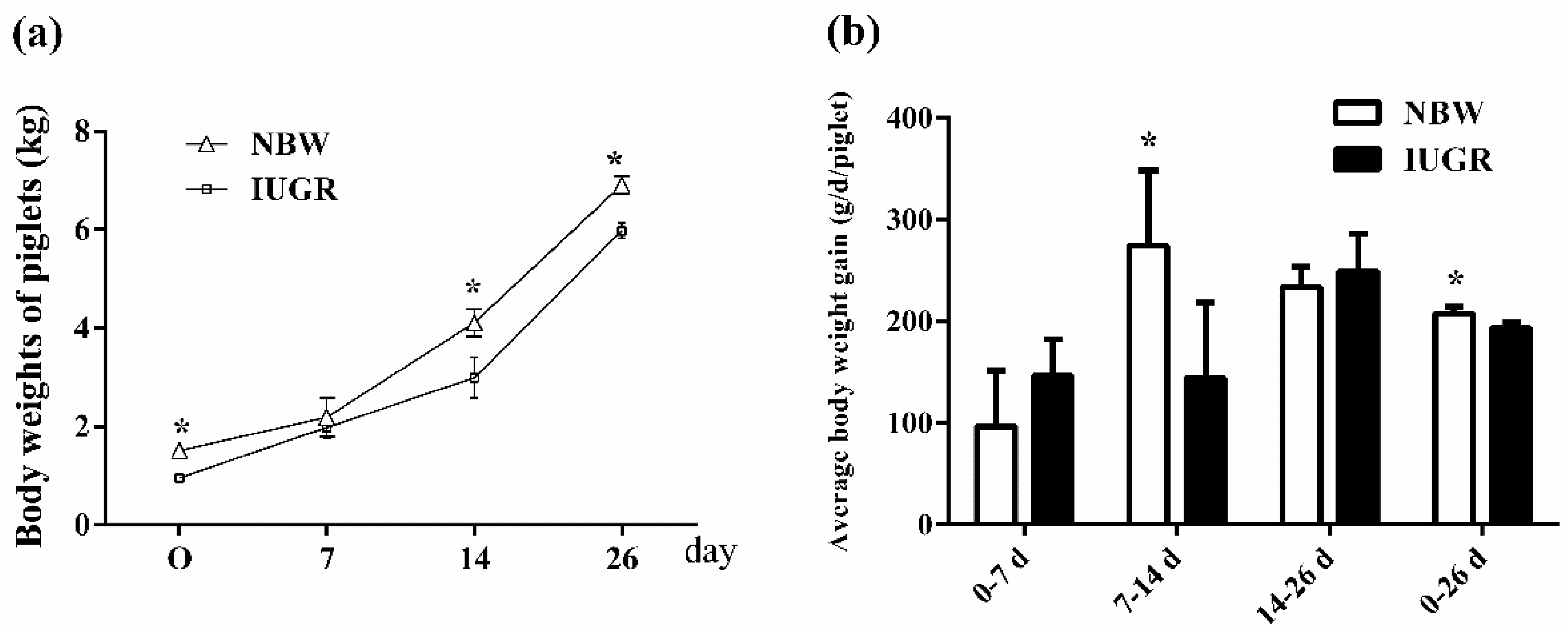
3.1. Growth Performance
3.2. Organ Index
3.3. Levels of Serum TNF-α, IL-1β and IL-6
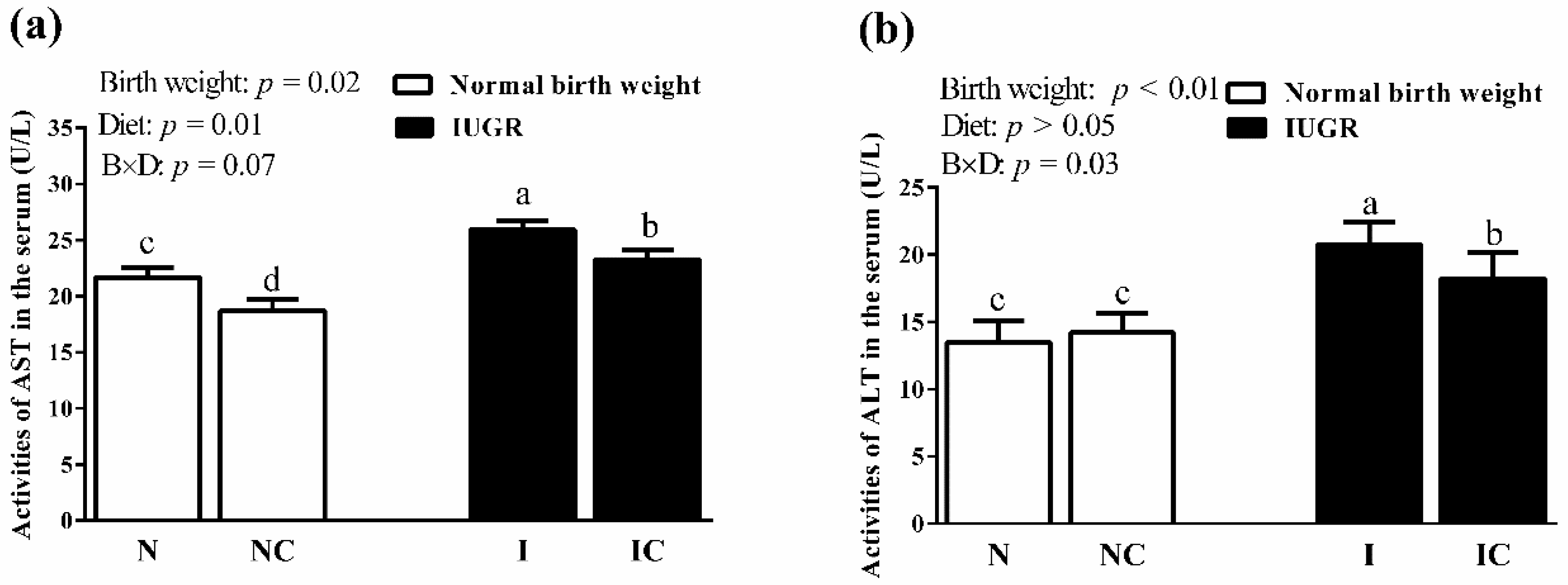
3.4. Activities of Serum AST and ALT
3.5. Serum Biochemistry Parameters
3.6. Hepatic Biochemistry Parameters
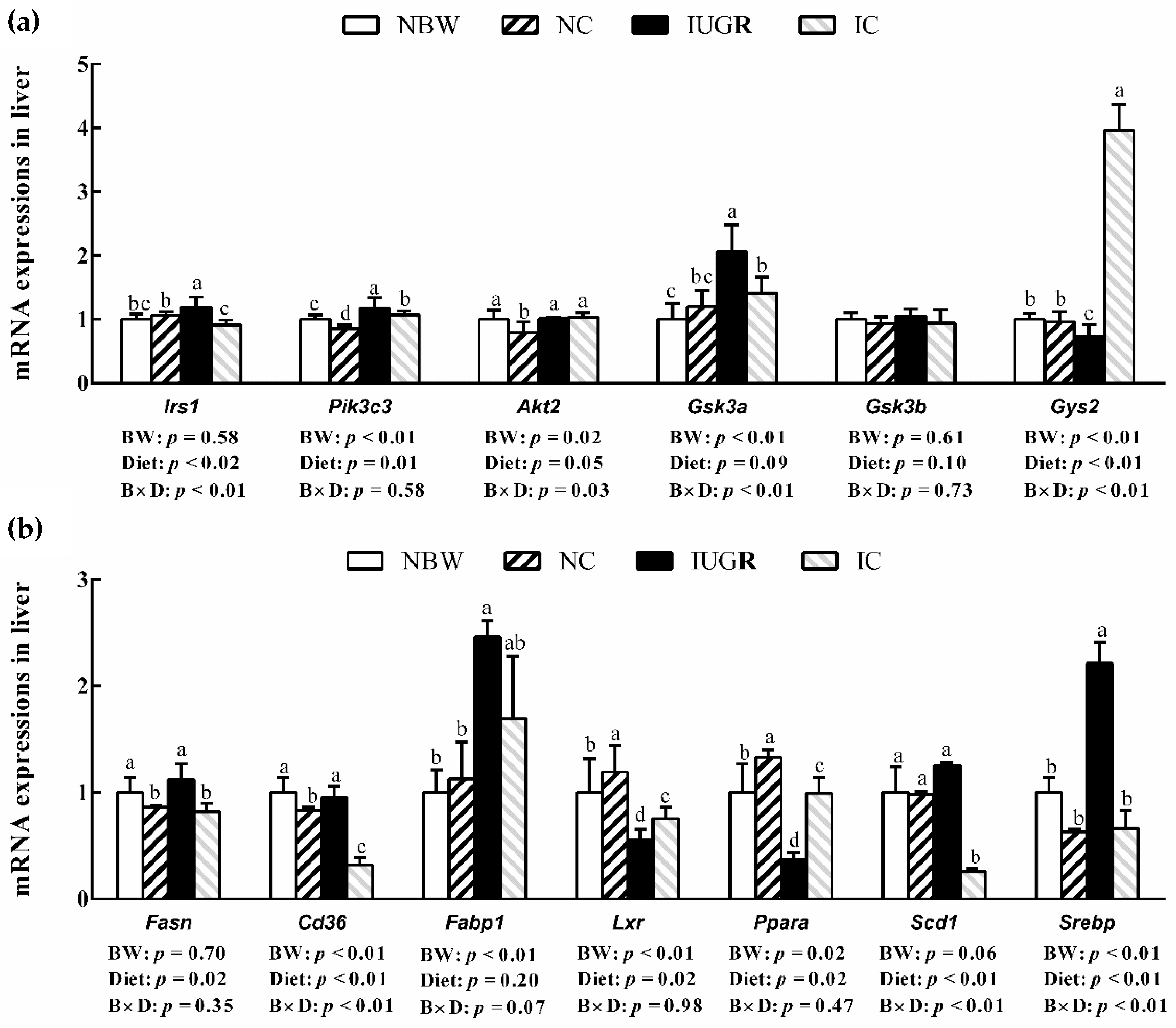
3.7. mRNA Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wu, G.; Bazer, F.W.; Walace, J.M.; Spencer, T.E. Board-invited review: Intrauterine growth retardation: Implications for the animal sciences. J. Anim. Sci. 2006, 84, 2316–2337. [Google Scholar] [CrossRef] [PubMed]
- Garite, T.J.; Clark, R.; Thorp, J.A. Intrauterine growth restriction increases morbidity and mortality among premature neonates. Am. J. Obstet. Gynecol. 2004, 191, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Barker, D.J.; Gluckman, P.D.; Godfrey, K.M.; Harding, J.E.; Owens, J.A.; Robinson, J.S. Fetal nutrition and cardiovascular disease in adult life. Lancet 1993, 341, 938–941. [Google Scholar] [CrossRef]
- Dong, L.; Zhong, X.; Zhang, L.; Kong, L.; Kong, Y.; Kou, T.; Wang, T. Impaired intestinal mucosal immunity is associated with the imbalance of T lymphocyte sub-populations in intrauterine growth-restricted neonatal piglets. Immunobiology 2015, 220, 775–781. [Google Scholar] [CrossRef]
- He, J.T.; Dong, L.; Xu, W.; Bai, K.W.; Lu, C.H.; Wu, Y.N.; Huang, Q.; Zhang, L.L.; Wang, T. Dietary tributyrin supplementation attenuates insulin resistance and abnormal lipid metabolism in suckling piglets with intrauterine growth retardation. PLoS ONE 2015, 10, e0136848. [Google Scholar] [CrossRef]
- Levy-Marchal, C.; Jaquet, D. Long-term metabolic consequences of being born small for gestational age. Pediatri. Diabetes 2004, 5, 147–153. [Google Scholar] [CrossRef]
- Widdowson, E.M. Intra-uterine growth retardation in the pig. I. Organ size and cellular development at birth and after growth to maturity. Biol. Neonate 1971, 19, 329–340. [Google Scholar] [CrossRef]
- Wang, J.; Chen, L.; Li, D.; Yin, Y.; Wang, X.; Li, P.; Dangott, L.J.; Hu, W.; Wu, G. Intrauterine growth restriction affects the proteomes of the small intestine, liver, and skeletal muscle in newborn pigs. J. Nutr. 2008, 138, 60–66. [Google Scholar] [CrossRef] [Green Version]
- Miłobȩdzka, J.; Kostanecki, S.V.; Lampe, V. Zur Kenntnis des Curcumins. Eur. J. Inorg. Chem. 1910, 43, 2163–2170. [Google Scholar]
- Xun, W.; Shi, L.; Zhou, H.; Hou, G.; Cao, T.; Zhao, C. Effects of curcumin on growth performance, jejunal mucosal membrane integrity, morphology and immune status in weaned piglets challenged with enterotoxigenic Escherichia coli. Int. Immunopharmacol. 2015, 27, 46–52. [Google Scholar] [CrossRef]
- Wei, S.; Xu, H.; Xia, D.; Zhao, R. Curcumin attenuates the effects of transport stress on serum cortisol concentration, hippocampal NO production, and BDNF expression in the pig. Swine Prod. 2010, 39, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Park, S.; Jeon, B.S.; Jang, W.S.; Lee, S.J.; Son, Y.; Rhim, K.J.; Lee, S.I.; Lee, S.S. Therapeutic effect of topical application of curcumin in treatment of radiation burns in a mini-pig model. J. Vet. Sci. 2016, 17, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Anderson, L.L.; Parker, R.O. Distribution and development of embryos in the pig. J. Reprod Fertil. 1976, 46, 363–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Research Council. Nutrient Requirements of Swine, 11th ed.; National Academy Press: Washington, DC, USA, 2012. [Google Scholar]
- Dong, L.; Zhong, X.; Ahmad, H.; Li, W.; Wang, Y.; Zhang, L.; Wang, T. Intrauterine growth restriction impairs small intestinal mucosal immunity in neonatal piglets. J. Histochem. Cytochem. 2014, 62, 510–518. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, L.; Zhou, G.; Liao, Z.; Ahmad, H.; Liu, W.; Wang, T. Dietary L-arginine supplementation improves the intestinal development through increasing mucosal Akt and mammalian target of rapamycin signals in intra-uterine growth retarded piglets. Br. J. Nutr. 2012, 108, 1371–1381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Xu, L.; Zhang, L.; Ying, Z.; Su, W.; Wang, T. Curcumin attenuates D-galactosamine/lipopolysaccharide-induced liver injury and mitochondrial dysfunction in mice. J. Nutr. 2014, 144, 1211–1218. [Google Scholar] [CrossRef] [Green Version]
- Bucolo, G.; David, H. Quantitative determination of serum triglycerides by the use of enzymes. Clin. Chem. 1973, 19, 476–482. [Google Scholar]
- Yokode, M.; Hammer, R.E.; Ishibashi, S.; Brown, M.S.; Goldstein, J.L. Diet-induced hypercholesterolemia in mice: Prevention by overexpression of LDL receptors. Science 1990, 250, 1273–1275. [Google Scholar] [CrossRef]
- Effect of Curcumin on Growth Performance, Inflammation, Insulin Level, and Lipid Metabolism in Weaned Piglets with IUGR. Available online: http://doi.org/10.5281/zenodo.3543510 (accessed on 15 November 2019).
- Enser, M. 2-The chemistry, biochemistry and nutritional importance of animal fats. Fats Anm. Nutr. 1984, 23–51. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Dong, L.; Zhong, X.; He, J.; Zhang, L.; Bai, K.; Xu, W.; Wang, T.; Huang, X. Supplementation of tributyrin improves the growth and intestinal digestive and barrier functions in intrauterine growth-restricted piglets. Clin. Nutr. 2015, 399–407. [Google Scholar] [CrossRef]
- He, J.; Niu, Y.; Wang, F.; Wang, C.; Cui, T.; Bai, K.; Zhang, J.; Zhong, X.; Zhang, L.; Wang, T. Dietary curcumin supplementation attenuates inflammation, hepatic injury and oxidative damage in a rat model of intra-uterine growth retardation. Br. J. Nutr. 2018, 120, 537–548. [Google Scholar] [CrossRef] [Green Version]
- Desai, M.; Gayle, D.; Babu, J.; Ross, M.G. Programmed obesity in intrauterine growth-restricted newborns: Modulation by newborn nutrition. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, R91–R96. [Google Scholar] [CrossRef] [PubMed]
- Desai, M.; Crowther, N.J.; Lucas, A.; Hales, C.N. Organ-selective growth in the offspring of protein-restricted mothers. Br. J. Nutr. 1996, 76, 591–603. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.F.; Hu, Z.P.; Lu, C.H.; Yang, M.X.; Zhang, L.L.; Wang, T. Dietary curcumin supplementation protects against heat-stress-impaired growth performance of broilers possibly through a mitochondrial pathway. J. Anim. Sci. 2015, 93, 1656–1665. [Google Scholar] [CrossRef] [PubMed]
- Caprau, D.M.; O’Grady, S.P.; Callaway, C.W.; Hale, M.A.; Ke, X.; Yu, X.; Men, P.; Mcknight, R.A.; Lane, R.H. 1A-4 IUGR alters IGF1 receptor gene expression, DNA methylation and histones acetylation in the brain. Early Hum. Dev. 2007, 83, S47. [Google Scholar] [CrossRef]
- Krajewski, P.; Sieroszewski, P.; Karowicz-Bilinska, A.; Kmiecik, M.; Chudzik, A.; Strzalko-Gloskowska, B.; Kwiatkowska, M.; Pokrzywnicka, M.; Wyka, K.; Chlapinski, J.; et al. Assessment of interleukin-6, interleukin-8 and interleukin-18 count in the serum of IUGR newborns. J. Matern. Fetal. Neonatal. Med. 2014, 27, 1142–1145. [Google Scholar] [CrossRef] [PubMed]
- Al-Azemi, M.; Raghupathy, R.; Azizieh, F. Pro-inflammatory and anti-inflammatory cytokine profiles in fetal growth restriction. Clin. Exp. Obstet. Gynecol. 2017, 44, 98–103. [Google Scholar] [PubMed]
- Dacaj, R.; Izetbegovic, S.; Stojkanovic, G.; Dreshaj, S. Elevated liver enzymes in cases of preeclampsia and intrauterine growth restriction. Med. Arch. 2016, 70, 44–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Shang, Y.; Li, M.; Han, X.; Wang, J.; Wang, J. Curcumin ameliorates asthmatic airway inflammation by activating nuclear factor-E2-related factor 2/haem oxygenase (HO)-1 signalling pathway. Clin. Exp. Pharmacol. Physiol. 2015, 42, 520–529. [Google Scholar] [CrossRef]
- Liu, C.; Lin, G.; Wang, X.Q.; Wang, T.J.; Wu, G.Y.; Li, D.F.; Wang, J.J. Intrauterine growth restriction alters the hepatic proteome in fetal pigs. J. Nutr. Biochem. 2013, 24, 954–959. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Li, W.; Zhang, H.; Zhang, L.; Wang, T. Effects of high dietary concentrations of choline on longissimus dorsi muscle glycolysis in intrauterine growth retardation pigs. J. Nanjing Agric. Univ. 2015, 38, 324–329. [Google Scholar]
- Fritsche, L.; Weigert, C.; Haring, H.U.; Lehmann, R. How insulin receptor substrate proteins regulate the metabolic capacity of the liver-implications for health and disease. Curr. Med. Chem. 2008, 15, 1316–1329. [Google Scholar] [CrossRef] [PubMed]
- Pearce, N.J.; Arch, J.R.; Clapham, J.C.; Coghlan, M.P.; Corcoran, S.L.; Lister, C.A.; Llano, A.; Moore, G.B.; Murphy, G.J.; Smith, S.A. Development of glucose intolerance in male transgenic mice overexpressing human glycogen synthase kinase-3 beta on a muscle-specific promoter. Metabolism 2004, 53, 1322–1330. [Google Scholar] [CrossRef] [PubMed]
- Morrison, J.L.; Duffield, J.A.; Muhlhausler, B.S.; Gentili, S.; Mcmillen, I.C. Fetal growth restriction, catch-up growth and the early origins of insulin resistance and visceral obesity. Pediatr. Nephrol. 2010, 25, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Patricia, V.; Elisabeth, R.; Bing, L.; Nir, B.; Rebecca, S. Hepatic insulin resistance precedes the development of diabetes in a model of intrauterine growth retardation. Diabetes 2004, 53, 2617–2622. [Google Scholar]
- Jang, E.M.; Choi, M.S.; Jung, U.J.; Kim, M.J.; Kim, H.J.; Jeon, S.M.; Shin, S.K.; Seong, C.N.; Lee, M.K. Beneficial effects of curcumin on hyperlipidemia and insulin resistance in high-fat-fed hamsters. Metabolism 2008, 57, 1576–1583. [Google Scholar] [CrossRef]
- Na, L.X.; Zhang, Y.L.; Li, Y.; Liu, L.Y.; Li, R.; Kong, T.; Sun, C.H. Curcumin improves insulin resistance in skeletal muscle of rats. Nutr. Metab. Cardiovasc. Dis. 2011, 21, 526–533. [Google Scholar] [CrossRef]
- Bustanji, Y.; Taha, M.O.; Almasri, I.M.; Al-Ghussein, M.A.; Mohammad, M.K.; Alkhatib, H.S. Inhibition of glycogen synthase kinase by curcumin: Investigation by simulated molecular docking and subsequent in vitro/in vivo evaluation. J. Enzyme. Inhib. Med. Chem. 2009, 24, 771–778. [Google Scholar] [CrossRef]
- Browning, J.D.; Horton, J.D. Molecular mediators of hepatic steatosis and liver injury. J. Clin. Investig. 2004, 114, 147–152. [Google Scholar] [CrossRef] [Green Version]
- Musso, G.; Gambino, R.; Cassader, M. Recent insights into hepatic lipid metabolism in non-alcoholic fatty liver disease (NAFLD). Prog. Lipid. Res. 2009, 48, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Asai, A.; Miyazawa, T. Dietary curcuminoids prevent high-fat diet-induced lipid accumulation in rat liver and epididymal adipose tissue. J. Nutr. 2001, 131, 2932–2935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magee, T.R.; Han, G.; Cherian, B.; Khorram, O.; Ross, M.G.; Desai, M. Down-regulation of transcription factor peroxisome proliferator-activated receptor in programmed hepatic lipid dysregulation and inflammation in intrauterine growth-restricted offspring. Am. J. Obstet. Gynecol. 2008, 199, 271.e1–271.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aml, E.; Salter, A.M.; Sculley, D.V.; Langley-Evans, S.C.; Bennett, A.J. Prenatal exposure to a low-protein diet programs disordered regulation of lipid metabolism in the aging rat. Am. J. Physiol. Endocrinol. Metab. 2007, 292, 1702–1714. [Google Scholar]
- Lee, S.; You, Y.A.; Kwon, E.J.; Jung, S.C.; Jo, I.; Kim, Y.J. Maternal food restriction during pregnancy and lactation adversely affect hepatic growth and lipid metabolism in three-week-old rat offspring. Int. J. Mol. Sci. 2016, 17, 2115. [Google Scholar] [CrossRef] [Green Version]
- Kang, O.H.; Kim, S.B.; Seo, Y.S.; Joung, D.K.; Mun, S.H.; Choi, J.G.; Lee, Y.M.; Kang, D.G.; Lee, H.S.; Kwon, D.Y. Curcumin decreases oleic acid-induced lipid accumulation via AMPK phosphorylation in hepatocarcinoma cells. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 2578–2586. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Ingredients | Ratio (%) | Nutrient Composition | |
|---|---|---|---|
| Corn | 57.70 | Digestible energy (MJ/kg) | 14.04 |
| Soybean meal | 12.50 | Crude protein (%) | 18.31 |
| Expanded corn | 8.00 | Lysine (%) | 1.31 |
| Full-fat soybean | 8.00 | Methionine (%) | 0.40 |
| Fermented soybean meal | 4.00 | Methionine + Cystine (%) | 0.70 |
| Whey powder | 3.00 | Threonine (%) | 0.80 |
| Fish meal (crude protein 67%) | 3.00 | Calcium (%) | 0.85 |
| Dicalcium phosphate | 1.80 | Total phosphorus (%) | 0.72 |
| Limestone | 0.50 | ||
| L-lysine (78%) | 0.30 | ||
| L-threonine | 0.10 | ||
| DL-methionine | 0.08 | ||
| Wheat middling and reddog | 0.02 | ||
| Premix * | 1 | ||
| Total | 100 |
| Genes | Accession No. | Primer, 5′-3′ |
|---|---|---|
| Irs1 | NM_001244489.1 | GCCACGGGAGAATGGGTTTA GTCGCACACAGTTTCAGCAG |
| Pik3c3 | XM_021093598.1 | GCATGTTTCGCCAAGGGATG CTGCTTGTTCTGCCAGGAGT |
| Akt2 | XM_011526619.1 | CTGACCTGCTGTCCGCAAAT GACACGCTGTCACCTAGCTT |
| Gsk3a | XM_021093339.1 | CAGTGCAAAGCAGTTGGTCC GGTGTAATCGGTGGCTCCAA |
| Gsk3b | NM_001128443 | CGAGACACACCTGCACTCTT CCGGCATTAGTATCTGAGGCT |
| Gys2 | NM_001195511.1 | TGGGAATTCTGTGGGAAGCC TAGGTGCACTTGATGCAGGG |
| Fasn | NM_213839.1 | TGATGCCCAAGTGACTGACC CAGCATGTTTCCGTTTGCCA |
| Cd36 | NM_001044622.1 | TAGGAATCCCACTGCCTCAC TGCTTCAAGTGCTGGGTCA |
| Fabp1 | AY960623.1 | GAGTAGCCTCATTGCCACCAT TGCACGATTTCCGATGTCCC |
| Lxrα | AB254405.1 | CCCTCTCTCGCTCAGCTCC GGAGCCCTGGACATTACCAA |
| Ppara | NM_001044526.1 | CTGGCCACATCCATCCAACA ATAACGGGCTTTCCAGGTCG |
| Scd1 | NM_213781.1 | TGCTGATCCCCACAATTCCC CTTTGACGGCTGGGTGTTTG |
| Srebp | AY338729.1 | GCTACCGCTCCTCCATCAAT CTGCTTGAGCTTCTGGTTGC |
| Actb | XM_003124280.4 | CAGTCGGTTGGATGGAGCAT AGGCAGGGACTTCCTGTAAC |
| Items | Experiment Groups | p-Value | |||||
|---|---|---|---|---|---|---|---|
| NBW | NC | IUGR | IC | B | D | B × D | |
| IBW (kg) | 6.96 ± 0.13 a | 6.97 ± 0.34 a | 6.02 ± 0.17 b | 6.08 ± 0.05 b | <0.01 | 0.60 | 0.68 |
| FBW (kg) | 13.01 ± 0.69 a | 12.55 ± 0.44 a | 10.48 ± 0.47 c | 11.38 ± 0.95 b | <0.01 | 0.36 | 0.01 |
| ADG (g/d/piglet) | 251.85 ± 27.78 a | 257.91 ± 16.29 a | 183.59 ± 19.35 b | 204.04 ± 31.08 b | <0.01 | 0.14 | 0.41 |
| ADFI (g/d/piglet) | 326.16 ± 13.78 a | 278.81 ± 20.74 a,b | 231.75 ± 17.84 b | 295.78 ± 17.95 a | 0.07 | 0.69 | <0.01 |
| FCR | 1.33 ± 0.11 a | 1.12 ± 0.05 b | 1.36 ± 0.06 a | 1.39 ± 0.12 a | <0.01 | 0.01 | <0.01 |
| Items | Experiment Groups | p-Value | |||||
|---|---|---|---|---|---|---|---|
| NBW | NC | IUGR | IC | B | D | B × D | |
| Liver weight (g) | 379.71 ± 22.42 a | 356.02 ± 29.56 a,b | 313.95 ± 23.26 c | 338.17 ± 26.40 b,c | <0.01 | 0.98 | 0.01 |
| Spleen weight (g) | 26.44 ± 6.87 | 25.55 ± 5.23 | 21.28 ± 2.26 | 27.46 ± 7.02 | 0.42 | 0.20 | 0.09 |
| Kidney weight (g) | 58.81 ± 2.93 a | 58.85 ± 5.16 a | 49.24 ± 4.86 b | 56.24 ± 2.91 a | <0.01 | 0.02 | 0.02 |
| Pancreas weight (g) | 31.28 ± 4.64 a | 24.96 ± 2.31 b | 22.74 ± 1.44 b | 22.99 ± 1.99 b | <0.01 | <0.01 | <0.01 |
| LRW (g/kg BW) | 30.06 ± 2.11 a | 27.89 ± 1.44 b | 28.67 ± 1.65 a,b | 29.44 ± 2.42 a,b | 0.91 | 0.31 | 0.04 |
| SRW (g/kg BW) | 2.15 ± 0.18 b | 2.04 ± 0.15 b | 2.06 ± 0.26 b | 2.46 ± 0.14 a | 0.02 | 0.04 | <0.01 |
| KRW (g/kg BW) | 4.77 ± 0.38 b | 5.01 ± 0.24 a | 4.78 ± 0.25 b | 5.39 ± 0.30 a | 0.08 | <0.01 | 0.09 |
| PRW (g/kg BW) | 2.37 ± 0.37 a | 2.14 ± 0.22 a,b | 2.08 ± 0.20 b | 2.01 ± 0.15 b | 0.03 | 0.09 | 0.35 |
| Items | Experiment Groups | p-Value | |||||
|---|---|---|---|---|---|---|---|
| NBW | NC | IUGR | IC | B | D | B × D | |
| TNF-α (pg/mL) | 250.60 ± 24.10 b | 214.26 ± 51.03 c | 287.40 ± 29.56 a | 244.58 ± 19.13 b | 0.02 | 0.01 | 0.81 |
| IL-1β (pg/mL) | 747.52 ± 101.98 b | 674.78 ± 106.50 c | 856.67 ± 128.92 a | 743.36 ± 57.37 b | 0.05 | 0.04 | 0.63 |
| IL-6 (pg/mL) | 1162.44 ± 281.32 b | 899.94 ± 139.67 b | 1306.12 ± 400.78 a | 1208.26 ± 133.94 b | 0.05 | 0.11 | 0.45 |
| Items | Experiment Groups | p-Value | |||||
|---|---|---|---|---|---|---|---|
| NBW | NC | IUGR | IC | B | D | B × D | |
| Insulin (mU/L) | 19.44 ± 3.01 b | 16.77 ± 4.14 c | 22.71 ± 3.80 a | 19.64 ± 3.67 b | 0.03 | 0.04 | 0.88 |
| Glucose (mmol/L) | 5.14 ± 0.89 a | 4.53 ± 0.52 b | 5.75 ± 0.42 a | 4.27 ± 0.58 b | 0.44 | <0.01 | 0.06 |
| HOMA-IR | 5.66 ± 1.06 b | 2.98 ± 0.51 d | 6.40 ± 1.06 a | 4.80 ± 1.55 c | <0.01 | <0.01 | 0.18 |
| TC (mmol/L) | 1.96 ± 0.36 a | 1.92 ± 0.08 b | 2.17 ± 0.04 a | 1.89 ± 0.09 b | 0.19 | 0.03 | 0.09 |
| TG (mmol/L) | 0.59 ± 0.13 | 0.55 ± 0.16 | 0.53 ± 0.17 | 0.50 ± 0.07 | 0.28 | 0.51 | 0.86 |
| HDL-C (mmol/L) | 1.23 ± 0.09 b | 1.55 ± 0.24 a | 1.59 ± 0.20 a | 1.32 ± 0.29 b | 0.42 | 0.75 | <0.01 |
| LDL-C (mmol/L) | 0.86 ± 0.15 a | 0.71 ± 0.12 b | 0.67 ± 0.10 b | 0.85 ± 0.16 a | 0.63 | 0.71 | <0.01 |
| NEFA (μmol/L) | 230.00 ± 21.46 a | 143.11 ± 34.15 c | 141.96 ± 18.10 c | 191.30 ± 21.03 b | 0.03 | 0.04 | <0.01 |
| Items | Experiment Groups | p-Value | |||||
|---|---|---|---|---|---|---|---|
| NBW | NC | IUGR | IC | B | D | B × D | |
| Glycogen (mg/g tissue) | 121.44 ± 20.58 c | 154.80 ± 22.24 b | 39.93 ± 18.72 d | 189.76 ± 37.56 a | 0.02 | <0.01 | <0.01 |
| Pyruvate (μmol/mg prot) | 0.04 ± 0.01 a | 0.03 ± 0.00 b | 0.04 ± 0.00 a | 0.03 ± 0.00 b | 0.31 | <0.01 | 0.45 |
| Lactate (mmol/g prot) | 0.24 ± 0.04 b | 0.38 ± 0.11 a | 0.43 ± 0.07 a | 0.37 ± 0.10 a | 0.01 | 0.17 | <0.01 |
| PK (U/g prot) | 17.17 ± 1.72 b | 18.63 ± 0.49 b | 22.27 ± 0.61 a | 18.59 ± 1.59 b | <0.01 | 0.02 | <0.01 |
| LDH (U/g prot) | 171.55 ± 23.53 a | 135.56 ± 13.76 b | 130.63 ± 8.68 b | 120.13 ± 14.11 b | <0.01 | <0.01 | 0.03 |
| TC (μmol/g prot) | 81.55 ± 19.51 b | 86.28 ± 13.92 b | 126.65 ± 22.83 a | 84.13 ± 12.52 b | <0.01 | 0.01 | <0.01 |
| TG (μmol/g prot) | 80.32 ± 6.44 b | 74.22 ± 5.34 c | 89.93 ± 9.12 a | 82.37 ± 8.93 b | <0.01 | 0.02 | 0.79 |
| NEFA (μmol/g prot) | 36.06 ± 7.93 b | 24.85 ± 3.03 c | 50.19 ± 11.18 a | 22.09 ± 3.40 c | 0.03 | <0.01 | <0.01 |
| LPL (U/mg prot) | 0.16 ± 0.02 b | 0.23 ± 0.04 a | 0.08 ± 0.01 c | 0.10 ± 0.03 c | <0.01 | <0.01 | 0.03 |
| HL (U/mg prot) | 0.11 ± 0.02 a | 0.07 ± 0.01 b | 0.09 ± 0.01 b | 0.12 ± 0.03 a | 0.23 | 0.54 | <0.01 |
| TL (U/mg prot) | 0.28 ± 0.03 b | 0.30 ± 0.04 a | 0.17 ± 0.01 d | 0.22 ± 0.04 c | <0.01 | <0.01 | 0.25 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, Y.; He, J.; Zhao, Y.; Shen, M.; Zhang, L.; Zhong, X.; Wang, C.; Wang, T. Effect of Curcumin on Growth Performance, Inflammation, Insulin level, and Lipid Metabolism in Weaned Piglets with IUGR. Animals 2019, 9, 1098. https://doi.org/10.3390/ani9121098
Niu Y, He J, Zhao Y, Shen M, Zhang L, Zhong X, Wang C, Wang T. Effect of Curcumin on Growth Performance, Inflammation, Insulin level, and Lipid Metabolism in Weaned Piglets with IUGR. Animals. 2019; 9(12):1098. https://doi.org/10.3390/ani9121098
Chicago/Turabian StyleNiu, Yu, Jintian He, Yongwei Zhao, Mingming Shen, Lili Zhang, Xiang Zhong, Chao Wang, and Tian Wang. 2019. "Effect of Curcumin on Growth Performance, Inflammation, Insulin level, and Lipid Metabolism in Weaned Piglets with IUGR" Animals 9, no. 12: 1098. https://doi.org/10.3390/ani9121098