Squalicorax Chips a Tooth: A Consequence of Feeding-Related Behavior from the Lowermost Navesink Formation (Late Cretaceous: Campanian-Maastrichtian) of Monmouth County, New Jersey, USA

Abstract
:1. Introduction
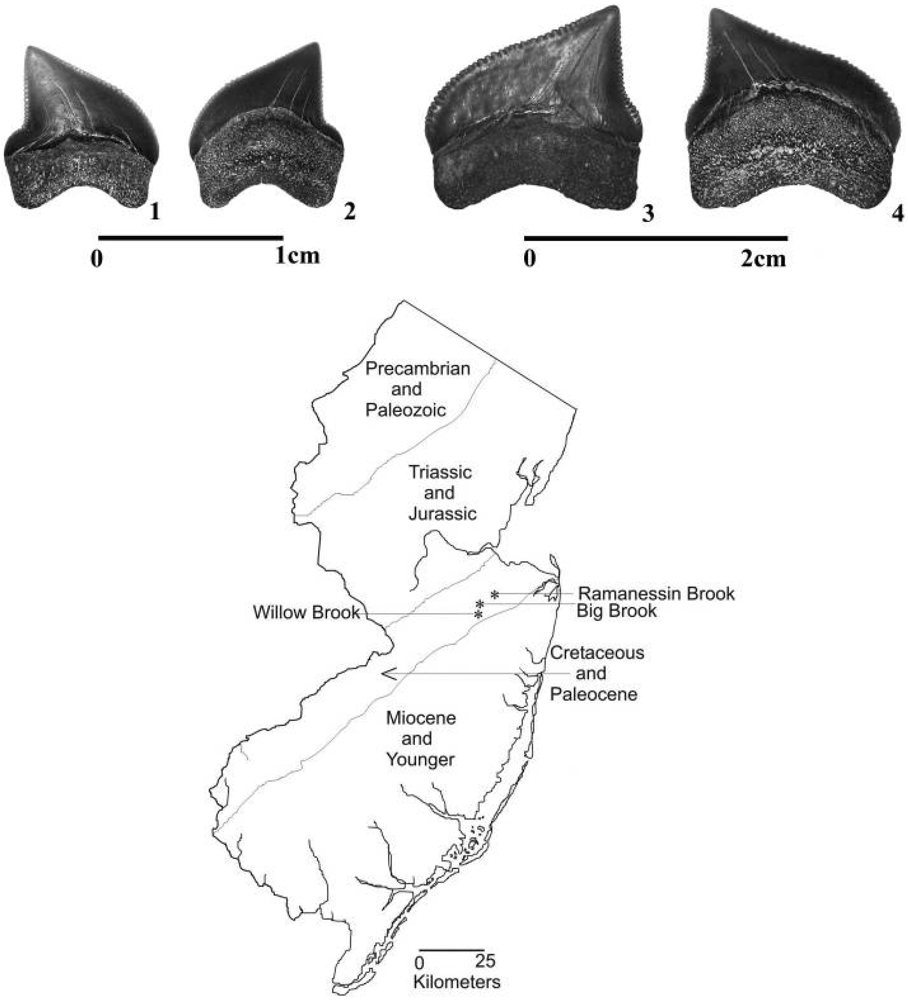
2. Teeth of Squalicorax

3. Materials and Methods
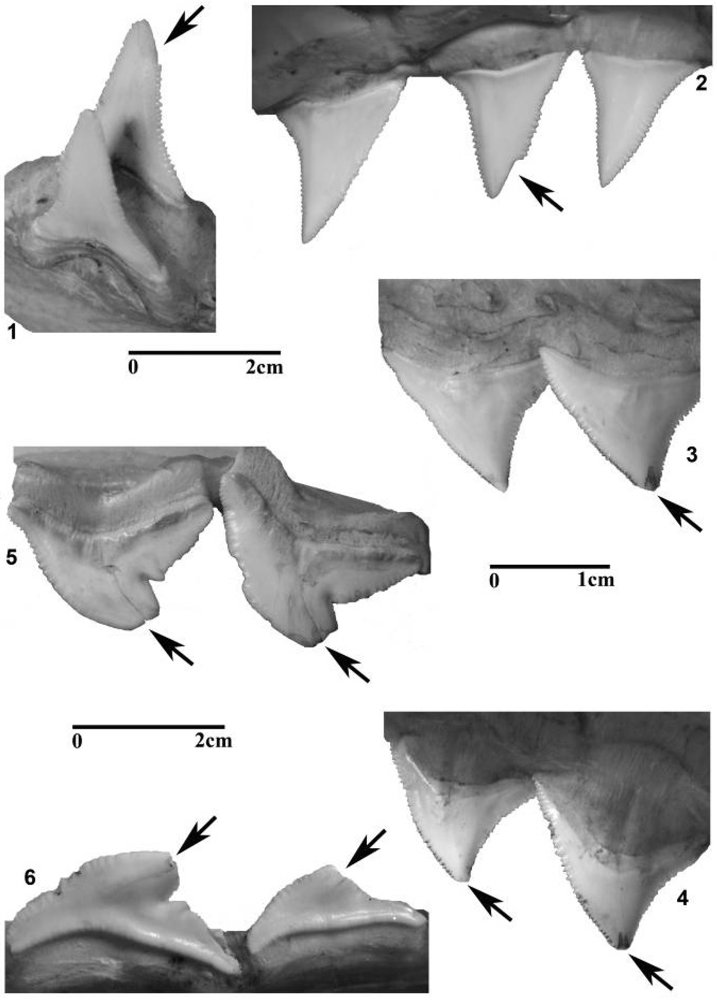
3.1. Modern Jaws and Teeth


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Functional Teeth | Pre-Functional Teeth | Indeterminate or Broken During Collecting | Total Number of Teeth Studied | |||
|---|---|---|---|---|---|---|---|
| Undamaged | Feeding Damage | Taphonomic Damage | Undamaged | Pathologically Deformed1 | |||
| Squalicorax kaupi | 187 | 36 | 16 | 1448 | 4 | 237 | 1928 |
| Squalicorax pristodontus | 13 | 20 | 30 | 326 | 1 | 87 | 477 |
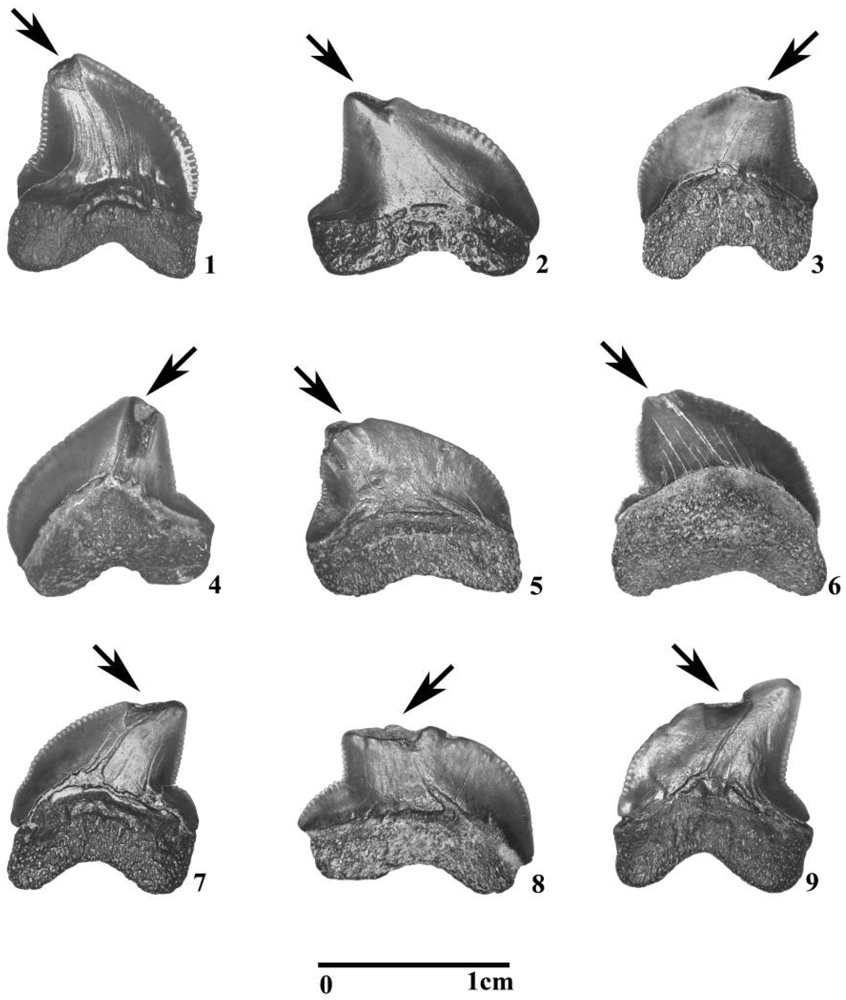
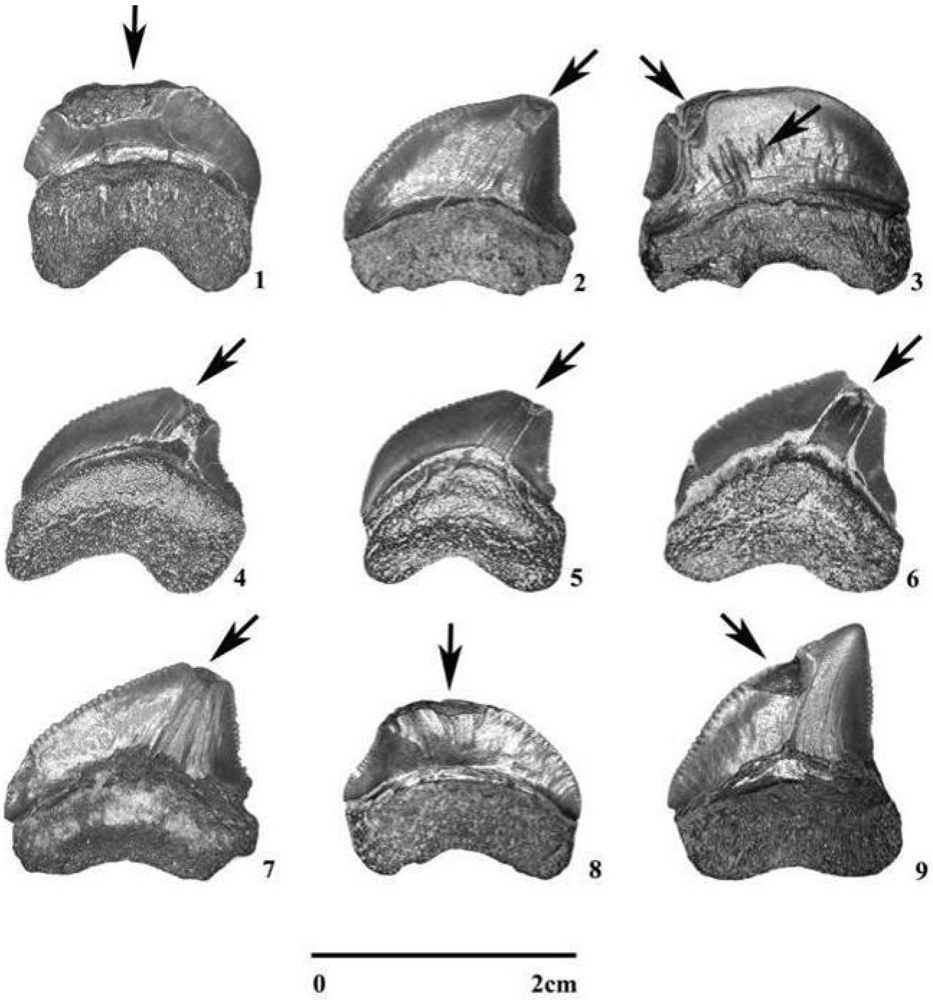
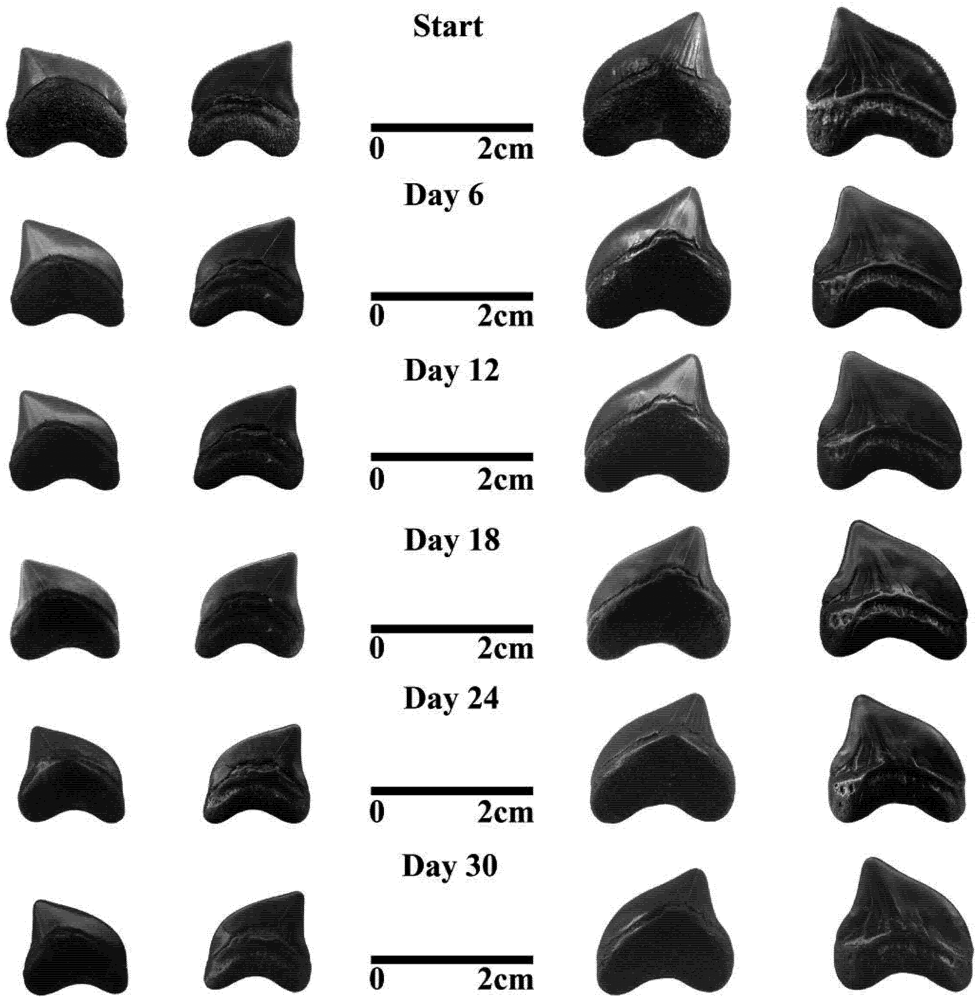
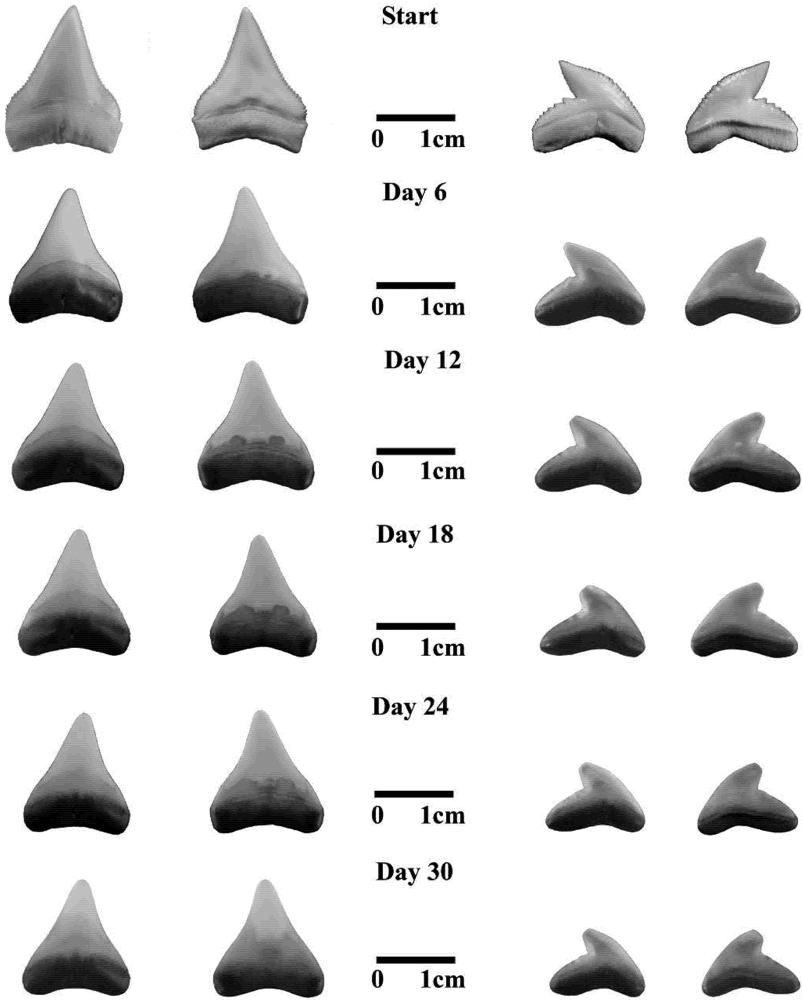
3.2. Fossil Teeth and Tumbler Experiments
4. Results
4.1. Sample Populations


4.2. Tumbling Experiments

| SPECIES | Mass Before Testing1 (gm) | Mass After Testing2 (gm) | % Loss3 | |
|---|---|---|---|---|
| FOSSIL | Squalicoras kaupi | 0.41 | 0.35 | 14.6 |
| Squalicorax pristodontus | 1.27 | 1.09 | 14.2 | |
| MODERN | Carcharodon carcharias | 2.49 | 1.43 | 42.6 |
| Carcharhinus leucas | 0.77 | 0.39 | 49.4 | |
| Galeocerdo cuvier | 0.29 | 0.15 | 43.3 | |



5. Discussion
5.1. Taphonomic Versus Feeding-Related Tooth Damage

5.2. Modern Versus Fossil Functional Teeth
5.3. Functional and Pre-Functional Teeth
5.4. Bite Force and Prey in Modern and Fossil Sharks with Serrated Tooth Shape
5.5. Diet of Fossil and Extant Sharks with Serrated Teeth
5.6. Serrated Teeth Replacement as an Evolutionary Advantage
Acknowledgments
References
- Agassiz, L. Recherches sur les Poissons Fossils, Volumes 1–5; Imprimerie de Neuchâtel: Soleure, Switzerland, 1833–1844; Volume 1, p. 420. [Google Scholar]
- Linnaeus, C. Systema Naturae per Regna Trianaturae, Secundum Classes, Ordines, Genera, Species cum Characteribus, Differentiis, Synonymis, Locis; Laurentius Salvius: Stockholm, Sweden, 1758; p. 824. [Google Scholar]
- Müller, J.; Henle, J. Systematische Beschreibung der Plagiostomen; Berlin, Germany, 1838–1841. [Google Scholar]
- Müller, J.; Henle, J. On the generic characters of cartilaginous fishes with descriptions of new genera. Mag. Nat. Hist. 1837, 2, 1–91. [Google Scholar]
- Compagno, L.J.V. Sharks of the World. An Annotated and Illustrated Catalogue of Shark Species Known to Date; Food and Agriculture Organization (FAO) Fisheries Synopsis: Rome, Italy, 1984; Volume 4, Part 1-Hexanchiformes to Lamniformes, pp. 1–250; Part 2-Carcharhiniformes, pp. 251–633. [Google Scholar]
- Mollet, H.; Cailliet, G.; Klimley, A.; Ebert, D.; Testi, A.; Compagno, L.V.G. A review of length validation methods and protocols to measure large white sharks. In Great White Sharks: The Biology of Carcharodon Carcharias; Klimley, A.P., Ainley, D.G., Eds.; Academic Press: San Diego, CA, USA, 1996; pp. 91–107. [Google Scholar]
- Shimada, K. The relationship between the tooth size and total body length in the white shark, Carcharodon carcharias (Lamniformes: Lamnidae). J. Fossil Res. 2003, 35, 28–33. [Google Scholar]
- Cruiz-Martinez, A.; Chiappa-Carrar, X.; Arenas-Fuentes, V. Age and growth of the Bull Shark, Carcharhinus leucas, from southern Gulf of Mexico. J. Northwest Atl. Fish. Sci. 2004, 35, 367–374. [Google Scholar]
- Goldman, K.J. Age and growth in elasmobranch fishes. In Management Techniques for Elasmobranch Fisheries; FAO Fisheries Technical Paper 474; Musick, J.A., Bonfil, R., Eds.; Food and Agricultural Organization of the United Nations: Rome, Italy, 2004; pp. 97–132. [Google Scholar]
- Tuma, R.E. An investigation of the feeding habits of the bull shark, Carcharias Leucas, in the Lake Nicaragua-Rio San Juan system. In Investigations of the Ichthyofauna of Nicaraguan Lakes; Thorson, T.B., Ed.; University of Nebraska Press: Lincoln, NE, USA, 1976; pp. 533–538. [Google Scholar]
- Ainley, D.G.; Strong, C.S.; Huber, H.R.; Lewis, T.J.; Morrell, S.H. Predation by sharks on pinnipeds at the Farallon Islands. Fish. B-NOAA 1981, 78, 941–945. [Google Scholar]
- Tricas, T.C.; Taylor, L.R.; Naftel, G. Diel behavior of the tiger shark, Galeocerdo cuvier, at French Frigate Shoals, Hawaiian Islands. Copeia 1981, 4, 904–908. [Google Scholar]
- Tricas, T.C.; McCosker, J.E. Predatory behavior of the White Shark Carcharodoncarcharias) with notes on its behavior. Proc. Calif. Acad. Sci. 1984, 42, 221–238. [Google Scholar]
- Long, D. Apparent predation by a White Shark Carcharodon carcharias on a pygmy sperm whale Kogia breviceps. Fish B-NOAA 1991, 89, 538–540. [Google Scholar]
- Rauzon, M.J. Sharks and albatross on laysan island. Ocean Realm 1993, 27–30. [Google Scholar]
- Dudley, S.F.J.; Anderson-Reade, M.D.; Thompson, G.S.; McMullen, P.B. Concurrentscavenging off a whale carcass by great white sharks, Carcharodon carcharias, and tiger sharks, Galeocerdo cuvier. Fish B-NOAA 2000, 98, 646–649. [Google Scholar]
- Fergusson, I.K.; Compagno, L.J.V.; Marks, M.A. Predation by white sharks Carcharodon carcharias (Chondrichthyes: Lamnidae) upon chelonians, with new records from the Mediterranean Sea and a first record of the ocean sunfish Mola mola (Osteichthyes: Molidae) as stomach contents. Environ. Biol. Fish. 2000, 58, 447–453. [Google Scholar] [CrossRef]
- Heithaus, M.R.; Dill, L.M.; Marshall, G.J.; Buhleier, B. Habitat use and foraging behavior of tiger sharks (Galeocerdo cuvier) in a seagrass ecosystem. Mar. Biol. 2002, 140, 237–249. [Google Scholar] [CrossRef]
- Hiethaus, M.R.; Frid, A.; Dill, L.M. Shark-inflicted injury frequencies, escape ability, and habitat use of green and loggerhead turtles. Mar. Biol. 2002, 140, 229–236. [Google Scholar] [CrossRef]
- Frazzetta, T.H. The mechanics of cutting and the form of shark teeth (Chondrichthyes, Elasmobranchii). Zoomorphology 1988, 108, 93–107. [Google Scholar] [CrossRef]
- Wroe, S.; Huber, D.R.; Lowry, M.; McHenry, C.; Moreno, K.; Clausen, P.; Ferrara, T.L.; Cunningham, E.; Dean, M.N.; Summers, A.P. Three-dimensional computer analysis of white shark jaw mechanics: How hard can a great white bite? J. Zool. 2008, 276, 336–342. [Google Scholar] [CrossRef]
- Huber, D.R.; Claes, J.M.; Mallefet, J.; Herrel, A. Is extreme bite performance associated with extreme morphologies in sharks? Physiol. Biochem. Zool. 2009, 82, 20–28. [Google Scholar] [CrossRef]
- Whitenack, L.B.; Simkins, D.C., Jr.; Motta, P.J. Biology meets engineering: The structural mechanics of fossil and extant shark teeth. J. Morphol. 2010, 272, 169–179. [Google Scholar]
- Whitenack, L.B.; Simkins, D.C., Jr.; Motta, P.J.; Hirai, M.; Kumar, A. Young’s modulus and hardness of shark tooth biomaterials. Arch. Oral Biol. 2010, 55, 203–209. [Google Scholar] [CrossRef]
- Breder, C.M., Jr. The shedding of teeth by Carcharias littoralis (Mitchill). Copeia 1942, 1, 42–44. [Google Scholar] [CrossRef]
- Reif, W.E.; McGill, D.; Motta, P. Tooth replacement rates of the sharks Triakissemifasciata and Ginglymostoma cirratum. Zool. Jahrb. 1978, 99, 151–156. [Google Scholar]
- Luer, C.A.; Blum, P.C.; Gilbert, P.W. Rate of tooth replacement in the nurse shark Ginglymostoma cirratum. Copeia 1990, 1, 182–191. [Google Scholar]
- Overstrom, N.A. Estimated tooth replacement in captive sand tiger sharks (Carchariastaurus Rafinesque 1810). Copeia 1991, 1, 525–526. [Google Scholar] [CrossRef]
- Shimada, K. Skeletal anatomy of the Late Cretaceous lamniform shark, Cretoxyrhinamantelli, from the Niobrara Chalk in Kansa. J. Vert. Paleontol. 1997, 17, 642–652. [Google Scholar] [CrossRef]
- Shimada, K. Skeletal and dental anatomy of lamniform shark, Cretalamna appendiculatafrom Upper Cretaceous Niobrara Chalk of Kansas. J. Vert. Paleontol. 2007, 27, 584–602. [Google Scholar] [CrossRef]
- Shimada, K.; Cicimurri, D.J. Skeletal anatomy of the Late Cretaceous shark, Squalicorax (Neoselachii: Anacoracidae). Palaeontol. Z. 2005, 79, 241–261. [Google Scholar]
- Welton, B.J.; Farish, R.F. The Collector’s Guide to Fossil Sharks and Rays from the Cretaceous of Texas; Before Time: Lewisville, TX, USA, 1993; pp. 1–204. [Google Scholar]
- Kent, B. Fossil Sharks of the Chesapeake Bay Region; Egan Rees and Boyer: Baltimore, MD, USA, 1994; pp. 1–146. [Google Scholar]
- Cappetta, H. Chondrichthyes II, Mesozoic and Cenozoic Elasmobranchii. In Handbook of Paleoichthyology; Schultze, H.P., Ed.; Gustav Fischer Verlag: Stuttgart, Germany, 1987; Volume 3B, pp. 1–193. [Google Scholar]
- Shimada, K.; Cicimurri, D.J. Skeletal anatomy of the Late Cretaceous shark, Squalicorax (Neoselachii: Anacoracidae). Palaeontol. Z. 2005, 79, 241–261. [Google Scholar]
- Shimada, K.; Cicimurri, D.J. The oldest record of the Late Cretaceous anacoracid shark, Squalicorax pristodontus (Agassiz) from the Western Interior, with comments on Squalicorax phylogeny. B. New Mexico Mus. Nat. Hist. Sci. 2006, 35, 177–184. [Google Scholar]
- Siverson, M.; Lindgren, J.; Kelley, L.S. Anacoracid sharks from the Albian (Lower Cretaceous) Pawpaw Shale of Texas. Palaeontology 2007, 50, 939–950. [Google Scholar] [CrossRef]
- Shimada, K. New anacoracid shark from Upper Cretaceous Niobrara Chalk of western Kansas, USA. J. Vertebr. Paleontol. 2008, 28, 1189–1194. [Google Scholar] [CrossRef]
- Schwimmer, D.R.; Stewart, J.D.; Williams, G.D. Scavenging by sharks of the Genus Squalicorax in the Late Cretaceous of North America. Palaios 1997, 12, 71–83. [Google Scholar] [CrossRef]
- Everhart, M.J. Late Cretaceous interaction between predators and prey: Evidence of feeding by two species of shark on a mosasaur. PalArch’s J. Vertebr. Palaeontol. 2004, 1, 1–7. [Google Scholar]
- Shimada, K.; Everhart, M.J. Shark-bitten Xiphactinus audax (Teleostei, Ichthyodectiformes) from the Niobrara Chalk (Upper Cretaceous) of Kansas. Mosasaur 2004, 4, 41–46. [Google Scholar]
- Cappetta, H.; Case, G.R. Contribution à l’étude des Sélaciens du groupe Monmouth (Campanien-Maestrichtien) du New Jersey. Palaeontogr. Abt. A 1975, 151, 1–46. [Google Scholar]
- Case, G.R. A new selachian fauna from the Judith River Formation (Campanian) of Montana. Palaeontogr. Abt. A 1978, 160, 76–205. [Google Scholar]
- Case, G.R. A new selachian fauna from the Late Campanian of Wyoming (Teapot Sandstone Member, Mesaverde Formation, Big Horn Basin). Palaeontogr. Abt. A 1987, 197, 1–37. [Google Scholar]
- Case, G.R.; Schwimmer, D.R. Late Cretaceous fish from the Blufftown Formation (Campanian) in western Georgia. J. Paleontol. 1988, 62, 290–301. [Google Scholar]
- Case, G.R.; Cappetta, H. A new selachian fauna from the Late Maastrichtian of Texas (Upper Cretaceous/Navarro Group; Kemp Formation). München Geowissenschaftliche Abh. A 1997, 34, 131–189. [Google Scholar]
- Robb, A.J., III. The upper cretaceous (campanian, black creek formation) fossil fish fauna of phoebus landing, Bladen County, North Carolina. Mosasaur 1989, 4, 75–92. [Google Scholar]
- Manning, E.M.; Dockery, D.T. A guide to the Frankstown vertebrate fossillocality (Upper Cretaceous), Prentiss County, Mississippi. Miss. Dept. Environ. Qual. Off. Geol. Circ. 1992, 4, 1–43. [Google Scholar]
- Becker, M.A.; Slattery, W.; Chamberlain, J.A., Jr. Mixing of santonian and campanian chondrichthyan and ammonite macrofossils along a transgressive lag deposit, Greene County, Western Alabama. Southeast. Geol. 1998, 37, 205–216. [Google Scholar]
- Becker, M.A.; Chamberlain, J.A., Jr.; Wolf, G. Chondrichthyans from the Arkadelphia formation (Upper Cretaceous: Upper Maastrichtian) of hot spring county Arkansas. J. Paleontol. 2006, 80, 700–716. [Google Scholar] [CrossRef]
- Hartstein, E.; Decina, L.; Keil, R. A late cretaceous (severn formation) vertebrate assemblage from Bowie, Maryland. Mosasaur 1999, 6, 17–24. [Google Scholar]
- Becker, M.A.; Chamberlain, R.B.; Chamberlain, J.A., Jr. Large carcharhinoid-type shark vertebrae in the Upper Cretaceous of New Jersey: Evidence for an anacoracid origin. Northeast. Geol. Environ. Sci. 2008, 30, 118–129. [Google Scholar]
- Moss, S.A. Feeding mechanisms in sharks. Am. Zool. 1977, 17, 355–364. [Google Scholar]
- Compagno, L.J.V. Sharks of the Order Carcharhiniformes; Princeton University Press: Princeton, NJ, USA, 1988; pp. 1–486. [Google Scholar]
- Shimada, K. Dental homologies in lamniform sharks (Chondrichthyes: Elasmobranchii). J. Morphol. 2002, 251, 38–72. [Google Scholar] [CrossRef]
- Becker, M.A.; Chamberlain, J.A., Jr.; Stoffer, P.W. Pathological tooth deformities in modern and late Cretaceous chondrichthyans: A consequence of feeding related injury. Lethaia 2000, 36, 1–16. [Google Scholar]
- Becker, M.A.; Slattery, W.; Chamberlain. J.A., Jr. Reworked Campanian and Maastrichtian macrofossils in a sequence bounding, transgressive lag deposit, Monmouth County, New Jersey. Northeast. Geol. Environ. Sci. 1996, 18, 234–252. [Google Scholar]
- Becker, M.A.; Wellner, R.W.; Mallery, C.S.; Chamberlain, J.A., Jr. Chondrichthyans from the lower ferron sandstone member of the mancos shale (upperCretaceous: Middle Turonian) of emery and carbon counties, Utah, USA. J. Paleontol. 2010, 84, 248–266. [Google Scholar] [CrossRef]
- Shimada, K.; Schumacher, B.A.; Parkin, J.A.; Palermo, J.M. Fossil marine vertebrates from the lowermost Greenhorn Limestone (Upper Cretaceous: Middle Cenomanian) in southeastern Colorado. J. Paleontol. 2006, 80, 1–45. [Google Scholar]
- Budker, P.; Whitehead, P. The Life of Sharks; Columbia University Press: New York, NY, USA, 1971; pp. 1–222. [Google Scholar]
- Hubble, G. Using tooth structure to determine the evolutionary history of the white shark. In Great White Sharks: The Biology ofCarcharodon carcharias; Klimley, A.P., Ainley, D.G., Eds.; Academic Press: San Diego, CA, USA, 1996; pp. 9–18. [Google Scholar]
- Shimada, K. Paleoecological relationships of the late Cretaceous lamniform shark, Cretoxyrhina mantelli (Agassiz). J. Paleontol. 1997, 71, 926–933. [Google Scholar]
- Renz, M. Megalodon: Hunting the Hunter; Paleo Press: Lehigh Acres, FL, USA, 2002; pp. 1–159. [Google Scholar]
- Zangerl, R. Chondrichthyes I: Paleozoic Elasmobranchii. In Handbook of Paleoichthyology; Schultze, H.P., Ed.; Gustav Fischer Verlag: New York, NY, USA, 1981; pp. 1–115. [Google Scholar]
- Maisey, J.G. Anatomical revision of the fossil shark, Hybodus fraasi (Chondrichthyes: Elasmobranchii). Am. Mus. Novit. 1986, 2857, 1–16. [Google Scholar]
- Maisey, J.G. Discovering Fossil Fishes; Henry Holt and Company: New York, NY, USA, 1996; p. 223. [Google Scholar]
- Williams, M.E. Tooth retention in cladodont sharks: With comparison between primitive grasping and swallowing, and modern cutting and gouging feeding mechanisms. J. Vertebr. Paleontol. 2001, 21, 214–226. [Google Scholar] [CrossRef]
- Botella, H. The oldest fossil evidence of dental lamina in sharks. J. Vertebr. Paleontol. 2006, 26, 1002–1003. [Google Scholar] [CrossRef]
- Botella, H.; Valenzuela-Ríos, J.I.; Martínez-Pérez, C. Tooth replacement rates in early chondrichthyans: A qualitative approach. Lethaia 2009, 42, 365–376. [Google Scholar] [CrossRef]
- Motta, P.J.; Wilga, C.D. Advances in the study of feeding behaviors, mechanisms, and mechanics of sharks. Environ. Biol. Fish. 2001, 60, 131–156. [Google Scholar] [CrossRef]
- Iscan, M.Y.; McCabe, B.Q. Analysis of human remains recovered from a shark. Forensic Sci. Int. 1995, 72, 15–23. [Google Scholar] [CrossRef]
- Randall, J.E. Review of the biology of the tiger shark (Galeocerdo cuvier). Aust. J. Mar. Fresh. Res. 1992, 43, 21–31. [Google Scholar] [CrossRef]
- Applegate, S.P.; Espinosa-Arrubarrena, L. The fossil history of Carcharodon and its possible ancestor, Cretolamna: A study in tooth identification. In Great White Sharks: The Biology of Carcharodon Carcharias; Klimley, A.P., Ainley, D.G., Eds.; Academic Press: San Diego, CA, USA, 1996; pp. 19–36. [Google Scholar]
- Gottfried, M.D.; Compagno, L.J.V.; Bowman, S.C. Size and skeletal anatomy of the giant “Megatooth” shark Carcharodon Megalodon. In Great White Sharks: The Biology of Carcharodon Carcharias; Klimley, A.P., Ainley, D.G., Eds.; Academic Press: San Diego, CA, USA, 1996; pp. 55–66. [Google Scholar]
- Klimley, A.P. The predatory behavior of the white shark. Am. Sci. 1994, 82, 122–133. [Google Scholar]
- Martin, R.A.; Hammerschlag, N.; Collier, R.S.; Fallows, C. Predatory behaviour of white sharks (Carcharodon carcharias) at Seal Island, South Africa. J. Mar. Biol. Assoc. UK 2005, 85, 1121–1135. [Google Scholar] [CrossRef]
- Randall, B.M.; Randall, R.M.; Compagno, L.V.J. Injuries to jackass penguins (Spheniscus demersus): Evidence for shark involvement. J. Zool. 1988, 214, 589–599. [Google Scholar] [CrossRef]
- Demere, T.A.; Cerutti, R.A. A Pliocene shark attack on a cethotheriid whale. J. Paleont. 1982, 56, 1480–1482. [Google Scholar]
- Cigala-Fulgosi, F. Predation (or possible scavenging) by a great white shark on an extinct species of bottlenosed dolphin in the Italian Pliocene. Tertiary Res. 1990, 12, 17–36. [Google Scholar]
- Ehret, D.J.; MacFadden, B.J.; Salas-Gismondi, R. Caught in the act: Trophic interactions between a 4-million year old white shark (Carcharodon) and a Mysticete whale from Peru. Palaios 2009, 24, 329–333. [Google Scholar] [CrossRef]
- Bianucci, G.; Sorce, G.B.; Storai, T.; Landini, W. Killing in the Pliocene: Shark attack on a dolphin from Italy. Palaeontology 2010, 53, 457–470. [Google Scholar] [CrossRef]
- Purdy, R.W. Paleoecology of fossil white sharks. In Great White Sharks. The Biology of Carcharodon carcharias; Klimley, A.P., Ainley, D., Eds.; Academic Press: San Diego, CA, USA, 1996; pp. 133–139. [Google Scholar]
- Schwimmer, D.R.; Stewart, J.D.; Williams, G.D. Scavenging by sharks of the genus Squalicorax in the Late Cretaceous of North America. Palaios 1997, 12, 71–83. [Google Scholar] [CrossRef]
- Neumann, C. Evidence of predation on Cretaceous sea stars from north-west Germany. Lethaia 2000, 33, 65–70. [Google Scholar] [CrossRef]
- Shimada, K.; Hooks, G.E. Shark-bitten Protostegid turtles from the Upper Cretaceous Mooreville Chalk, Alabama. J. Paleontol. 2004, 78, 205–210. [Google Scholar] [CrossRef]
- Becker, M.A.; Chamberlain, J.A., Jr.; Goldstein, L. Evidence for a shark-bitten turtle costal from the lowermost Navesink Formation (Campanian-Maastrichtian) Monmouth County, New Jersey. Northeast. Geol. Environ. Sci. 2006, 28, 174–181. [Google Scholar]
- Becker, M.A.; Meier, J.; Slattery, W. Spiral coprolites from the Upper Cretaceous Wenonah-Mt. Laurel and Navesink formations in the northern coastal plain of New Jersey. Northeast. Geol. Environ. Sci. 1999, 21, 181–187. [Google Scholar]
- Reif, W.E. Evolution of dermal skeleton and dentition invertebrates: The odontoderegulation theory. Evol. Biol. 1982, 15, 287–368. [Google Scholar]
- Cuny, G. Primitive neoselachian sharks: A survey. Oryctos 1998, 1, 3–21. [Google Scholar]
- Kriwet, J.; Klug, S. Diversity and biogeography patterns of Late Jurassicneoselachians (Chondrichthys: Elasmobranchii). Geol. Soc. Lond. Spec. Publ. 2008, 295, 55–70. [Google Scholar] [CrossRef]
- Underwood, C.J. Diversification of the Neoselachii (Chondrichthyes) during the Jurassic and Cretaceous. Palaeobiology 2006, 32, 215–235. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Becker, M.A.; Chamberlain, J.A., Jr. Squalicorax Chips a Tooth: A Consequence of Feeding-Related Behavior from the Lowermost Navesink Formation (Late Cretaceous: Campanian-Maastrichtian) of Monmouth County, New Jersey, USA. Geosciences 2012, 2, 109-129. https://doi.org/10.3390/geosciences2020109
Becker MA, Chamberlain JA Jr. Squalicorax Chips a Tooth: A Consequence of Feeding-Related Behavior from the Lowermost Navesink Formation (Late Cretaceous: Campanian-Maastrichtian) of Monmouth County, New Jersey, USA. Geosciences. 2012; 2(2):109-129. https://doi.org/10.3390/geosciences2020109
Chicago/Turabian StyleBecker, Martin A., and John A. Chamberlain, Jr. 2012. "Squalicorax Chips a Tooth: A Consequence of Feeding-Related Behavior from the Lowermost Navesink Formation (Late Cretaceous: Campanian-Maastrichtian) of Monmouth County, New Jersey, USA" Geosciences 2, no. 2: 109-129. https://doi.org/10.3390/geosciences2020109