Distribution and Diversity of Carboniferous and Permian Colonial Rugose Coral Faunas in Western North America: Clues for Placement of Allochthonous Terranes

Abstract
:1. Introduction


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GENERA | MISSISSIPPIAN | PENNSYLVANIAN | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Cord. | Arctic | Alex. | Stikine | Cord. | Arctic | Alex. | Stikine | Klamath | |
| Acrocyathus | + | + | |||||||
| Actinocyathus | + | + | |||||||
| Corwenia? | + | + | |||||||
| Cystolonsdaleia | + | + | + | ||||||
| "Diphyphyllum" | + | + | + | ||||||
| Dorlodotia? | + | ||||||||
| Durhamina | + | ||||||||
| Eastonastraea | + | ||||||||
| Fedorowskiella | + | ||||||||
| Fomichevella | + | + | |||||||
| Heintzella | + | + | + | ||||||
| Heritschioides | ? | + | + | ||||||
| Lonsdaleia | + | ||||||||
| Lonsdaleoides | + | ||||||||
| Nemistium | + | + | |||||||
| Paraheritschioides | + | + | + | ||||||
| Pararachnastraea | ? | + | |||||||
| Petalaxis | + | + | |||||||
| "Pseudodorlodotia" | + | ||||||||
| Schoenophyllum | + | ||||||||
| Sciophyllum | + | + | + | ||||||
| Siphonodendron | + | + | + | ||||||
| Stelechophyllum | + | + | + | ||||||
| "Thysanophyllum" | + | + | + | ||||||
| 2 new genera | + | ||||||||
| Species | Canada | Arctic | Alex | Species | Canada | Arctic | Alex |
|---|---|---|---|---|---|---|---|
| Acrocyathus pennsylvanica | + | + | Siphonodendron warreni | + | + | + | |
| Actinocyathus peratrovichensis | + | Stelechophyllum banffensis | + | + | + | ||
| "Diphyphyllum" klawockensis | + | Stelechophyllum birdi | + | + | |||
| "Diphyphyllum" venosum | + | Stelechophyllum macouni | ? | + | |||
| Sciophyllum alaskaensis | + | + | Stelechophyllum mclareni | + | + | ||
| Sciophyllum lambarti | + | + | "Thysanophyllum" astraeiforme | + | + | + | |
| Siphonodendron sinuosum | + | + | "Thysanophyllum" orientale | ? | + | ||
| Siphonodendron succinctus | + |
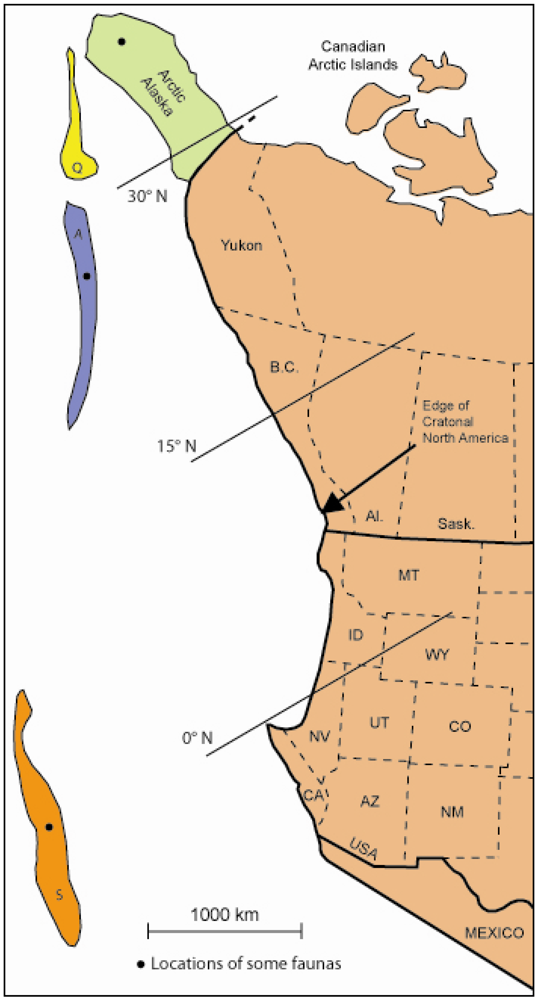
2. Mississippian Faunas
2.1. Western Interior Province of Canada
2.2. Faunas of Allochthonous Terranes
2.3. Faunal Relationships
2.4. Interpretation of Terrane Placement

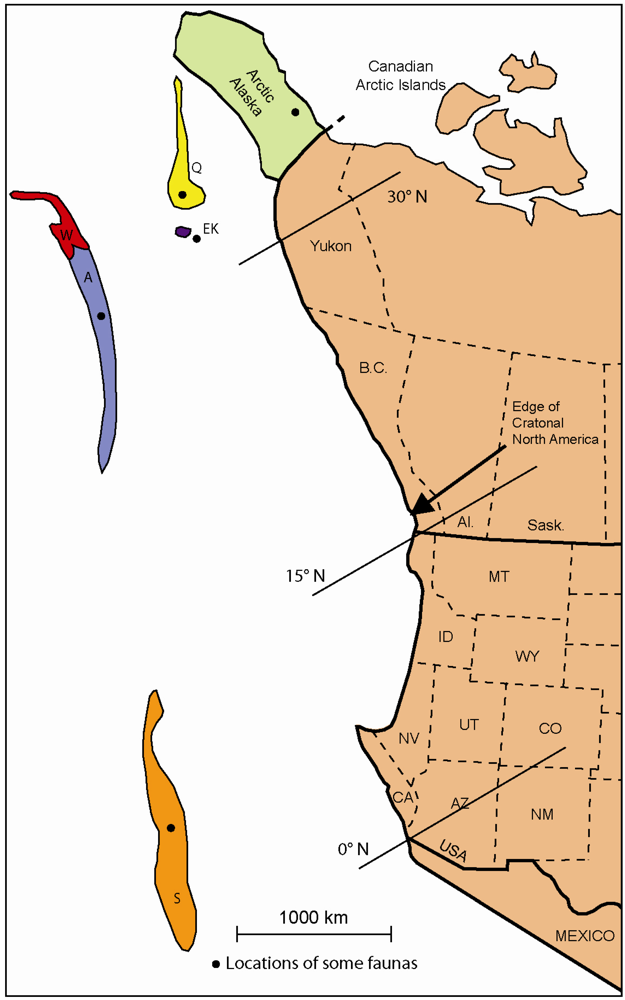
3. Pennsylvanian Faunas
3.1. Faunas of Cratonal North America
3.2. Faunas of Allochthonous Terranes and Arctic Alaska
3.3. Faunal Relationships
3.4. Interpretation of Terrane Placements in the Pennsylvanian

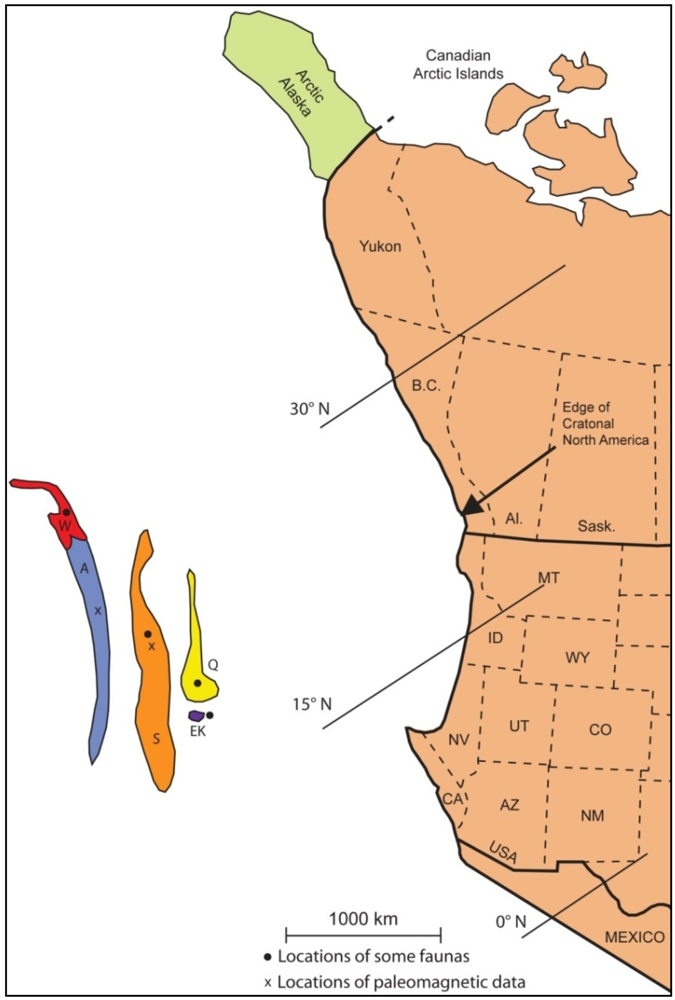
4. Early Permian Faunas
4.1. Faunas of cratonal North America
4.2. Faunas of the Terranes
4.3. Faunal Relationships
4.4. Interpretation of Terrane Placements in the Early Permian

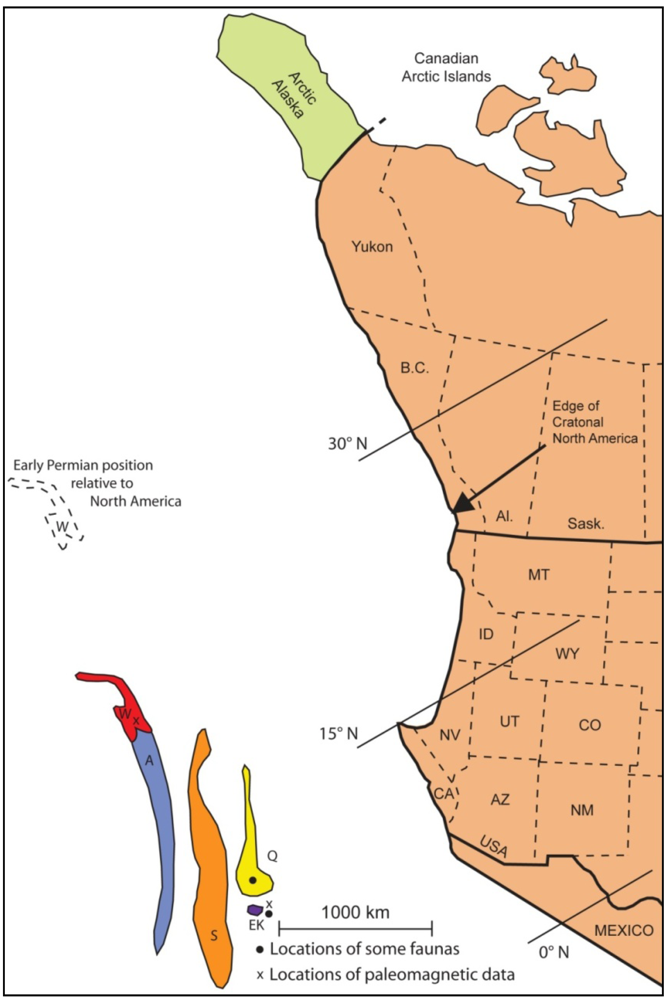
5. Middle Permian Faunas
5.1. Faunas in Terranes
5.2. Interpretation of Terrane Placement in the Middle Permian

6. Comparison of Terrane Placement with Previous Interpretations
7. Conclusions
Acknowledgments
References
- Kistler, R.W. Mesozoic Paleogeography of California: A viewpoint from isotope geology. In Mesozoic Paleogeography of the Western United States, Pacific Coast Paleogeography, Symposium 2; Howell, D.G., McDougall, K.A., Eds.; Pacific Section SEPM: Los Angeles, CA, USA, 1978; pp. 75–84. [Google Scholar]
- Stevens, C.H.; Stone, P.; Kistler, R.W. A speculative reconstruction of the middle Paleozoic continental margin of southwestern North America. Tectonics 1992, 11, 405–419. [Google Scholar] [CrossRef]
- Mankinen, E.A.; Irwin, W.P. Review of paleomagnetic data from the Klamath Mountains, Blue Mountains, and Sierra Nevada; Implications for paleogeographic reconstructions. Geol. Soc. Am. 1990, 255, 397–409. [Google Scholar]
- Colpron, M.; Nelson, J.L. A Palaeozoic NW passage and the Timanian, Caledonian, and Uralian connections of some exotic Teranes in the North American Cordillera. Geol. Soc. Lond. 2011, 35, 463–484. [Google Scholar] [CrossRef]
- Stevens, C.H.; Yancey, T.E.; Hanger, R.A. Significance of the provincial signature of Early Permian faunas of the eastern Klamath Terrane. Geol. Soc. Am. 1990, 255, 201–218. [Google Scholar]
- Armstrong, A.K. Mississippian Rugose Corals, Peratrovich Formation, West Coast, Prince of Wales Island, Southeastern Alaska; Geological Survey Professional Paper 534; US Government Printing Office: Washington, DC, USA, 1970a.
- Armstrong, A.K. Carbonate Facies and the Lithostrotionid Corals of the Mississippian Kogruk Formation, DeLong Mountains, Northwestern Alaska; Geological Survey Professional Paper 664; US Government Printing Office: Washington, DC, USA, 1970b.
- Sando, W.J.; Bamber, E.W. Coral zonation of the Mississippian System in the Western Interior Province of North America; Geological Survey Professional Paper 1134; US Government Printing Office: Washington, DC, USA, 1985. [Google Scholar]
- Wu, W.S.; Stevens, C.H.; Bamber, E.W. New Carboniferous and Permian Tethyan and Boreal corals from northwestern British Columbia, Canada. J. Paleontol. 1985, 59, 1489–1504. [Google Scholar]
- Gunning, M.H.; Fedorowski, J.; Bamber, E.W. Paleogeographic significance of mid-Carboniferous limestone, northwestern Stikine Terrane, British Columbia. In Paleogeography of the North American Cordillera: Evidence for and against Large-Scale Displacements, Special Paper; Haggart, J.W., Enkin, R.J., Monger, J.W.H., Eds.; Geological Association of Canada: St. John’s, Newfoundland, Canada, 2006; 46, pp. 59–69. [Google Scholar]
- Fedorowski, J.; Bamber, E.W.; Stevens, C.H. Lower Permian Colonial Rugose Corals, Western and Northwestern Pangaea: Taxonomy and Distribution; NRC Research Press: Ottawa, Canada, 2007. [Google Scholar]
- Stevens, C.H.; Belasky, P. Nature of Permian faunas in western North America: A key to the understanding of the history of Allochthonous Terranes. In Geomorphology and Plate Tectonics; Ferrari, D.M., Guiseppi, A.R., Eds.; Nova Scientific Publication: New York, NY, USA, 2009; pp. 275–310, Chapter 10. [Google Scholar]
- Stevens, C.H. Corals from a dismembered late Paleozoic Paleo-Pacific Plateau. Geology 1983, 11, 603–606. [Google Scholar] [CrossRef]
- McGhee, G.R., Jr.; Bottjer, D.J.; Droser, M.L. Ecological ranking of Phanerozoic biodiversity crises: The Serpukhovian (early Carboniferous) crisis had a greater ecological impact than the end-Ordovician. Geology 2012, 40, 147–150. [Google Scholar] [CrossRef]
- Scotese, C.R.; McKerrow, W.S. Revised world maps and introduction. Geol. Soc. Lond. 1990, 12. [Google Scholar] [CrossRef]
- Stevens, C.H. A new Middle Pennsylvanian species of Petalaxis (Rugosa) from eastern California. J. Paleontol. 1995, 69, 787–789. [Google Scholar]
- Bamber, E.W. Personal Communication. Geological Survey of Canada: Calgary, Canada, 15 January 2012. [Google Scholar]
- Stevens, C.H. Fasciculate rugose corals from Gzhelian and Lower Permian strata, Pequop Mountains, northeast Nevada. J. Paleontol. 2008, 82, 1190–1200. [Google Scholar] [CrossRef]
- Armstrong, A.K. Pennsylvanian Carbonates, Paleoecology and Rugose Colonial Corals, North Flank, Eastern Brooks Range, Arctic Alaska; US Geological Survey Professional Paper; US Government Printing Office: Washington, DC, USA, 1972; Volume 747. [Google Scholar]
- Katvala, E. Personal Communication. University of Calgary: Calgary, Canada, 10 April 2010. [Google Scholar]
- Kawamura, T.; Stevens, C.H. Middle Pennssylvanian rugose corals from the Baird Formation, Klamath Mountains, northwestern California. J. Paleontol. 2012, 86, 513–520. [Google Scholar]
- Gardner, M.C.; Bergman, S.C.; Cushing, G.W.; MacKevett, E.M., Jr.; Plafker, G.; Campbell, R.B.; Dodds, C.J.; McClelland, W.C.; Mueller, P.A. Pennsylvanian pluton stitching of Wrangellia and the Alexander Terrane, Wrangell Mountains, Alaska. Geology 1988, 16, 967–971. [Google Scholar]
- Bacon, C.R.; Vazquez, J.A.; Wooden, J.L. Peninsular Terrane basement ages recorded by Paleozoic and Paleoproterozoic zircon in Gabbro Xenoliths and Andesite from Redoubt Volcano, Alaska. Geol. Soc. Am. Bull. 2012, 124, 24–34. [Google Scholar]
- Stevens, C.H. Occurrence and migration of the “Northern” massive Rugosa in the Early Permian. In Drevnei Cnidaria; Sokolov, B.S., Ed.; pp. 197–204, Akademiia Nauk SSR, Sibirskoe Otdelenie, 1975; Tom 2, vyp. 202.
- Miller, M.M. Dispersed remnants of a northeast Pacific fringing arc: Upper Paleozoic Terranes of Permian McCloud faunal affinity, western United States. Tectonics 1987, 6, 807–830. [Google Scholar]
- Stevens, C.H. Permian colonial rugose corals from the Wrangellian terrane in Alaska. J. Paleontol. 2008, 82, 1043–1050. [Google Scholar] [CrossRef]
- Belasky, P.; Stevens, C.H. Permian faunas of Westernmost North America: Paleobiogeographic Constraints on the Permian positions of Cordilleran Terranes. In Paleogeography of the North American Cordillera: Evidence for and against Large-Scale Displacement, Special Paper; Haggart, J.W., Enkin, R.J., Monger, J.W.H., Eds.; Geological Association of Canada: St. John’s, Newfoundland, Canada, 2006; Volume 46, pp. 71–80. [Google Scholar]
- Irving, E.; Monger, J.W.H. Preliminary paleomagnetic results from the Permian Asitka Group, British Columbia. Can. J. Earth Sci. 1987, 24, 1490–1497. [Google Scholar] [CrossRef]
- Butler, R.F.; Gehrels, G.E.; Bazard, D.R. Paleomagnetism of Paleozoic strata of the Alexander Terrane, southeastern Alaska. Geol. Soc. Am. Bull. 1997, 109, 1372–1388. [Google Scholar]
- Monger, J.W.H.; Ross, C.A. Distribution of fusulinaceans in the western Canadian Cordillera. Can. J. Earth Sci. 1971, 8, 259–278. [Google Scholar] [CrossRef]
- Beauchamp, B.; Desrochers, A. Permian warm-to very cold-water carbonates and cherts in northwestern Pangea. In Cool-Water Carbonates; James, N.P., Clark, J.A.D., Eds.; Society for Sedimentary Geology: Tulsa, OK, USA, 1997; pp. 327–347, Special Publcations No. 56. [Google Scholar]
- Wardlaw, B.R. Middle-Late Permian paleogeography of Idaho, Montana, Nevada, Utah, and Wyoming. AAPG Bull. 1980, 64, 353–361. [Google Scholar]
- Nelson, S.J.; Nelson, E.R. Allochthonous Permian micro- and macrofauna, Kamloops area, British Columbia. Can. J. Earth Sci. 1985, 22, 442–451. [Google Scholar]
- Stevens, C.H.; Miller, M.M.; Nestell, M. A new Permian waagenophyllid coral from the Klamath Mountains, California. J. Paleontol. 1985, 61, 691–699. [Google Scholar]
- Monger, J.W.H. Stratigraphy and structure of the Upper Paleozoic rocks, northeast Dease Lake map area, British Columbia (104J). Geol. Surv. Canada 1969, 41, Paper 68-48. [Google Scholar]
- Panuska, B.C.; Stone, D.B. Latitudinal motion of Wrangellia and Alexander Terranes and the southern Alaska Superterrane. In Tectonostratigraphic Terranes of the Circum-Pacific Region; Howell, D.B., Ed.; Circum-Pacific Council on Energy and Mineral Resources: Houston, TX, USA, 1985; pp. 109–119. [Google Scholar]
- Skinner, J.W.; Wilde, G.L. New fusulinids from the Permian of West Texas. J. Paleontol. 1955, 29, 927–940. [Google Scholar]
- Miller, E.L.; Kuznetsov, N.; Soboleva, A.; Udoratina, O.; Grove, M.J.; Gehrels, G. Baltica in the Cordillera? Geology 2011, 39, 791–794. [Google Scholar] [CrossRef]
- Stevens, C.H. A giant Permian fusulinid from east-central Alaska with comparisons of all giant fusulinids in western North America. J. Paleontol. 1995c, 69, 805–812. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Stevens, C.H. Distribution and Diversity of Carboniferous and Permian Colonial Rugose Coral Faunas in Western North America: Clues for Placement of Allochthonous Terranes. Geosciences 2012, 2, 42-63. https://doi.org/10.3390/geosciences2020042
Stevens CH. Distribution and Diversity of Carboniferous and Permian Colonial Rugose Coral Faunas in Western North America: Clues for Placement of Allochthonous Terranes. Geosciences. 2012; 2(2):42-63. https://doi.org/10.3390/geosciences2020042
Chicago/Turabian StyleStevens, Calvin H. 2012. "Distribution and Diversity of Carboniferous and Permian Colonial Rugose Coral Faunas in Western North America: Clues for Placement of Allochthonous Terranes" Geosciences 2, no. 2: 42-63. https://doi.org/10.3390/geosciences2020042