1. Introduction
In a dynamic and noisy world, it is best to attend to reliable cues and when possible disregard those that may be irrelevant, misleading, or at times redundant [
1]. The more distantly removed—in space or time—a secondary cue is from a corresponding target, the less likely that cue will reliably convey information about the target. In other words, it is best to rely relatively more on direct target cues and relatively less on indirect cues. However, flexibly attending and responding to different sources of information can provide coping mechanisms in complex environments [
2,
3]. Bees depend upon floral resources in constant flux of bloom, depletion, and expiration, which provide multiple signal cues, such as color, shape, taste, and size. Additionally, the environment provides many potential indirect or secondary indicators of nearby targets, including: foliage and vegetation; conspecifics and other animals; and airborne scent. Any one of these cues may signal nectar or pollen, but in redundant combinations not all may elicit equal attention.
Secondary cues may be learned as being generally associated with certain types of stimuli or specifically associated with a specific target. General correspondents provide flexible information about common associations among similar types of stimuli, such as the higher likelihood of flowers being located in areas with foliage than areas without. Specific correspondents provide information about individual stimuli, such as the learning of particular landmarks along a route to a known flower patch [
4]. Delayed matching tasks test the capacity to see a cue, separated from a target, and then from multiple stimuli select a target or location corresponding to that cue directly or symbolically. The task has been adapted in studies of animal cognition for a variety of species to determine what is remembered and for how long [
5]. Bumblebees can match-to-sample (cue remains visible when choosing among stimuli) in a ‘same-different’ task [
6]. They can also learn to respond to contextual priming color cues when choosing between two subsequently presented stimuli as long as the pairs of colors were presented at two different places [
7]. Honeybees can learn to solve both symbolic [
8] and non-symbolic delayed matching to sample (DMTS) tasks including categorization [
9,
10]. Honeybees are further able to learn to disregard an additional irrelevant cue in a DMTS task, and transfer this learning to a novel cue but only when the relevant cue is provided in a consistent place or position in a sequence [
11]. Delayed matching to sample in bumblebees, however, has remained elusive. Possible obstacles include foraging strategies [
12] and the abilities in color discrimination and object detection [
13] of bumblebees, which differ from those of honeybees. Nonetheless, hints of success, usually by one or a few individuals, suggest that this capacity or variations may yet be elicited in bumblebees under some circumstances.
One significant challenge for bumblebees in tests of DMTS may be the location variation needed to test ‘true’ matching to sample instead of positional learning, similar to landmark cue use. However, this condition is relatively unnatural; while bees may face challenges of flower growth, depletion, and expiration, flowers do not move. In a confined laboratory setting, the constantly changing location of known stimuli may undermine tests of delayed matching. Context can provide necessary discriminating information for bumblebees when faced with a delayed priming task [
7] and tasks requiring matching of one cue to a reward and another to their nest [
14,
15]. Stable location of target stimuli may facilitate cue to target correspondence in a variation of traditional DMTS whereby a non-spatial cue can correspond to a predictable location [
16].
The use of a picture cue in a delayed matching task may further facilitate bumblebee performance. Previous findings have determined that bumblebees can both easily differentiate an object from a corresponding picture but also perceive a relation between them [
17]. As such, photographs can play a unique role in cue learning: (i) potentially being more likely to trigger memory for, and therefore approach to, the corresponding target; (ii) and lessen the risk that experience with an unrewarding cue will diminish the strength of rewarded training to the target, as may occur when the cue and target are exactly the same. In other words, the subject can learn that the picture of the object (the cue) is unrewarding but the object itself (target) is rewarding, as opposed to the object (cue) seen first being unrewarding but that same object seen later (target) being rewarding. It has been suggested that honeybee performance on DMTS may improve by a ‘win-stay’ approach, whereby honeybees are more likely to approach more familiar stimuli [
12,
18]. Honeybees and bumblebees display significant differences in foraging strategy and behavior, and it is possible that among bumblebees, a familiar but unrewarding stimulus (cue) could reduce rather than increase the likelihood of approaching the same type of stimulus again (target).
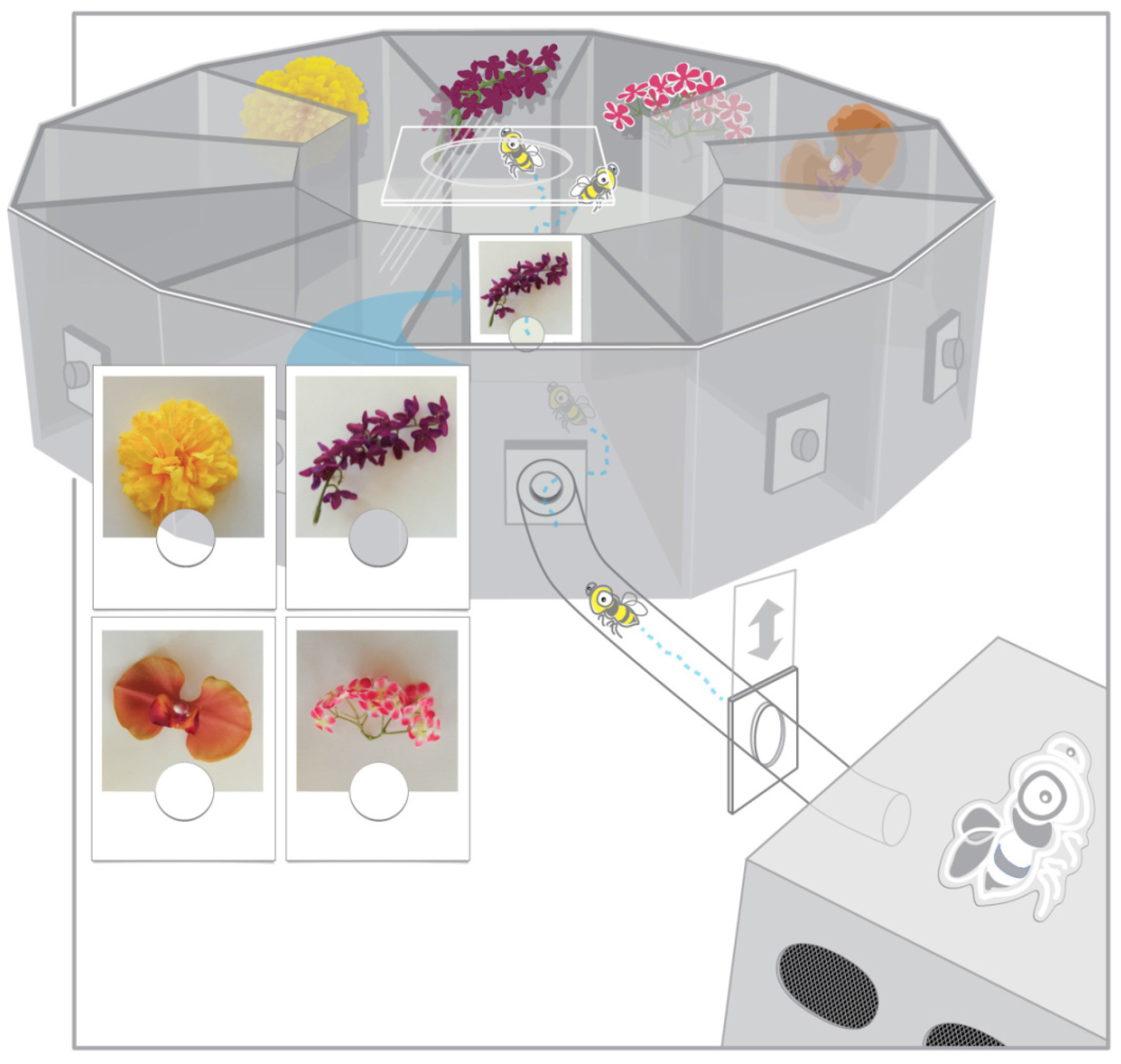
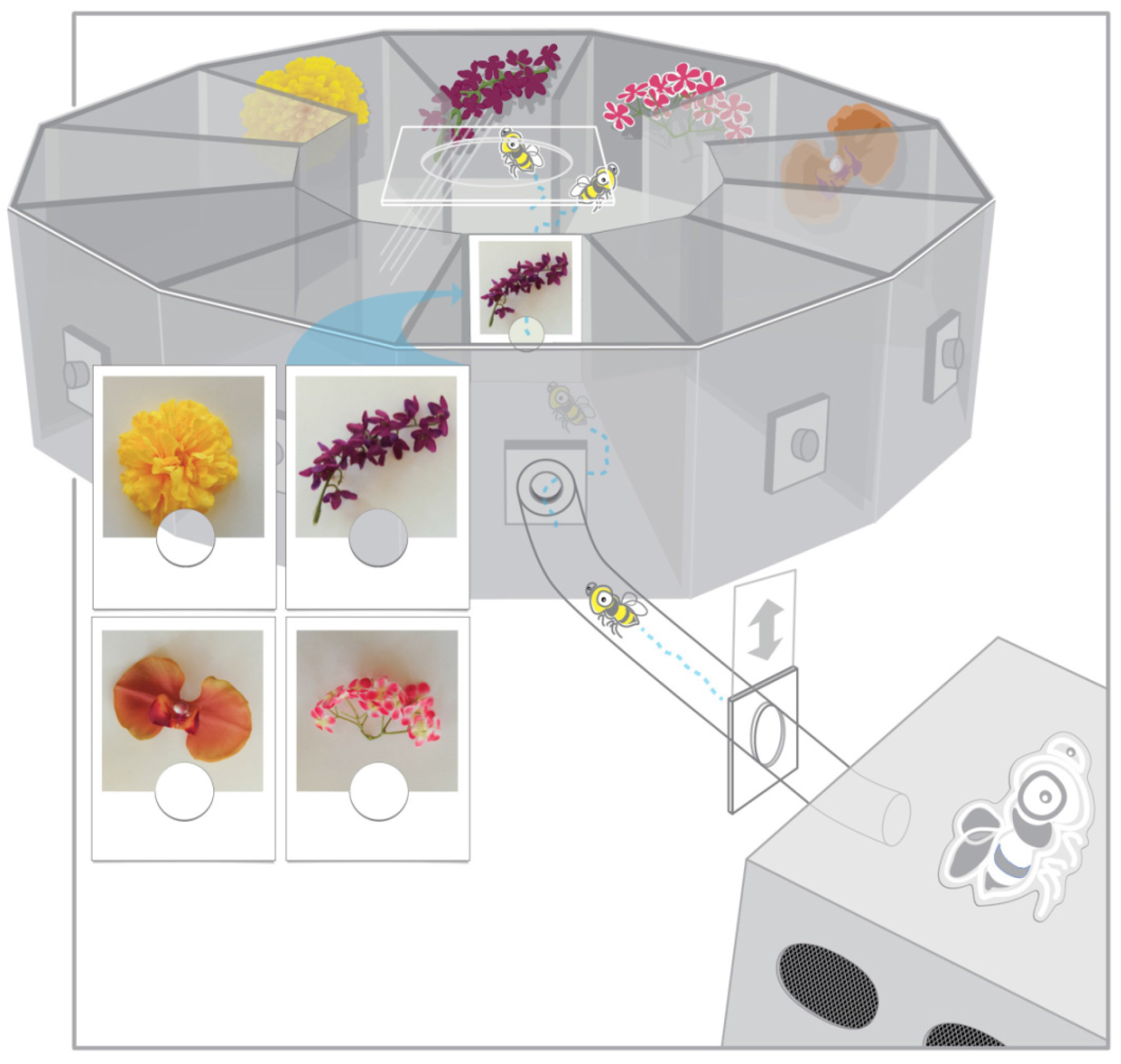
In the present study, photographic cues corresponding to a selection of target stimuli test cue use by bumblebees in an alternative DMTS task with stable target location whereby a cue signals a rewarding target among four stimuli but all four stimuli remain consistently positioned [
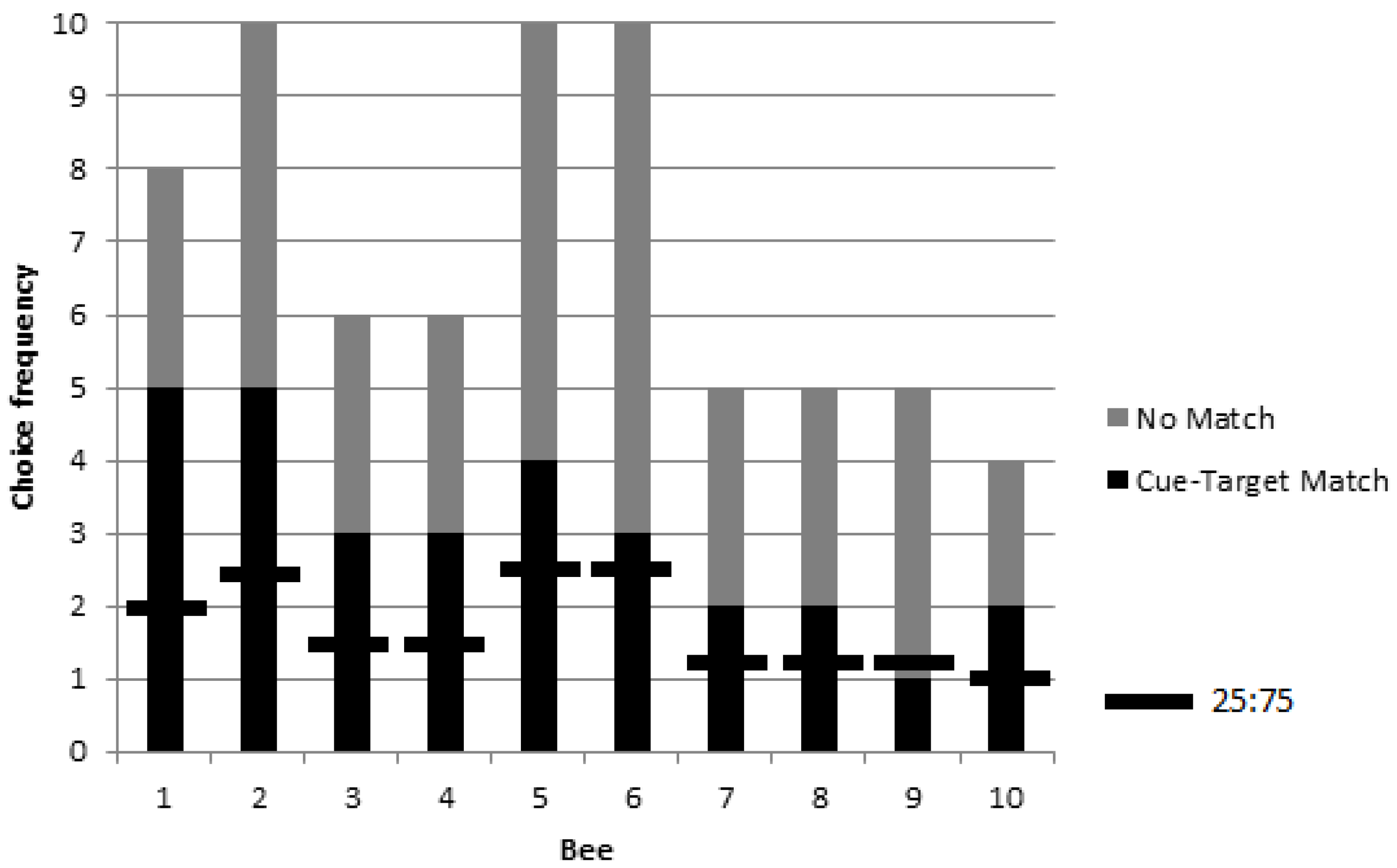
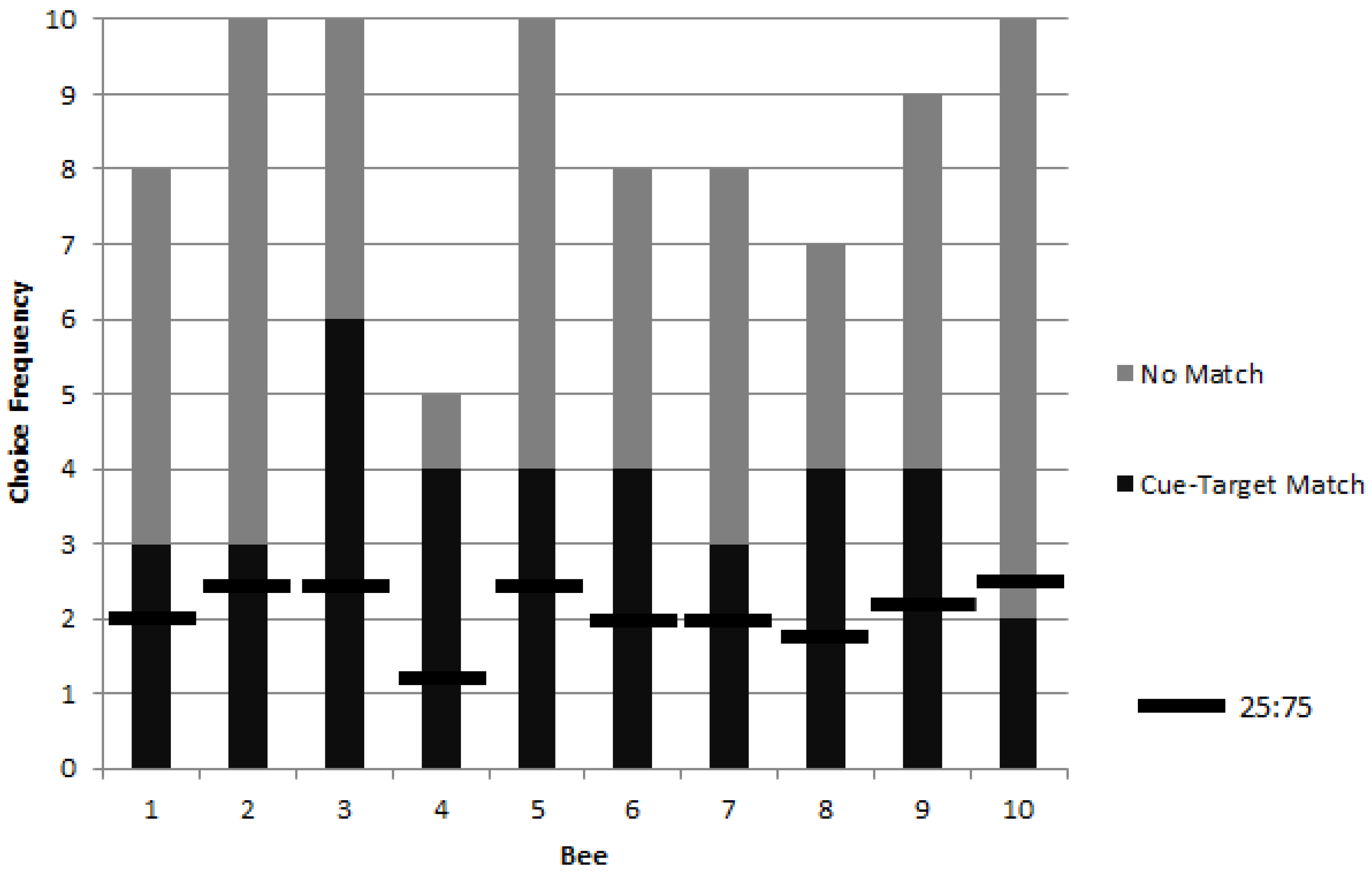
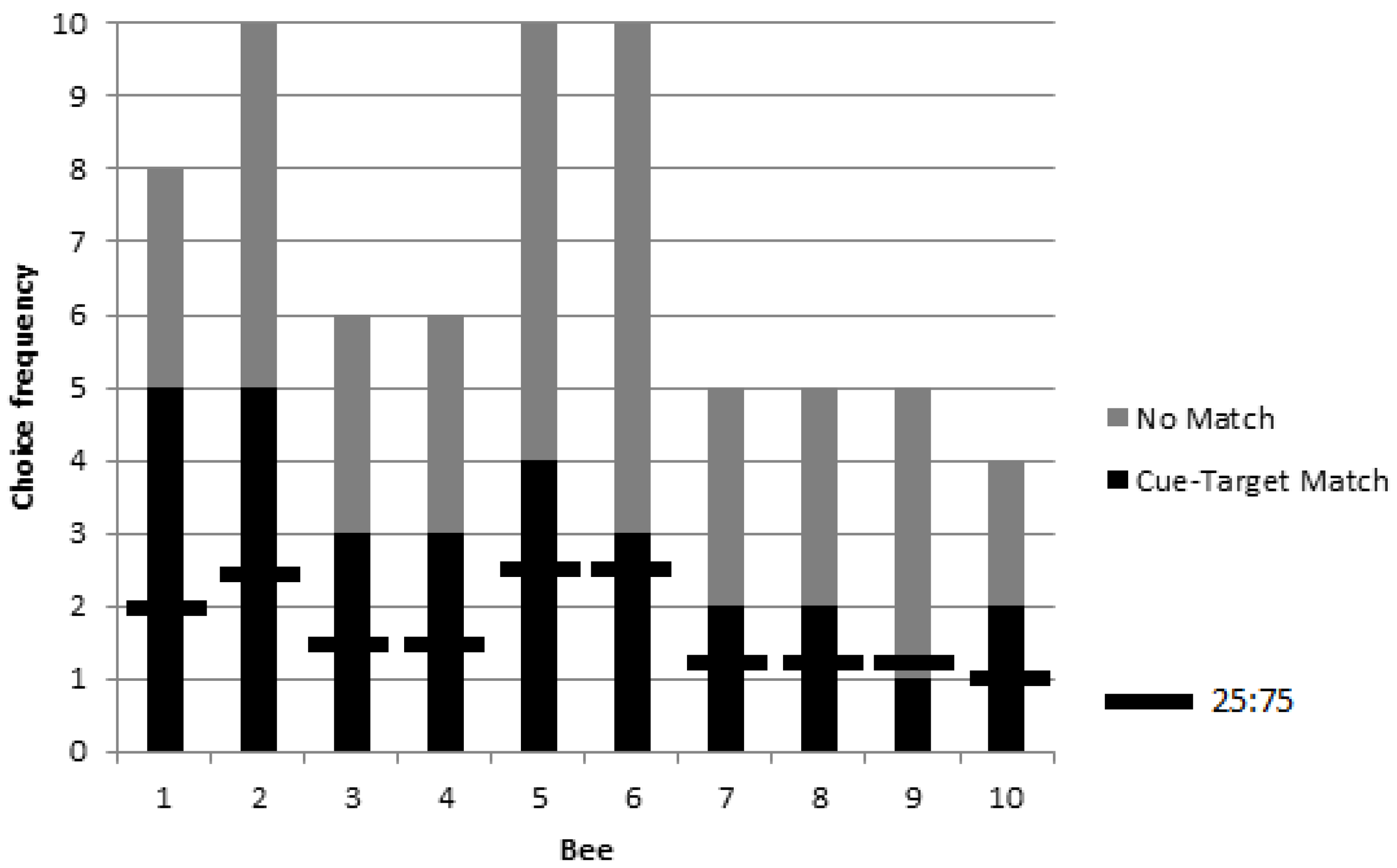
16]. The photograph cue-target combination was chosen to also facilitate association between cue and target and potential novel transfer to never before seen photograph and object stimuli during testing. Cue-target matching was defined as approaching the object corresponding to a photographic cue immediately following a cue-target combination change. Bees displaying matching at levels above chance during training were subsequently tested for rewarded, unrewarded, and novel preference for targets matching picture cues.
4. Discussion
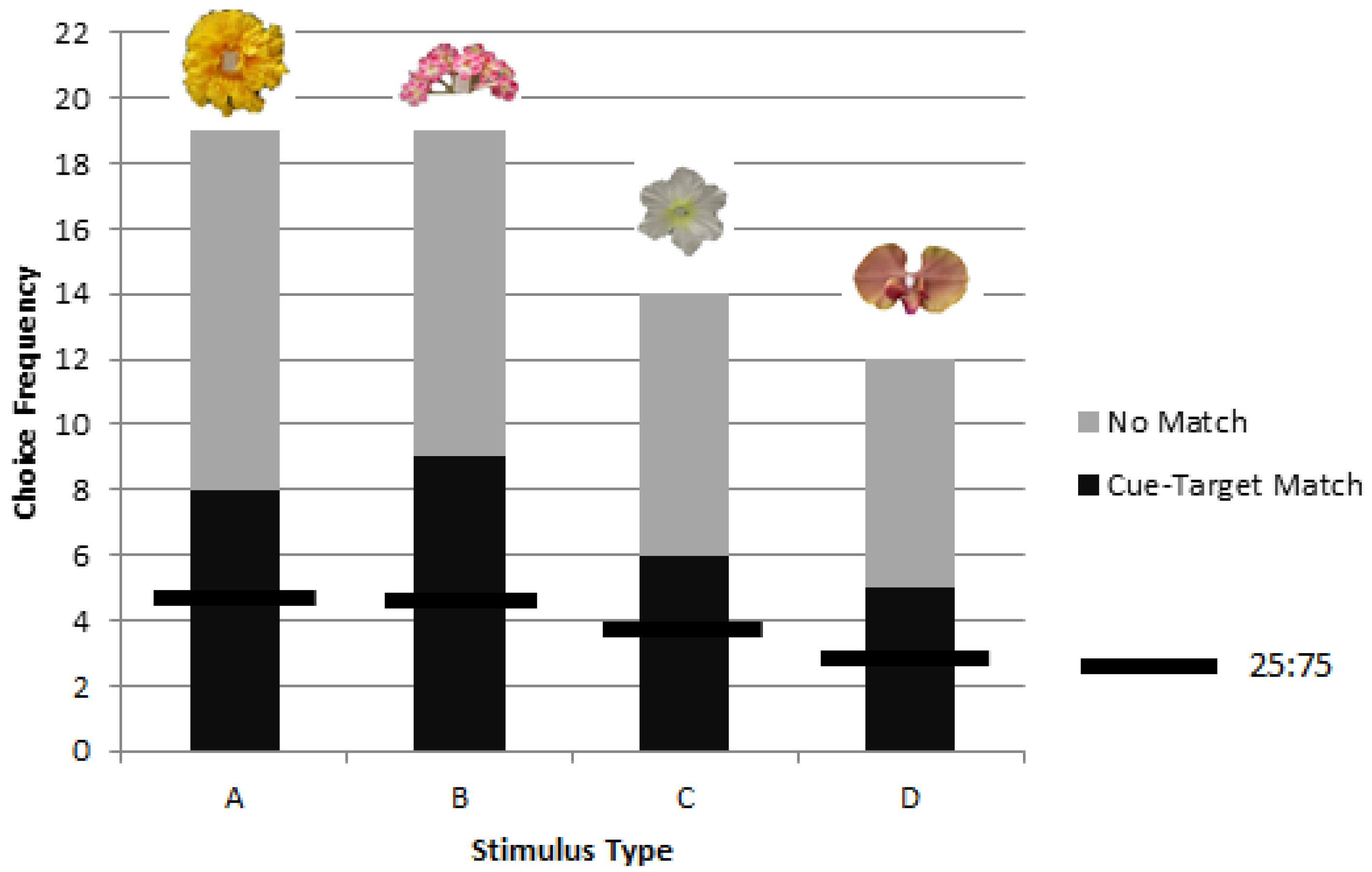
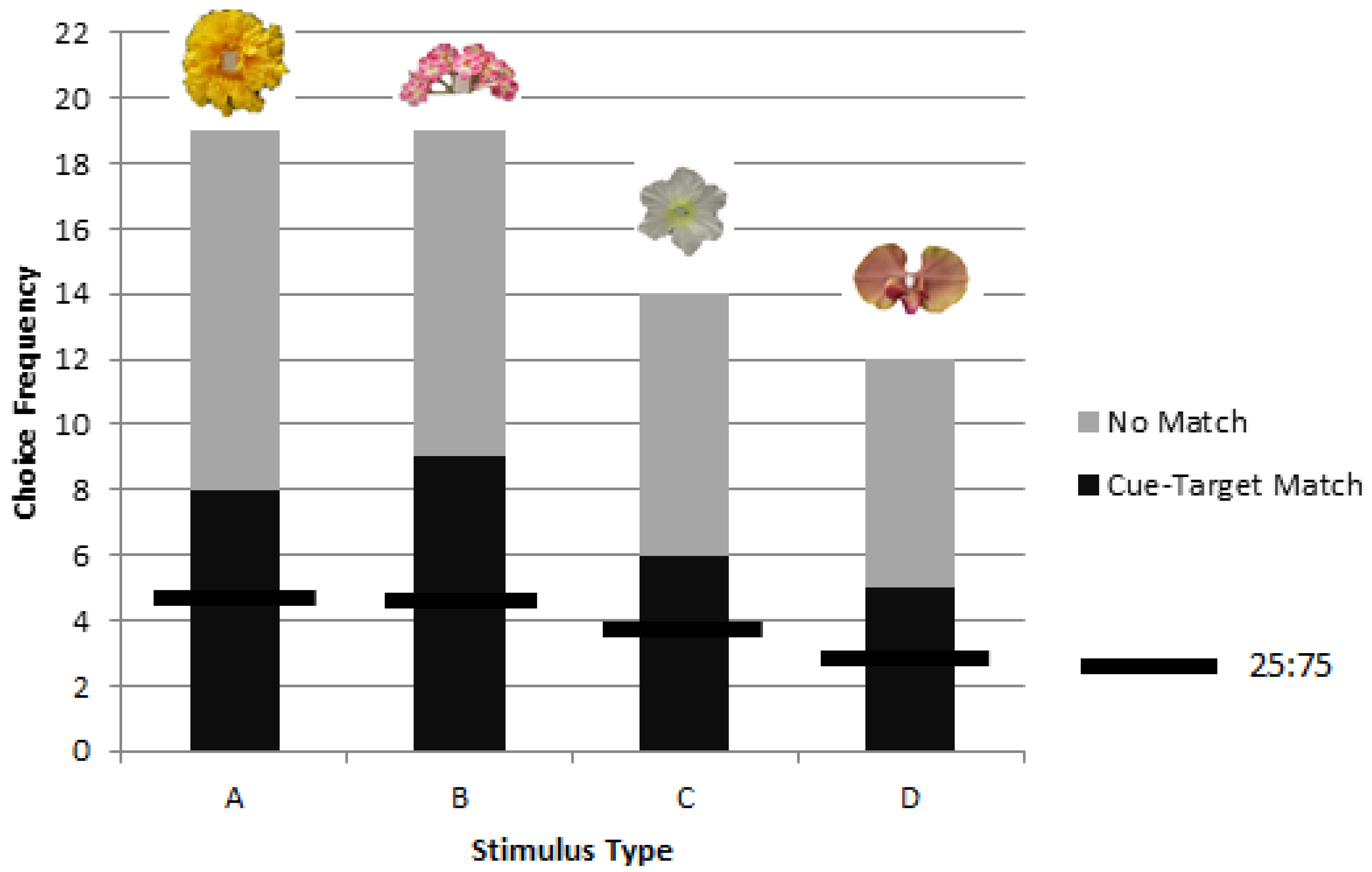
Bumblebees can learn to use a picture cue to find rewarding stimuli while foraging in a delayed matching task when the stimuli remain in consistent positions. Both rewarded and unrewarded testing showed significant preference for object stimuli matching picture cues. We obtained no evidence that matching would appear to transfer to novel cue-target combinations but the number of choices was limited to a single test trial for each of eight bees, and so there is no claim here to have demonstrated an inability in transfer learning. Without evidence of novel transfer or location change, the present results of delayed matching may correspond to delayed cue-target matching, cue-location matching, or a combination whereby the cue became associated with the location facilitated by picture to target correspondence. In the future, methods used in research on spatial and non-spatial coding of objects by vertebrates might be adapted for use with invertebrates to rule on the question of the contents of learning [
22]. In the present study, only the cue predicted which target would be rewarding. Although floral constancy, the tendency to approach relatively more familiar stimuli, could also explain the choice pattern of ‘matching’ corresponding objects to picture stimuli, it is unlikely given that the image cues were never rewarding. Positioning of the cue at the top or the bottom of the gate was random and provided no information about reward. Unrewarded testing of picture cue-target matching was consistent with DMTS proportions found for honeybees [
23], and this study could serve as a stepping stone towards a systematic species comparison in bee cognition. However, the bumblebees in the present study may have been using the picture cues as route cues, signaling the location of reward and not necessarily matching the cue and object. In the future, tests with targets placed in novel locations, or new stimuli placed in old locations, might serve to distinguish between these possibilities. The delayed matching task employed in this study retained unchanging position for target stimuli within the maze to reduce the previously suggested difficulty reflected by constantly changing ‘flower’ locations within a small, familiar environment.
Past research had determined that bumblebees perceive both a difference and similarity between pictures and corresponding object targets [
17]. The results of the current study further show that pictures can also be used by bumblebees in a delayed matching task. It is possible that the picture cue became associated with both a known location and the corresponding target therein. Multimodal floral signaling can increase learning speed, persistence, and facilitate memory among foragers [
24,
25]. Secondary or associative cues can further provision serial priming cues to facilitate detection before a target becomes visible [
26]. Airborne scent in particular is believed to trigger memory and foraging for the corresponding, but out of sight, flower [
27]. While naturally learned secondary cues, specific to a route or location, would not necessarily resemble the target, directed foraging may benefit from using cues similar to the target itself, by triggering memory for the similar, corresponding stimulus. How bees judge the similarity between a 3D flower and its 2D image is not well understood. In our previous research [
17], we found that for one flower in particular, used again in the present study (stimulus ‘B’), neither the black and white silhouette nor the drawing of the flower were judged as similar to the object. The image somehow failed to capture some important elements of the object itself. The same may have been true here when colony 1 failed to match stimulus ‘B’ to the cue. Given that another group succeeded, the question of possible colony differences, as documented in other learning tasks [
28], also remain to be addressed.
Observations made throughout this study highlight some additional components that may contribute to delayed matching by bumblebees but require further exploration. The present results, in combination with past difficulty training bumblebees to DMTS [
12], may suggest additional conditions required for bumblebee learning in a delayed matching task. Below, we speculate how the following aspects of our task design may have contributed to success: (i) a history of foraging experience; (ii) high reward inconsistency; (iii) and stationary location of familiar stimuli.
(i) Experience
Training in the current study required days (from 3 to 11) of exposure before bees exhibited a preference for stimuli matching the photograph cue. A meta-analysis of DMTS among many animals including honeybees [
23] suggested that amount of training improves signal to noise differentiation and recall for memories of cues. Other studies have shown that facilitation or pre-training can influence learning of an otherwise impossible task or shift strategic or attentional processes. Experience or facilitated learning has been found to be beneficial or essential in the performance of the bumblebee: reversal learning; use of global over local cues in navigation and rotated image discrimination [
12,
29,
30]. Facilitated learning may well have contributed in this study to experienced bees later exhibiting matching but it cannot be ruled out that any form of foraging experience may facilitate delayed matching. A general history of foraging experience may simply encourage bees to remain active despite high levels of inconsistency and difficulty finding reward.
(ii) Reward Inconsistency
Selective attention best facilitates foraging or hunting for consistent resources but when resources change, either declining or increasing in availability, broadening of attention becomes advantageous [
31]. Changing conditions can alter the relative worth of various resources as well as the corresponding sources of information signaling those resources by re-directing attention and reinforcing memories or recognition of alternatives [
25,
32].
Successful delayed matching by bumblebees in this study may have required conditions of high reward inconsistency. Bumblebees do show flexibility under inconsistent or changing circumstances, and outperform the more persistent honeybee on tasks of reversal learning with repeated experience [
12]. In the current study, when the same target was rewarded twice or more in a row (blocked trials), bees exhibited a strong preference for the previously rewarded stimulus, disregarding the cue. However, when the cue and rewarding target combination were changed following every foraging visit for each bee (alternating trials), matching was observed and evidenced by both rewarded and unrewarded testing. Foraging animals often rely on selective attention to best attend to relevant over irrelevant stimuli but still gather information broadly when needed due to resource change, loss, or depletion [
31]. It has been found that blue jays used a predictive signal when choosing between two stimuli but, as with the bumblebees in the present study, only when the signal was reliable and under inconsistent environmental conditions [
33]. The signal was ignored if unpredictable and the environment was consistent. The possible effect of inconsistent resources influencing delayed matching has also been suggested for the spider
Misumena vatia [
34].
While floral constancy may represent a more energy efficient and focused strategy when compared with sampling, information about alternate resources may still be retained for later use when resource state changes and sampling becomes necessary [
35]. In this study, during blocked trials, bees could rely on a relatively successful perseverating strategy because change occurred only following repeated reward with the same stimulus. Only when perseverating was never successful, during alternating trials, did the bees begin to exhibit matching and use of the picture cue.
Lastly, interference may be reduced with increased number of cues and reward variability among cues. Repeated or prolonged exposure to a cue can interfere with acquisition of alternate cues. Greater numbers of stimuli are known to reduce interference in DMTS [
16], and picture stimuli changed more frequently may further reduce interference, reducing prolonged exposure to any one stimulus. In other words, the use of four stimuli and corresponding cues for which reward was changed frequently (alternating trials) may have reduced the potential for interference.
(iii) Location
As mentioned, the delayed matching evidenced in this study may correspond to either a cue-target match or a cue-location match. Changing target stimuli location previously resulted in significantly decreased foraging activity, but this does not necessarily suggest that bees were learning a cue-location match. In a confined laboratory space, the movement of stimuli within a familiar environment corresponds poorly with natural challenges of floral cycle. Field study may be better able to replicate stimuli change and thereby test for DMTS without stable location of target stimuli. However, it is also likely that the bees were learning to use the picture cues as spatial cues to direct location or route choice rather than matching the picture to the corresponding object [
7,
36]. While secondary, environmental cues may have limited value predicting rewarding resources, spatial cues used along a route to rewarding resources are commonly used in bee navigation [
7,
9,
12,
36]. Airborne scent cues likewise rely on directing attention to the corresponding flower of a known location to elicit foraging for a specific flower [
32].
5. Conclusions
The results of the present study do evidence a capacity among bumblebees for learned picture cue use when foraging in a delayed matching task, under conditions of high environmental inconsistency without variation in target location. Experience is likely needed for foraging bumblebees to persist with a task in which only one stimulus is rewarding at a time and the rewarding stimulus changes constantly. This likely corresponds to a preferred reliance on primary cues, those directly presented by the potentially rewarding stimulus (flower color, size, location, etc.), instead of relying on secondary or associated cues, likely to be less predictive or reliable (e.g., presence of foliage). Switching from a preferred to a less preferred strategy has been observed in bee navigation, whereby bees will disregard landmark cues in favor of route memory unless that route memory is unreliable [
37]. Similarly, although illumination is most often disregarded as a misleading feature (which changes throughout the day), bees can learn to attend to and use this cue when it is the only predictive cue for reward [
38]. The results of the current study likewise suggest that bumblebees prefer alternative strategies to delayed matching but can learn to use cues when foraging if preferred behavior patterns fail in a highly inconsistent environment. Perseveration is a relatively less risky and costly method of foraging when conditions favor repeated visits to the same or similar flowers, but under conditions when perseveration is unsuccessful alternative methods, such as the use of associative cues, could be beneficial.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}