Acute Administration of Caffeine: The Effect on Motor Coordination, Higher Brain Cognitive Functions, and the Social Behavior of BLC57 Mice

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Tests
2.2.1. Rotarod (RR)
2.2.2. Elevated Plus Maze Test (EPM)
2.2.3. Morris Water Maze Test (MWM)
2.2.4. Three-Chambers Social Apparatus (Crawley’s Sociability and Preference for Social Novelty Test) (3C)
3. Statistical Analysis
4. Results
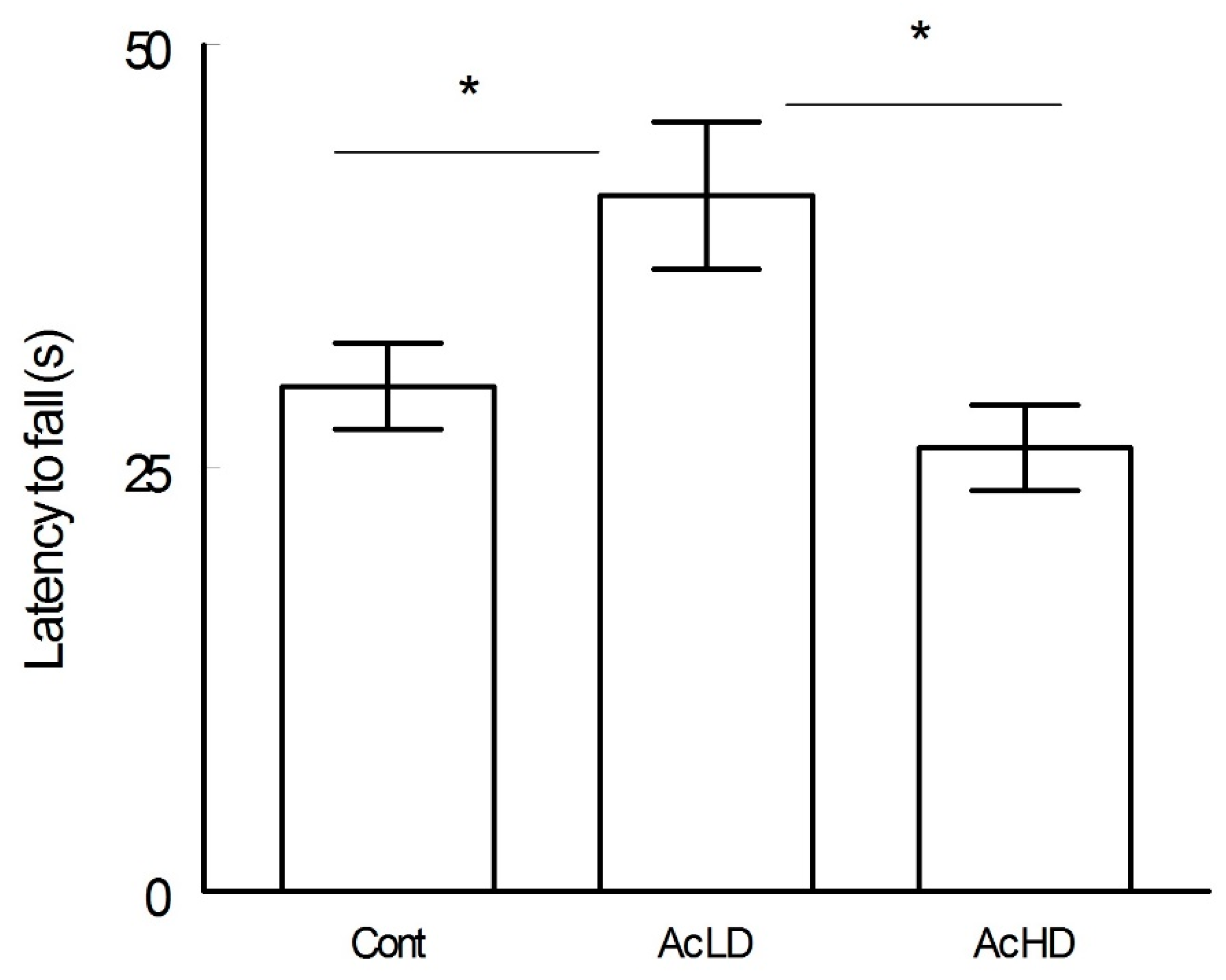
4.1. Rotarod Test: Ac MD Displayed Better Motor Coordination than the Other Groups
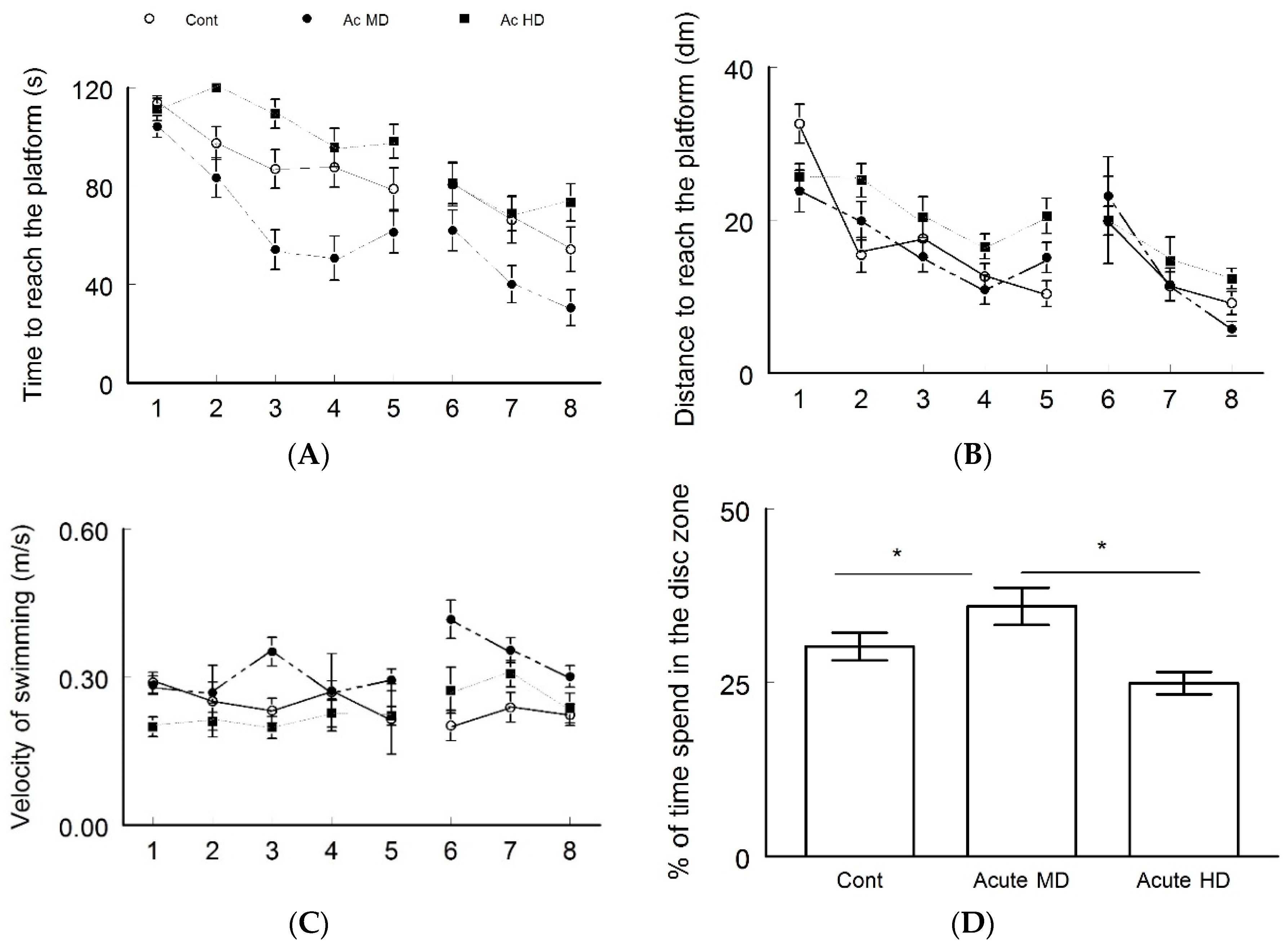
4.2. Improved Performance Displayed by Ac MD Group in Morris Water Maze Test
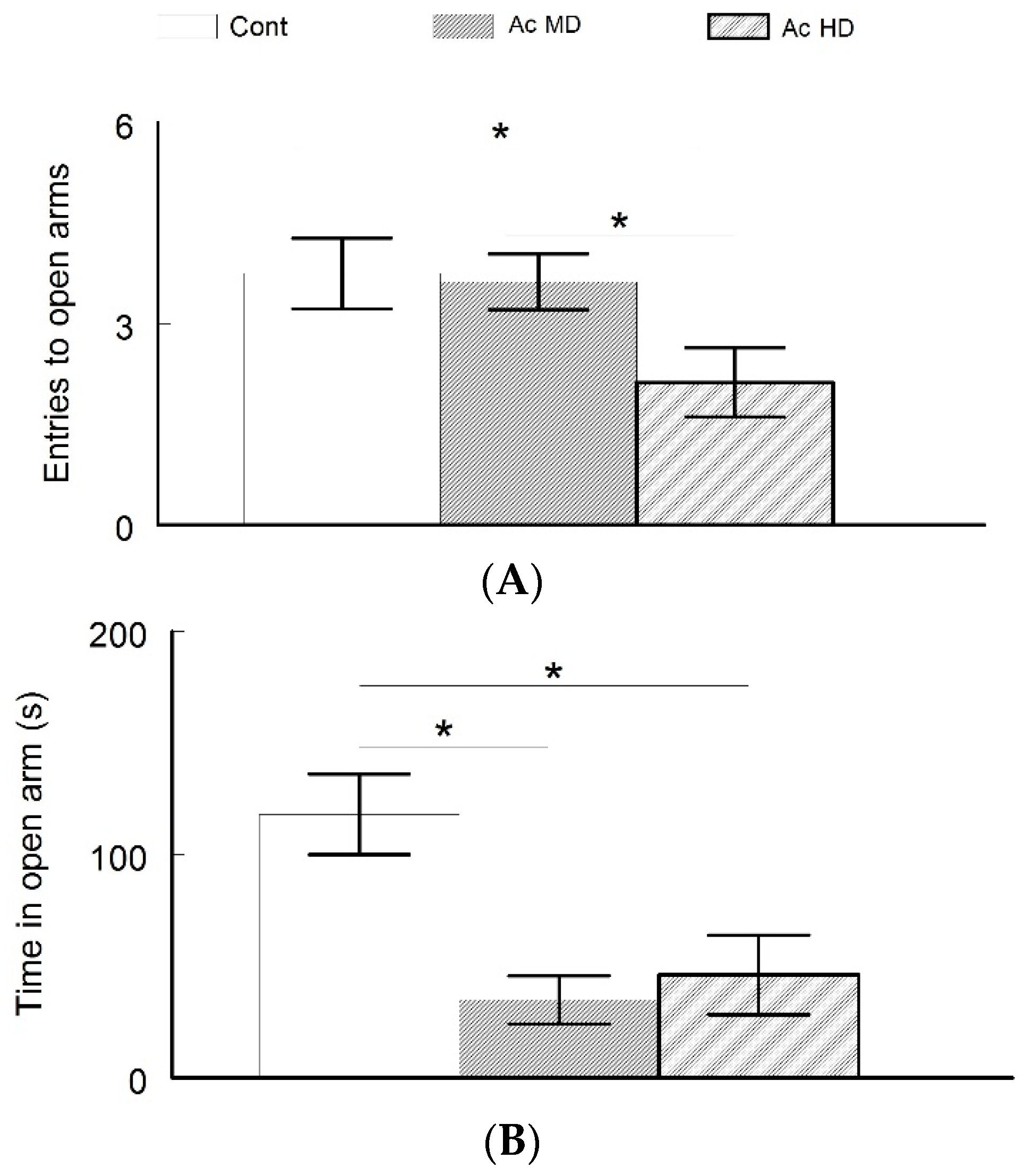
4.3. Increased Anxiety in Caffeine-Treated Mice
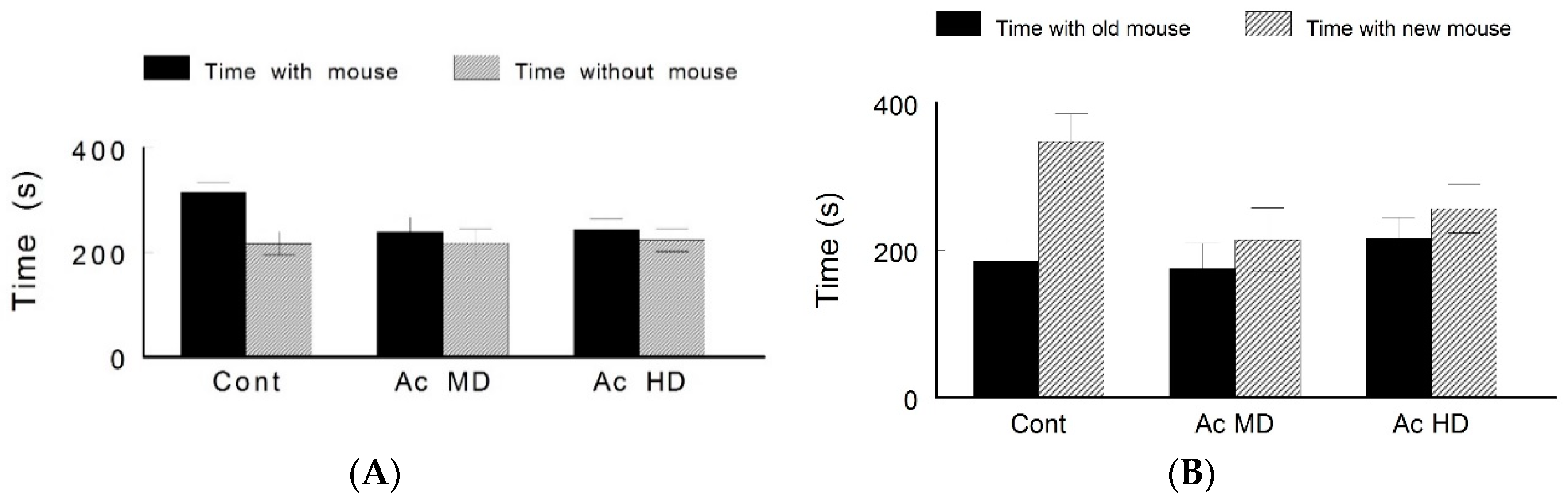
4.4. Lack of Sociability and Preference for Social Novelty in Caffeine-Treated Mice
5. Discussion and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fredholm, B.B.; Bättig, K.; Holmén, J.; Nehlig, A.; Zvartau, E.E. Actions of caffeine in the brain with special reference to factors that contribute to its widespread use. Pharmacol. Rev. 1999, 51, 83–133. [Google Scholar] [PubMed]
- Ardais, A.P.; Borges, M.F.; Rocha, A.S.; Sallaberry, C.; Cunha, R.A.; Porciúncula, L.O. Caffeine triggers behavioral and neurochemical alterations in adolescent rats. Neuroscience 2014, 270, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Bolton, S.; Null, G. Caffeine: Psychological Effects, Use and Abuse. Orthomol. Psychiatry 1981, 10, 202–211. [Google Scholar]
- Gallagher, M.; Burwell, R.; Burchinal, M. Severity of spatial learning impairment in aging: Development of a learning index for performance in the Morris water maze. Behav. Neurosci. 1993, 107, 618–626. [Google Scholar] [CrossRef] [PubMed]
- Ellenbogen, J.M.; Payne, J.D.; Stickgold, R. The role of sleep in declarative memory consolidation: Passive, permissive, active or none? Curr. Opin. Neurobiol. 2006, 16, 716–722. [Google Scholar] [CrossRef] [PubMed]
- Seugnet, L.; Galvin, J.E.; Suzuki, Y.; Gottschalk, L.; Shaw, P.J. Persistent short-term memory defects following sleep deprivation in a Drosophila model of Parkinson disease. Sleep 2009, 32, 984–992. [Google Scholar] [CrossRef] [PubMed]
- Pompili, M.; Innamorati, M.; Forte, A.; Longo, L.; Mazzetta, C.; Erbuto, D.; Ricci, F.; Palermo, M.; Stefani, H.; Seretti, M.E.; et al. Insomnia as a predictor of high-lethality suicide attempts. Int. J. Clin. Pract. 2013, 67, 1311–1316. [Google Scholar] [CrossRef] [PubMed]
- Angelucci, M.E.M.; Vital, M.A.B.F.; Cesário, C.; Zadusky, C.R.; Rosalen, P.L.; Da Cunha, C. The effect of caffeine in animal models of learning and memory. Eur. J. Pharmacol. 1999, 373, 135–140. [Google Scholar] [CrossRef]
- Ferré, S. An update on the mechanisms of the psychostimulant effects of caffeine. J. Neurochem. 2008, 105, 1067–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paterson, L.M.; Wilson, S.J.; Nutt, D.J.; Hutson, P.H.; Ivarsson, M. A translational, caffeine-induced model of onset insomnia in rats and healthy volunteers. Psychopharmacology 2007, 191, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Stein, M.B.; Stein, D.J. Social anxiety disorder. Lancet 2008, 371, 1115–1125. [Google Scholar] [CrossRef]
- Lara, D.R. Caffeine, mental health, and psychiatric disorders. J. Alzheimer’s Dis. 2010, 20 (Suppl. 1), S239–S248. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.-J.; Yoon, C.-H.; Ko, H.-J.; Kim, H.-M.; Kim, A.-S.; Moon, H.-N.; Jung, S.-P. The Relationship of Caffeine Intake with Depression, Anxiety, Stress, and Sleep in Korean Adolescents. Korean J. Fam. Med. 2016, 37, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Pechlivanova, D.M.; Tchekalarova, J.D.; Alova, L.H.; Petkov, V.V.; Nikolov, R.P.; Yakimova, K.S. Effect of long-term caffeine administration on depressive-like behavior in rats exposed to chronic unpredictable stress. Behav. Pharmacol. 2012, 23, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Watson, E.J.; Coates, A.M.; Kohler, M.; Banks, S. Caffeine consumption and sleep quality in Australian adults. Nutrients 2016, 8, 479. [Google Scholar] [CrossRef] [PubMed]
- Babson, K.A.; Feldner, M.T.; Trainor, C.D.; Smith, R.C. An Experimental Investigation of the Effects of Acute Sleep Deprivation on Panic-Relevant Biological Challenge Responding. Behav. Ther. 2009, 40, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Babson, K.A.; Trainor, C.D.; Feldner, M.T.; Blumenthal, H. A test of the effects of acute sleep deprivation on general and specific self-reported anxiety and depressive symptoms: An experimental extension. J. Behav. Ther. Exp. Psychiatry 2010, 41, 297–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; (DSM-5); American Psychiatric Publishing: Washington, DC, USA, 2013. [Google Scholar] [CrossRef]
- Wittchen, H.U. Generalized anxiety disorder: Prevalence, burden, and cost to society. Depress. Anxiety 2002, 16, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Clementz, G.L.; Dailey, J.W. Psychotropic effects of caffeine. Am. Fam. Physician 1988, 37, 167–172. [Google Scholar] [PubMed]
- Loke, W.H. Effects of caffeine on mood and memory. Physiol. Behav. 1988, 44, 367–372. [Google Scholar] [CrossRef]
- Haskell, C.F.; Kennedy, D.O.; Wesnes, K.A.; Scholey, A.B. Cognitive and mood improvements of caffeine in habitual consumers and habitual non-consumers of caffeine. Psychopharmacology 2005, 179, 813–825. [Google Scholar] [CrossRef] [PubMed]
- Lieberman, H.R.; Tharion, W.J.; Shukitt-Hale, B.; Speckman, K.L.; Tulley, R. Effects of caffeine, sleep loss, and stress on cognitive performance and mood during U.S. Navy SEAL training. Psychopharmacology 2002, 164, 250–261. [Google Scholar] [CrossRef] [PubMed]
- Komada, M.; Takao, K.; Miyakawa, T. Elevated plus maze for mice. J. Vis. Exp. 2008, e1088. [Google Scholar] [CrossRef] [PubMed]
- Onaolapo, O.J.; Onaolapo, A.Y.; Akanmu, M.A.; Olayiwola, G. Caffeine/Sleep-Deprivation interaction in mice produces complex memory effects. Ann. Neurosci. 2015, 22, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Lindner, M.D. Reliability, distribution, and validity of age-related cognitive deficits in the Morris water maze. Neurobiol. Learn. Mem. 1997, 68, 203–220. [Google Scholar] [CrossRef] [PubMed]
- Abreu, R.V.; Silva-Oliveira, E.M.; Moraes, M.F.D.; Pereira, G.S.; Moraes-Santos, T. Chronic coffee and caffeine ingestion effects on the cognitive function and antioxidant system of rat brains. Pharmacol. Biochem. Behav. 2011, 99, 659–664. [Google Scholar] [CrossRef] [PubMed]
- Kaidanovich-Beilin, O.; Lipina, T.; Vukobradovic, I.; Roder, J.; Woodgett, J.R. Assessment of social interaction behaviors. J. Vis. Exp. 2011, e2473. [Google Scholar] [CrossRef] [PubMed]
- Nikodijevic, O.; Jacobson, K.A.; Daly, J.W. Locomotor activity in mice during chronic treatment with caffeine and withdrawal. Pharmacol. Biochem. Behav. 1993, 44, 199–216. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, S.K.; Satyan, K.S.; Chakrabarti, A. Anxiogenic action of caffeine: An experimental study in rats. J. Psychopharmacol. 1997, 11, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Daly, J.W.; Shi, D.; Nikodijevic, O.; Jacobson, K.A. The role of adenosine receptors in the central action of caffeine. Pharmacopsychoecologia 1994, 7, 201–213. [Google Scholar] [PubMed]
- Daly, J. Mechanism of action of caffeine. In Caffeine, Coffee and Health; Raven Press: New York, NY, USA, 1993. [Google Scholar]
- Smith, A. Effects of caffeine on human behavior. Food Chem. Toxicol. 2002, 40, 1243–1255. [Google Scholar] [CrossRef]
- Nehlig, A.; Daval, J.L.; Debry, G. Caffeine and the central nervous system: Mechanisms of action, biochemical, metabolic and psychostimulant effects. Brain Res. Rev. 1992, 17, 139–170. [Google Scholar] [CrossRef]
- McPhersonx, P.S.; Kim, Y.K.; Valdivia, H.; Knudson, C.M.; Takekura, H.; Franzini-Armstrong, C.; Coronadot, R.; Campbell, K.P. The brain ryanodine receptor: A caffeine-sensitive calcium release channel. Neuron 1991, 7, 17–25. [Google Scholar] [CrossRef]
- Milbrandt, J. A nerve growth factor-induced gene encodes a possible transcriptional regulatory factor. Science 1987, 238, 797–799. [Google Scholar] [CrossRef] [PubMed]
- Milbrandt, J. Nerve growth factor induces a gene homologous to the glucocorticoid receptor gene. Neuron 1988, 1, 183–188. [Google Scholar] [CrossRef]
- Svenningsson, P.; le Moine, C.; Kull, B.; Sunahara, R.; Bloch, B.; Fredholm, B.B. Cellular expression of adenosine A2A receptor messenger RNA in the rat central nervous system with special reference to dopamine innervated areas. Neuroscience 1997, 80, 1171–1185. [Google Scholar] [CrossRef]
- Briley, M. Biochemical strategies in the search for cognition enhancers. Pharmacopsychiatry 1990, 23 (Suppl. 2), 75–80. [Google Scholar] [CrossRef] [PubMed]
- Carter, A.J.; O’Connor, W.T.; Carter, M.J.; Ungerstedt, U. Caffeine enhances acetylcholine release in the hippocampus in vivo by a selective interaction with adenosine A1 receptors. J. Pharmacol. Exp. Ther. 1995, 273, 637–642. [Google Scholar] [PubMed]
- Morton, R.A.; Davies, C.H. Regulation of muscarinic acetylcholine receptor-mediated synaptic responses by adenosine receptors in the rat hippocampus. J. Physiol. 1997, 502, 75–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Everitt, B.J.; Robbins, T.W. Central cholinergic systems and cognition. Annu. Rev. Psychol. 1997, 48, 649–684. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.J.; Li, W.; Chen, J.F. Normal and abnormal functions of adenosine receptors in the central nervous system revealed by genetic knockout studies. Biochim. Biophys. Acta Biomembr. 2011, 1808, 1358–1379. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.F. Adenosine receptor control of cognition in normal and disease. Int. Rev. Neurobiol. 2014, 119, 257–307. [Google Scholar] [CrossRef] [PubMed]
- Angelucci, M.E.; Cesario, C.; Hiroi, R.H.; Rosalen, P.L.; da Cunha, C. Effects of caffeine on learning and memory in rats tested in the Morris water maze. Braz. J. Med. Biol. Res. 2002, 35, 1201–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, N.J.; Simões, A.P.; Silva, H.B.; Ardais, A.P.; Kaster, M.P.; Garção, P.; Rodrigues, D.I.; Pochmann, D.; Santos, A.I.; Araújo, I.M.; et al. Caffeine Reverts Memory But Not Mood Impairment in a Depression-Prone Mouse Strain with Up-Regulated Adenosine A2A Receptor in Hippocampal Glutamate Synapses. Mol. Neurobiol. 2017, 54, 1552–1563. [Google Scholar] [CrossRef] [PubMed]
- Hughes, R.N.; Hancock, N.J. Effects of acute caffeine on anxiety-related behavior in rats chronically exposed to the drug, with some evidence of possible withdrawal-reversal. Behav. Brain Res. 2017, 321, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Rainnie, D.; Grunze, H.; McCarley, R.; Greene, R. Adenosine inhibition of mesopontine cholinergic neurons: Implications for EEG arousal. Science 1994, 263, 689–692. [Google Scholar] [CrossRef] [PubMed]
- Cunha, R.A.; Ferre, S.; Vaugeois, J.M.; Chen, J.F. Potential therapeutic interest of adenosine A2A receptors in psychiatric disorders. Curr. Pharm. Des. 2008, 14, 1512–1524. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, H.A.; File, S.E. Caffeine-induced anxiogenesis: The role of adenosine, benzodiazepine and noradrenergic receptors. Pharmacol. Biochem. Behav. 1989, 32, 181–186. [Google Scholar] [CrossRef]
- Hilakivi, L.A.; Durcan, M.J.; Lister, R.G. Effects of caffeine on social behavior, exploration and locomotor activity: Interactions with ethanol. Life Sci. 1989, 44, 543–553. [Google Scholar] [CrossRef]
- Prediger, R.D.S.; Batista, L.C.; Takahashi, R.N. Adenosine A1 receptors modulate the anxiolytic-like effect of ethanol in the elevated plus-maze in mice. Eur. J. Pharmacol. 2004, 499, 147–154. [Google Scholar] [CrossRef] [PubMed]
- File, S.E.; Baldwin, H.A.; Johnston, A.L.; Wilks, L.J. Behavioral effects of acute and chronic administration of caffeine in the rat. Pharmacol. Biochem. Behav. 1988, 30, 809–815. [Google Scholar] [CrossRef]
- Lopez, F.; Miller, L.G.; Greenblatt, D.J.; Kaplan, G.B.; Shader, R.I. Interaction of caffeine with the GABAAreceptor complex: Alterations in receptor function but not ligand binding. Eur. J. Pharmacol. Mol. Pharmacol. 1989, 172, 453–459. [Google Scholar] [CrossRef]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almosawi, S.; Baksh, H.; Qareeballa, A.; Falamarzi, F.; Alsaleh, B.; Alrabaani, M.; Alkalbani, A.; Mahdi, S.; Kamal, A. Acute Administration of Caffeine: The Effect on Motor Coordination, Higher Brain Cognitive Functions, and the Social Behavior of BLC57 Mice. Behav. Sci. 2018, 8, 65. https://doi.org/10.3390/bs8080065
Almosawi S, Baksh H, Qareeballa A, Falamarzi F, Alsaleh B, Alrabaani M, Alkalbani A, Mahdi S, Kamal A. Acute Administration of Caffeine: The Effect on Motor Coordination, Higher Brain Cognitive Functions, and the Social Behavior of BLC57 Mice. Behavioral Sciences. 2018; 8(8):65. https://doi.org/10.3390/bs8080065
Chicago/Turabian StyleAlmosawi, Sayed, Hasan Baksh, Abdulrahman Qareeballa, Faisal Falamarzi, Bano Alsaleh, Mallak Alrabaani, Ali Alkalbani, Sadiq Mahdi, and Amer Kamal. 2018. "Acute Administration of Caffeine: The Effect on Motor Coordination, Higher Brain Cognitive Functions, and the Social Behavior of BLC57 Mice" Behavioral Sciences 8, no. 8: 65. https://doi.org/10.3390/bs8080065