Wildlife Habitat Quality (Sward Structure and Ground Cover) Response of Mixed Native Warm-Season Grasses to Harvesting

 , ,
, ,
Abstract
:1. Introduction
1.1. Sward Structure for Ground-Nesting Birds
1.2. Justification and Objectives
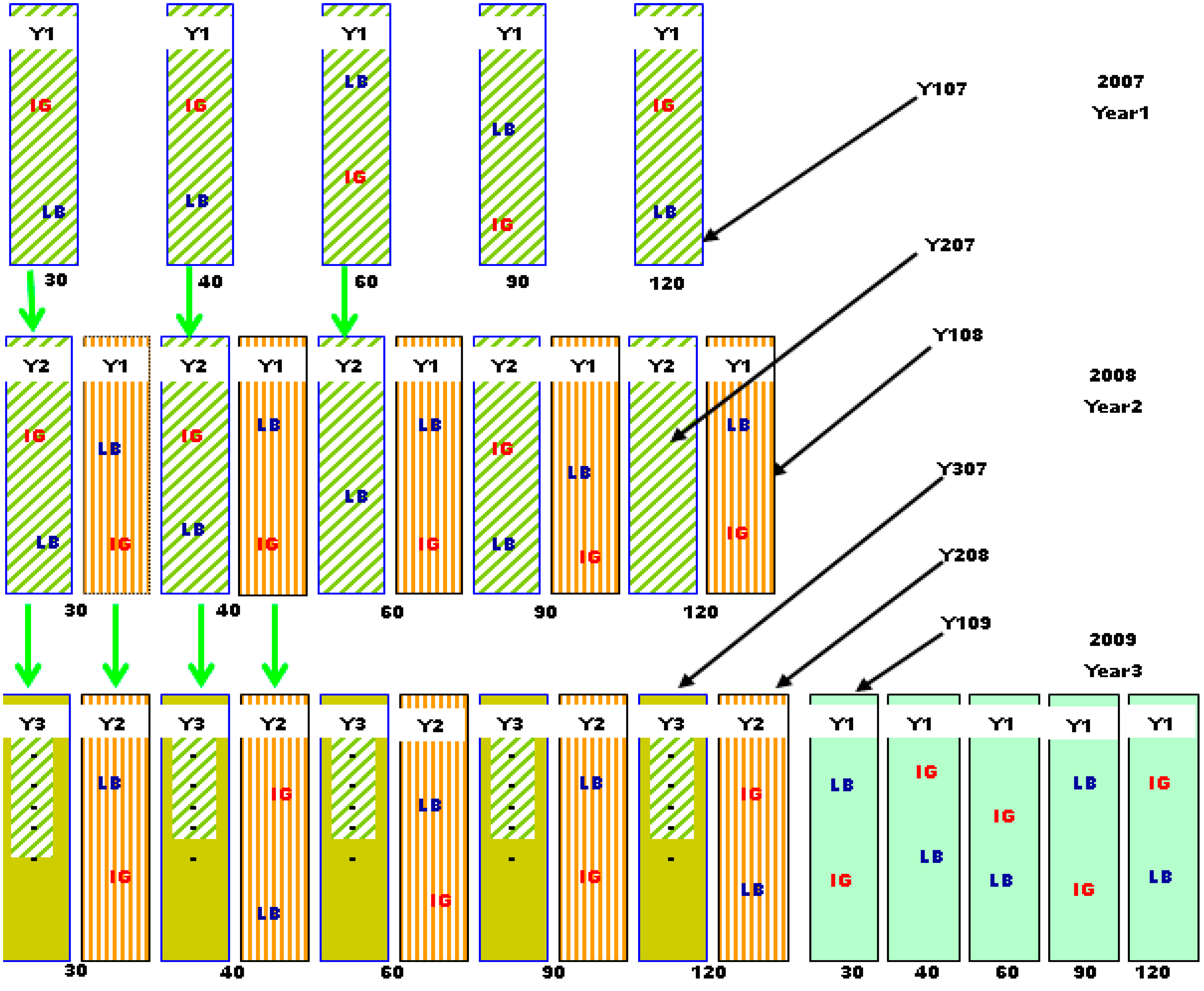
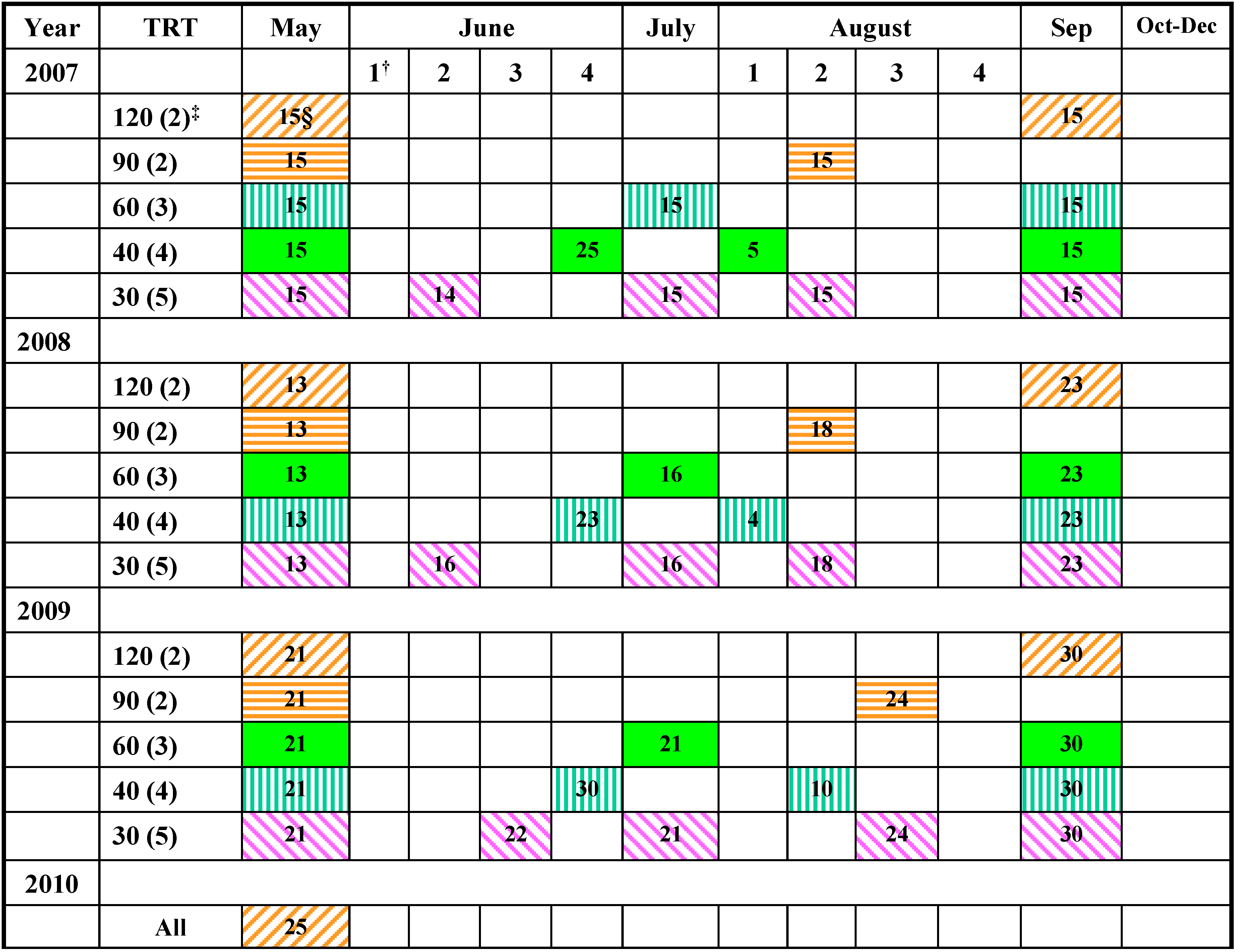
2. Experimental Section
2.1. Location and Field Layout


2.2. Data Analyses
3. Results and Discussion
3.1. Harvesting Effects on Ground Cover
3.1.1. Cover by Vegetation

{kind=link}
{kind=link}
{kind=link}
| Treatment | Cover Attributes | ||||
|---|---|---|---|---|---|
| Grass | Forb | Litter | Bare Ground | ||
 | |||||
| Harvest interval | |||||
| Year 2009 | |||||
| Control | 38 | 26 | 36a ¶ | 1c | |
| 120(2) § | 37 | 21 | 14b | 29a | |
| 90(2) | 39 | 21 | 17b | 23ab | |
| 60(3) | 44 | 19 | 16b | 21b | |
| 40(4) | 44 | 17 | 14b | 25ab | |
| 30(5) | 41 | 19 | 13b | 26ab | |
| Pr > α # | 0.29 | 0.33 | <0.01 | <0.01 | |
| Year 2010 | |||||
| Control | 57c | 25a | 7a | 10ab | |
| 120(2) § | 71b | 15b | 3c | 11a | |
| 90(2) | 76ab | 15b | 3bc | 6b | |
| 60(3) | 75ab | 13b | 4b | 8ab | |
| 40(4) | 77a | 12b | 4bc | 7b | |
| 30(5) | 78a | 12b | 3bc | 7b | |
| Pr > α | <0.01 | <0.01 | <0.01 | 0.02 | |
| Harvest duration | |||||
| Year 2009 | |||||
| Control (Y109 ††) | 38b | 26 | 36a | 1b | |
| Y208 | 36b | 20 | 17b | 27a | |
| Y307 | 46a | 19 | 13b | 22a | |
| Pr > α | <0.01 | 0.12 | <0.01 | <0.01 | |
| Year 2010 | |||||
| Control | 57c | 25a | 7a | 10 | |
| Y209 | 72b | 15b | 4b | 9 | |
| Y308 | 79a | 12b | 3b | 6 | |
| Y407 | 75ab | 12b | 4b | 9 | |
| Pr > α | <0.01 | <0.01 | <0.01 | 0.07 | |

3.1.2. Ground Cover by Litter
3.1.3. Bare-Ground Space
3.2. Harvest Duration Effects on Ground Cover
3.2.1. Harvest Duration Effects in 2009
3.2.2. Harvest Duration Effects in 2010
3.3. Sward Heights
| Harvest Interval | May 2009 | May 2010 | ||||
|---|---|---|---|---|---|---|
| Y109 § | Y208 | Y307 | Y209 | Y308 | Y407 | |
| Sward Heights | ||||||
 | ||||||
| Control | 58 | 58a ¶ | 58a | 45a | 45a | 45 |
| 120(2) # | 56 | 41b | 41b | 29b | 28b | 30 |
| 90(2) | 53 A | 38b B | 35b B | 28b | 29b | 31 |
| 60(3) | 52 A | 39b AB | 34b B | 28b | 28b | 34 |
| 40(4) | 54 A | 35b B | 32b B | 27b | 25b | 32 |
| 30(5) | 56 A | 32b B | 32b B | 28b | 25b | 32 |
| Pr > α †† | 0.91 | 0.02 | <0.01 | <0.01 | <0.01 | 0.06 |
| Light interception | ||||||
 | ||||||
| Control | 61 | 61a | 61a | 62 | 62 | 62 |
| 120(2) | 58 A | 33b B | 40b B | 46 | 42 | 48 |
| 90(2) | 63 A | 34b B | 36b B | 55 | 45 | 45 |
| 60(3) | 63 A | 36b B | 40b B | 53 | 44 | 45 |
| 40(4) | 56 A | 32b B | 31b B | 56 | 44 | 42 |
| 30(5) | 54 A | 32b B | 37b B | 53 | 42 | 44 |
| Pr > α | 0.64 | <0.01 | <0.01 | 0.90 | 0.52 | 0.72 |
3.4. Sward Canopy Closure
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wilson, J.D.; Whittingham, M.J.; Bradbury, R.B. The management of crop structure: A general approach to reversing the impacts of agricultural intensification on birds. IBIS 2005, 47, 453–463. [Google Scholar]
- Knapp, A.K.; Seastedt, T.R. Detritus accumulation limits productivity of tallgrass prairie. BioScience 1986, 36, 662–668. [Google Scholar] [CrossRef]
- Shea, A.B. The Return of Native Grasses to Tennessee. 1999. Available online: http://www.tn.gov/environment/conservationist/archive/grass.htm (accessed on 12 August 2009). [Google Scholar]
- Atkinson, P.W.; Buckingham, D.; Morris, A.J. What factors determine where invertebrate-feeding birds forage in dry agricultural grasslands? IBIS 2004, 146, 99–107. [Google Scholar] [CrossRef]
- Brenna, L.A. How can we reverse the northern bobwhite population declines? Wildl. Soc. Bull. 1991, 19, 544–555. [Google Scholar]
- Riffell, S.; Scognamillo, D.; Burger, L.W. Effects of the Conservation Reserve Program on northern bobwhite and grassland birds. Environ. Monit. Assess. 2008, 146, 309–323. [Google Scholar] [CrossRef]
- Harper, C.A.; Moorman, C.E. Qualifying native warm-season grasses and early succession habitat. In Proceedings of the 11th Triennial National Wildlife and Fisheries Extension Specialists Conference, Big Sky, MT, USA, 14–18 October 2006; Timm, R.M., Harper, C.A., Higginbotham, B.J., Parkhurst, J.A., Eds.; p. 10.
- White, B.; Graham, P.; Pierce, R.A., II. Missouri Bobwhite Quail. Habitat Appraisal Guide: Assessing Your Farm’s Potential for Bobwhites. 2005. Available online: http://extension.missouri.edu/explorepdf/miscpubs/mp0902.pdf (accessed on 25 October 2008).
- Lusk, J.J.; Smith, S.G.; Fuhlendorf, S.D.; Guthery, F.S. Factors influencing northern bobwhite nest-site selection and fate. J. Wildlife. Manag. 2006, 70, 564–571. [Google Scholar] [CrossRef]
- Jones, J.; Coggin, D.S.; Cummins, J.L.; Hill, J. Restoring and Managing Native Prairies. A Handbook for Mississippi Landowners; Wildlife Mississippi: Starkville, MS, USA, 2007. [Google Scholar]
- McCoy, T.D.; Kurzejeski, E.W.; Burger, L.W., Jr.; Ryan, M.R. Effects of conservation practice, mowing, and temporal changes on vegetation structure on CRP fields in northern Missouri. Wildl. Soc. Bull. 2001, 29, 979–987. [Google Scholar]
- Greenfield, K.C.; Burger, L.W., Jr.; Chamberlain, M.J.; Kurzejeski, E.W. Vegetation management practices on conservation reserve program fields to improve northern bobwhite habitat quality. Wildl. Soc. Bull. 2002, 30, 527–538. [Google Scholar]
- Greenfield, K.C.; Chamberlain, M.J.; Burger, L.W., Jr.; Kurzejeski, E.W. Effects of burning and discing Conservation Reserve Program fields to improve habitat quality for northern bobwhite (Colinus virginianus). Am. Midl. Nat. 2003, 149, 344–353. [Google Scholar] [CrossRef]
- Helzer, C. Using Defoliation of Dominant Grasses to Increase Prairie Plant Diversity. 2011. Available online: http://prairieecologist.com/2011/03/15/using-defoliation-of-dominant-grasses-to-increase-prairie-plant-diversity (accessed on 25 March 2011).
- Ferraro, D.O.; Oesterheld, M. Effect of defoliation on grass growth. A quantitative review. Oikos 2002, 98, 125–133. [Google Scholar] [CrossRef]
- Alexander, K.I.; Thompson, K. The effect of clipping frequency on the competitive interaction between two perennial grass species. Oecologia (Berlin) 1982, 53, 251–254. [Google Scholar]
- Woodis, J.E.; Jackson, R.D. The effects of clipping height and frequency on net primary production of Andropogon gerardii (C4 grass) and Bromus inermis (C3 grass) in greenhouse experiments. Grass Forage Sci. 2008, 63, 458–466. [Google Scholar] [CrossRef]
- Bonham, C.D.; Mergen, D.E.; Montoya, S. Plant cover estimation: A contiguous daubenmire frame. Rangelands 2004, 26, 17–22. [Google Scholar] [CrossRef]
- Pedreira, C.G.S.; Sollenberger, L.E.; Mislevy, P. Botanical composition, light interception, and carbohydrate reserve status of grazed ‘Florakirk’ Bermudagras. Agron. J. 2000, 92, 194–199. [Google Scholar] [CrossRef]
- SAS Institute. SAS Version 9.2, SAS Institute: Cary, NC, USA, 2007.
- Burger, L.W., Jr.; Ryan, M.R.; Kurzejeski, E.W.; Dailey, T.V. Factors affecting the habitat value of conservation reserve program lands for northern bobwhite in northern Missouri. In Proceedings of the NCT-163 Post Conservation Reserve Land Use Conference, Denver, CO, USA, 10–11 January 1994; Dicks, M., Monsoon, M., Eds.; pp. 142–156.
- Manske, L.L. Biologically Effective Grazing Management. 2003. Available online: http://www.ag.ndsu.edu/archive/dickinso/research/2003/range03a.htm (accessed on 10 July 2008).
- Damhoureyeh, S.A.; Hartnett, D.C. Effects of bison and cattle on growth, reproduction, and abundances of five tallgrass prairie forbs. Am. J. Bot. 1997, 84, 1719. [Google Scholar] [CrossRef]
- Hubbard, J.A. Fire-Grazing Interactions in a Mixed Grass Prairie. Ph.D. Dissertation, Texas A&M University, College Station, TX, USA, 2003. [Google Scholar]
- Bremer, D.J.; Ham, J.M. Effect of spring burning on the surface energy balance in a tallgrass prairie. Agric. For. Meteorol. 1999, 97, 43–54. [Google Scholar] [CrossRef]
- Carver, A.V.; Burger, L.W., Jr.; Palmer, W.P.; Brennan, L.A. Vegetation characteristics in seasonal-disked fields and at bobwhite brood locations. In Proceedings of the Annual Conference Southeastern Association Fish and Wild Agencies, Louisville, KY, USA, 28 October–1 November 2001; Eversole, A.G., Ed.; pp. 436–444.
- Temu, V.W.; Rude, B.J.; Baldwin, B.S. Yield response of native warm-season forage grasses to harvest intervals and durations in mixed stands. Agronomy 2014, 4, 90–107. [Google Scholar] [CrossRef]
- Briske, D.D. Plant response to defoliation: Morphological Considerations and Allocation Priorities. In Rangelands: A Resource Under Siege; Joss, P.J., Lynch, P.W., Williams, O.B., Eds.; Cambridge University Press: Cambridge, UK, 1986; pp. 425–427. [Google Scholar]
- Briske, D.D.; Richards, J.H. Plant Responses to Defoliation: A Physiological, Morphological and Demographic Evaluation. In Wildland Plants: Physiological Ecology and Developmental Morphology; Bedunah, D.J., Sosebee, R.E., Eds.; Society for Range Management: Denver, CO, USA, 1995; pp. 635–710. [Google Scholar]
- Michalk, D.L.; Herbert, P.K. Assessment of four techniques for estimating yield on dryland pastures. Agron. J. 1977, 69, 864–868. [Google Scholar] [CrossRef]
- Harmoney, K.R.; Moore, K.J.; George, J.R.; Brummer, E.C.; Russell, J.R. Determination of pasture biomass using four indirect methods. Agron. J. 1997, 89, 665–672. [Google Scholar] [CrossRef]
- Jatimliansky, J.R.; Gimenez, D.O.; Bujan, A. Herbage yield, tiller number and root system activity after defoliation of prairie grass (Bromus catharticus Vahl). Grass Forage Sci. 1997, 52, 52–62. [Google Scholar]
- Richards, J.H. Root growth response to defoliation in two Agropyron bunchgrasses: Field observations with an improved root periscope. Oecologia (Berlin) 1984, 64, 21–25. [Google Scholar] [CrossRef]
- McNaughton, S.J. Laboratory-simulated grazing: Interactive effects of defoliation and canopy closure on Serengeti grasses. Ecology 1992, 73, 170–182. [Google Scholar] [CrossRef]
- Palmer, W.E. Effects of Modern Pesticides and Farming Systems on Northern Bobwhite Quail Brood Ecology. Ph.D. Dissertation, North Carolina State University, Raleigh, NC, USA, 1995. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Temu, V.W.; Baldwin, B.S.; Reddy, K.R.; Riffell, S.; Burger, L.W. Wildlife Habitat Quality (Sward Structure and Ground Cover) Response of Mixed Native Warm-Season Grasses to Harvesting. Environments 2014, 1, 75-91. https://doi.org/10.3390/environments1010075
Temu VW, Baldwin BS, Reddy KR, Riffell S, Burger LW. Wildlife Habitat Quality (Sward Structure and Ground Cover) Response of Mixed Native Warm-Season Grasses to Harvesting. Environments. 2014; 1(1):75-91. https://doi.org/10.3390/environments1010075
Chicago/Turabian StyleTemu, Vitalis W., Brian S. Baldwin, K. Raja Reddy, Samuel Riffell, and Loren W. Burger. 2014. "Wildlife Habitat Quality (Sward Structure and Ground Cover) Response of Mixed Native Warm-Season Grasses to Harvesting" Environments 1, no. 1: 75-91. https://doi.org/10.3390/environments1010075