
Classical Optimization of Cellulase and Xylanase Production by a Marine Streptomyces Species


Abstract
:1. Introduction
2. Materials and Methods
2.1. Source of Bacteria
2.2. Identification of the Bacterial Isolate by 16S rDNA
2.3. Media Composition and Fermentation Conditions
2.4. Determination of Optimal Parameters for Enzyme Production
2.4.1. Effect of Initial pH on Cellulase and Xylanase Production
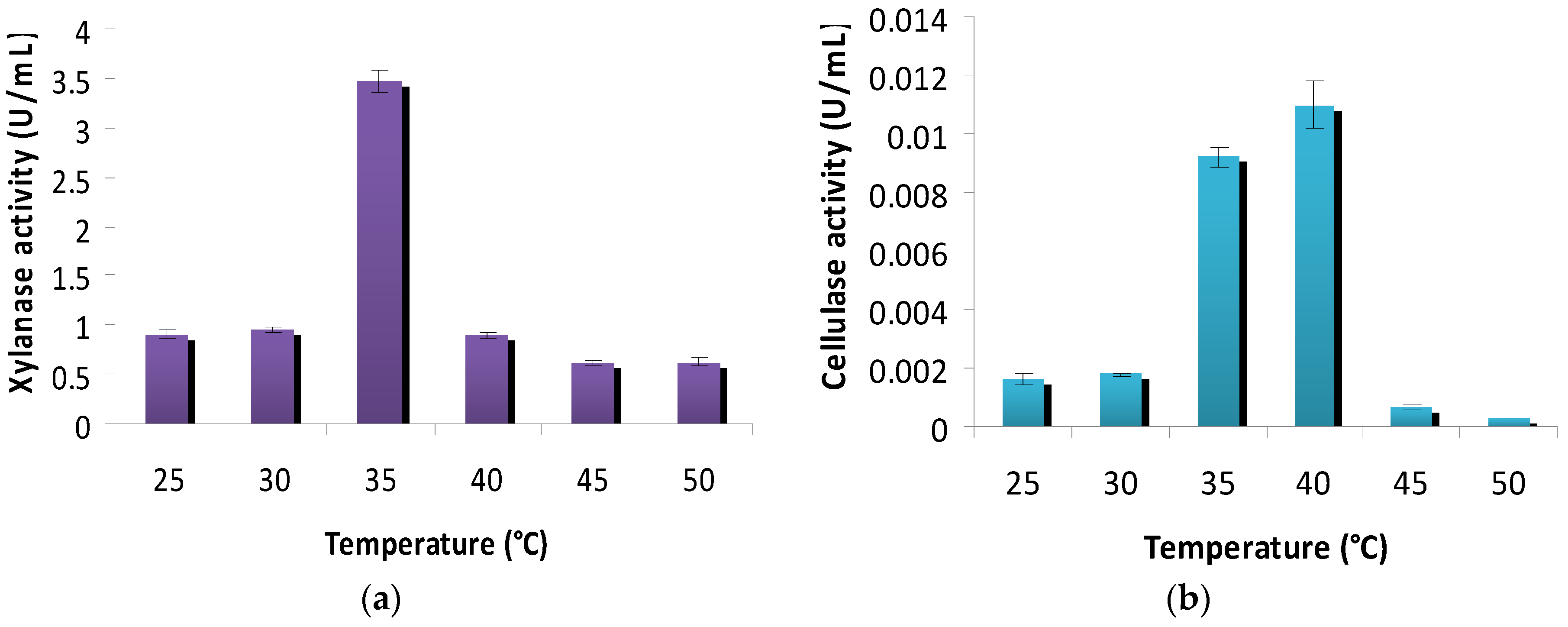
2.4.2. Effect of Incubation Temperature on Cellulase and Xylanase Production
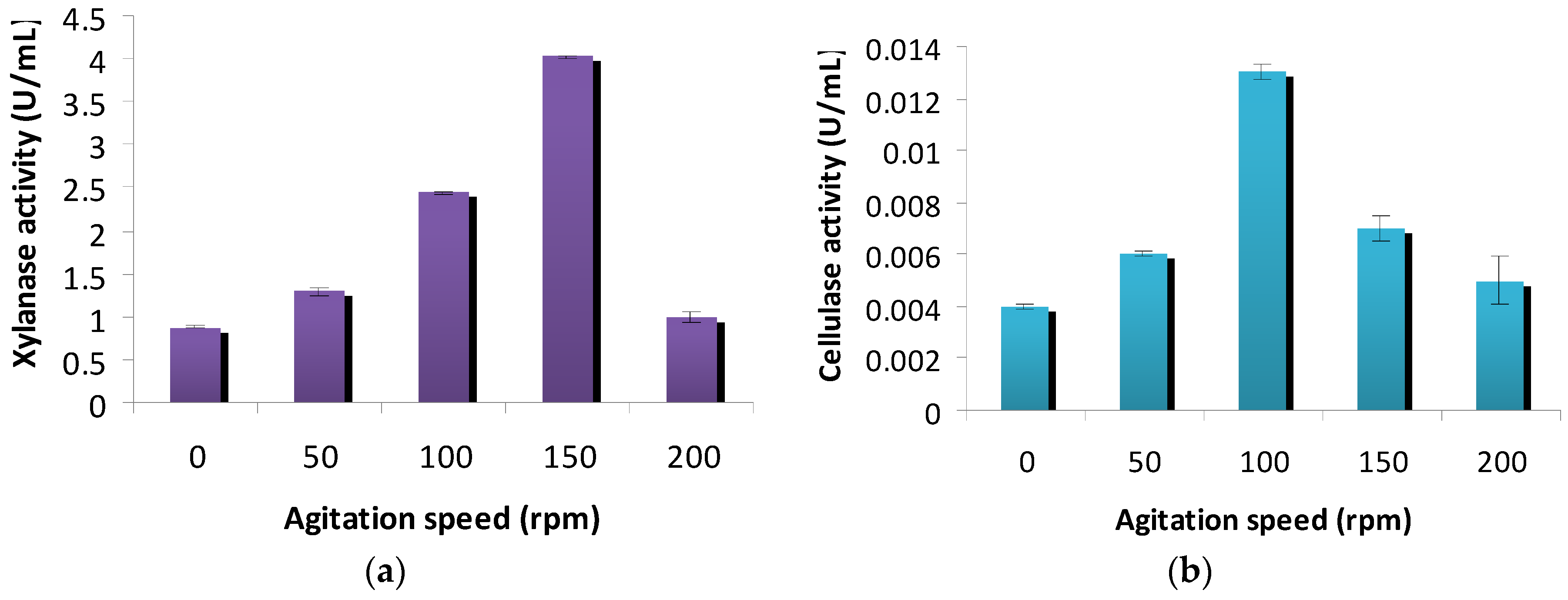
2.4.3. Effect of Agitation Speed on Cellulase and Xylanase Production
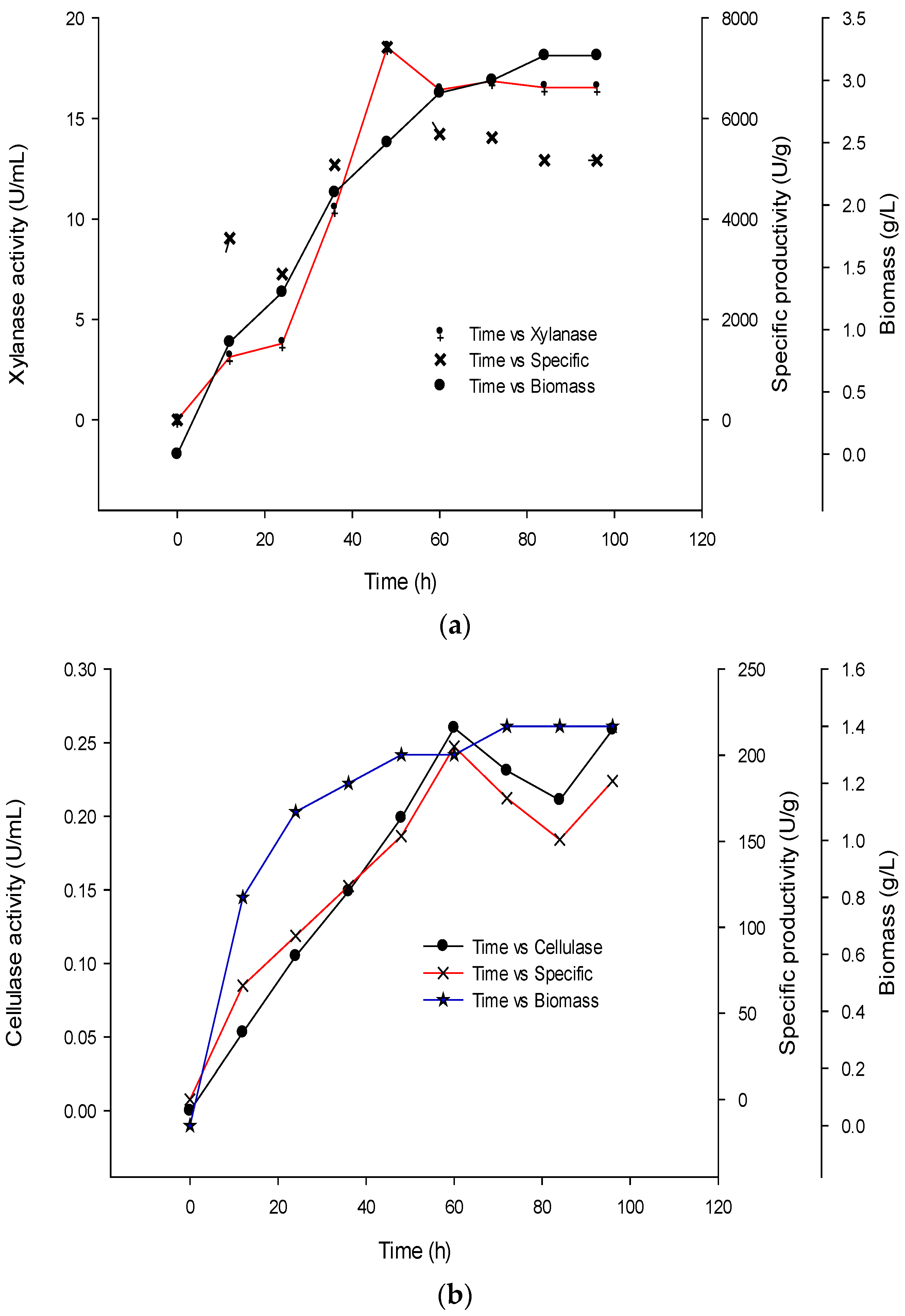
2.5. Time Course and Growth Kinetic for Production of Cellulase and Xylanase
2.6. Inducing Effect of Some Mono- and Polysaccharides on Enzyme Production
2.7. Enzyme Assay
2.8. Statistical Analysis
3. Results
3.1. Identification of Bacterial Strain
3.2. Effect of Initial pH of Fermentation Medium on Cellulase and Xylanase Production
3.3. Effect of Incubation Temperature on Cellulase and Xylanase Production
3.4. Effect of Agitation on Cellulase and Xylanase Production
3.5. Time Course and Kinetics for Production of Cellulase and Xylanase
3.6. Inducing Effect of Carbon Source Inducers on Holocellulase Production
4. Discussion
4.1. Effect of Initial pH of Fermentation Medium on Cellulase and Xylanase Production
4.2. Effect of Incubation Temperature on Xylanase and Cellulase Production
4.3. Effect of Agitation on Cellulase and Xylanase Production
4.4. Time Course and Kinetics for Production of Cellulase and Xylanase
4.5. Effect of Carbon Source Inducers on Cellulase and Xylanase Production
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Maki, M.L.; Idrees, A.; Leung, K.T.; Qin, W. Newly Isolated and Characterized Bacteria with Great Application Potential for Decomposition of Lignocellulosic Biomass. J. Mol. Microbiol. Biotechnol. 2012, 22, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Howard, R.R.; Abotsi, E.; Jansen van Renshurg, E.L.; Howard, S. Lignocellulose biotechnology: Issues of bioconversion and enzyme production. Afr. J. Biotechnol. 2003, 2, 602–619. [Google Scholar] [CrossRef]
- Kuhad, R.C.; Gupta, R.; Singh, A. Microbial cellulases and their industrial applications. Enzym. Res. 2011, 2011, 280696. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, H.M.N.; Asgher, M.; Ahmed, I.; Hussain, S. Media optimization for Hyper-production of carboxymethyl cellulase using proximally analyzed agro-industrial residue with Trichoderma harzianum under SSF. IJAVMS 2013, 4, 47–55. [Google Scholar]
- Agbor, V.B.; Cicek, N.; Sparling, R.; Berlin, A.; Levin, D.B. Biomass pretreatment: Fundamentals toward application. Biotechnol. Adv. 2011, 29, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Shallom, D.; Shoham, Y. Microbial hemicellulases. Curr. Opin. Microbiol. 2003, 6, 219–228. [Google Scholar] [CrossRef]
- Fontes, C.M.; Gilbert, H.J.; Hazlewood, G.P.; Clarke, J.H.; Prates, J.A.M.; McKie, V.A.; Nagy, T.; Fernandes, T.H.; Ferreira, L.M. A novel Cellvibrio mixtus family 10 xylanase that is both intracellular and expressed under non-inducing conditions. Microbiology 2000, 146, 1959–1967. [Google Scholar] [CrossRef] [PubMed]
- Viikari, L.; Vehmaanpera, J.; Koivula, A. Lignocellulosic ethanol: From science to industry. Biomass Bioeng. 2012, 46, 13–24. [Google Scholar] [CrossRef]
- Banerjee, G.; Scott-Craig, S.J.; Walton, D.J. Improving enzymes for biomass conversion: A basic research perspective. Bioenerg. Res. 2010, 3, 82–92. [Google Scholar] [CrossRef]
- Biswas, R. Production of cellulolytic enzymes. In Bioprocessing of Renewable Resources to Commodity Bioproducts, 1st ed.; Bisaria, V.S., Kondo, A., Eds.; John Wiley & Sons, Inc.: New York, NY, USA, 2014; pp. 105–132. [Google Scholar]
- Ray, A.K.; Bairagi, A.; Ghosh, K.S.; Sen, S.K. Optimization of fermentation conditions for cellulase production by Bacillus subtilis CY5 and Bacillus circulans TP3 isolated from fish gut. Acta Ichthyol. Piscat. 2007, 3, 47–53. [Google Scholar] [CrossRef]
- Ritter, C.E.T.; Camassola, M.; Zampieri, D.; Silveira, M.M.; Dillon, A.J.P. Cellulase and xylanase production by Penicillium echinulatum in submerged media containing cellulose amended with sorbitol. Enzym. Res. 2013, 9, 240219. [Google Scholar] [CrossRef]
- Adhyaru, N.D.; Bhatt, S.N.; Modi, A.H. Optimization of upstream and downstream process parameters for cellulase-poor-thermosolvent-stable xylanase production by Apergillus tubingensis FDHN1. Bioresour. Bioprocess. 2015, 2, 1–14. [Google Scholar] [CrossRef]
- Okeke, C.B.; Lu, J. Characterization of a defined cellulolytic and xylanolytic bacterial consortium for bioprocessing of cellulose and hemicelluloses. J. Appl. Biochem. 2011, 163, 869–881. [Google Scholar] [CrossRef] [PubMed]
- Dionisi, H.M.; Lozada, M.; Olivera, N.L. Bioprospection of marine microorganisms: Biotechnological applications and methods. Rev. Argent. Microbiol. 2012, 44, 49–60. [Google Scholar]
- Ekborg, N.A.; Gonzalez, J.M.; Howard, M.B.; Taylor, L.E.; Hutcheson, S.W.; Weiner, R.M. Saccharophagus degradans gen. nov., sp. nov., a versatile marine degrader of complex polysaccharides. Int. J. Syst. Evol. Microbiol. 2005, 55, 1545–1549. [Google Scholar] [CrossRef] [PubMed]
- Michel, G.; Czjzek, M. Polysaccharide-degrading enzymes from marine bacteria. In Marine Enzymes for Biocatalysis: Sources, Biocatalytic Characteristics and Bioprocesses of Marine Enzymes; Trincone, A., Ed.; Woodhead Publishing: Oxford, UK, 2013; pp. 429–464. [Google Scholar]
- Adhi, T.P.; Korus, R.A.; Crawford, D.L. Production of major extracellular enzymes during lignocellulose degradation by two Streptomycetes in agitated submerged culture. Appl. Environ. Microbiol. 1989, 55, 1165–1168. [Google Scholar] [PubMed]
- Trincone, A. Marine biocatalysts: Enzymatic features and applications. Mar. Drugs 2011, 9, 478–499. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie, C.R.; Bilous, D.; Schneider, H.; Johnson, K.G. Induction of cellulolytic and xylanolytic enzyme systems in Streptomyces spp. Appl. Environ. Microbiol. 1987, 53, 2835–2839. [Google Scholar] [PubMed]
- Herai, S.; Hashimoto, Y.; Higashibata, H.; Maseda, H.; Ikeda, H.; Omura, S.; Kobayashi, M. Hyper-inducible expression system for streptomycetes. Proc. Natl. Acad. Sci. USA 2004, 101, 14031–14035. [Google Scholar] [CrossRef] [PubMed]
- Mincer, T.J.; Jensen, P.R.; Kauffman, C.A.; Fenical, W. Widespread and Persistent populations of a major New Marine Actinomycete Taxon in Ocean Sediments. Appl. Environ. Microbiol. 2002, 68, 5005–5011. [Google Scholar] [CrossRef] [PubMed]
- Cosa, S.; Mabinya, L.V.; Ademola, O.; Olaniran, A.O.; Okoh, A.I. Production and characterization of bioflocculant produced by Halobacillus sp. Mvuyo isolated from bottom sediment of Algoa Bay. Environ. Technol. 2011, 33, 967–973. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Kumar, V.; Naik, B.; Bisht, G.S. Screening of xylanase activity of Streptomyces albidoflavus PSM-3n isolated from Uttarakhand. Eurasia J. Biosci. 2013, 7, 30–40. [Google Scholar] [CrossRef]
- Battan, B.; Sharma, J.; Dhiman, S.S.; Kuhad, R.C. Enhanced production of cellulase-free thermostable xylanase by Bacillus pumilus ASH and its potential application in paper industry. Enzym. Microb. Technol. 2007, 41, 733–739. [Google Scholar] [CrossRef]
- Yassien, M.A.-M.; Jiman-Fatani, A.A.M.; Asfour, H.Z. Production, purification and characterization of cellulase from Streptomyces sp. Afr. J. Microbiol. Res. 2014, 8, 348–354. [Google Scholar]
- Godden, B.; Legon, T.; Helvenstein, P.; Penninck, M. Regulation of the production of hemicellulolytic and cellulolytic enzymes by a Streptomyces sp. growing on lignocellulose. J. Gen. Microbiol. 1989, 135, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Saha, B.C.; Iten, L.B.; Cotta, M.A.; Wu, Y.V. Dilute acid pretreatment, enzymatic saccharification and fermentation of wheat straw to ethanol. Proc. Biochem. 2005, 40, 3693–3700. [Google Scholar] [CrossRef]
- Saratale, G.D.; Saratale, R.G.; Oh, S.E. Production and characterization of multiple cellulolytic enzymes by isolated Streptomyces sp. MDS. Biomass Bioenerg. 2012, 47, 302–315. [Google Scholar] [CrossRef]
- Millar, G.L. Use of dinitrosalicyclic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Da-Vinha, F.N.M.; Gravina, O.M.P.; Franco, M.N.; Macrae, A.; Coelho, R.R.R. Cellulase production by Streptomyces viridobrunneus SCPE-09 using lignocellulosic biomass as inducer substrate. Appl. Biochem. Biotechnol. 2011, 164, 256–267. [Google Scholar] [CrossRef] [PubMed]
- Brzezinska, M.S.; Jankiewicz, J.; Burkowsk, A. Purification and characterization of Streptomyces albidoflavus antifungal components. Appl. Biochem. Microbiol. 2013, 49, 451–457. [Google Scholar] [CrossRef]
- Das, P.; Solanki, R.; Khanna, M. Isolation and Screening of cellulolytic Actinomycetes from diverse habitats. Int. J. Adv. Biotechnol. Res. 2014, 5, 438–451. [Google Scholar]
- Gupta, U.; Kar, R. Optimization and scale up of cellulase free endo xylanase production by solid state fermentation on corn cob and by immobilized cells of a thermotolerant bacterial isolate. Jordan J. Biosci. 2008, 1, 129–134. [Google Scholar]
- Olson, E.R. Influence of pH on bacterial gene expression. Mol. Microbiol. 1993, 8, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Tuncer, M.; Kuru, A.; Isikli, M.; Sahin, N.; Çelenk, F.G. Optimization of extracellular endoxylanase, endoglucanase and peroxidase production by Streptomyces sp. F2621 isolated in Turkey. J. Appl. Microbiol. 2004, 97, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Heck, J.X.; Hertz, P.F.; Ayub, M.A.Z. Cellulase and xylanase production by isolated Amazon Bacillus strains using soya been industrial residue based solid-state cultivation. Braz. J. Microbiol. 2002, 33, 213–218. [Google Scholar] [CrossRef]
- Solingen, P.; Meijer, D.; Kleij, W.A.; Barnett, C.; Bolle, R.; Power, S.D.; Jones, B.E. Cloning and expression of an endocellulase gene from a novel streptomycete isolated from an East African soda lake. Extremophiles 2001, 5, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Acharya, S.; Chaudhary, A. Optimization of fermentation conditions for cellulases production by Bacillus licheniformis MVS1 and Bacillus sp. MVS3 Isolated from Indian Hot Spring. Braz. Arch. Biol. Technol. 2012, 55, 497–503. [Google Scholar] [CrossRef]
- Immanuel, G.; Dhanusha, R.; Prema, P.; Palavesam, A. Effect of different growth parameters on endoglucanase enzyme activity by bacteria isolated from coir retting effluents of estuarine environment. Int. J. Environ. Sci. Technol. 2006, 3, 25–34. [Google Scholar] [CrossRef]
- Bhalla, A.; Bischoff, K.M.; Sani, R.K. Highly thermostable xylanase production from a thermophilic Geobacillus strain WSUCF1 utilizing lignocellulosic biomass. Front. Bioeng. Biotechnol. 2015, 3, 84. [Google Scholar] [CrossRef] [PubMed]
- Juturu, V.; Wu, J.C. Microbial cellulases: Engineering, production and applications. Renew. Sustain. Energy Rev. 2014, 33, 188–203. [Google Scholar] [CrossRef]
- Abdelwahed, N.A.M.; El-Naggar, N.E.; Saber, W.I.A. Factors and correlations controlling cellulase-free xylanase production by Streptomyces halstedii NRRL B-1238 in submerged culture. Aust. J. Basic Appl. Sci. 2011, 5, 45–53. [Google Scholar]
- Thanongsak, C.; Ampin, K.; Charin, T.; Noppol, L.; Phisit, S.; Prasert, H. Optimization of cellulase-free xylanase production by thermophilic Streptomyces thermovulgaris TISTER1948 through Plackett-Burman and response surface methodological approaches. Biosci. Biotechnol. Biochem. 2011, 75, 531–537. [Google Scholar]
- Chellapandi, P.; Jani, H.M. Production of endoglucanase by the native strains of Streptomyces isolates in submerged fermentation. Braz. J. Microbiol. 2008, 39, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.; Kumar, M.R.; Kumar, D.J.M.; Balashanmugam, P.; Kumaran, B.M.D.; Kalaichelvan, P.T. Cellulase production by Bacillus subtilis isolated from cow dung. Arch. Appl. Sci. Res. 2012, 4, 269–279. [Google Scholar]
- Jaradat, Z.; Dawagreh, A.; Ababneh, Q.; Saadoun, I. Influence of culture conditions on cellulase production by Streptomyces Sp. (Strain J2). Jordan J. Biol. Sci. 2008, 1, 141–146. [Google Scholar]
- Techapun, C.; Charoenrat, T.; Poosaran, N.; Watanabe, M.; Sasaki, K. Thermostable and alkaline-tolerant cellulase-free xylanase produced by thermotolerant Streptomyces sp. Ab106. J. Biosci. Bioeng. 2002, 93, 431–433. [Google Scholar] [CrossRef]
- Sepahy, A.A.; Ghazi, S.; Sepahy, M.A. Cost-effective production and optimization of alkaline xylanase by indigenous Bacillus mojavensis AG137 fermented on waste. Enzym. Res. 2011, 2011, 593624. [Google Scholar] [CrossRef]
- Jang, H.-D.; Chang, K.-S. Thermostable cellulases from Streptomyces sp.: Scale-up production in a 50-l fermenter. Biotechnol. Lett. 2005, 27, 239–242. [Google Scholar] [CrossRef] [PubMed]
- Balan, V.; Jin, M.; Culbertson, A.; Uppugundla, N. The saccharification step: Trichoderma reseei cellulase hyper producer strains. In Lignocellulose Conversion Enzymatic and Microbial Tools for Bioethanol Production; Faraco, V., Ed.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 65–92. [Google Scholar]
- Xia, X.; Lin, S.; Xia, X.-X.; Cong, F.-S.; Zhong, J.-J. Significance of agitation-induced shear stress on mycelium morphology and lavendamycin production by engineered Streptomyces flocculus. Appl. Microbiol. Biotechnol. 2014, 98, 4399–4407. [Google Scholar] [CrossRef] [PubMed]
- Papagianni, M.; Moo-Young, M. Protease secretion in glucoamylase producer Aspergillus niger cultures: Fungal morphology and inoculum effects. Proc. Biochem. 2002, 37, 1271–1278. [Google Scholar] [CrossRef]
- George, J.; Arunachalam, R.; Paulkumar, K.; Wisely, E.G.; Shiburaj, S.; Annadura, G. Characterization and Phylogenetic Analysis of Cellulase Producing Streptomyces noboritoensis SPKC1. Interdiscip. Sci. Comput. Life Sci. 2010, 2, 205–212. [Google Scholar] [CrossRef] [PubMed]
- El-Naggar, N.E.-A.; Abdelwahed, N.A.M. Optimization of process parameters for the production of alkali-tolerant carboxymethyl cellulase by newly isolated Streptomyces sp. strain NEAE-D. Afr. J. Biotechnol. 2012, 11, 1185–1196. [Google Scholar]
- Nagar, S.; Mittal, A.; Kumar, D.; Kumar, L.; Kuhad, R.C.; Gupta, V.K. Hyper production of alkali stable xylanase in lesser duration by Bacillus pumilus SV-85S using wheat bran under solid state fermentation. New Biotechnol. 2011, 28, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Prakash, P.; Jayalakshmi, S.K.; Prakash, B.; Rubul, M.; Sreeramulu, K. Production of alkaliphilic, halotolerent, thermostable cellulase free xylanase by Bacillus halodurans PPKS-2 using agro waste: Single step purification and characterization. World J. Microbiol. Biotechnol. 2011, 28, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Gaur, R.; Tiwari, S.; Rai, P.; Srivastava, V. Isolation, production, and characterization of thermotolerant xylanase from solvent tolerant Bacillus vallismortis RSPP 15. Int. J. Polym. Sci. 2015, 2015, 986324. [Google Scholar] [CrossRef]
- Porsuk, İ.; Özakin, S.; Bali, B.; Yilmaz, E.İ. A cellulase-free, thermoactive, and alkali xylanase production by terrestrial Streptomyces sp. CA24. Turk. J. Biol. 2013, 37, 370–375. [Google Scholar] [CrossRef]
- McKee, S.L.; Pena, J.M.; Rogowski, A.; Jackson, A.; Lewis, J.R.; York, S.W.; Krogh, K.B.; Viksø-Nielsen, A.; Skjøt, M.; Gilbert, H.J.; et al. Introducing endo-xylanase activity into an exo-acting arabinofuranosidase that targets side chains. Proc. Natl. Acad. Sci. USA 2012, 109, 6537–6542. [Google Scholar] [CrossRef] [PubMed]
- Olsson, L.; Christensen, T.M.I.E.; Hansen, K.P.; Palmqvist, E.A. Influence of the carbon source on the production of cellulases, hemicellulases and pectinases by Trichoderma reesei Rut C-30. Enzym. Microb. Technol. 2003, 33, 612–619. [Google Scholar] [CrossRef]
- Motta, F.L.; Andrade, C.C.P.; Santana, M.H.A. A review of xylanase production by the fermentation of xylan: Classification, characterization and applications. In Sustainable Degradation of Lignocellulosic Biomass; Chandel, A.K., da Silva, S.S., Eds.; INTECH: Rijeka, Croatia, 2013; pp. 251–275. [Google Scholar]
- Singh, K.; Richa, K.; Bose, H.; Karthik, L.; Kumar, G.; Rao, B.V.K. Statistical media optimization and cellulase production from marine Bacillus VITRKHB. 3 Biotech 2014, 4, 591–598. [Google Scholar] [CrossRef]
- Shankar, T.; Isaiarasu, L. Cellulase production by Bacillus pumilus EWBCM1 under varying cultural conditions. Middle-East J. Sci. Res. 2011, 8, 40–45. [Google Scholar]
- Khandeparkar, R.D.D.; Bhosle, N.B. Isolation, purification and characterization of the xylanase produced by Arthrobacter sp. MTCC 5214 when grown in solid-state fermentation. Enzym. Microb. Technol. 2006, 39, 732–742. [Google Scholar] [CrossRef]
- Bhosale, J.H.; Sukalkar, R.S.; Uzma, Z.M.S.; Kadam, A.T. Production of xylanase by Streptomyces rameus grown on agricultural wastes. Biotechnol. Bioinf. Bioeng. 2011, 1, 505–512. [Google Scholar]






{kind=link}
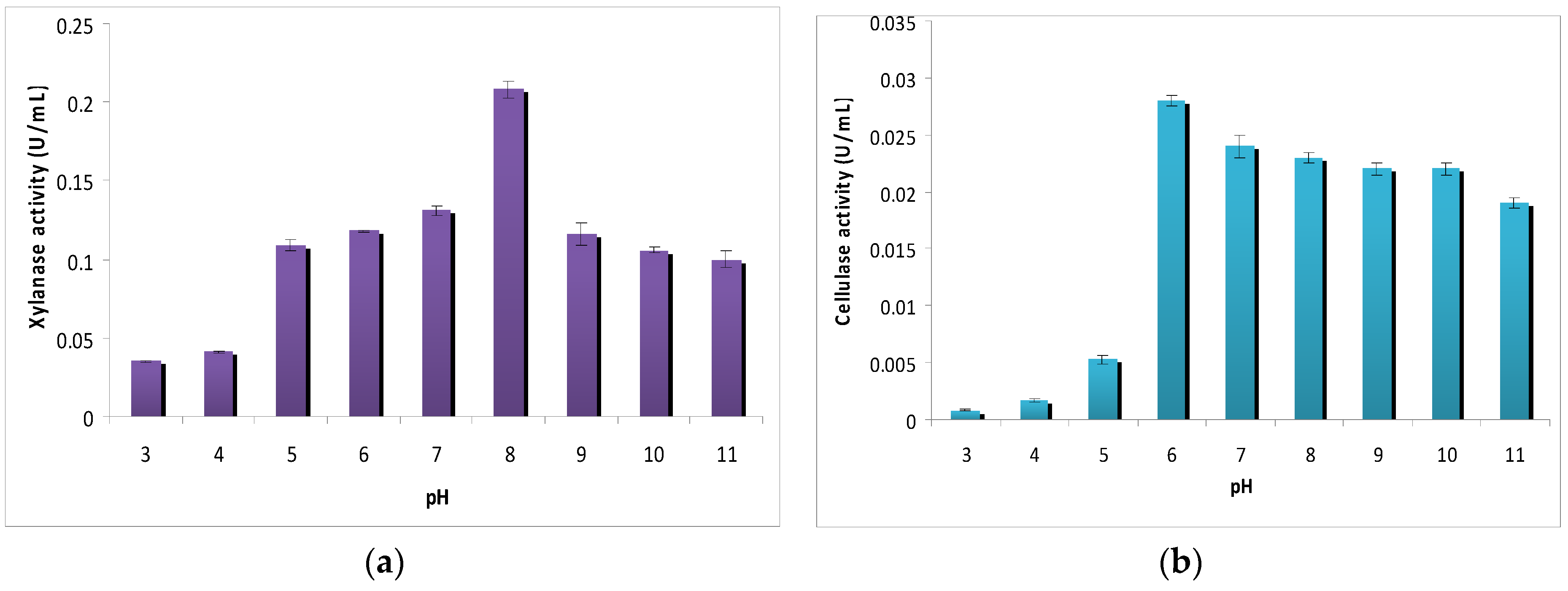{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Carbon Source | Cellulase Activity (U/mL) | Xylanase Activity (U/mL) |
|---|---|---|
| Monosaccharides | ||
| Arabinose | 0.061 b ± 0.01 | 0.30 a ± 0.01 |
| Galactose | ND | 10.35 a ± 0.65 |
| Glucose | ND | 2.55 a ± 0.36 |
| Mannose | 1.33 b ± 0.014 | 5.92 a ± 0.57 |
| Xylose | ND | 8.02 a ± 0.65 |
| Disaccharide | ||
| Cellobiose | ND | 4.39 a ± 0.09 |
| Polysaccharide | ||
| Avicel® | ND | 1.66 a ± 0.13 |
| CMC | 0.070 b ± 0.01 | 2.58 a ± 0.15 |
| Xylan | ND | 14.15 a ± 0.07 |
| Lignocellulose | ||
| Wheat straw | 0.01 b ± 0.002 | 14.85 a ± 0.56 |
| Saw dust | ND | 2.29 a ± 0.35 |
| Control * | ND | 0.17 ± 0.01 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fatokun, E.N.; Nwodo, U.U.; Okoh, A.I. Classical Optimization of Cellulase and Xylanase Production by a Marine Streptomyces Species. Appl. Sci. 2016, 6, 286. https://doi.org/10.3390/app6100286
Fatokun EN, Nwodo UU, Okoh AI. Classical Optimization of Cellulase and Xylanase Production by a Marine Streptomyces Species. Applied Sciences. 2016; 6(10):286. https://doi.org/10.3390/app6100286
Chicago/Turabian StyleFatokun, Evelyn N., Uchechukwu U. Nwodo, and Anthony I. Okoh. 2016. "Classical Optimization of Cellulase and Xylanase Production by a Marine Streptomyces Species" Applied Sciences 6, no. 10: 286. https://doi.org/10.3390/app6100286