Aging, Aerobic Activity and Interhemispheric Communication


Abstract
:1. Introduction
2. Methods
2.1. Participants

{kind=link}
{kind=link}
| Active Middle-Age | Sedentary Middle-Age | |
|---|---|---|
| Age | 51.3 ± 5.9 (41–60) | 52.6 ± 6.9 (41–60) |
| N/Gender | 17/8 Female | 21/12 Female |
| Education | 16.1 ± 2.25 (12–20) | 16.4 ± 2.38 (12–20) |
| BMI * | 22.1 ± 2.5 * (18–25) | 26.7 ± 3.9 * (22–32) |
| VO2max * | 49.8 ± 12.7 * (36–69) | 29.6 ± 6.6 * (20–37) |
| Weekly Activity (min/bout) * | 146 ± 23.4 * (45–180) | 36.7 ± 12.5 * (15–45) |
2.2. Assessments
2.2.1. Motor Assessments
2.2.2. Estimated VO2max
| <80 bpm * | 80–90 bpm * | 90–100 bpm * | >100 bpm * | |
|---|---|---|---|---|
| Stage 1 | 125 watts | 100 watts | 75 watts | 50 watts |
| Stage 2 | 150 watts | 125 watts | 100 watts | 75 watts |
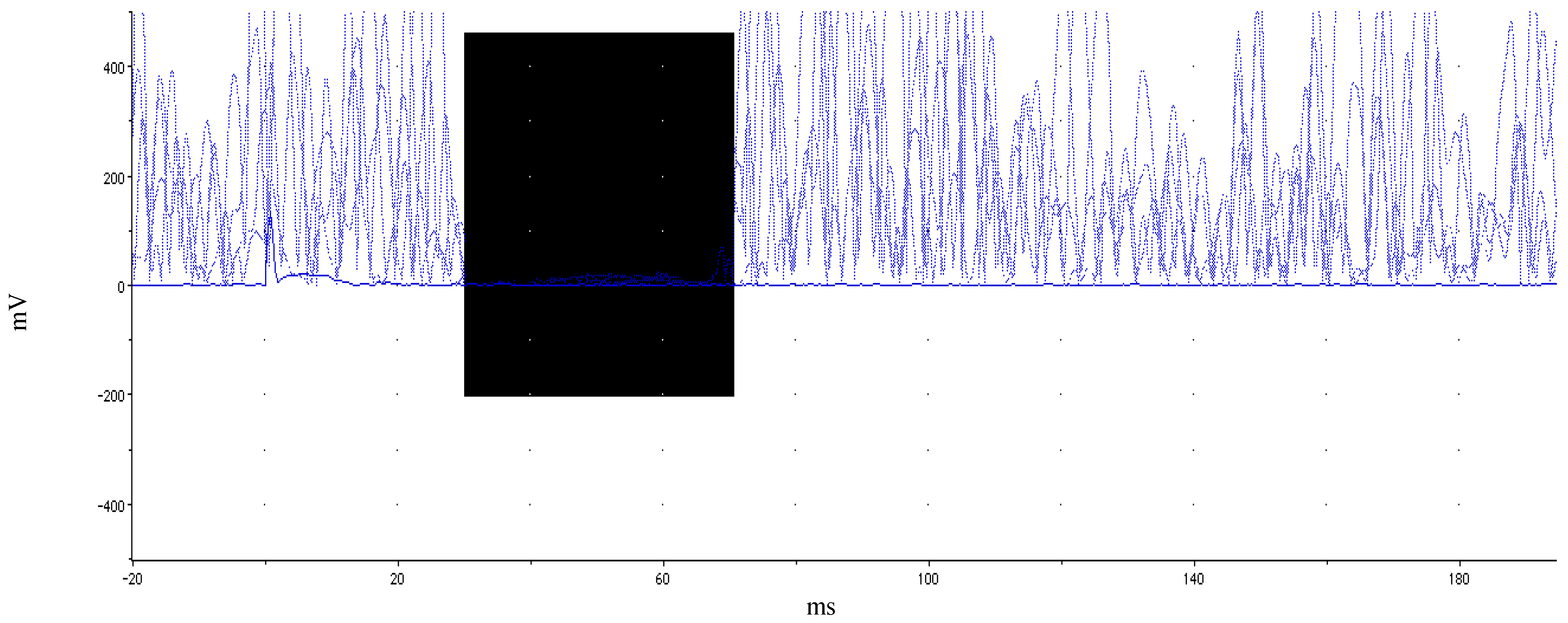
2.2.3. Single-Pulse Transcranial Magnetic Stimulation

2.3. Data Analysis
3. Results
3.1. Behavioral Results
| Active Middle-Age | Sedentary Middle-Age | |
|---|---|---|
| 9-hole Pegboard * (s) | 16.2 ± 1.76 * | 18.79 ± 3.03 * |
| Halstead Finger Tapping * (count) | 52.29 ± 7.3 * | 43.82 ± 8.79 * |
| Simple Reaction Time (ms) | 269 ± 32.25 | 289 ± 33.3 |
| Coin Rotation-Right (s) | 13.25 ± 2.88 | 14.99 ± 3.77 |
| Coin Rotation-Left * (s) | 13.05 ± 2.18 * | 15.3 ± 3.3 * |
| Bimanual Task-Right (s) | 13.16 ± 0.63 | 12.95 ±0.55 |
| Bimanual Task-Left (s) | 12.73 ± 1.39 | 12.61 ± 2.34 |
| Task Difference-Right * (s) | 0.31 ± 2.02 * | −2.74 ± 2.03 * |
| Task Difference-Left * (s) | 0.09 ± 1.83 * | −2.03 ± 3.2 * |
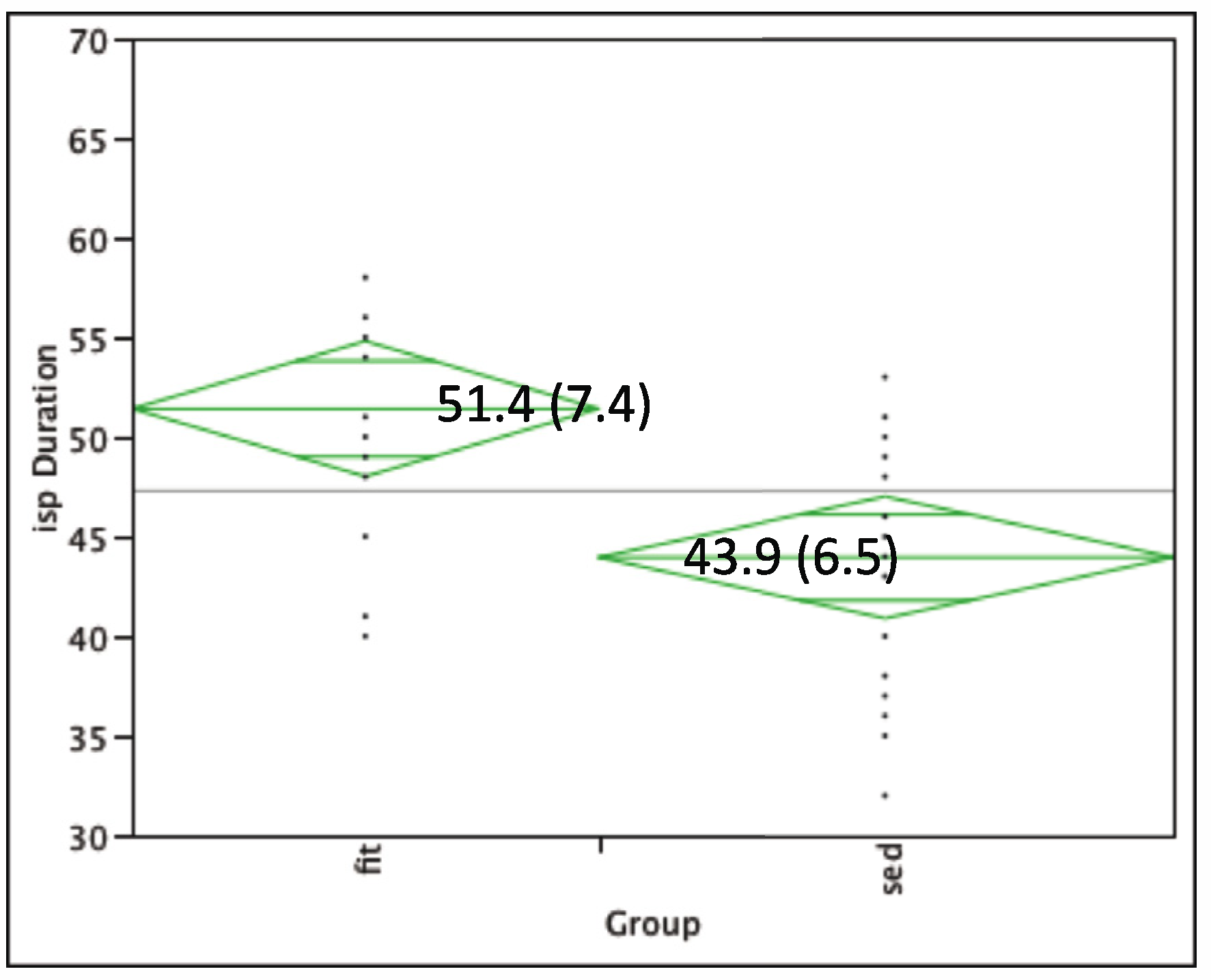
3.2. TMS Results
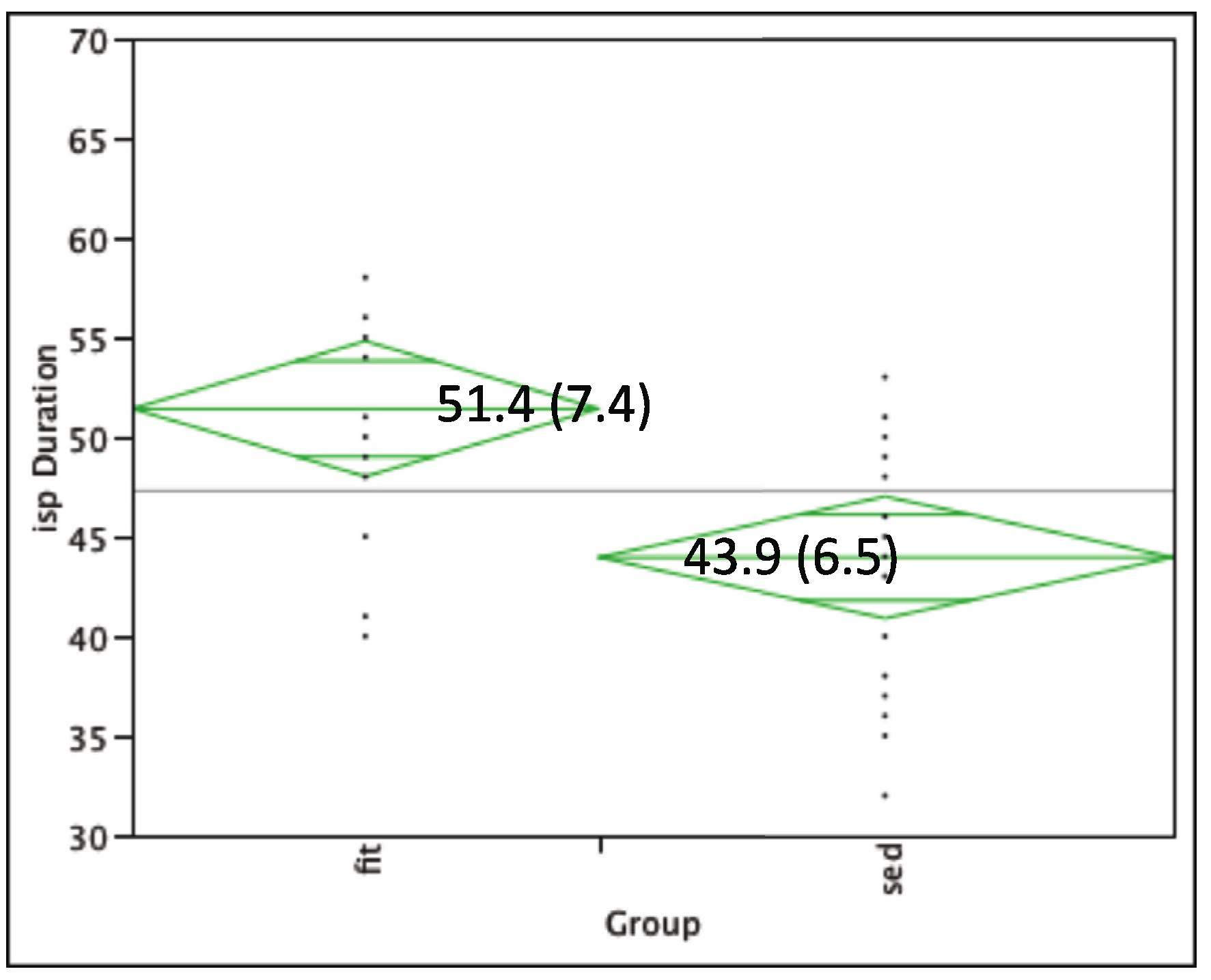
3.3. TMS and Behavioral Correlation Results
4. Discussion
5. Conclusions
Acknowledgments
Conflict of Interest
References
- Ward, N.S. Compensatory mechanisms in the aging motor system. Ageing Res. Rev. 2006, 5, 239–254. [Google Scholar] [CrossRef]
- Voelcker-Rehage, C.; Alberts, J.L. Age-related changes in grasping force modulation. Exp. Brain Res. 2005, 166, 61–70. [Google Scholar] [CrossRef]
- Seidler, R.D. Differential effects of age on sequence learning and sensorimotor adaptation. Brain Res. Bull. 2006, 70, 337–346. [Google Scholar] [CrossRef]
- Bernard, J.A.; Seidler, R.D. Evidence for motor cortex dedifferentiation in older adults. Neurobiol. Aging 2012, 33, 1890–1899. [Google Scholar] [CrossRef]
- Szabo, A.N.; Bangert, A.S.; Reuter-Lorenz, P.A.; Seidler, R.D. Physical activity is related to timing performance in older adults. Neuropsychol. Dev. Cogn. B Aging Neuropsychol. Cogn. 2012. [Google Scholar] [CrossRef]
- Hugle, T.; Geurts, J.; Nuesch, C.; Muller-Gerbl, M.; Valderrabano, V. Aging and osteoarthritis: An inevitable encounter? J. Aging Res. 2012, 2012. [Google Scholar] [CrossRef]
- Kuettner, K.E.; Cole, A.A. Cartilage degeneration in different human joints. Osteoarthritis Cartilage 2005, 13, 93–103. [Google Scholar] [CrossRef]
- Christou, E.A. Aging and variability of voluntary contractions. Exerc. Sport Sci. Rev. 2011, 39, 77–84. [Google Scholar] [CrossRef]
- Clark, B.C.; Taylor, J.L. Age-related changes in motor cortical properties and voluntary activation of skeletal muscle. Curr. Aging Sci. 2011, 4, 192–199. [Google Scholar] [CrossRef]
- Aoki, T.; Fukuoka, Y. Finger tapping ability in healthy elderly and young adults. Med. Sci. Sports Exerc. 2010, 42, 449–455. [Google Scholar]
- Riecker, A.; Groschel, K.; Ackermann, H.; Steinbrink, C.; Witte, O.; Kastrup, A. Functional significance of age-related differences in motor activation patterns. NeuroImage 2006, 32, 1345–1354. [Google Scholar] [CrossRef]
- McGregor, K.M.; Zlatar, Z.; Kleim, E.; Sudhyadhom, A.; Bauer, A.; Phan, S.; Seeds, L.; Ford, A.; Manini, T.M.; White, K.D.; et al. Physical activity and neural correlates of aging: A combined TMS/fMRI study. Behav. Brain Res. 2011, 222, 158–168. [Google Scholar] [CrossRef]
- Fujiyama, H.; Garry, M.I.; Levin, O.; Swinnen, S.P.; Summers, J.J. Age-related differences in inhibitory processes during interlimb coordination. Brain Res. 2009, 1262, 38–47. [Google Scholar]
- Fujiyama, H.; Hinder, M.R.; Schmidt, M.W.; Garry, M.I.; Summers, J.J. Age-related differences in corticospinal excitability and inhibition during coordination of upper and lower limbs. Neurobiol. Aging 2012, 33, 1484.e1–1484.e14. [Google Scholar] [CrossRef]
- Talelli, P.; Waddingham, W.; Ewas, A.; Rothwell, J.C.; Ward, N.S. The effect of age on task-related modulation of interhemispheric balance. Exp. Brain Res. 2008, 186, 59–66. [Google Scholar] [CrossRef]
- Talelli, P.; Ewas, A.; Waddingham, W.; Rothwell, J.C.; Ward, N.S. Neural correlates of age-related changes in cortical neurophysiology. NeuroImage 2008, 40, 1772–1781. [Google Scholar] [CrossRef]
- Peinemann, A.; Lehner, C.; Conrad, B.; Siebner, H.R. Age-related decrease in paired-pulse intracortical inhibition in the human primary motor cortex. Neurosci. Lett. 2001, 313, 33–36. [Google Scholar] [CrossRef]
- Meyer, B.U.; Roricht, S.; Woiciechowsky, C. Topography of fibers in the human corpus callosum mediating interhemispheric inhibition between the motor cortices. Ann. Neurol. 1998, 43, 360–369. [Google Scholar] [CrossRef]
- Kurth, F.; Mayer, E.A.; Toga, A.W.; Thompson, P.M.; Luders, E. The right inhibition? callosal correlates of hand performance in healthy children and adolescents callosal correlates of hand performance. Hum. Brain Mapp. 2012. [Google Scholar] [CrossRef]
- Sale, M.V.; Semmler, J.G. Age-related differences in corticospinal control during functional isometric contractions in left and right hands. J. Appl. Physiol. 2005, 99, 1483–1493. [Google Scholar] [CrossRef]
- Kramer, A.F.; Hahn, S.; Cohen, N.J.; Banich, M.T.; McAuley, E.; Harrison, C.R.; Chason, J.; Vakil, E.; Bardell, L.; Boileau, R.A.; Colcombe, A. Ageing, fitness and neurocognitive function. Nature 1999, 400, 418–419. [Google Scholar] [CrossRef]
- McGregor, K.; Craggs, J.; Benjamin, M.; Crosson, B.; White, K. Age-related changes in motor control during unimanual movements. Brain Imaging Behav. 2009, 3, 317–331. [Google Scholar] [CrossRef]
- Kossev, A.R.; Schrader, C.; Dauper, J.; Dengler, R.; Rollnik, J.D. Increased intracortical inhibition in middle-aged humans; a study using paired-pulse transcranial magnetic stimulation. Neurosci. Lett. 2002, 333, 83–86. [Google Scholar] [CrossRef]
- Gonzales, M.M.; Tarumi, T.; Kaur, S.; Nualnim, N.; Fallow, B.A.; Pyron, M.; Tanaka, H.; Haley, A.P. Aerobic fitness and the brain: Increased N-acetyl-aspartate and choline concentrations in endurance-trained middle-aged adults. Brain Topogr. 2012. [Google Scholar] [CrossRef]
- Louis, E.D.; Ottman, R. Study of possible factors associated with age of onset in essential tremor. Mov. Disord. 2006, 21, 1980–1986. [Google Scholar] [CrossRef]
- Taylor, C.A.; Saint-Hilaire, M.H.; Cupples, L.A.; Thomas, C.A.; Burchard, A.E.; Feldman, R.G.; Myers, R.H. Environmental, medical, and family history risk factors for Parkinson’s disease: A New England-based case control study. Am. J. Med. Genet. 1999, 88, 742–749. [Google Scholar] [CrossRef]
- Thomas, S. Revision of the Physical Activity Readiness Questionnaire (PAR-Q). Can. J. Sports Sci. 1992, 17, 338–345. [Google Scholar]
- Lamb, K.L.; Brodie, D.A. The assessment of physical activity by leisure-time physical activity questionnaires. Sports Med. 1990, 10, 159–180. [Google Scholar] [CrossRef]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Mathiowetz, V.; Weber, K.; Kashman, N.; Volland, G. Adult Norms for the nine hole peg test of finger dexterity. Occup. Ther. J. Res. 1985, 5, 24–38. [Google Scholar]
- Hanna-Pladdy, B.; Mendoza, J.E.; Apostolos, G.T.; Heilman, K.M. Lateralised motor control: Hemispheric damage and the loss of deftness. J. Neurol. Neurosurg. Psychiatry 2002, 73, 574–577. [Google Scholar] [CrossRef]
- Mendoza, J.E.; Apostolos, G.T.; Humphreys, J.D.; Hanna-Pladdy, B.; O’Bryant, S.E. Coin rotation task (CRT): A new test of motor dexterity. Arch. Clin. Neuropsychol. 2009, 24, 287–292. [Google Scholar] [CrossRef]
- Garatachea, N.; Cavalcanti, E.; Garcia-Lopez, D.; Gonzalez-Gallego, J.; de Paz, J.A. Estimation of energy expenditure in healthy adults from the YMCA submaximal cycle ergometer test. Eval. Health Prof. 2007, 30, 138–149. [Google Scholar] [CrossRef]
- Garvey, M.A.; Ziemann, U.; Becker, D.A.; Barker, C.A.; Bartko, J.J. New graphical method to measure silent periods evoked by transcranial magnetic stimulation. Clin. Neurophysiol. 2001, 112, 1451–1460. [Google Scholar] [CrossRef]
- Christensen, C.L.; Payne, V.G.; Wughalter, E.H.; Yan, J.H.; Henehan, M.; Jones, R. Physical activity, physiological, and psychomotor performance: A study of variously active older adult men. Res. Q. Exerc. Sport 2003, 74, 136–142. [Google Scholar]
- Spirduso, W.W. Reaction and movement time as a function of age and physical activity level. J. Gerontol. 1975, 30, 435–440. [Google Scholar]
- Seeman, T.E.; Berkman, L.F.; Charpentier, P.A.; Blazer, D.G.; Albert, M.S.; Tinetti, M.E. Behavioral and psychosocial predictors of physical performance: MacArthur studies of successful aging. J. Gerontol. A Biol. Sci. Med. Sci. 1995, 50, M177–M183. [Google Scholar]
- Baliz, Y.; Armatas, C.; Farrow, M.; Hoy, K.E.; Fitzgerald, P.B.; Bradshaw, J.L.; Georgiou-Karistianis, N. The influence of attention and age on the occurrence of mirror movements. J. Int. Neuropsychol. Soc. 2005, 11, 855–862. [Google Scholar]
- Addamo, P.K.; Farrow, M.; Hoy, K.E.; Bradshaw, J.L.; Georgiou-Karistianis, N. The effects of age and attention on motor overflow production—A review. Brain Res. Rev. 2007, 54, 189–204. [Google Scholar] [CrossRef]
- Wu, T.; Hallett, M. The influence of normal human ageing on automatic movements. J. Physiol. 2005, 562, 605–615. [Google Scholar]
- Addamo, P.K.; Farrow, M.; Hoy, K.E.; Bradshaw, J.L.; Georgiou-Karistianis, N. The influence of task characteristics on younger and older adult motor overflow. Q. J. Exp. Psychol. (Hove) 2009, 62, 239–247. [Google Scholar] [CrossRef]
- Fling, B.W.; Kwak, Y.; Peltier, S.J.; Seidler, R.D. Differential relationships between transcallosal structural and functional connectivity in young and older adults. Neurobiol. Aging 2012, 33, 2521–2526. [Google Scholar] [CrossRef]
- Langan, J.; Peltier, S.J.; Bo, J.; Fling, B.W.; Welsh, R.C.; Seidler, R.D. Functional implications of age differences in motor system connectivity. Front. Syst. Neurosci. 2010, 4, 17. [Google Scholar]
- Giovannelli, F.; Borgheresi, A.; Balestrieri, F.; Zaccara, G.; Viggiano, M.P.; Cincotta, M.; Ziemann, U. Modulation of interhemispheric inhibition by volitional motor activity: An ipsilateral silent period study. J. Physiol. 2009, 587, 5393–5410. [Google Scholar] [CrossRef]
- Bodwell, J.A.; Mahurin, R.K.; Waddle, S.; Price, R.; Cramer, S.C. Age and features of movement influence motor overflow. J. Am. Geriatr. Soc. 2003, 51, 1735–1739. [Google Scholar] [CrossRef]
- Mayston, M.J.; Harrison, L.M.; Stephens, J.A. A neurophysiological study of mirror movements in adults and children. Ann. Neurol. 1999, 45, 583–594. [Google Scholar] [CrossRef]
- Krams, M.; Quinton, R.; Mayston, M.J.; Harrison, L.M.; Dolan, R.J.; Bouloux, P.M.; Stephens, J.A.; Frackowiak, R.S.; Passingham, R.E. Mirror movements in X-linked Kallmann’s syndrome. II. A PET study. Brain 1997, 120, 1217–1228. [Google Scholar] [CrossRef]
- Mayston, M.J.; Harrison, L.M.; Quinton, R.; Stephens, J.A.; Krams, M.; Bouloux, P.M. Mirror movements in X-linked Kallmann’s syndrome. I. A neurophysiological study. Brain 1997, 120, 1199–1216. [Google Scholar] [CrossRef]
- Beck, S.; Shamim, E.A.; Richardson, S.P.; Schubert, M.; Hallett, M. Inter-hemispheric inhibition is impaired in mirror dystonia. Eur. J. Neurosci. 2009, 29, 1634–1640. [Google Scholar] [CrossRef]
- Erol, F.S.; Ucler, N.; Yakar, H. The association of Chiari type III malformation and Klippel-Feil syndrome with mirror movement: A case report. Turk. Neurosurg. 2011, 21, 655–658. [Google Scholar]
- Erickson, K.I.; Prakash, R.S.; Voss, M.W.; Chaddock, L.; Heo, S.; McLaren, M.; Pence, B.D.; Martin, S.A.; Vieira, V.J.; Woods, J.A.; et al. Brain-derived neurotrophic factor is associated with age-related decline in hippocampal volume. J. Neurosci. 2010, 30, 5368–5375. [Google Scholar]
- Erickson, K.I.; Miller, D.L.; Roecklein, K.A. The aging hippocampus: Interactions between exercise, depression, and BDNF. Neurosci. 2012, 18, 82–97. [Google Scholar] [CrossRef]
- Pedersen, B.K. Exercise-induced myokines and their role in chronic diseases. Brain Behav. Immun. 2011, 25, 811–816. [Google Scholar] [CrossRef]
- Kaliman, P.; Parrizas, M.; Lalanza, J.F.; Camins, A.; Escorihuela, R.M.; Pallas, M. Neurophysiological and epigenetic effects of physical exercise on the aging process. Ageing Res. Rev. 2011, 10, 475–486. [Google Scholar] [CrossRef]
- Bloor, C.M. Angiogenesis during exercise and training. Angiogenesis 2005, 8, 263–271. [Google Scholar] [CrossRef]
- Colcombe, S.J.; Kramer, A.F.; Erickson, K.I.; Scalf, P.; McAuley, E.; Cohen, N.J.; Webb, A.; Jerome, G.J.; Marquez, D.X.; Elavsky, S. Cardiovascular fitness, cortical plasticity, and aging. Proc. Natl. Acad. Sci. USA 2004, 101, 3316–3321. [Google Scholar]
- Erickson, K.I.; Voss, M.W.; Prakash, R.S.; Basak, C.; Szabo, A.; Chaddock, L.; Kim, J.S.; Heo, S.; Alves, H.; White, S.M.; et al. Exercise training increases size of hippocampus and improves memory. Proc. Natl. Acad. Sci. USA 2011, 108, 3017–3022. [Google Scholar]
- Hötting, K.; Schauenburg, G.; Röder, B. Long-term effects of physical exercise on verbal learning and memory in middle-aged adults: Results of a one-year follow-up study. Brain Sci. 2012, 2, 332–346. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
McGregor, K.M.; Heilman, K.M.; Nocera, J.R.; Patten, C.; Manini, T.M.; Crosson, B.; Butler, A.J. Aging, Aerobic Activity and Interhemispheric Communication. Brain Sci. 2012, 2, 634-648. https://doi.org/10.3390/brainsci2040634
McGregor KM, Heilman KM, Nocera JR, Patten C, Manini TM, Crosson B, Butler AJ. Aging, Aerobic Activity and Interhemispheric Communication. Brain Sciences. 2012; 2(4):634-648. https://doi.org/10.3390/brainsci2040634
Chicago/Turabian StyleMcGregor, Keith M., Kenneth M. Heilman, Joe R. Nocera, Carolynn Patten, Todd M. Manini, Bruce Crosson, and Andrew J. Butler. 2012. "Aging, Aerobic Activity and Interhemispheric Communication" Brain Sciences 2, no. 4: 634-648. https://doi.org/10.3390/brainsci2040634