Is There Release from Masking from Isomorphism between Perception and Action?

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Ideomotor Theory and Perceptual Resonance
1.2. Strong Dimensional Overlap
2. Experimental Section

2.1. Experiment 1: No Dimensional Overlap versus Bottom-Up Salience
2.1.1. Participants
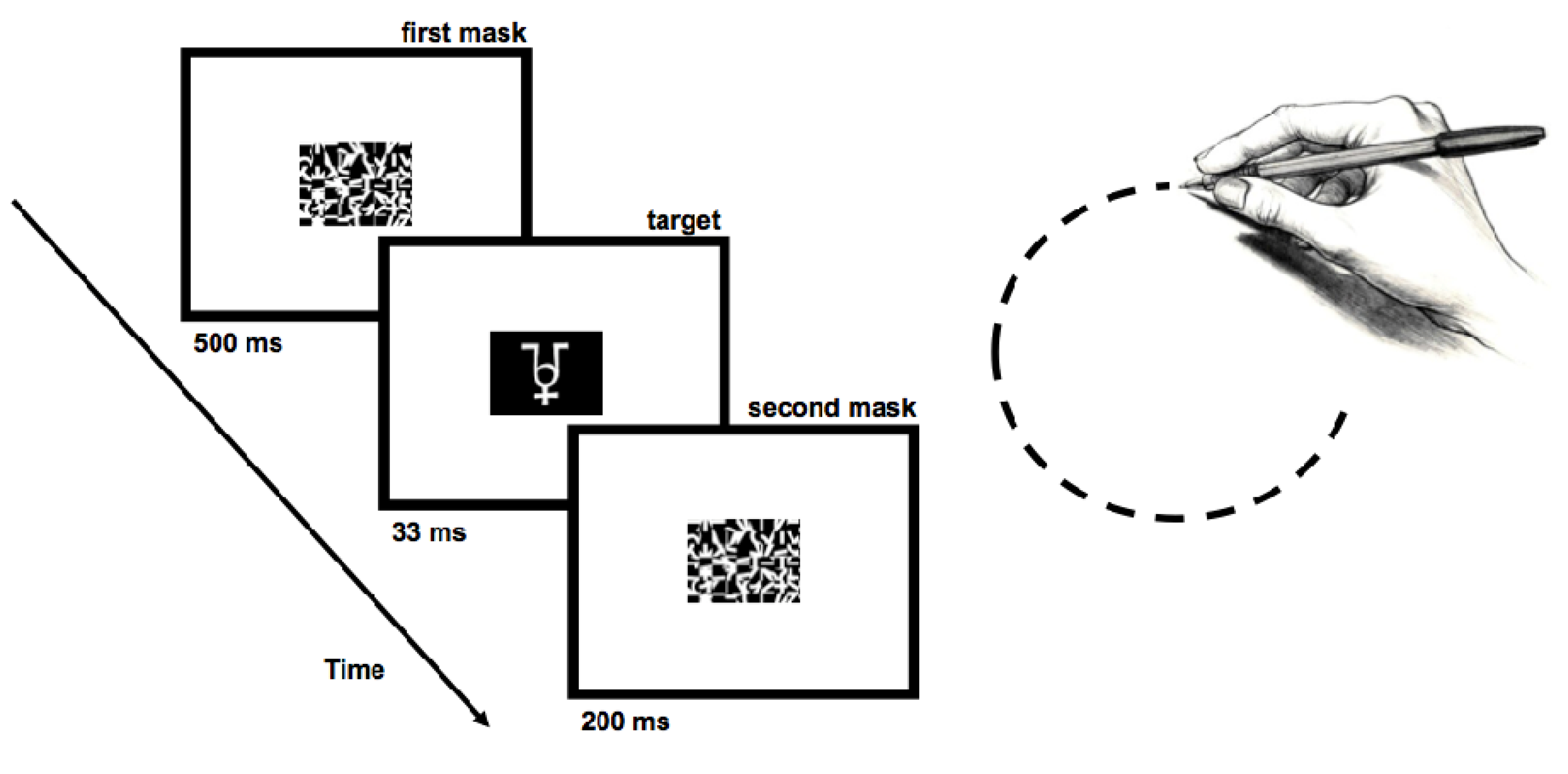
2.1.2. Stimuli

2.1.3. Procedures
2.2. Experiment 2: Weak Dimensional Overlap
2.2.1. Participants
2.2.2. Procedures
2.3. Experiment 3: Strong Dimensional Overlap

2.3.1. Participants
2.3.2. Procedures
3. Results and Discussion
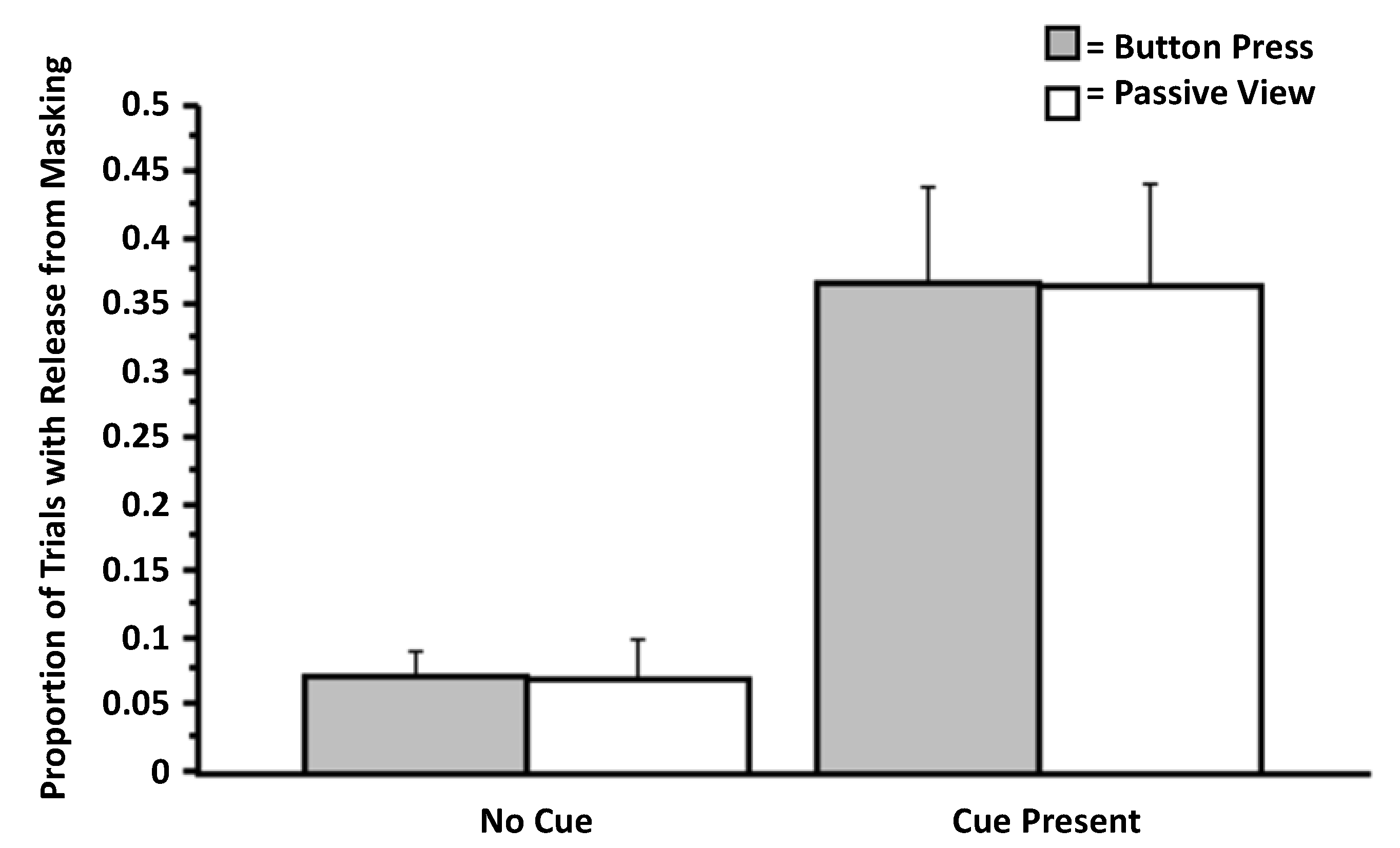
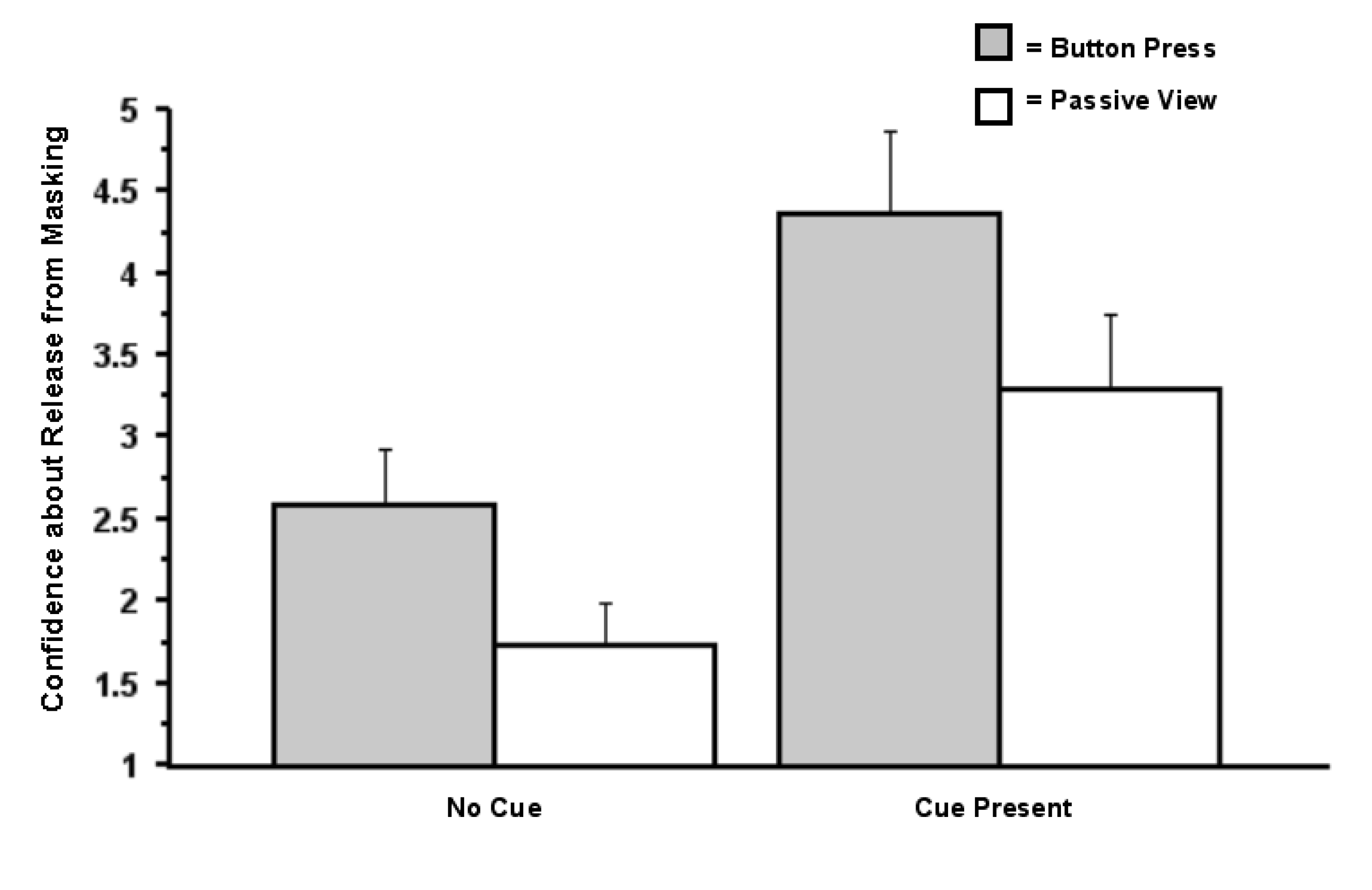
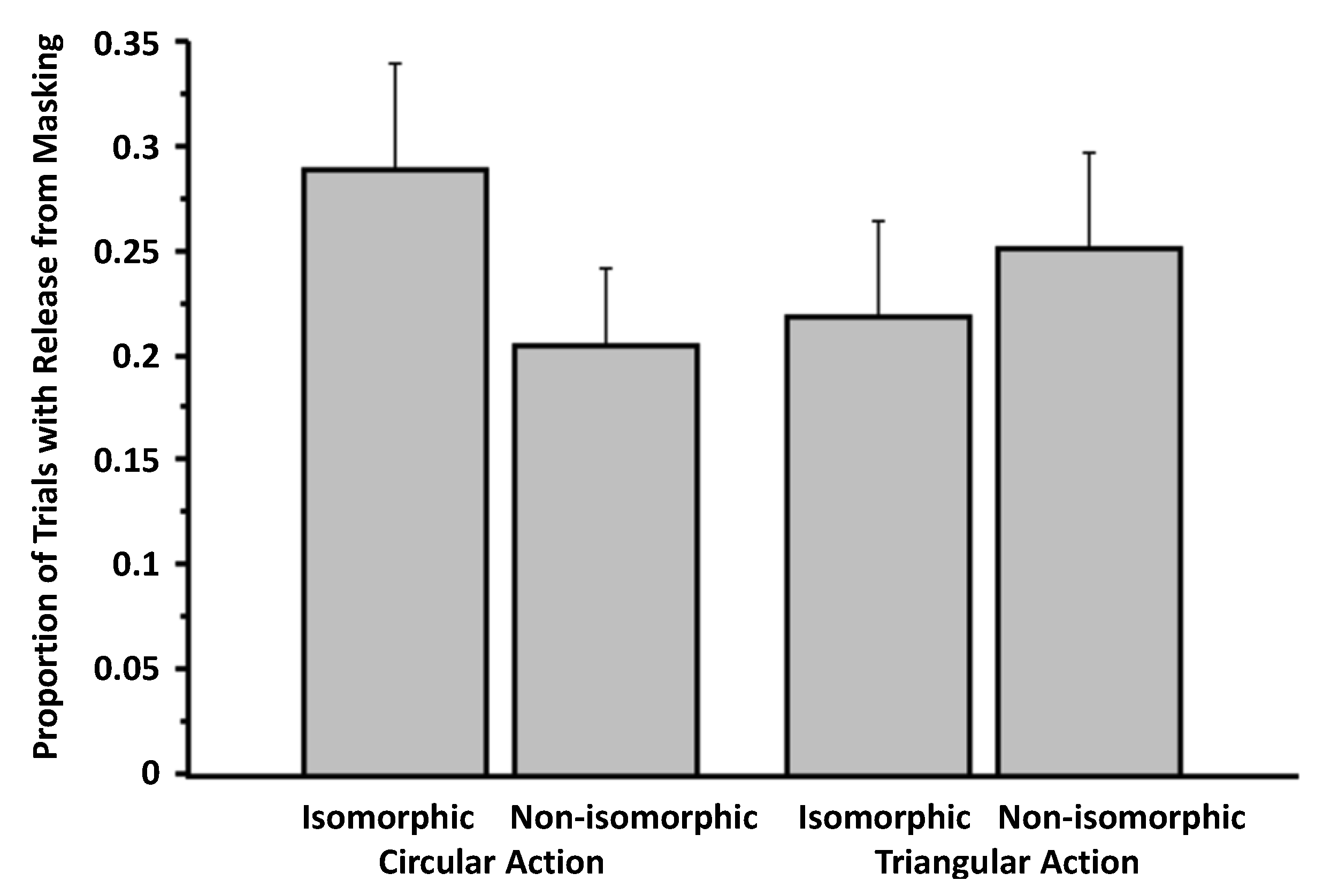
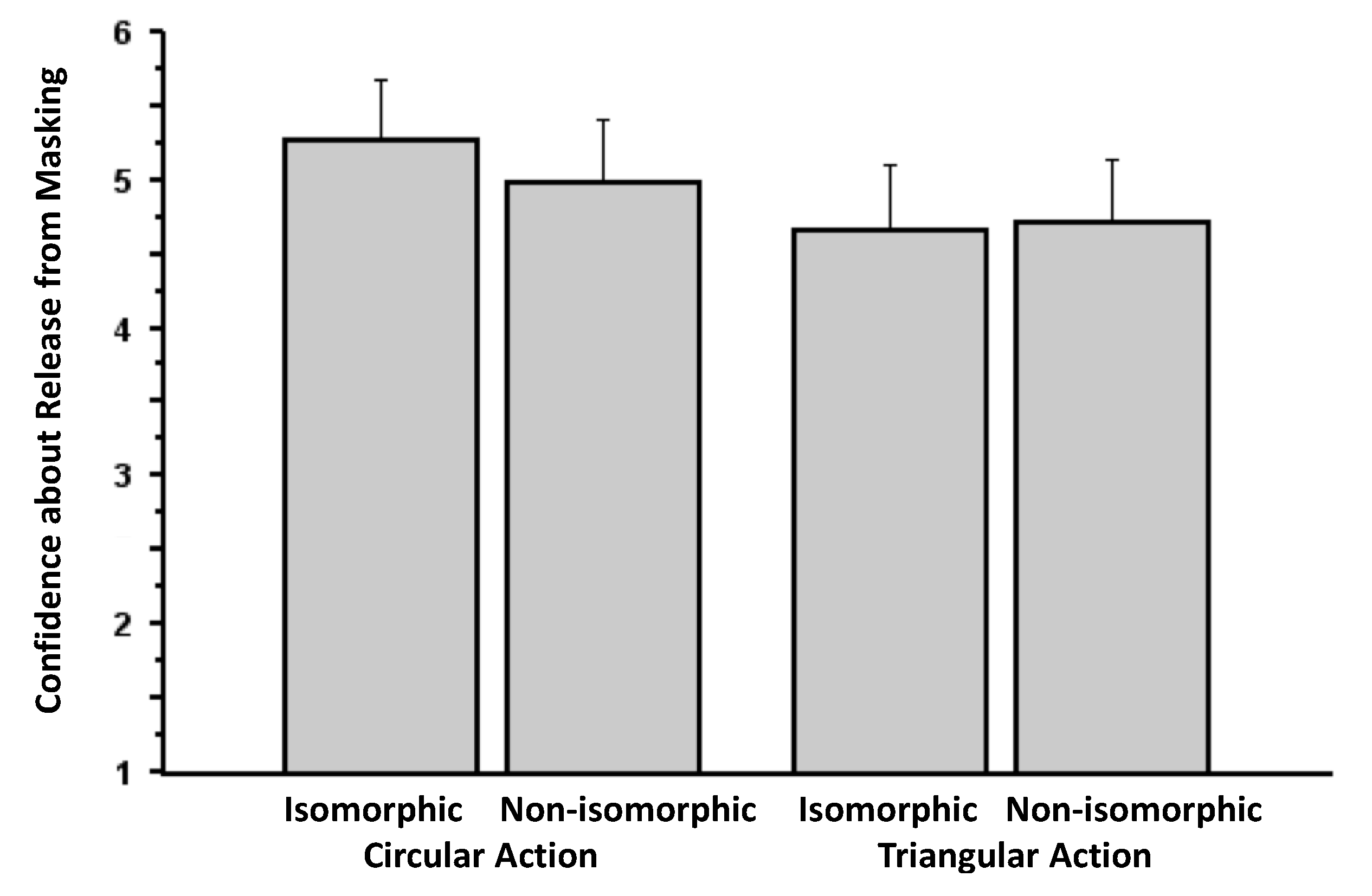
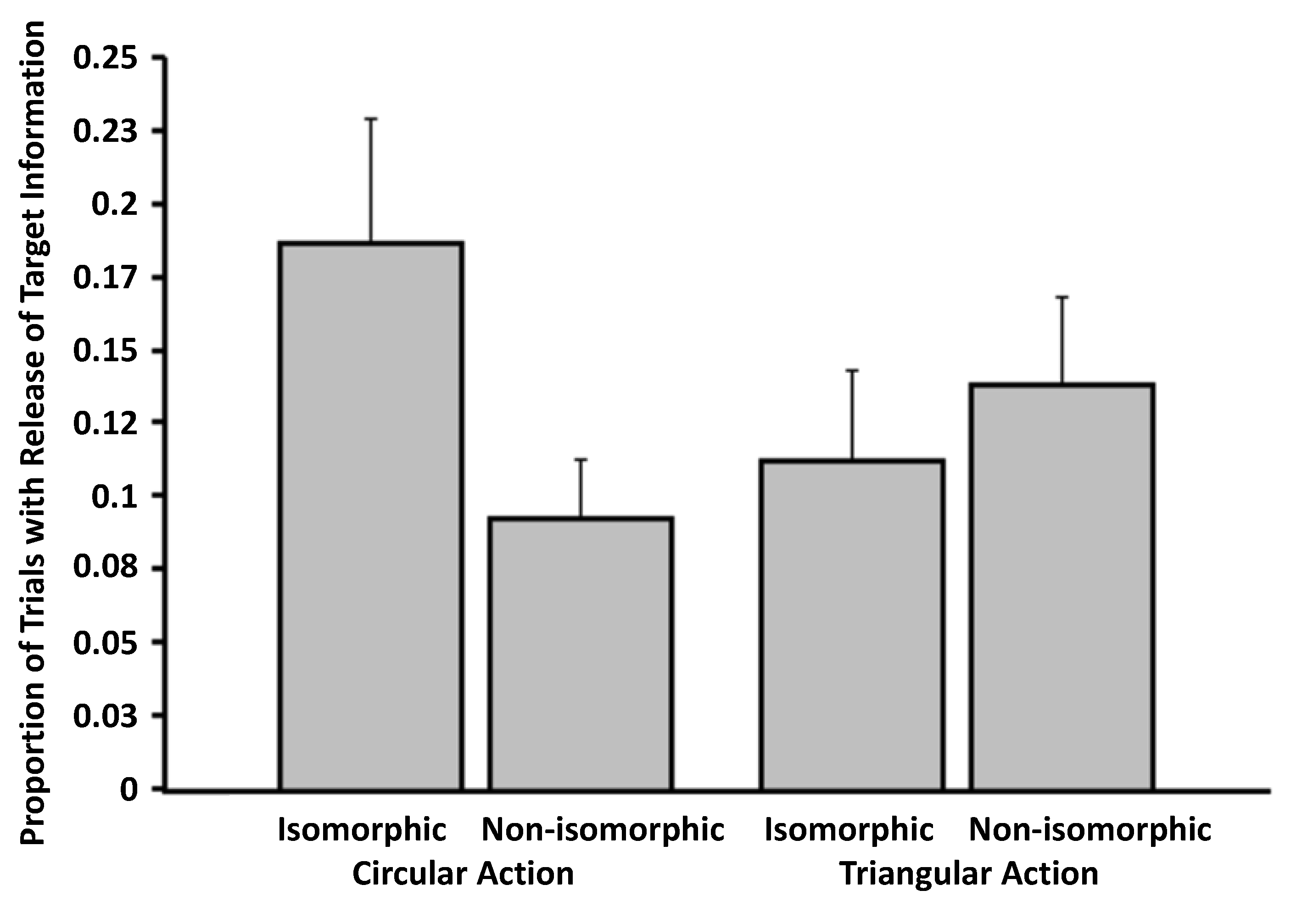
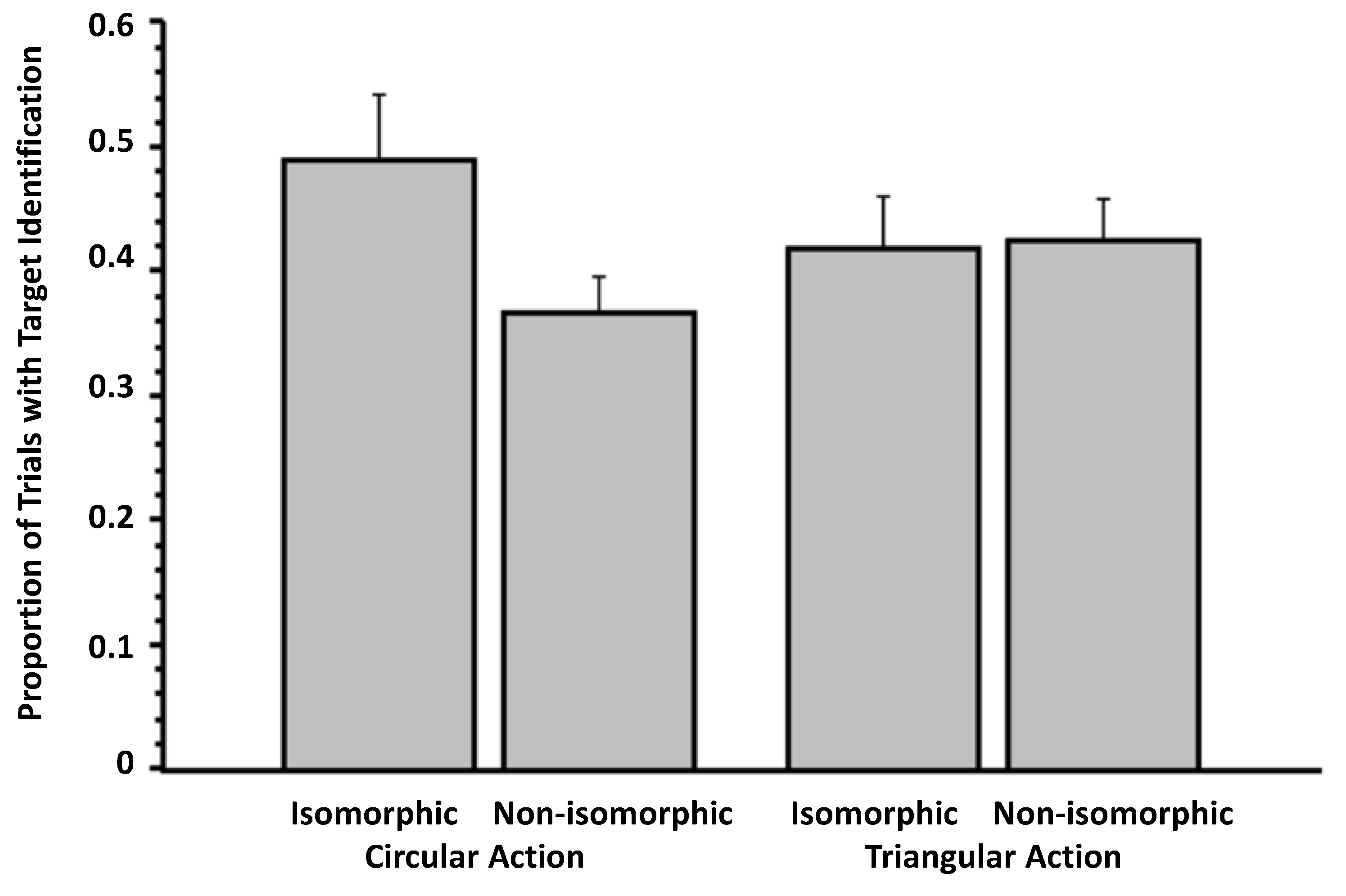
3.1. Experiment 1


3.2. Experiment 2
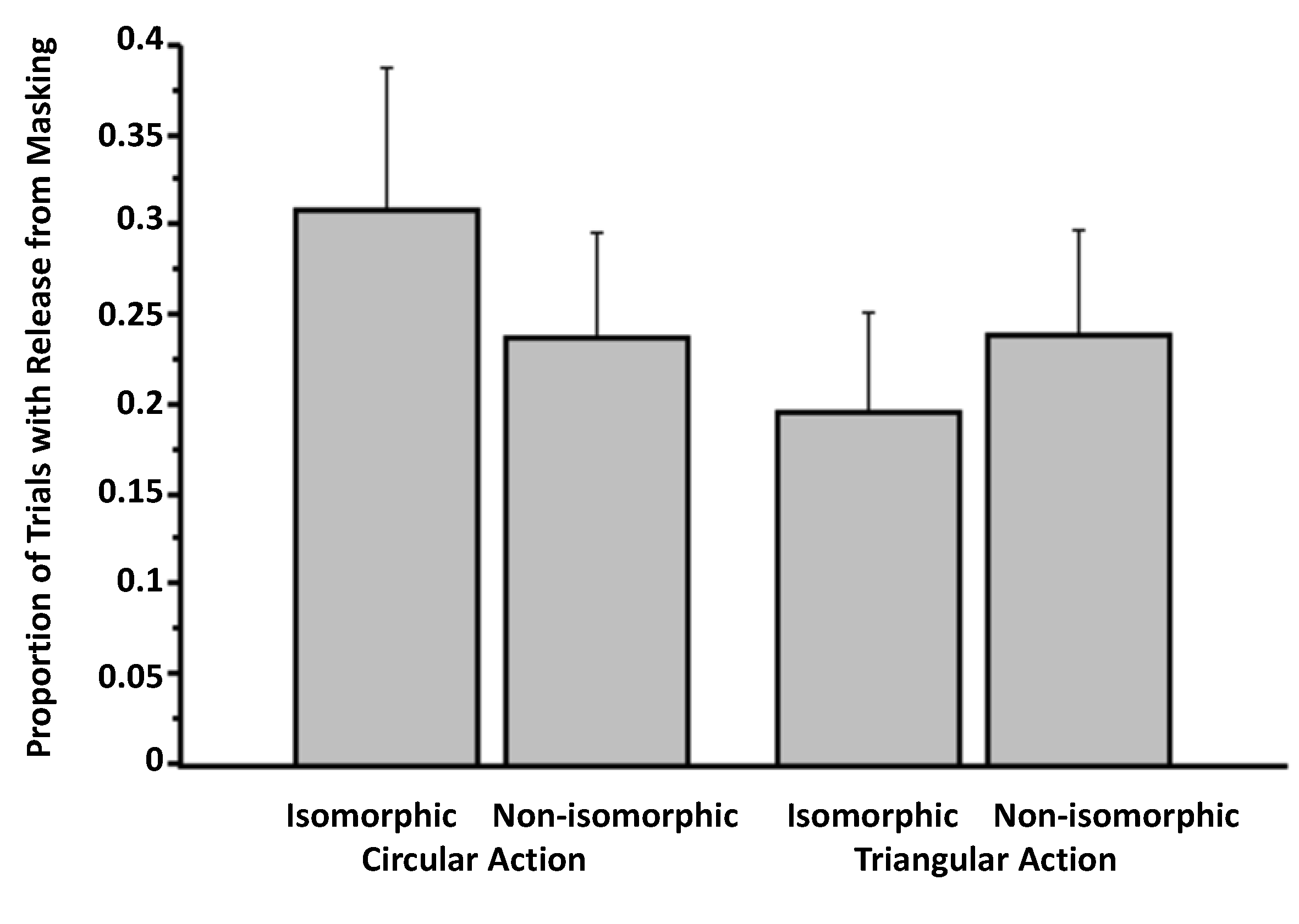
3.3. Experiment 3




4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Di Lollo, V.; Enns, J.T.; Rensink, R.A. Competition for consciousness among visual events: The psychophysics of reentrant visual processes. J. Exp. Psychol. Gen. 2000, 129, 481–507. [Google Scholar] [CrossRef]
- Mathewson, K.; Fabiani, M.; Gratton, G.; Beck, D.; Lleras, A. Rescuing stimuli from invisibility: Inducing a momentary release from visual masking with pre-target entrainment. Cognition 2009, 115, 186–191. [Google Scholar]
- Crick, F.; Koch, C. A framework for consciousness. Nat. Neurosci. 2003, 6, 119–126. [Google Scholar] [CrossRef]
- Spence, C.; Parise, C. Prior-entry: A review. Conscious Cogn. 2010, 19, 364–379. [Google Scholar] [CrossRef]
- Gazzaley, A.; D’Esposito, M. Top-down modulation and normal aging. Ann. N. Y. Acad. Sci. 2007, 1097, 67–83. [Google Scholar] [CrossRef]
- Most, S.B.; Scholl, B.J.; Clifford, E.R.; Simons, D.J. What you see is what you set: Sustained inattentional blindness and the capture of awareness. Psychol. Rev. 2005, 112, 217–242. [Google Scholar] [CrossRef]
- Neisser, U. Cognitive Psychology; AppleCenturyCrofts: New York, NY, USA, 1967. [Google Scholar]
- Raymond, J.E.; Shapiro, K.L.; Arnell, K.M. Temporary suppression of visual processing in an RSVP task: An attentional blink? J. Exp. Psychol. Hum. Percept. Perform. 1992, 18, 849–860. [Google Scholar] [CrossRef]
- Morsella, E.; Ben-Zeev, A.; Lanska, M.; Bargh, J. The spontaneous thoughts of the night: How future tasks breed intrusive cognitions. Soc. Cogn. 2010, 28, 640–649. [Google Scholar]
- Gayet, S.; Paffen, C.L.E.; van der Stigchel, S. Information matching the content of visual working memory is prioritized for conscious accesss. Psychol. Sci. 2013. [Google Scholar] [CrossRef]
- Allport, D. Visual Attention; MIT: Cambridge, MA, USA, 1989. [Google Scholar]
- Baddeley, A. Working Memory, Thought and Action; Oxford University Press: Oxford, UK, 2007. [Google Scholar]
- Hamker, F.H. The reentry hypothesis: Linking eye movements to visual perception. J. Vis. 2003, 3, 808–816. [Google Scholar] [CrossRef]
- Morsella, E. The function of phenomenal states: Supramodular interaction theory. Psychol. Rev. 2005, 112, 1000–1021. [Google Scholar] [CrossRef]
- Neumann, O.; Hillsdale, N. Beyond Capacity: A Functional View of Attention. In Perspectives on Perception and Action; Heuer, H., Sanders, A.F., Eds.; Lawrence Erlbaum: Mahwah, NJ, USA, 1987; pp. 361–394. [Google Scholar]
- James, W. The Principles of Psychology; Dover: New York, NY, USA, 2007. [Google Scholar]
- Rosenbaum, D.A. The Cinderella of psychology: The neglect of motor control in the science of mental life and behavior. Am. Psychol. 2005, 60, 308–317. [Google Scholar] [CrossRef]
- The Mechanisms of Human Action: Introduction and Background. In Oxford Handbook of Human Action; Morsella, E.; Bargh, J.A.; Gollwitzer, P.M. (Eds.) Oxford University Press: Oxford, UK, 2009.
- Baars, B.J. Some essential differences between consciousness and attention, perception, and working memory. Conscious Cogn. 1997, 6, 363–371. [Google Scholar] [CrossRef]
- Hommel, B. Action control according to TEC (theory of event coding). Psychol. Res. 2009, 73, 512–526. [Google Scholar] [CrossRef]
- Hommel, B.; Müsseler, J.; Aschersleben, G.; Prinz, W. The Theory of Event Coding (TEC): A framework for perception and action planning. Behav. Brain Sci. 2001, 24, 849–878. [Google Scholar] [CrossRef]
- Harleß, E. Der apparat des willens. Z. Philos. Philos. Krit. 1861, 38, 50–73. (in Dutch). [Google Scholar]
- Lotze, R. Medizinische Psychologie oder Physiologie der Seele; (in German). Weidmannsche Buchhandlung: Frankfurt, German, 1852. [Google Scholar]
- Grossberg, S. The link between brain learning, attention, and consciousness. Conscious Cogn. 1999, 8, 1–44. [Google Scholar] [CrossRef]
- Kunde, W. Response-effect compatibility in manual choice reaction tasks. J. Exp. Psychol. Hum. Percept. Perform. 2001, 27, 387–394. [Google Scholar] [CrossRef]
- Gray, J. Consciousness: Creeping up on the Hard Problem; Oxford University Press: Oxford, UK, 2004. [Google Scholar]
- Schütz-Bosbach, S.; Prinz, W. Perceptual resonance: Action-induced modulation of perception. Trends Cogn. Sci. 2007, 11, 349–355. [Google Scholar] [CrossRef]
- Wohlschläger, A. Visual motion priming by invisible actions. Vis. Res. 2000, 40, 925–930. [Google Scholar] [CrossRef]
- Maruya, K.; Yang, E.; Blake, R. Voluntary action influences visual competition. Psychol. Sci. 2007, 18, 1090–1098. [Google Scholar] [CrossRef]
- Repp, B.H.; Knoblich, G. Action can affect auditory perception. Psychol. Sci. 2007, 18, 6–7. [Google Scholar] [CrossRef]
- Doesburg, S.M.; Green, J.J.; McDonald, J.J.; Ward, L.M. Rhythms of consciousness: Binocular rivalry reveals large-scale oscillatory network dynamics mediating visual perception. PLoS One 2009, 4, e6142. [Google Scholar] [CrossRef]
- Boly, M.; Garrido, M.I.; Gosseries, O.; Bruno, M.-A.; Boveroux, P.; Schnakers, C.; Massimini, M.; Litvak, V.; Laureys, S.; Friston, K. Preserved feedforward but impaired top-down processes in the vegetative state. Science 2011, 332, 858–862. [Google Scholar] [CrossRef]
- Cardoso-Leite, P.; Mamassian, P.; Schütz-Bosbach, S.; Waszak, F. A new look at sensory attenuation: Action-effect anticipation affects sensitivity, not response bias. Psychol. Sci. 2010, 21, 1740–1745. [Google Scholar] [CrossRef]
- Gray, J.R.; Bargh, J.A.; Morsella, E. Neural correlates of the essence of conscious conflict: fMRI of sustaining incompatible intentions. Exp. Brain Res. 2013, 229, 453–465. [Google Scholar] [CrossRef]
- Müller, J. Elements of Physiology; Lea and Blanchard: Philadelphia, PA, USA, 1843. [Google Scholar]
- Morsella, E.; Bargh, J. What is an output? Psychol. Inq. 2010, 21, 354–370. [Google Scholar] [CrossRef]
- Goodale, M.; Milner, D. Sight Unseen: An Exploration of Conscious and Unconscious Vision; Oxford University Press: Oxford, UK, 2004. [Google Scholar]
- Desmurget, M.; Reilly, K.T.; Richard, N.; Szathmari, A.; Mottolese, C.; Sirigu, A. Movement intention after parietal cortex stimulation in humans. Science 2009, 324, 811–813. [Google Scholar] [CrossRef]
- Tallon-Baudry, C. On the neural mechanisms subserving attention and consciousness and attention. Front. Psychol. 2012, 2, 397. [Google Scholar] [CrossRef]
- Chambon, V.; Wenke, D.; Fleming, S.M.; Prinz, W.; Haggard, P. An online neural substrate for sense of agency. Cereb. Cortex 2013, 23, 1031–1037. [Google Scholar] [CrossRef]
- Christensen, M.S.; Lundbye-Jensen, J.; Geertsen, S.S.; Petersen, T.H.; Paulson, O.B.; Nielsen, J.B. Premotor cortex modulates somatosensory cortex during voluntary movements without proprioceptive feedback. Nat. Neurosci. 2007, 10, 417–419. [Google Scholar]
- Neisser, U. Cognition and Reality: Principles and Implications of Cognitive Psychology; W. H. Freeman: San Francisco, CA, USA, 1976. [Google Scholar]
- Knuf, L.; Aschersleben, G.; Prinz, W. An analysis of ideomotor action. J. Exp. Psychol. Gen. 2001, 130, 779–798. [Google Scholar] [CrossRef]
- Morsella, E.; Krauss, R.M. The role of gestures in spatial working memory and speech. Am. J. Psychol. 2004, 117, 411–424. [Google Scholar] [CrossRef]
- Alais, D.; Blake, R. Binocular Rival; MIT: Cambridge, MA, USA, 2005. [Google Scholar]
- Breitmeyer, B.G.; Ögmen, H. Visual Masking: Time Slices through Conscious and Unconscious Vision; Oxford University Press: Oxford, UK, 2006. [Google Scholar]
- Mack, A.; Rock, I. Inattentional Blindness; The MIT Press: Cambridge, MA, USA, 1998. [Google Scholar]
- Wolfe, J.M. Reversing ocular dominance and suppression in a single flash. Vis. Res. 1984, 24, 471–478. [Google Scholar] [CrossRef]
- Pessiglione, M.; Petrovic, P.; Daunizeau, J.; Palminteri, S.; Dolan, R.J.; Frith, C.D. Subliminal instrumental conditioning demonstrated in the human brain. Neuron 2008, 59, 561–567. [Google Scholar] [CrossRef]
- Tzelgov, J.; Porat, Z.; Henik, A. Automaticity and consciousness: Is perceiving the word necessary for reading it? Am. J. Psychol. 1997, 110, 429–448. [Google Scholar] [CrossRef]
- Gray, J. The contents of consciousness: A neuropsychological conjecture. Behav. Brain Sci. 1995, 18, 659–676. [Google Scholar] [CrossRef]
- Morsella, E.; Lanska, M.; Berger, C.; Gazzaley, A. Indirect cognitive control through top-down activation of perceptual symbols. Eur. J. Soc. Psychol. 2009, 39, 1173–1177. [Google Scholar] [CrossRef]
- Fukuda, K.; Vogel, E.K. Human variation in overriding attentional capture. J. Neurosci. 2009, 29, 8726–8733. [Google Scholar] [CrossRef]
- Cohen, J.; MacWhinney, B.; Flatt, M.; Provost, J. PsyScope: A new graphic interactive environment for designing psychology experiments. Behav. Res. Methods Instrum. Comput. 1993, 25, 257–271. [Google Scholar] [CrossRef]
- Zajonc, R. Attitudinal effects of mere exposure. J. Pers. Soc. Psychol. 1968, 9, 1–27. [Google Scholar] [CrossRef]
- Bargh, J.A.; Chartrand, T.L. The Mind in the Middle: A Practical Guide to Priming and Automaticity Research. In Handbook of Research Methods in Social and Personality Psychology; Cambridge University Press: Cambridge, UK, 2000. [Google Scholar]
- Eitam, B.; Yeshurun, Y.; Hassan, K. Blinded by irrelevance: Pure irrelevance induced “blindness.”. J. Exp. Psychol. Hum. Percept. Perform. 2013, 39, 611–615. [Google Scholar] [CrossRef]
- Block, N. Perceptual consciousness overflows cognitive access. Trends Cogn. Sci. 2011, 15, 567–575. [Google Scholar] [CrossRef]
- Blakemore, S.-J.; Wolpert, D.; Frith, C. Why you can’t tickle yourself? NeuroReport 2000, 11, r11–r16. [Google Scholar] [CrossRef]
- Müsseler, J.; Hommel, B. Blindness to response-compatible stimuli. J. Exp. Psychol. Hum. Percept. Perform. 1997, 23, 861–887. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Dennehy, T.C.; Cooper, S.; Molapour, T.; Morsella, E. Is There Release from Masking from Isomorphism between Perception and Action? Brain Sci. 2014, 4, 220-239. https://doi.org/10.3390/brainsci4020220
Dennehy TC, Cooper S, Molapour T, Morsella E. Is There Release from Masking from Isomorphism between Perception and Action? Brain Sciences. 2014; 4(2):220-239. https://doi.org/10.3390/brainsci4020220
Chicago/Turabian StyleDennehy, Tara C., Shanna Cooper, Tanaz Molapour, and Ezequiel Morsella. 2014. "Is There Release from Masking from Isomorphism between Perception and Action?" Brain Sciences 4, no. 2: 220-239. https://doi.org/10.3390/brainsci4020220