
Long-Term Task- and Dopamine-Dependent Dynamics of Subthalamic Local Field Potentials in Parkinson’s Disease

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Recruitment
2.2. DBS Surgery
2.3. Data Collection
2.4. Behavioral Tasks
2.5. Analysis
2.6. Statistics
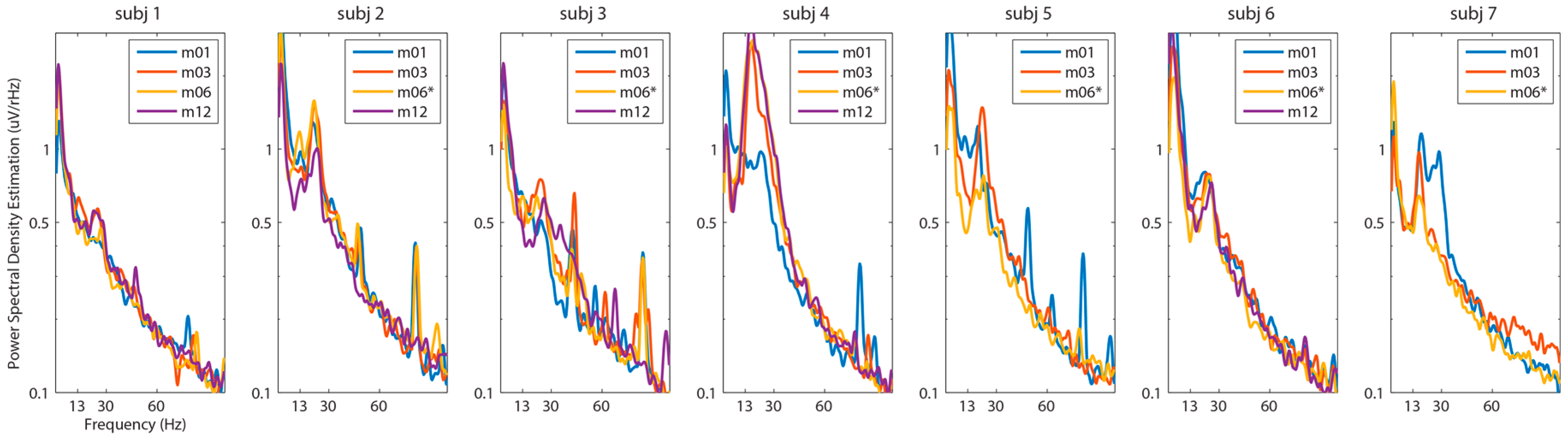
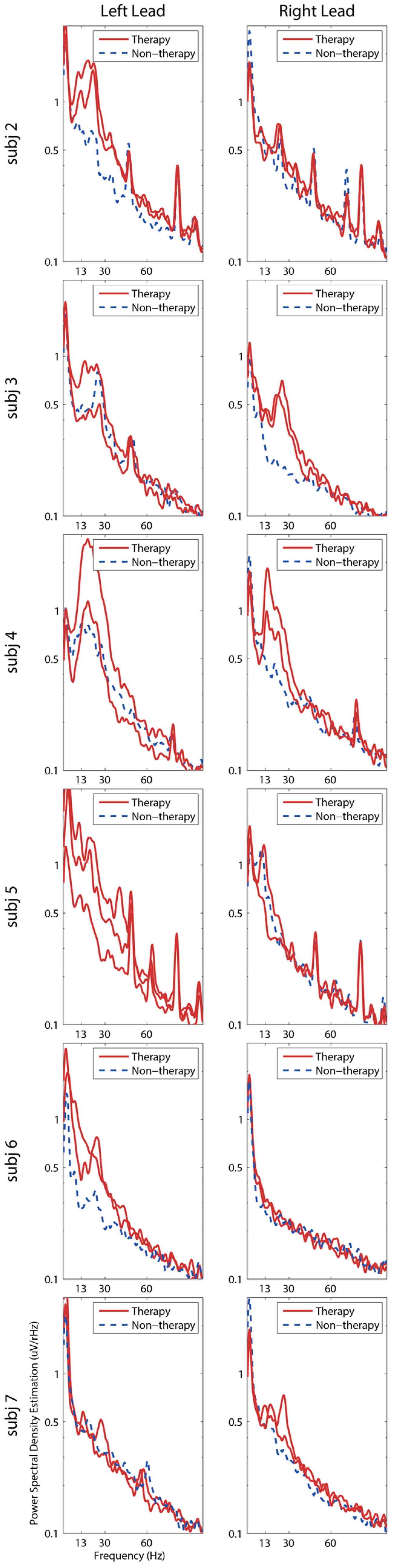
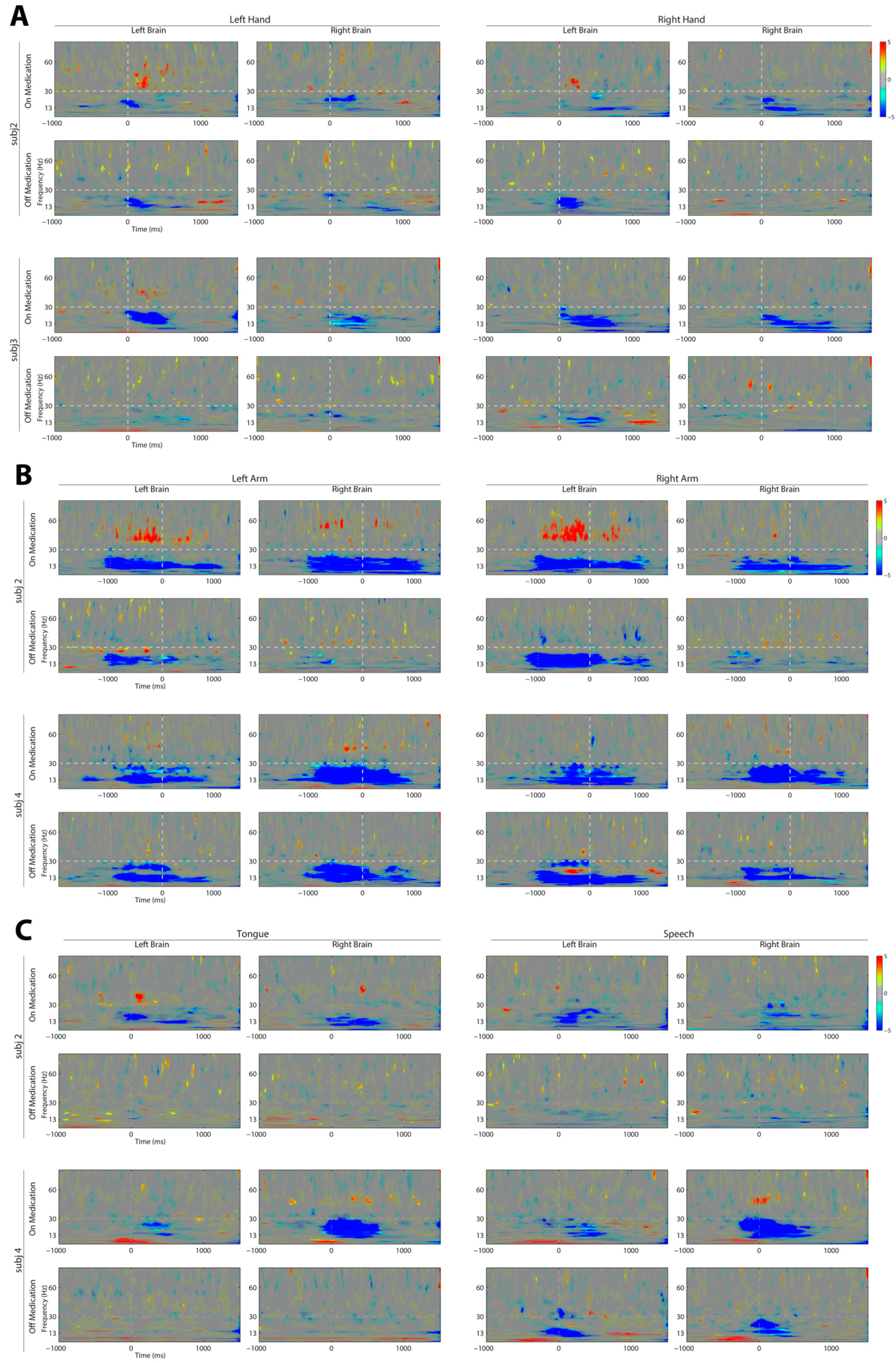
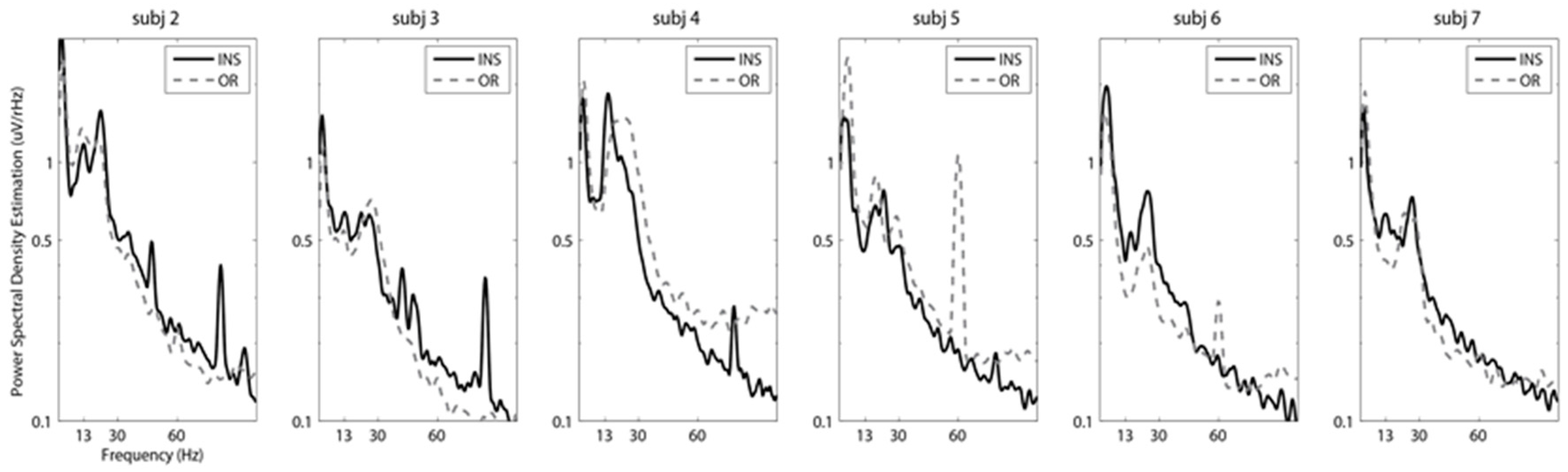
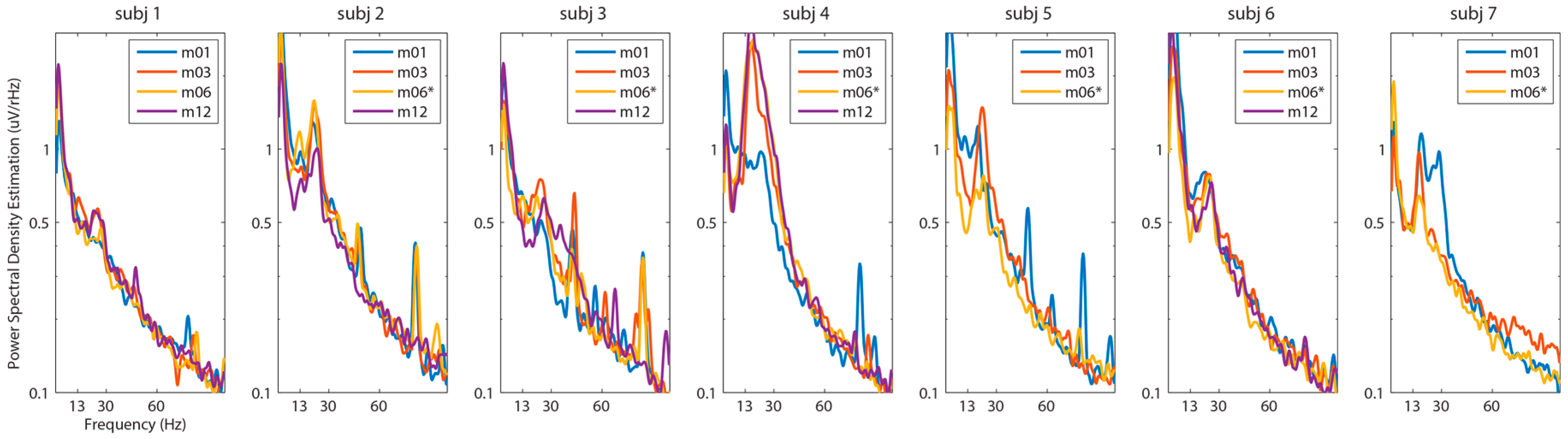
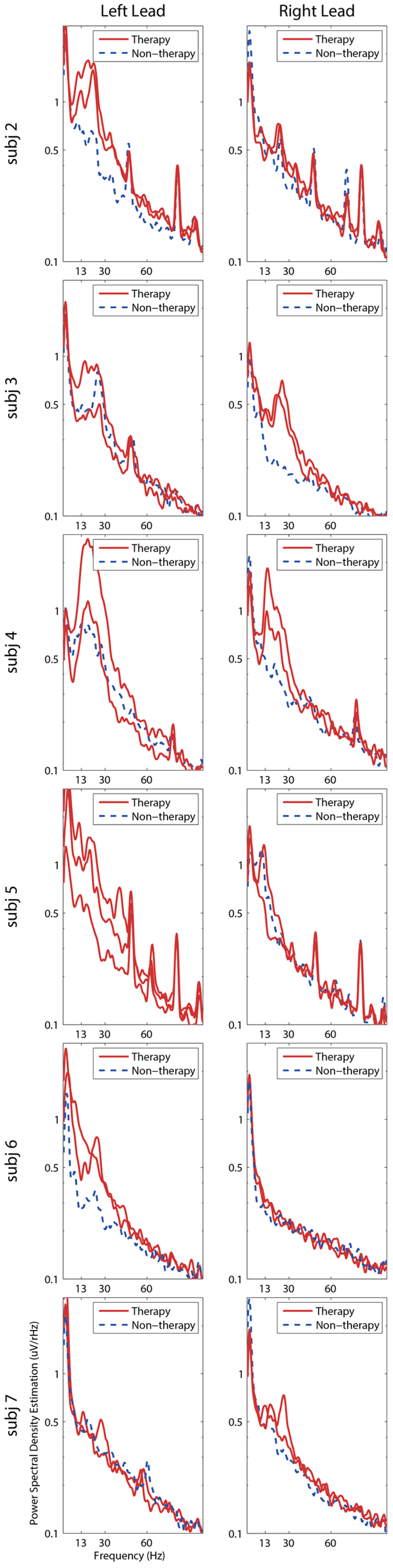
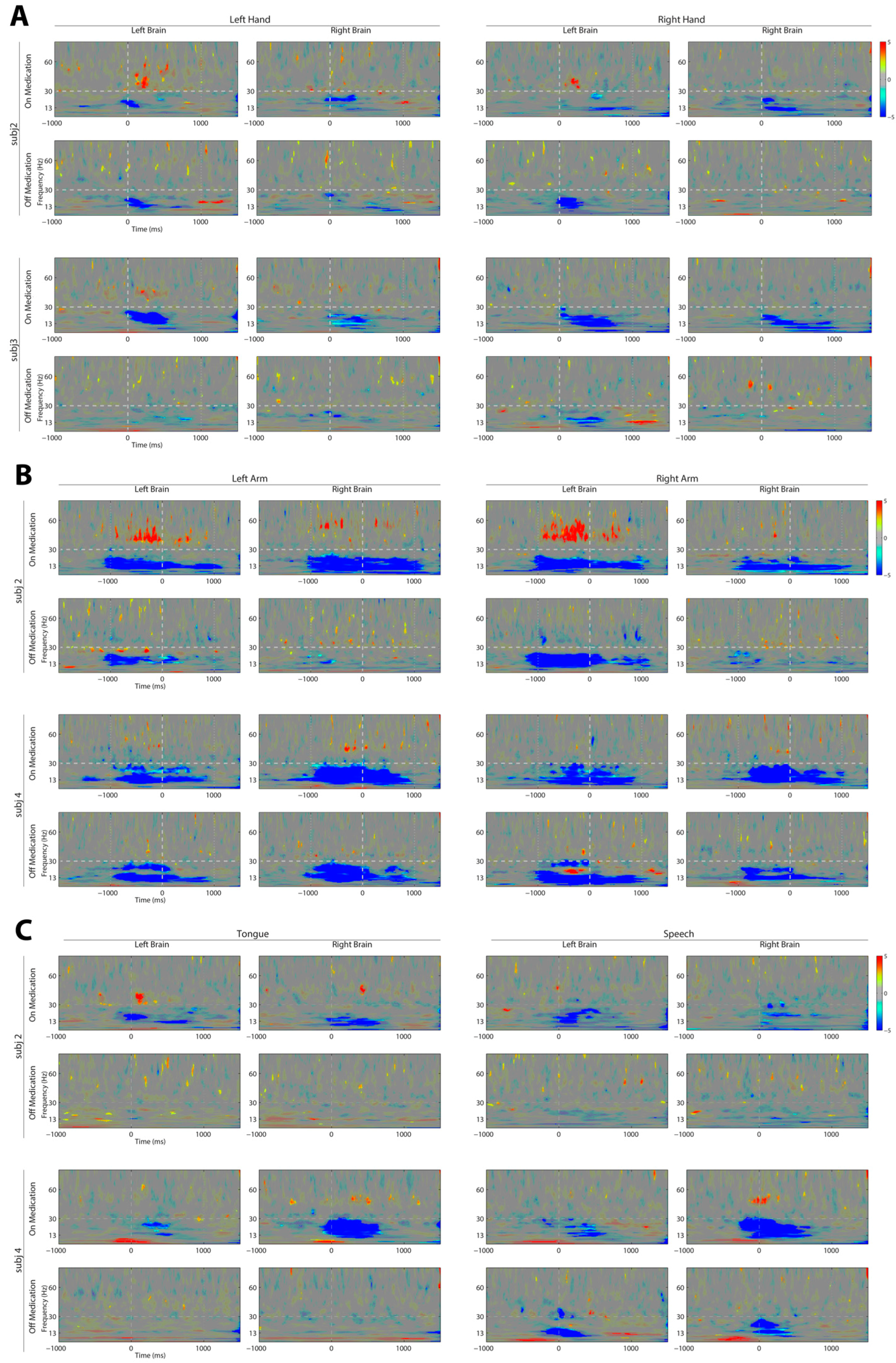
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Li, Q.; Qian, Z.-M.; Arbuthnott, G.W.; Ke, Y.; Yung, W.-H. Cortical effects of deep brain stimulation: Implications for pathogenesis and treatment of Parkinson disease. JAMA Neurol. 2014, 71, 100–103. [Google Scholar] [CrossRef] [PubMed]
- Bordini, B.J.; Garg, A.; Gallagher, C.L.; Bell, B.; Garell, P.C. Neuropsychological effects of bilateral deep brain stimulation of the subthalamic nucleus in Parkinson’s disease. Stereotact. Funct. Neurosurg. 2007, 85, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Tripoliti, E.; Zrinzo, L.; Martinez-Torres, I.; Frost, E.; Pinto, S.; Foltynie, T.; Holl, E.; Petersen, E.; Roughton, M.; Hariz, M.I.; et al. Effects of subthalamic stimulation on speech of consecutive patients with Parkinson disease. Neurology 2011, 76, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Morrison, C.E.; Borod, J.C.; Perrine, K.; Beric, A.; Brin, M.F.; Rezai, A.; Kelly, P.; Sterio, D.; Germano, I.; Weisz, D.; et al. Neuropsychological functioning following bilateral subthalamic nucleus stimulation in Parkinson’s disease. Arch. Clin. Neuropsychol. 2004, 19, 165–181. [Google Scholar] [CrossRef]
- Fleury, V.; Pollak, P.; Gere, J.; Tommasi, G.; Romito, L.; Combescure, C.; Bardinet, E.; Chabardes, S.; Momjian, S.; Krainik, A.; et al. Subthalamic stimulation may inhibit the beneficial effects of levodopa on akinesia and gait. Mov. Disord. 2016, 31, 1389–1397. [Google Scholar] [CrossRef] [PubMed]
- Hariz, M.I.; Rehncrona, S.; Quinn, N.P.; Speelman, J.D.; Wensing, C.; Multicentre Advanced Parkinson’s Disease Deep Brain Stimulation Group. Multicenter study on deep brain stimulation in Parkinson’s disease: An independent assessment of reported adverse events at 4 years. Mov. Disord. 2008, 23, 416–421. [Google Scholar] [CrossRef] [PubMed]
- Arlotti, M.; Rossi, L.; Rosa, M.; Marceglia, S.; Priori, A. An external portable device for adaptive deep brain stimulation (aDBS) clinical research in advanced Parkinson’s disease. Med. Eng. Phys. 2016, 38, 498–505. [Google Scholar] [CrossRef] [PubMed]
- Beudel, M.; Little, S.; Pogosyan, A.; Ashkan, K.; Foltynie, T.; Limousin, P.; Zrinzo, L.; Hariz, M.; Bogdanovic, M.; Cheeran, B.; et al. Tremor Reduction by Deep Brain Stimulation Is Associated With Gamma Power Suppression in Parkinson’s Disease. Neuromodulation 2015, 18, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Mahlknecht, P.; Limousin, P.; Foltynie, T. Deep brain stimulation for movement disorders: Update on recent discoveries and outlook on future developments. J. Neurol. 2015, 262, 2583–2595. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, A.; Zamani, M.; Bayford, R.; Demosthenous, A. Patient specific Parkinson’s disease detection for adaptive deep brain stimulation. In Proceedings of the 2015 37th Annual International Conference of the IEEE Engineering in Medicine and Biology Society, Milan, Italy, 25–29 August 2015; pp. 1528–1531.
- Hebb, A.O.; Zhang, J.J.; Mahoor, M.H.; Tsiokos, C.; Matlack, C.; Chizeck, H.J.; Pouratian, N. Creating the Feedback Loop. Neurosurg. Clin. N. Am. 2014, 25, 187–204. [Google Scholar] [CrossRef] [PubMed]
- Giannicola, G.; Rosa, M.; Servello, D.; Menghetti, C.; Carrabba, G.; Pacchetti, C.; Zangaglia, R.; Cogiamanian, F.; Scelzo, E.; Marceglia, S.; et al. Subthalamic local field potentials after seven-year deep brain stimulation in Parkinson’s disease. Exp. Neurol. 2012, 237, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Friston, K.J.; Bastos, A.M.; Pinotsis, D.; Litvak, V. LFP and oscillations-what do they tell us? Curr. Opin. Neurobiol. 2015, 31, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Little, S.; Brown, P. What brain signals are suitable for feedback control of deep brain stimulation in Parkinson’s disease? Ann. N. Y. Acad. Sci. 2012, 1265, 9–24. [Google Scholar] [CrossRef] [PubMed]
- Gmel, G.E.; Parker, J.L.; Hamilton, T.J. A new biomarker for closed-loop deep brain stimulation in the subthalamic nucleus for patients with Parkinson’s disease. In Proceedings of the 2014 IEEE Biomedical Circuits and Systems Conference (BioCAS), Lausanne, Switzerland, 22–24 October 2014; pp. 500–503.
- Timmermann, L.; Florin, E. Parkinson’s disease and pathological oscillatory activity: Is the beta band the bad guy?—New lessons learned from low-frequency deep brain stimulation. Exp. Neurol. 2012, 233, 123–125. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.; Tijssen, M.; Van Bruggen, G.; Bosch, A.; Insola, A.; Di Lazzaro, V.; Mazzone, P.; Oliviero, A.; Quartarone, A.; Speelman, H.; et al. Dopamine-dependent changes in the functional connectivity between basal ganglia and cerebral cortex in humans. Brain J. Neurol. 2002, 125, 1558–1569. [Google Scholar] [CrossRef]
- Brown, P.; Oliviero, A.; Mazzone, P.; Insola, A.; Tonali, P.; Lazzaro, V.D. Dopamine Dependency of Oscillations between Subthalamic Nucleus and Pallidum in Parkinson’s Disease. J. Neurosci. 2001, 21, 1033–1038. [Google Scholar] [PubMed]
- Loukas, C.; Brown, P. Online prediction of self-paced hand-movements from subthalamic activity using neural networks in Parkinson’s disease. J. Neurosci. Methods 2004, 137, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Niketeghad, S.; Hebb, A.O.; Nedrud, J.; Hanrahan, S.J.; Mahoor, M.H. Single trial behavioral task classification using subthalamic nucleus local field potential signals. In Proceedings of the 2014 36th Annual International Conference of the IEEE Engineering in Medicine and Biology Society, Chicago, IL, USA, 26–30 August 2014; pp. 3793–3796.
- Hebb, A.O.; Darvas, F.; Miller, K.J. Transient and state modulation of beta power in human subthalamic nucleus during speech production and finger movement. Neuroscience 2012, 202, 218–233. [Google Scholar] [CrossRef] [PubMed]
- Flint, R.D.; Wright, Z.A.; Scheid, M.R.; Slutzky, M.W. Long term, stable brain machine interface performance using local field potentials and multiunit spikes. J. Neural Eng. 2013, 10, 56005. [Google Scholar] [CrossRef] [PubMed]
- Abosch, A.; Lanctin, D.; Onaran, I.; Eberly, L.; Spaniol, M.; Ince, N.F. Long-term recordings of local field potentials from implanted deep brain stimulation electrodes. Neurosurgery 2012, 71, 804–814. [Google Scholar] [CrossRef] [PubMed]
- Quinn, E.J.; Blumenfeld, Z.; Velisar, A.; Koop, M.M.; Shreve, L.A.; Trager, M.H.; Hill, B.C.; Kilbane, C.; Henderson, J.M.; Brontë-Stewart, H. Beta oscillations in freely moving Parkinson’s subjects are attenuated during deep brain stimulation. Mov. Disord. 2015, 30, 1750–1758. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, M.-J.; Vidailhet, M.; Loc’h, C.; Dupel, C.; Nguyen, J.P.; Ponchant, M.; Dollé, F.; Peschanski, M.; Hantraye, P.; Cesaro, P.; et al. Dopaminergic function and dopamine transporter binding assessed with positron emission tomography in Parkinson disease. Arch. Neurol. 2002, 59, 580–586. [Google Scholar] [CrossRef] [PubMed]
- Bronte-Stewart, H.; Barberini, C.; Koop, M.M.; Hill, B.C.; Henderson, J.M.; Wingeier, B. The STN beta-band profile in Parkinson’s disease is stationary and shows prolonged attenuation after deep brain stimulation. Exp. Neurol. 2009, 215, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Levy, R.; Ashby, P.; Hutchison, W.D.; Lang, A.E.; Lozano, A.M.; Dostrovsky, J.O. Dependence of subthalamic nucleus oscillations on movement and dopamine in Parkinson’s disease. Brain J. Neurol. 2002, 125, 1196–1209. [Google Scholar] [CrossRef]
- Marceglia, S.; Foffani, G.; Bianchi, A.M.; Baselli, G.; Tamma, F.; Egidi, M.; Priori, A. Dopamine-dependent non-linear correlation between subthalamic rhythms in Parkinson’s disease. J. Physiol. 2006, 571, 579–591. [Google Scholar] [CrossRef] [PubMed]
- Weinberger, M.; Mahant, N.; Hutchison, W.D.; Lozano, A.M.; Moro, E.; Hodaie, M.; Lang, A.E.; Dostrovsky, J.O. Beta oscillatory activity in the subthalamic nucleus and its relation to dopaminergic response in Parkinson’s disease. J. Neurophysiol. 2006, 96, 3248–3256. [Google Scholar] [CrossRef] [PubMed]
- Marsden, J.F.; Limousin-Dowsey, P.; Ashby, P.; Pollak, P.; Brown, P. Subthalamic nucleus, sensorimotor cortex and muscle interrelationships in Parkinson’s disease. Brain J. Neurol. 2001, 124, 378–388. [Google Scholar] [CrossRef]
- De Hemptinne, C.; Ryapolova-Webb, E.S.; Air, E.L.; Garcia, P.A.; Miller, K.J.; Ojemann, J.G.; Ostrem, J.L.; Galifianakis, N.B.; Starr, P.A. Exaggerated phase-amplitude coupling in the primary motor cortex in Parkinson disease. Proc. Natl. Acad. Sci. USA 2013, 110, 4780–4785. [Google Scholar] [CrossRef] [PubMed]
- Foffani, G.; Priori, A.; Egidi, M.; Rampini, P.; Tamma, F.; Caputo, E.; Moxon, K.A.; Cerutti, S.; Barbieri, S. 300-Hz subthalamic oscillations in Parkinson’s disease. Brain J. Neurol. 2003, 126, 2153–2163. [Google Scholar] [CrossRef] [PubMed]
- Cassidy, M.; Mazzone, P.; Oliviero, A.; Insola, A.; Tonali, P.; Di Lazzaro, V.; Brown, P. Movement-related changes in synchronization in the human basal ganglia. Brain J. Neurol. 2002, 125, 1235–1246. [Google Scholar] [CrossRef]
- Jenkinson, N.; Brown, P. New insights into the relationship between dopamine, beta oscillations and motor function. Trends Neurosci. 2011, 34, 611–618. [Google Scholar] [CrossRef] [PubMed]
- Paradiso, G.; Cunic, D.; Saint-Cyr, J.A.; Hoque, T.; Lozano, A.M.; Lang, A.E.; Chen, R. Involvement of human thalamus in the preparation of self-paced movement. Brain J. Neurol. 2004, 127, 2717–2731. [Google Scholar] [CrossRef] [PubMed]
- Meziane, H.B.; Moisello, C.; Perfetti, B.; Kvint, S.; Isaias, I.U.; Quartarone, A.; Di Rocco, A.; Ghilardi, M.F. Movement preparation and bilateral modulation of beta activity in aging and Parkinson’s disease. PLoS ONE 2015, 10, e0114817. [Google Scholar] [CrossRef] [PubMed]
- Doyle, L.M.F.; Kühn, A.A.; Hariz, M.; Kupsch, A.; Schneider, G.-H.; Brown, P. Levodopa-induced modulation of subthalamic beta oscillations during self-paced movements in patients with Parkinson’s disease. Eur. J. Neurosci. 2005, 21, 1403–1412. [Google Scholar] [CrossRef] [PubMed]
- Pfurtscheller, G.; Lopes da Silva, F.H. Event-related EEG/MEG synchronization and desynchronization: Basic principles. Clin. Neurophysiol. 1999, 110, 1842–1857. [Google Scholar] [CrossRef]
- Chomiak, T.; Pereira, F.V.; Meyer, N.; de Bruin, N.; Derwent, L.; Luan, K.; Cihal, A.; Brown, L.A.; Hu, B. A new quantitative method for evaluating freezing of gait and dual-attention task deficits in Parkinson’s disease. J. Neural Transm. 2015, 122, 1523–1531. [Google Scholar] [CrossRef] [PubMed]
- Nocera, J.R.; Roemmich, R.; Elrod, J.; Altmann, L.J.P.; Hass, C.J. Effects of cognitive task on gait initiation in Parkinson disease: Evidence of motor prioritization? J. Rehabil. Res. Dev. 2013, 50, 699–708. [Google Scholar] [CrossRef] [PubMed]
- Wessel, J.R.; Jenkinson, N.; Brittain, J.-S.; Voets, S.H.E.M.; Aziz, T.Z.; Aron, A.R. Surprise disrupts cognition via a fronto-basal ganglia suppressive mechanism. Nat. Commun. 2016, 7, 11195. [Google Scholar] [CrossRef] [PubMed]
- Powell, D.; Hanson, N.; Threlkeld, A.J.; Fang, X.; Xia, R. Enhancement of parkinsonian rigidity with contralateral hand activation. Clin. Neurophysiol. 2011, 122, 1595–1601. [Google Scholar] [CrossRef] [PubMed]
- Rektor, I.; Baláz, M.; Bocková, M. Cognitive activities in the subthalamic nucleus. Invasive studies. Parkinsonism Relat. Disord. 2009, 15 (Suppl. S3), S83–S86. [Google Scholar] [CrossRef]
- Stanslaski, S.; Afshar, P.; Cong, P.; Giftakis, J.; Stypulkowski, P.; Carlson, D.; Linde, D.; Ullestad, D.; Avestruz, A.-T.; Denison, T. Design and validation of a fully implantable, chronic, closed-loop neuromodulation device with concurrent sensing and stimulation. IEEE Trans. Neural Syst. Rehabil. Eng. 2012, 20, 410–421. [Google Scholar] [CrossRef] [PubMed]
- Connolly, A.T.; Muralidharan, A.; Hendrix, C.; Johnson, L.; Gupta, R.; Stanslaski, S.; Denison, T.; Baker, K.B.; Vitek, J.L.; Johnson, M.D. Local field potential recordings in a non-human primate model of Parkinsons disease using the Activa PC + S neurostimulator. J. Neural Eng. 2015, 12, 66012. [Google Scholar] [CrossRef] [PubMed]
- Schalk, G.; McFarland, D.J.; Hinterberger, T.; Birbaumer, N.; Wolpaw, J.R. BCI2000: A general-purpose brain-computer interface (BCI) system. IEEE Trans. Biomed. Eng. 2004, 51, 1034–1043. [Google Scholar] [CrossRef] [PubMed]
- Oswal, A.; Litvak, V.; Brücke, C.; Huebl, J.; Schneider, G.-H.; Kühn, A.A.; Brown, P. Cognitive Factors Modulate Activity within the Human Subthalamic Nucleus during Voluntary Movement in Parkinson’s Disease. J. Neurosci. 2013, 33, 15815–15826. [Google Scholar] [CrossRef] [PubMed]
- Waldert, S.; Vigneswaran, G.; Philipp, R.; Lemon, R.N.; Kraskov, A. Modulation of the Intracortical LFP during Action Execution and Observation. J. Neurosci. 2015, 35, 8451–8461. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.A.; Nebeck, S.D.; Muralidharan, A.; Johnson, M.D.; Baker, K.B.; Vitek, J.L. Closed-Loop Deep Brain Stimulation Effects on Parkinsonian Motor Symptoms in a Non-Human Primate—Is Beta Enough? Brain Stimul. 2016, 9, 892–896. [Google Scholar] [CrossRef] [PubMed]
- Watson, P.; Montgomery, E.B. The relationship of neuronal activity within the sensori-motor region of the subthalamic nucleus to speech. Brain Lang. 2006, 97, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Van der Walt, S.; Colbert, S.C.; Varoquaux, G. The NumPy Array: A Structure for Efficient Numerical Computation. Comput. Sci. Eng. 2011, 13, 22–30. [Google Scholar] [CrossRef]
- Jones, E.; Oliphant, T.; Peterson, P. SciPy: Open Source Scientific Tools for Python. Available online: http://www.scipy.org/ (accessed on 5 November 2014).
- Allen, D.P.; Stegemöller, E.L.; Zadikoff, C.; Rosenow, J.M.; Mackinnon, C.D. Suppression of deep brain stimulation artifacts from the electroencephalogram by frequency-domain Hampel filtering. Clin. Neurophysiol. 2010, 121, 1227–1232. [Google Scholar] [CrossRef] [PubMed]
- Erez, Y.; Tischler, H.; Moran, A.; Bar-Gad, I. Generalized framework for stimulus artifact removal. J. Neurosci. Methods 2010, 191, 45–59. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, T.; Elder, C.M.; Vitek, J.L. A template subtraction method for stimulus artifact removal in high-frequency deep brain stimulation. J. Neurosci. Methods 2002, 113, 181–186. [Google Scholar] [CrossRef]
- Thompson, J.A.; Lanctin, D.; Ince, N.F.; Abosch, A. Clinical Implications of Local Field Potentials for Understanding and Treating Movement Disorders. Stereotact. Funct. Neurosurg. 2014, 92, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Holdefer, R.N.; Cohen, B.A.; Greene, K.A. Intraoperative local field recording for deep brain stimulation in Parkinson’s disease and essential tremor. Mov. Disord. 2010, 25, 2067–2075. [Google Scholar] [CrossRef] [PubMed]
- Alegre, M.; Alonso-Frech, F.; Rodríguez-Oroz, M.C.; Guridi, J.; Zamarbide, I.; Valencia, M.; Manrique, M.; Obeso, J.A.; Artieda, J. Movement-related changes in oscillatory activity in the human subthalamic nucleus: Ipsilateral vs. contralateral movements. Eur. J. Neurosci. 2005, 22, 2315–2324. [Google Scholar] [CrossRef] [PubMed]
- Cheung, T.; Nuño, M.; Hoffman, M.; Katz, M.; Kilbane, C.; Alterman, R.; Tagliati, M. Longitudinal Impedance Variability in Patients with Chronically Implanted DBS Devices. Brain Stimul. 2013, 6, 746–751. [Google Scholar] [CrossRef] [PubMed]
- Beudel, M.; Brown, P. Adaptive deep brain stimulation in Parkinson’s disease. Parkinsonism Relat. Disord. 2016, 22, S123–S126. [Google Scholar] [CrossRef] [PubMed]
- Swann, N.C.; de Hemptinne, C.; Miocinovic, S.; Qasim, S.; Wang, S.S.; Ziman, N.; Ostrem, J.L.; San Luciano, M.; Galifianakis, N.B.; Starr, P.A. Gamma Oscillations in the Hyperkinetic State Detected with Chronic Human Brain Recordings in Parkinson’s Disease. J. Neurosci. 2016, 36, 6445–6458. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Wang, J.; Deng, B.; Wei, X.; Yu, H.; Li, H.; Fietkiewicz, C.; Loparo, K. Closed-loop Control of Tremor-predominant Parkinsonian State Based on Parameter Estimation: A Computational Study. IEEE Trans. Neural Syst. Rehabil. Eng. 2016, 24, 1109–1121. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subject | Age (years) | Sex | Handedness | Pre-op UPDRS III 1 | Post-op UPDRS III 2 | Diagnosis and Predominant Symptom | Clinical DBS Settings Left STN | Clinical DBS Settings Right STN | Bipolar Channels for Behavioral Recording |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 59 | F | R | 37/22 | 35/24/16/13 | PD, bradykinesia | E3⁺, E2−, 3.6 V, 60 µs, 130 Hz | C⁺, E1−, 3.6 V, 60 us, 130 Hz | L: 2/3, R: 0/1 |
| 2 | 65 | M | R | 48/14 | 49/23/17/16 | PD, rigidity | E1⁺, E2−, 2.6 V, 60 µs, 150 Hz | C⁺, E2−, 2.4 V, 60 us, 150 Hz | L: 2/3, R: 2/3 |
| 3 | 63 | F | R | 23/6 | 32/25/24/16 | PD, dyskinesias | E3⁺, E2−, 2.4 V, 60 µs, 130 Hz | E3⁺, E2−, 2.5 V, 60 us, 130 Hz | L: 1/2, R: 2/3 |
| 4 | 71 | M | R | 31/7 | 38/37/21/20 | PD, gait disturbance | E3⁺, E2−, 3.6 V, 60 µs, 130 Hz | E3⁺, E2−, 3.6 V, 60 µs, 130 Hz | L: 2/3, R: 2/3 |
| 5 | 44 | M | R | 38/20 | 36/16/29/11 | PD, tremor | C⁺, E1−, E2−, 2.9 V, 60 µs, 130 Hz | C⁺, E2−, 3.2 V, 60 µs, 130 Hz | L: 1/2, R: 2/3 |
| 6 | 62 | M | L | 31/20 | 24/16/21/14 | PD, tremor | C⁺, E1−, 2.6 V, 60 µs, 130 Hz | C⁺, E1−, 3.9 V, 70 µs, 135 Hz | L: 1/2, R: 2/3 |
| 7 | 68 | M | R | 67/40 | -/-/-/- | PD, bradykinesia | C⁺, E1−, 2.2 V, 60 µs, 130 Hz | C⁺, E2−, 2.0 V, 60 µs, 130 Hz | L: 2/3, R: 2/3 |
| OR | INS | |
|---|---|---|
| Input impedance | >100 MOhm | 1 MOhm |
| Range | −250 mV–250 mV | −10 V–10 V |
| Filters used | 0.5–2000 Hz | 0.5–100 Hz |
| Sampling Rate | 4800 Hz | 422 Hz |
| Noise Floor | <0.3 µV RMS (0.1–10 Hz) | Min signal to detect 1 μV RMS differential with noise floor <0.3 µV RMS |
| Recording Session | Intra-Operative | 1 Month | 3 Month | 6 Month | 12 Month |
|---|---|---|---|---|---|
| Recording Type | OR | INS | INS | INS | INS |
| Medication State | Off | On | On | Off | On |
| Non-behavioral Montage Recording | 6 bipolar channels | 6 bipolar channels | 6 bipolar channels | 6 bipolar channels | 6 bipolar channels |
| Behavioral Recording | - | - | 2 bipolar channels | 2 bipolar channels | - |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hanrahan, S.J.; Nedrud, J.J.; Davidson, B.S.; Farris, S.; Giroux, M.; Haug, A.; Mahoor, M.H.; Silverman, A.K.; Zhang, J.J.; Hebb, A.O. Long-Term Task- and Dopamine-Dependent Dynamics of Subthalamic Local Field Potentials in Parkinson’s Disease. Brain Sci. 2016, 6, 57. https://doi.org/10.3390/brainsci6040057
Hanrahan SJ, Nedrud JJ, Davidson BS, Farris S, Giroux M, Haug A, Mahoor MH, Silverman AK, Zhang JJ, Hebb AO. Long-Term Task- and Dopamine-Dependent Dynamics of Subthalamic Local Field Potentials in Parkinson’s Disease. Brain Sciences. 2016; 6(4):57. https://doi.org/10.3390/brainsci6040057
Chicago/Turabian StyleHanrahan, Sara J., Joshua J. Nedrud, Bradley S. Davidson, Sierra Farris, Monique Giroux, Aaron Haug, Mohammad H. Mahoor, Anne K. Silverman, Jun Jason Zhang, and Adam Olding Hebb. 2016. "Long-Term Task- and Dopamine-Dependent Dynamics of Subthalamic Local Field Potentials in Parkinson’s Disease" Brain Sciences 6, no. 4: 57. https://doi.org/10.3390/brainsci6040057