l-Carnitine Modulates Epileptic Seizures in Pentylenetetrazole-Kindled Rats via Suppression of Apoptosis and Autophagy and Upregulation of Hsp70

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Study Groups
2.3. Pentylenetetrazole-Kindled Rat Model and Scoring of Epileptic Seizure
2.4. Euthanasia and Collection of Brain Samples
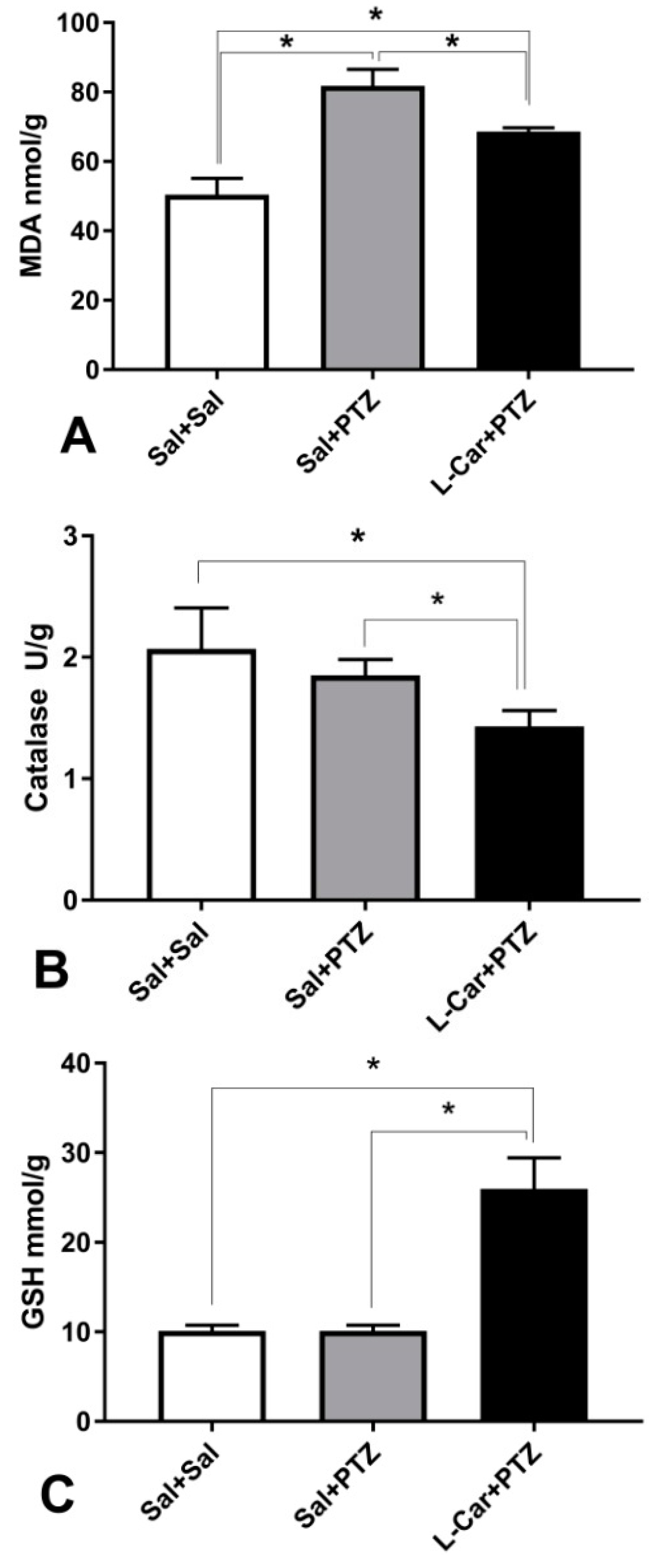
2.5. Assay of Lipid Peroxidations Marker (MDA) and Antioxidants (GSH Activity) and Catalase in Brain Tissues
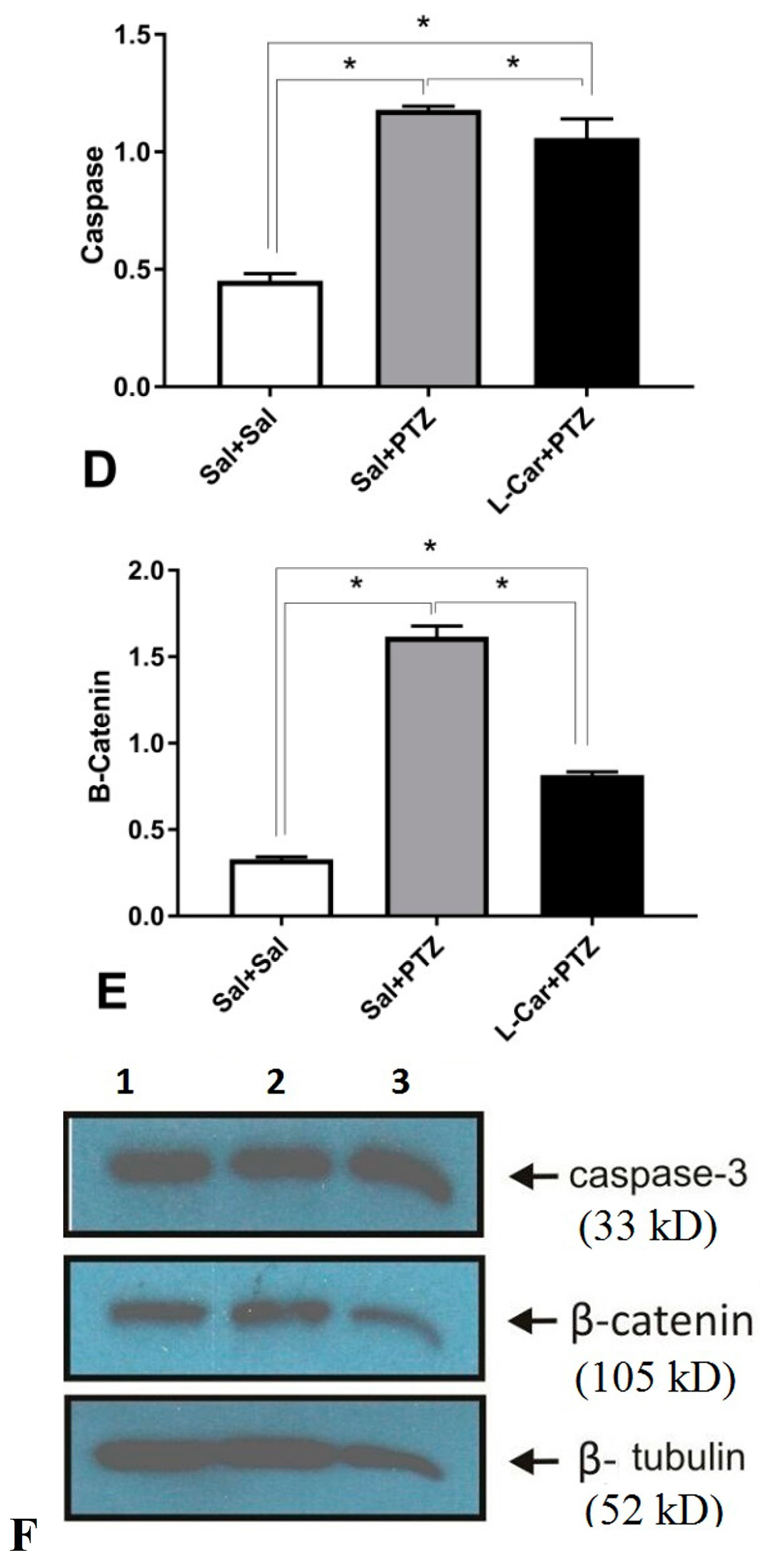
2.6. Gel Electrophoresis and Western Blotting for Caspase-3 and β-Catenin
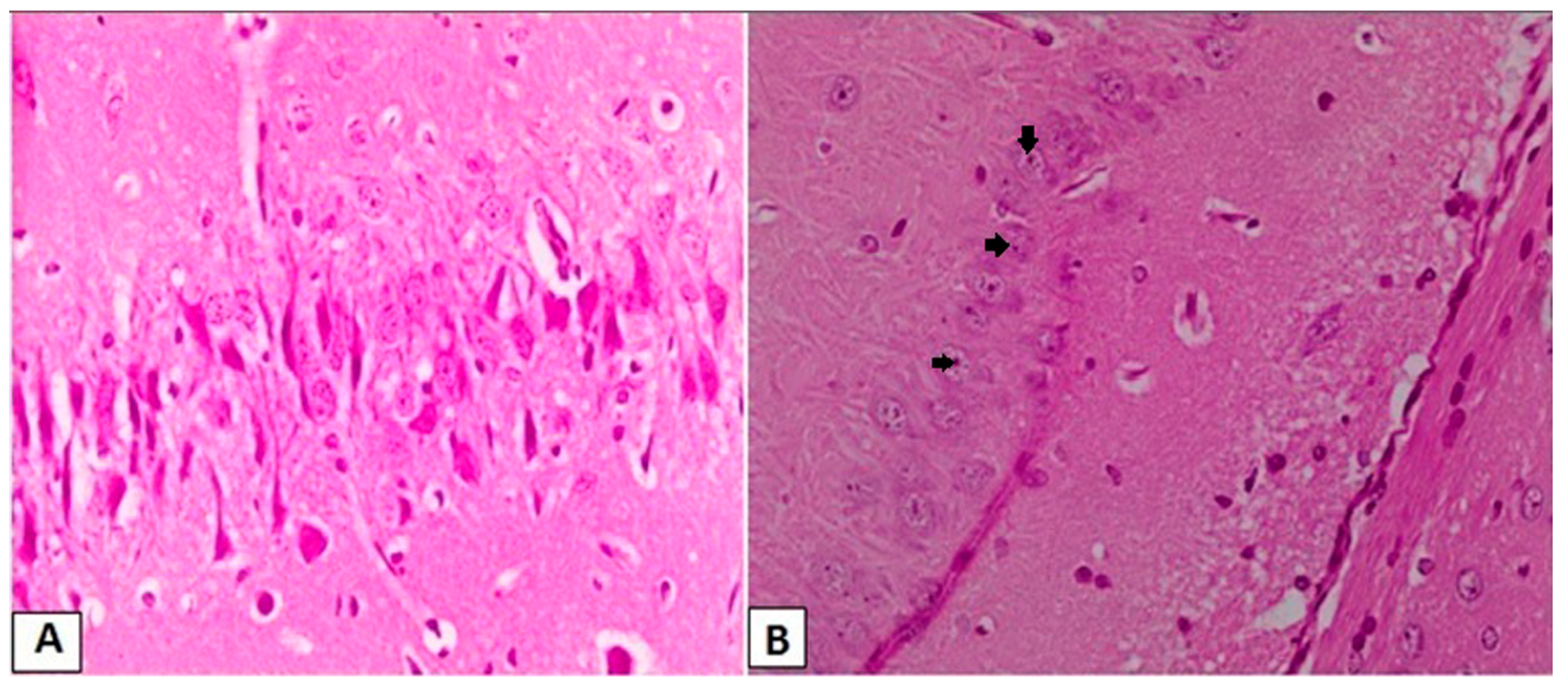
2.7. Histopathological Examination of Hippocampal Neurons by Hematoxylin and Eosin
2.8. Measurement of Expression of Heat Shock Protein (Hsp) 70 and Microtubule-Associated Protein 1A/1B-Light Chain 3 (LC3) by Immunohistochemistry in Hippocampus
2.9. Statistical Analysis
3. Results
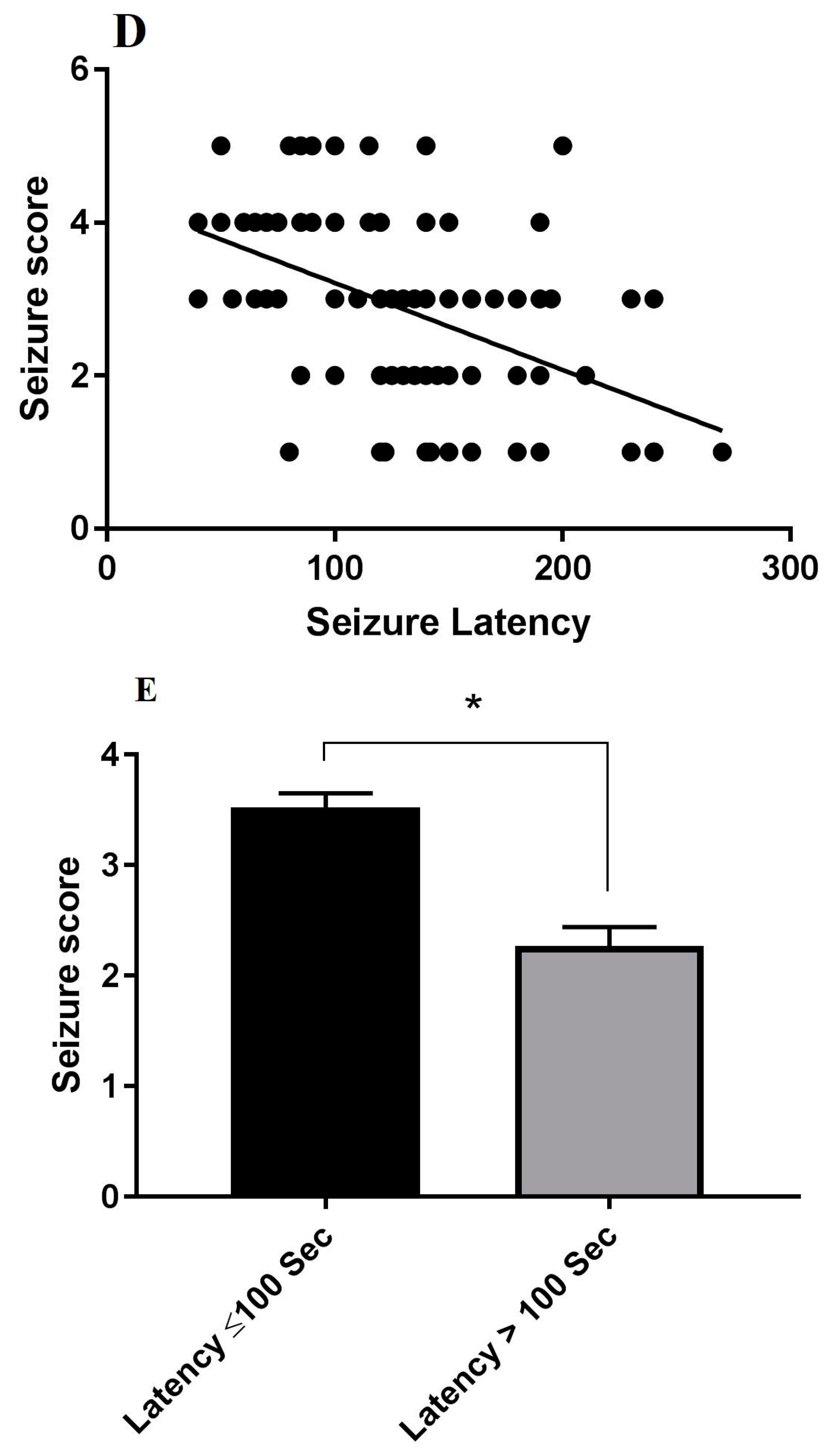
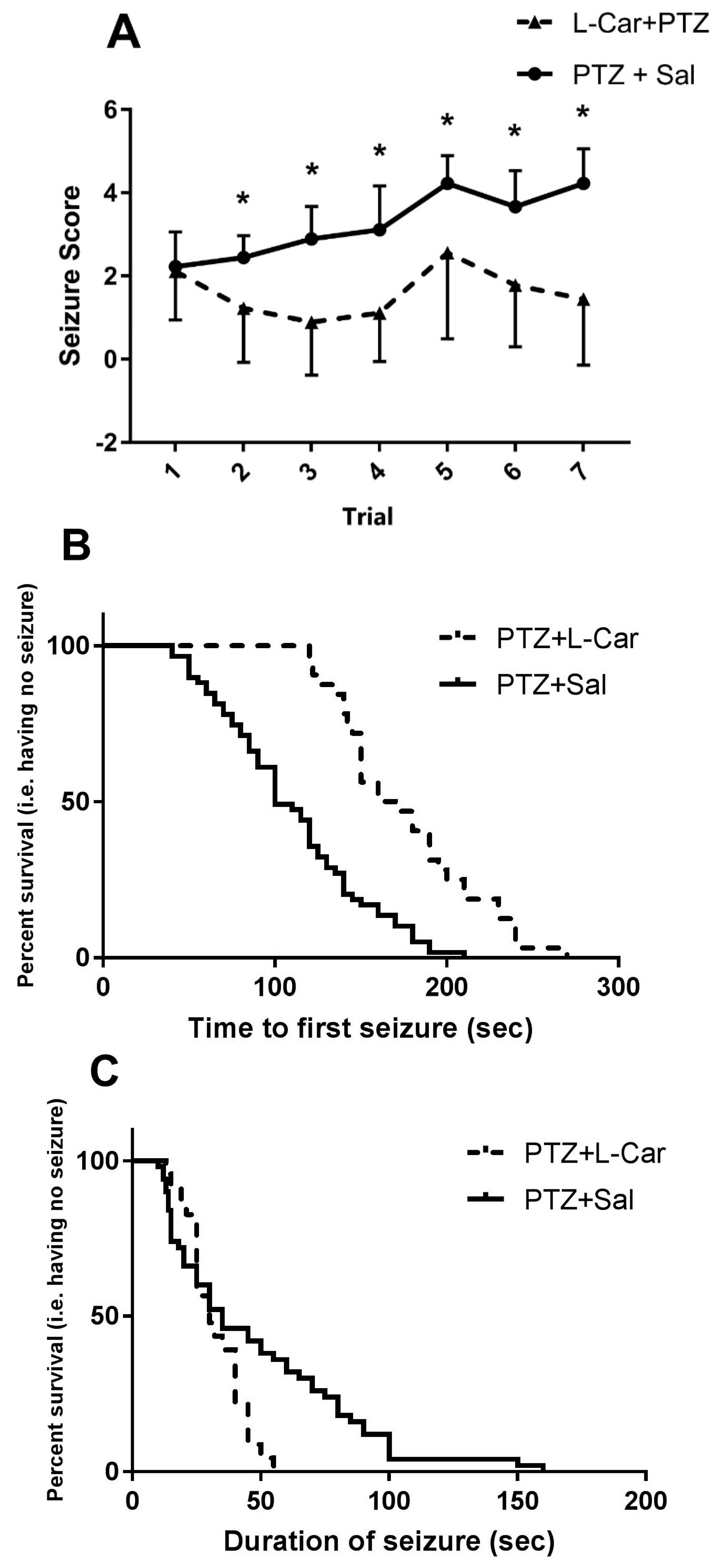
3.1. The Behavioral Effects of l-Car on PTZ-Induced Seizure
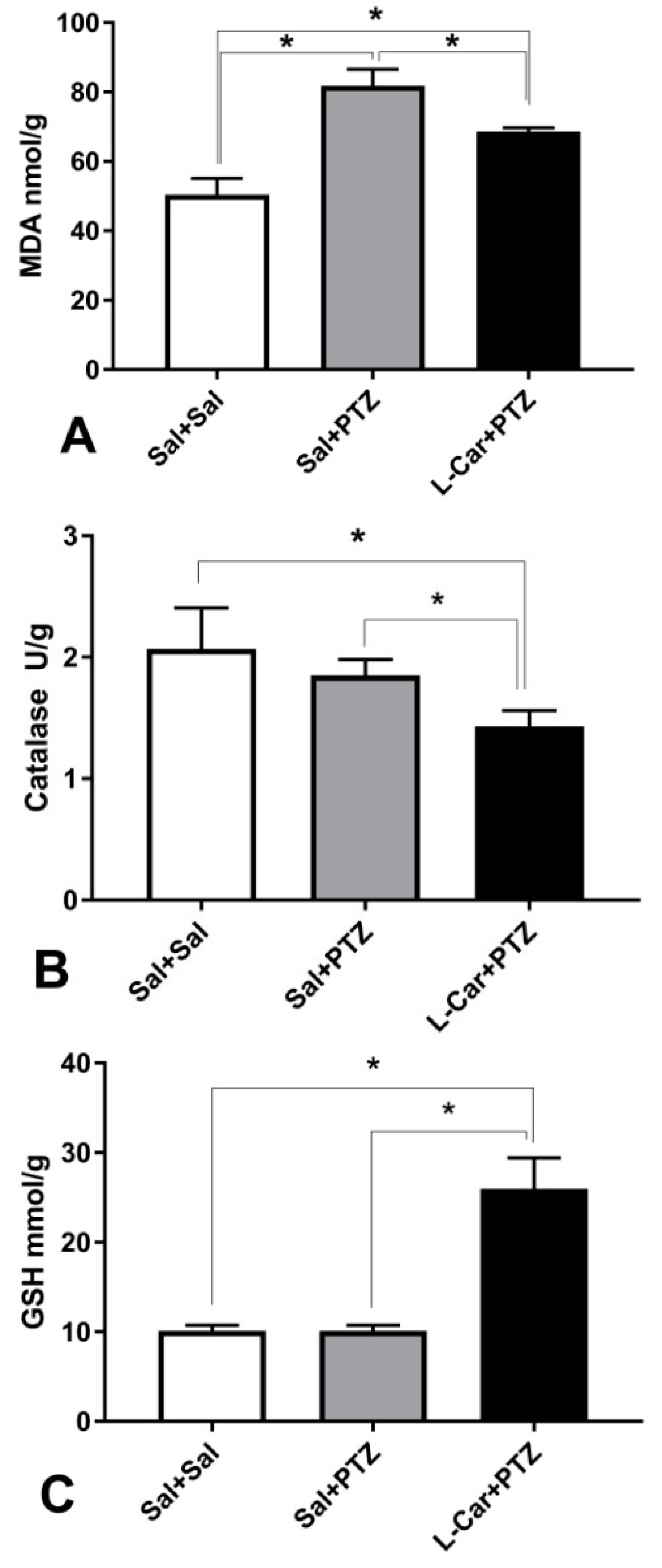
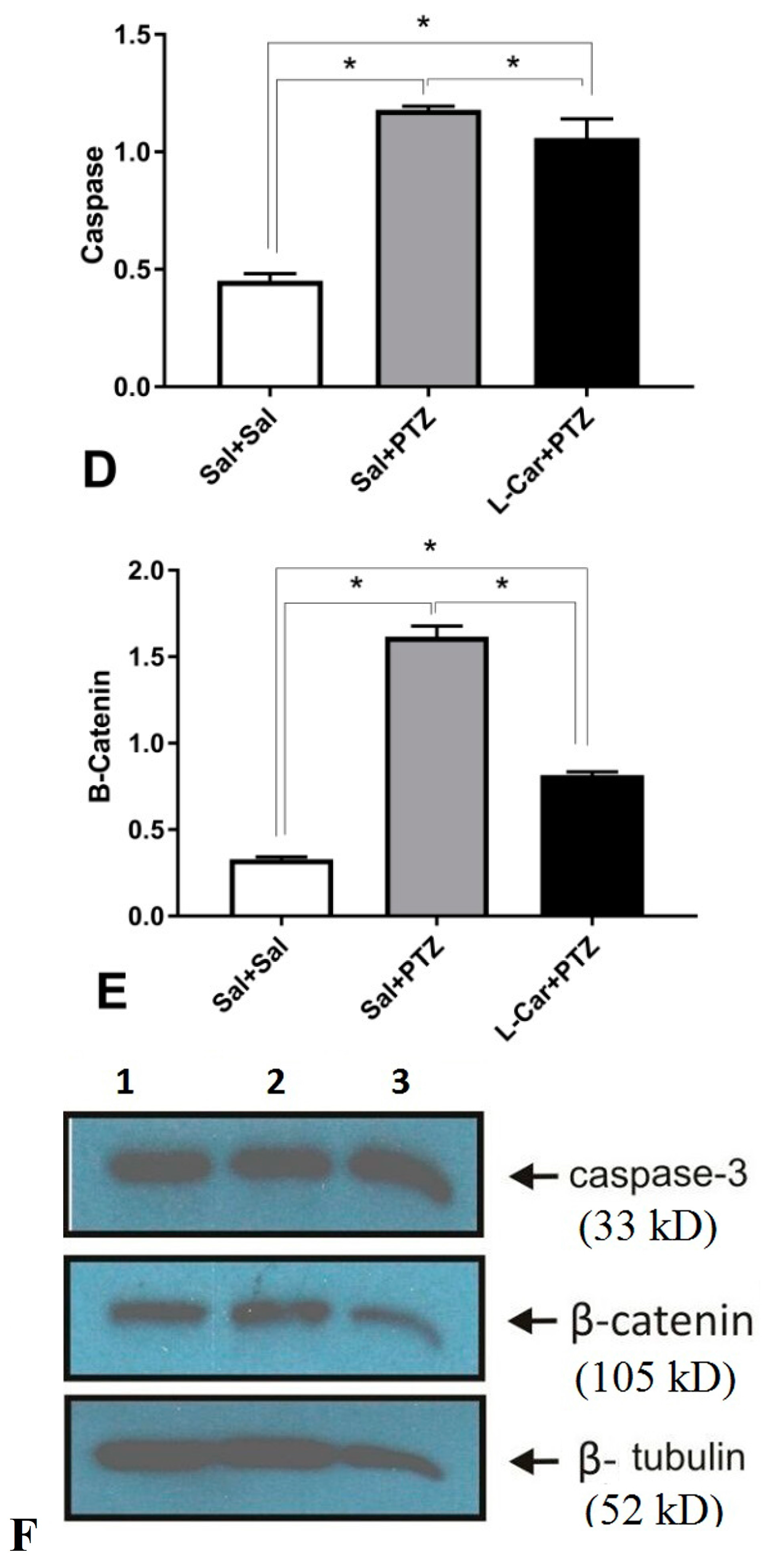
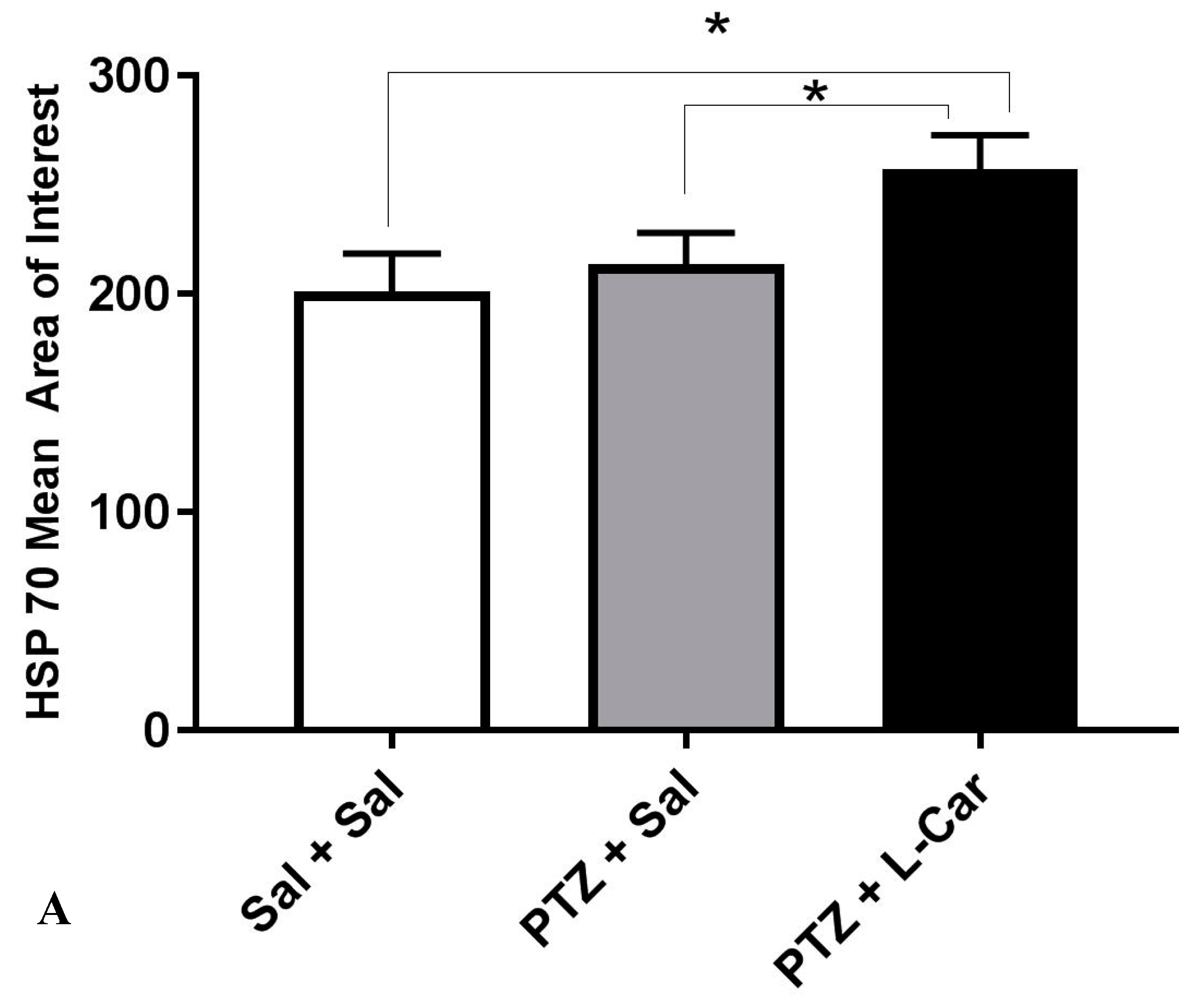
3.2. The Molecular Effects of l-Car on MDA, GSH, Catalase Enzyme, Caspase-3 and β-Catenin
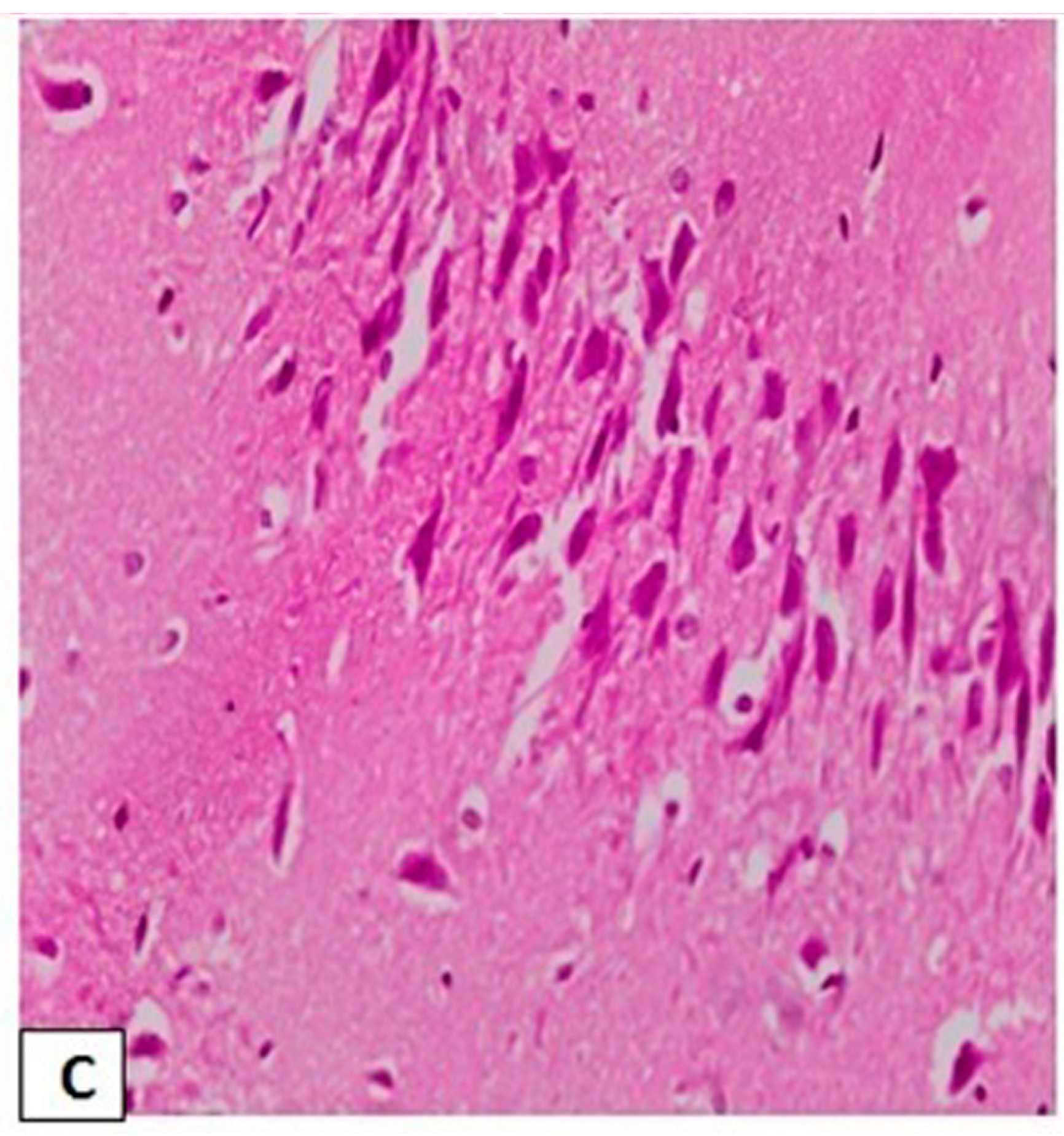
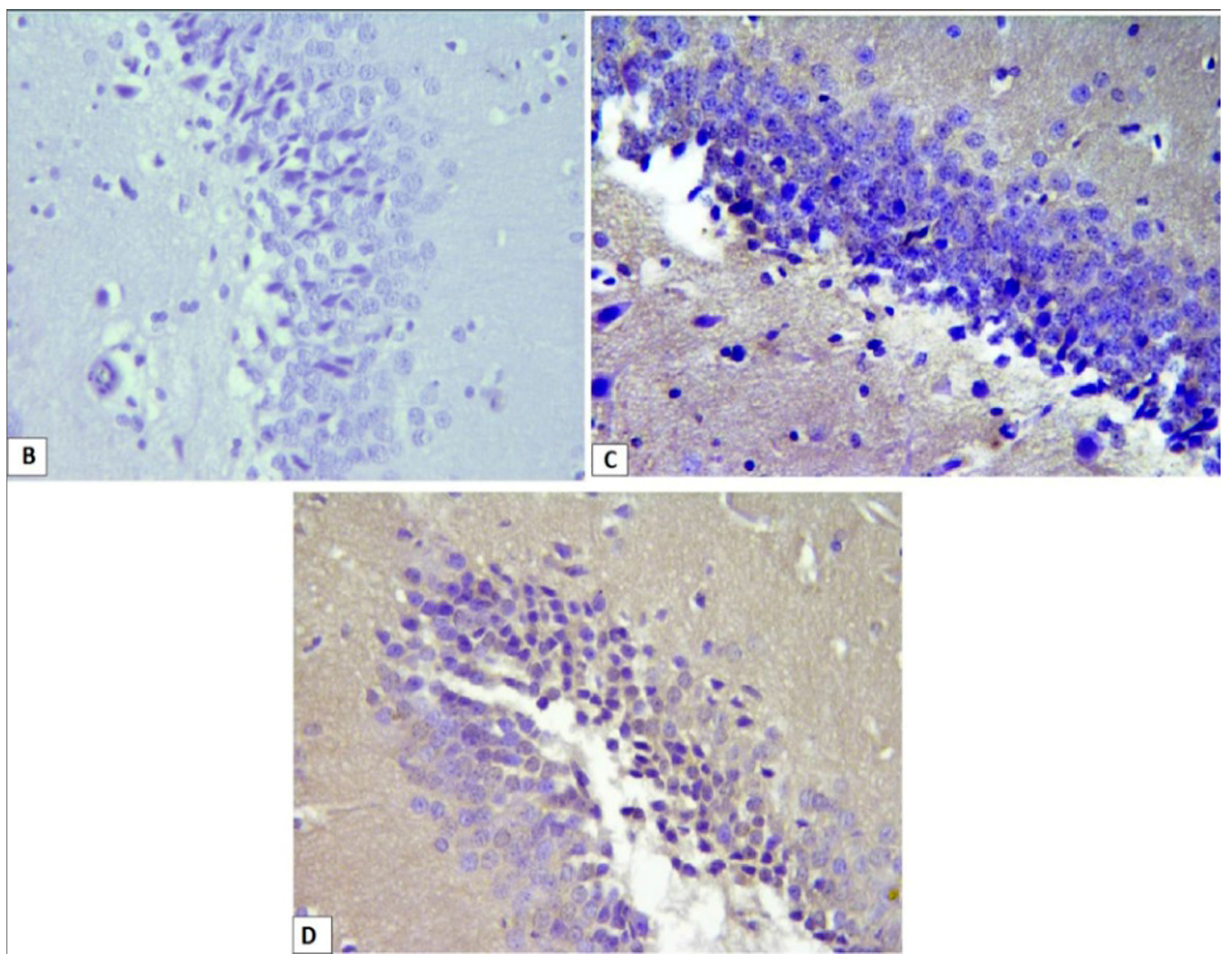
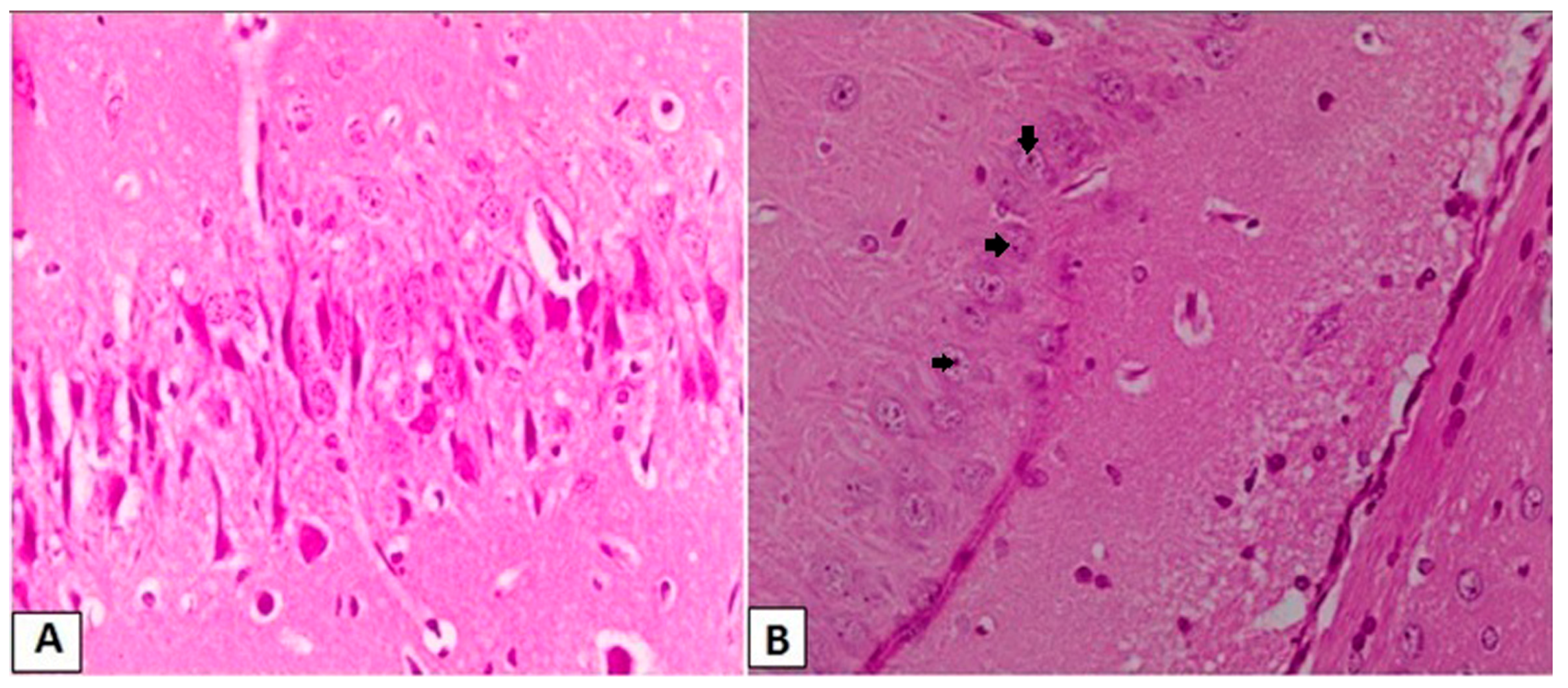
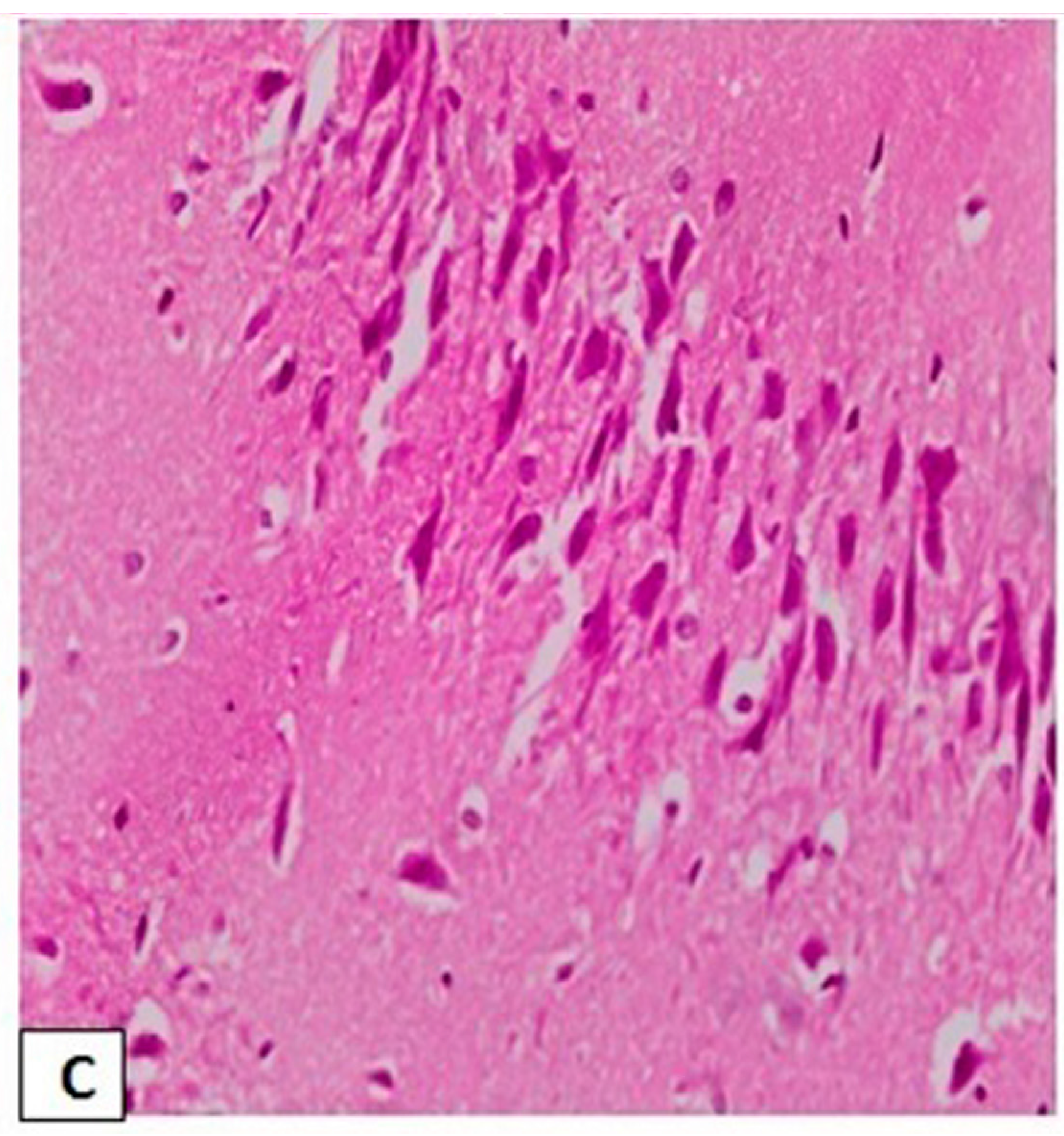
3.3. Effects of l-Car on the Morphology of Neurons in the CA3 Region of Hippocampus
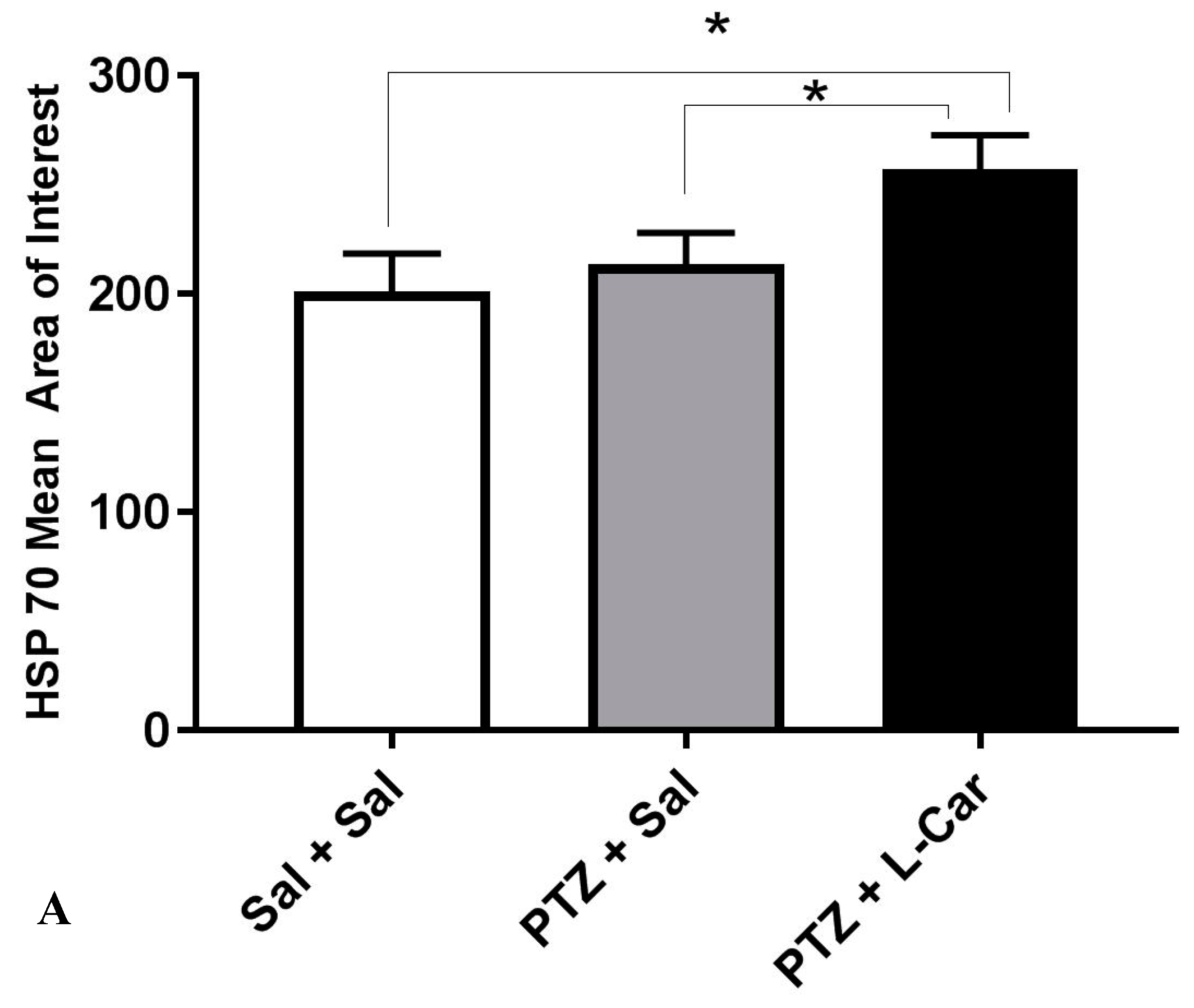
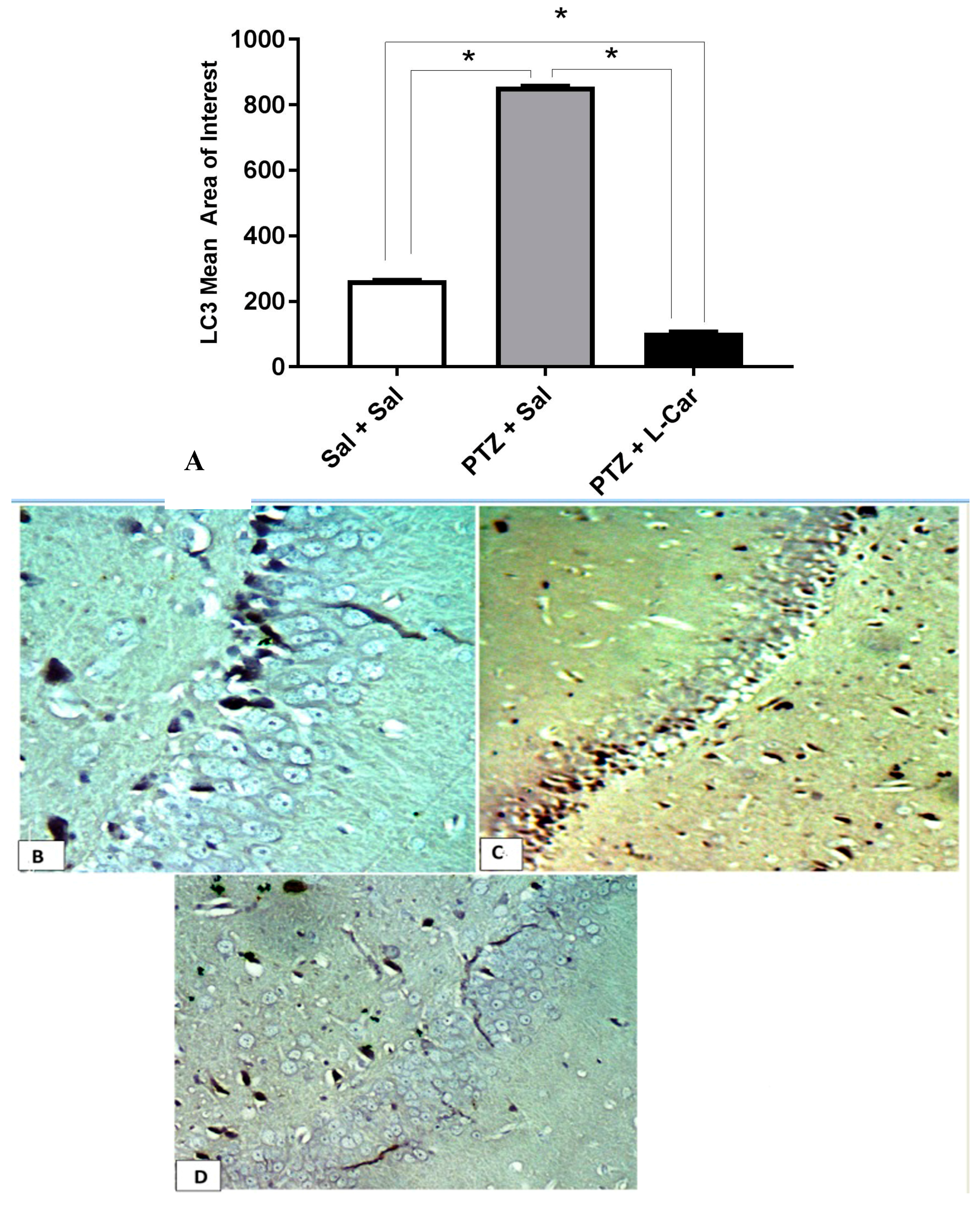
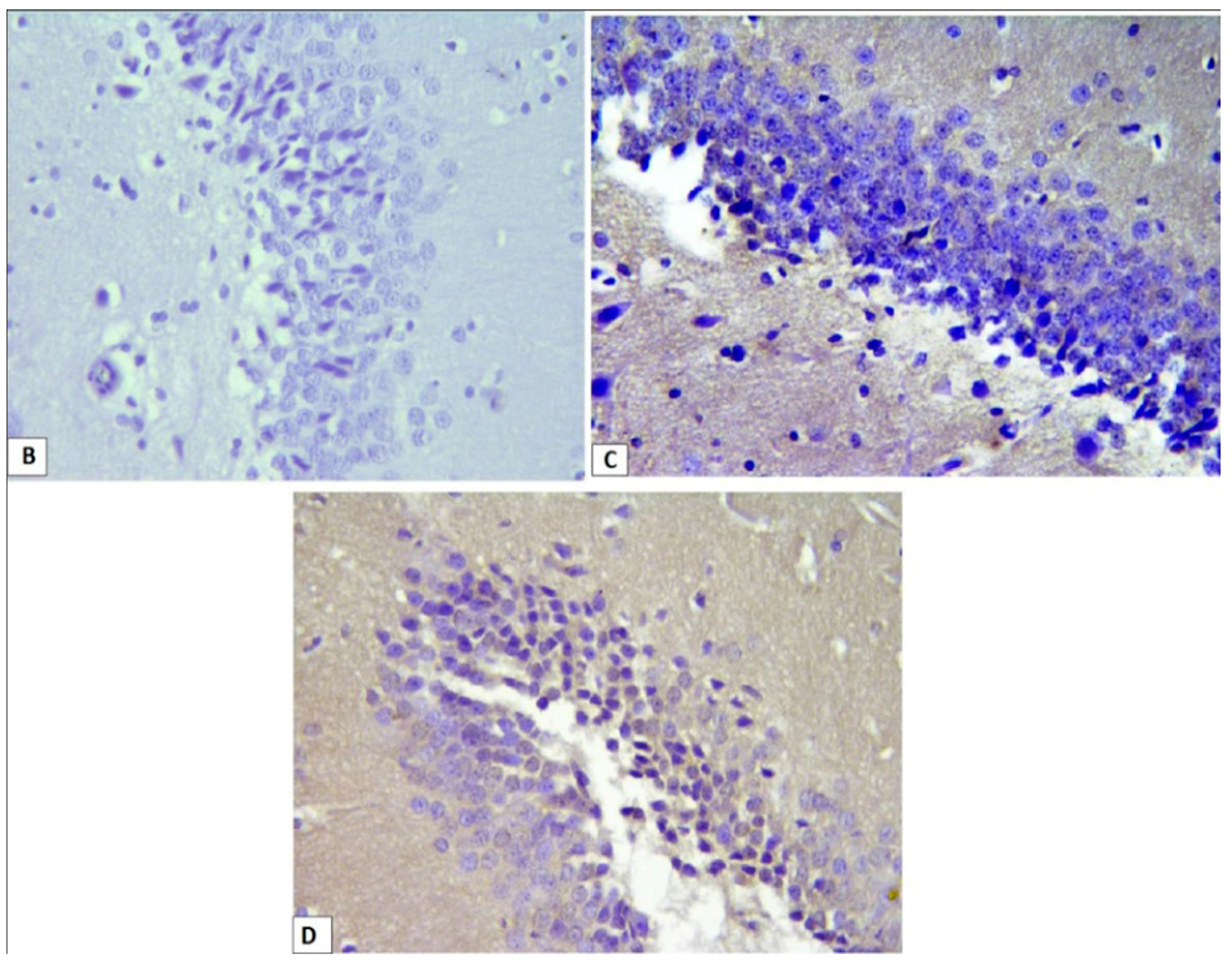
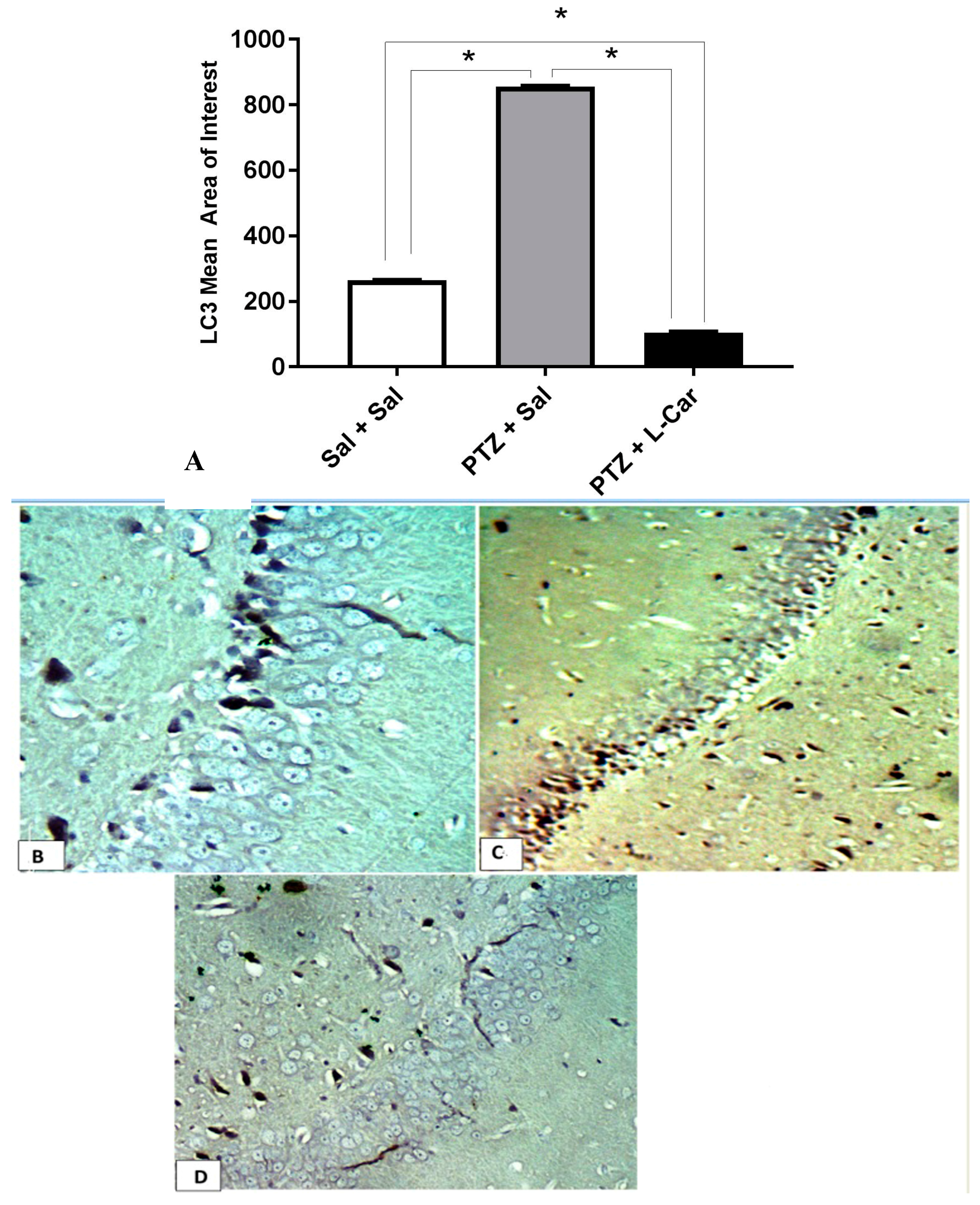
3.4. Effects of l-Car on Hsp70 and LC3 Expression in the CA3 Region of Hippocampus
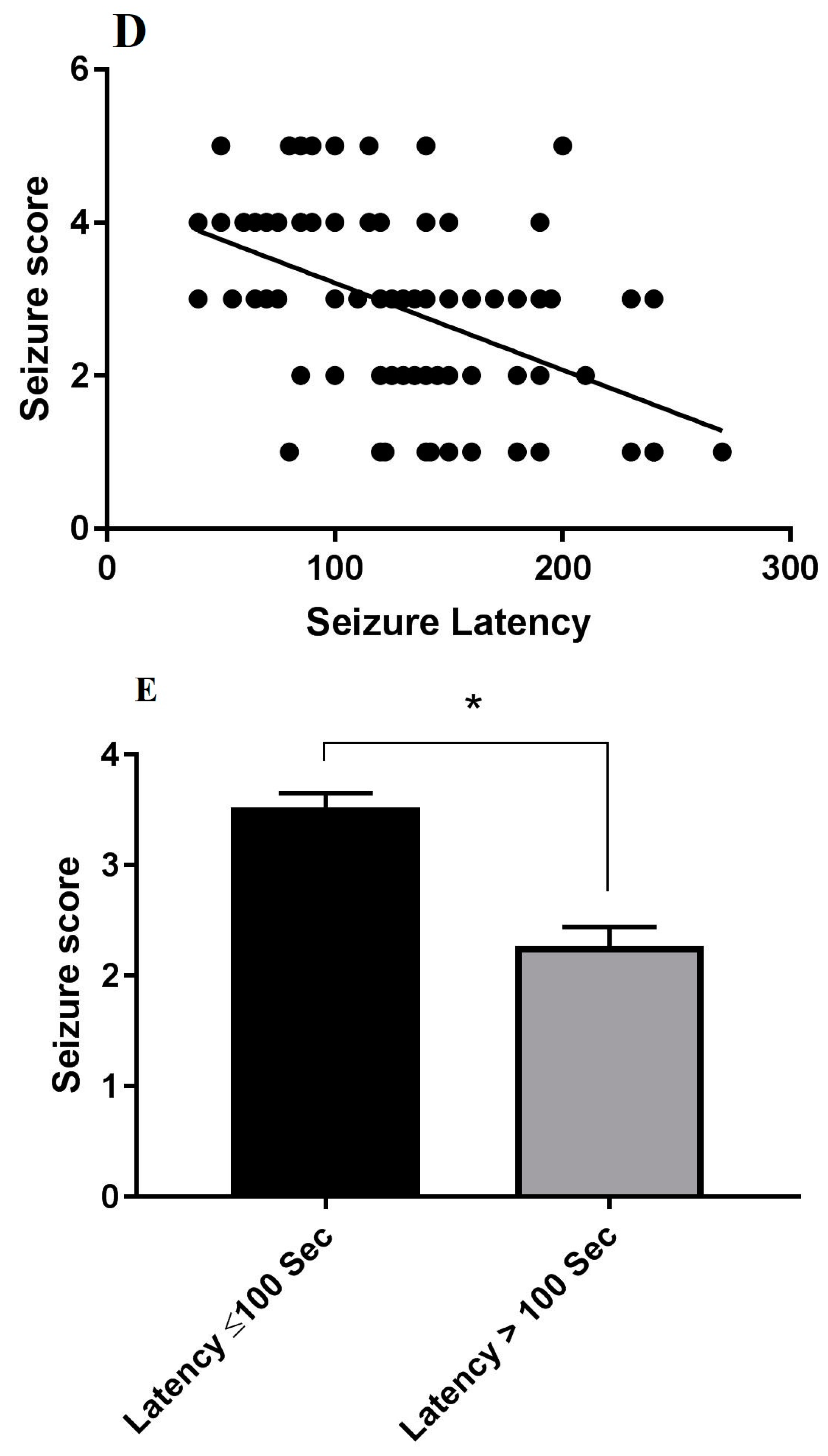
3.5. Correlations between the Stage of Seizure, Oxidative Stress Markers, Hsp70 Expression and LC3 Expression in Hippocampus
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Forsgren, L.; Beghi, E.; Oun, A.; Sillanpaa, M. The epidemiology of epilepsy in Europe—A systematic review. Eur. J. Neurol. 2005, 12, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Hauser, W.A.; Annegers, J.F.; Rocca, W.A. Descriptive epidemiology of epilepsy: Contributions of population-based studies from Rochester, Minnesota. Mayo Clin. Proc. 1996, 71, 576–586. [Google Scholar] [CrossRef] [PubMed]
- Eadie, M. Epilepsy-from the Sakikku to hughlings Jackson. J. Clin. Neurosci. 1995, 2, 156–162. [Google Scholar] [CrossRef]
- Fisher, R.S.; Boas, W.V.; Blume, W.; Elger, C.; Genton, P.; Lee, P.; Engel, J. Epileptic seizures and epilepsy: Definitions proposed by the International League Against Epilepsy (ILAE) and the International Bureau for Epilepsy (IBE). Epilepsia 2005, 46, 470–472. [Google Scholar] [CrossRef] [PubMed]
- Engel, J., Jr. Introduction to temporal lobe epilepsy. Epilepsy Res. 1996, 26, 141–150. [Google Scholar] [CrossRef]
- Pitkanen, A.; Lukasiuk, K. Mechanisms of epileptogenesis and potential treatment targets. Lancet Neurol. 2011, 10, 173–186. [Google Scholar] [CrossRef]
- Cunliffe, V.T.; Baines, R.A.; Giachello, C.N.; Lin, W.H.; Morgan, A.; Reuber, M.; Russell, C.; Walker, M.C.; Williams, R.S. Epilepsy research methods update: Understanding the causes of epileptic seizures and identifying new treatments using non-mammalian model organisms. Seizure 2015, 24, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Tang, F.; Hartz, A.M.S.; Bauer, B. Drug-Resistant Epilepsy: Multiple Hypotheses, Few Answers. Front. Neurol. 2017, 8, 301. [Google Scholar] [CrossRef] [PubMed]
- Levira, F.; Thurman, D.J.; Sander, J.W.; Hauser, W.A.; Hesdorffer, D.C.; Masanja, H.; Odermatt, P.; Logroscino, G. Premature mortality of epilepsy in low- and middle-income countries: A systematic review from the Mortality Task Force of the International League Against Epilepsy. Epilepsia 2017, 58, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Zaccara, G.; Giannasi, G.; Oggioni, R.; Rosati, E.; Tramacere, L.; Palumbo, P.; Convulsive Status Epilepticus Study Group of the Uslcentro Toscana, Italy. Challenges in the treatment of convulsive status epilepticus. Seizure 2017, 47, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Kwan, P.; Brodie, M.J. Early identification of refractory epilepsy. N. Engl. J. Med. 2000, 342, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, K.; Eaton, S. Mitochondrial beta-oxidation. Eur. J. Biochem. 2004, 271, 462–469. [Google Scholar] [CrossRef] [PubMed]
- Kido, Y.; Tamai, I.; Ohnari, A.; Sai, Y.; Kagami, T.; Nezu, J.; Nikaido, H.; Hashimoto, N.; Asano, M.; Tsuji, A. Functional relevance of carnitine transporter OCTN2 to brain distribution of L carnitine and acetyl-l-carnitine across the blood-brain barrier. J. Neurochem. 2001, 79, 959–969. [Google Scholar] [CrossRef] [PubMed]
- Smeland, O.S.; Meisingset, T.W.; Sonnewald, U. Dietary supplementation with acetyl-l-carnitine improves brain energy metabolism in healthy mice and increases noradrenaline and serotonin content. Neurochem. Int. 2012, 61, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Hansen, S.L.; Nielsen, A.H.; Knudsen, K.E.; Artmann, A.; Petersen, G.; Kristiansen, U.; Hansen, H.S. Ketogenic diet is antiepileptogenic in pentylenetetrazole kindled mice and decrease levels of N-acylethanolamines in hippocampus. Neurochem. Int. 2009, 54, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Couturier, A.; Ringseis, R.; Mooren, F.C.; Kruger, K.; Most, E.; Eder, K. Carnitine supplementation to obese Zucker rats prevents obesity-induced type II to type I muscle fiber transition and favors an oxidative phenotype of skeletal muscle. Nutr. Metab. (Lond.) 2013, 10, 48. [Google Scholar] [CrossRef] [PubMed]
- Moloney, T.C.; Dockery, P.; Windebank, A.J.; Barry, F.P.; Howard, L.; Dowd, E. Survival and immunogenicity of mesenchymal stem cells from the green fluorescent protein transgenic rat in the adult rat brain. Neurorehabil. Neural Repair 2010, 24, 645–656. [Google Scholar] [CrossRef] [PubMed]
- Hussein, A.M.; Abbas, K.M.; Abuoleseod, O. Effects of Ferulic Acid on Oxidative Stress, Heat Shock Protein 70, Connexin 43 and Monoamines in Hippocampus of Pentylenetetrazole-Kindled Rats. J. Physiol. Pharmacol. 2017, 97, 579–585. [Google Scholar] [CrossRef] [PubMed]
- Forloni, G.; Angeretti, N.; Smiroldo, S. Neuroprotective activity of acetyl-l-carnitine: Studies in vitro. J. Neurosci. Res. 1994, 37, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Aureli, T.; Miccheli, A.; Ricciolini, R.; Di Cocco, M.E.; Ramacci, M.T.; Angelucci, L.; Ghirardi, O.; Conti, F. Aging brain: Effect of acetyl-l-carnitine treatment on rat brain energy and phospholipid metabolism. A study by 31P and 1H NMR spectroscopy. Brain Res. 1990, 526, 108–112. [Google Scholar] [CrossRef]
- Aureli, T.; Di Cocco, M.E.; Puccetti, C.; Ricciolini, R.; Scalibastri, M.; Miccheli, A.; Manetti, C.; Conti, F. Acetyl-l-carnitine modulates glucose metabolism and stimulates glycogen synthesis in rat brain. Brain Res. 1998, 796, 75–81. [Google Scholar] [CrossRef]
- Scafidi, S.; Fiskum, G.; Lindauer, S.L.; Bamford, P.; Shi, D.; Hopkins, I.; McKenna, M.C. Metabolism of acetyl-l-carnitine for energy and neurotransmitter synthesis in the immature rat brain. J. Neurochem. 2010, 114, 820–831. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Killilea, D.W.; Ames, B.N. Age-associated mitochondrial oxidative decay: Improvement of carnitine acetyltransferase substrate-binding affinity and activity in brain by feeding old rats acetyl-l-carnitine and/or R-alpha-lipoic acid. Proc. Natl. Acad. Sci. USA 2002, 99, 1876–1881. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Iryo, Y.; Matsuoka, M.; Igisu, H.; Ikeda, M. Suppression of pentylenetetrazol-induced seizures by carnitine in mice. Naunyn-Schmiedeberg’s Arch. Pharmacol. 1997, 355, 545–549. [Google Scholar] [CrossRef]
- Dhir, A. Pentylenetetrazol (PTZ) Kindling Model of Epilepsy. Curr. Protoc. Neurosci. 2012, 58, 9–37. [Google Scholar] [CrossRef]
- Rajabzadeh, A.; Bideskan, A.E.; Fazelm, A.; Sankian, M.; Rafatpanah, H.; Haghir, H. The effect of PTZ-induced epileptic seizures on hippocampal expression of PSA-NCAM in offspring born to kindled rats. J. Biomed. Sci. 2012, 19, 56. [Google Scholar] [CrossRef] [PubMed]
- Saha, L.; Bhandari, S.; Bhatia, A.; Banerjee, D.; Chakrabarti, A. Anti-kindling Effect of Bezafibrate, a Peroxisome Proliferator-activated Receptors Alpha Agonist, in Pentylenetetrazole Induced Kindling Seizure Model. J. Epilepsy Res. 2014, 4, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Sudha, K.; Rao, A.V.; Rao, A. Oxidative stress and antioxidants in epilepsy. Clin. Chim. Acta. 2001, 303, 19–24. [Google Scholar] [CrossRef]
- Danjo, S.; Ishihara, Y.; Watanabe, M.; Nakamura, Y.; Itoh, K. Pentylenetrazole-induced loss of blood-brain barrier integrity involves excess nitric oxide generation by neuronal nitric oxide synthase. Brain Res. 2013, 1530, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Shehata, A.M. Neurophysiological Studies on the Effect of Acetone on Pentylenetetrazole-Induced Seizure in Rats. Bull. Egypt. Soc. Physiol. Sci. 2011, 31, 135–146. [Google Scholar]
- Ahmed, M.F.; Mahmoud, M.A. Effect of l-Carnitine on Pilocarpine-Induced Seizures in Rats. J. Am. Sci. 2012, 8, 612–618. [Google Scholar]
- Zhao, X.; Liu, G.; Shen, H.; Gao, B.; Li, X.; Fu, J.; Zhou, J.; Ji, Q. Liraglutide inhibits autophagy and apoptosis induced by high glucose through GLP-1R in renal tubular epithelial cells. Int. J. Mol. Med. 2015, 35, 684–692. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Han, R.; Liang, Z.Q.; Wu, J.C.; Zhang, X.D.; Gu, Z.L.; Qin, Z.H. An autophagic mechanism is involved in apoptotic death of rat striatal neurons induced by the non-N-methyl-Daspartate receptor agonist kainic acid. Autophagy 2008, 4, 214–226. [Google Scholar] [CrossRef] [PubMed]
- Shacka, J.J.; Lu, J.; Xie, Z.L.; Uchiyama, Y.; Roth, K.A.; Zhang, J. Kainic acid induces early and transient autophagic stress in mouse hippocampus. Neurosci. Lett. 2007, 414, 57–60. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Xu, J.; Lin, Y.; Zhao, X.; Liu, X.; Chi, Z. Autophagy is upregulated in rats with status epilepticus and partly inhibited by vitamin E. Biochem. Biophys. Res. Commun. 2009, 379, 949–953. [Google Scholar] [CrossRef] [PubMed]
- Scherz-Shouval, R.; Elazar, Z. ROS, mitochondria and the regulation of autophagy. Trends Cell Biol. 2007, 17, 422–427. [Google Scholar] [CrossRef] [PubMed]
- Scherz-Shouval, R.; Shvets, E.; Fass, E.; Shorer, H.; Gil, L.; Elazar, Z. Reactive oxygen species are essential for autophagy and specifically regulate the activity of Atg4. EMBO J. 2007, 26, 1749–1760. [Google Scholar] [CrossRef] [PubMed]
- Simonian, N.A.; Getz, R.L.; Leveque, J.C.; Konradi, C.; Coyle, J.T. Kainic acid induces apoptosis in neurons. Neuroscience 1996, 75, 1047–1055. [Google Scholar] [CrossRef]
- Naseer, M.I.; Ullah, I.; Ullah, N.; Lee, H.Y.; Cheon, E.W.; Chung, J.; Kim, M.O. Neuroprotective effect of vitamin C against PTZ induced apoptotic neurodegeneration in adult rat brain. Pak. J. Pharm. Sci. 2011, 24, 263–268. [Google Scholar] [PubMed]
- Green, D.R.; Reed, J.C. Mitochondria and apoptosis. Science 1998, 281, 1309–1312. [Google Scholar] [CrossRef] [PubMed]
- Soti, C.; Nagy, E.; Giricz, Z.; Vígh, L.; Csermely, P.; Ferdinandy, P. Heat shock proteins as emerging therapeutic targets. Br. J. Pharmacol. 2005, 146, 769–780. [Google Scholar] [CrossRef] [PubMed]
- Foster, J.A.; Brown, I.R. Differential induction of heat shock mRNA in oligodendrocytes, microglia, and astrocytes following hyperthermia. Brain Res. Mol. Brain Res. 1997, 45, 207–218. [Google Scholar] [CrossRef]
- Gass, P.; Prior, P.; Kiessling, M. Correlation between seizure intensity and stress protein expression after limbic epilepsy in the rat brain. Neuroscience 1995, 65, 27–36. [Google Scholar] [CrossRef]
- Zhang, X.; Boulton, A.A.; Yu, P.H. Expression of heat shock protein-70 and limbic seizure-induced neuronal death in the rat brain. Eur. J. Neurosci. 1996, 8, 1432–1440. [Google Scholar] [CrossRef] [PubMed]
- Thom, M.; Seetah, S.; Sisodiya, S.; Koepp, M.; Scaravilli, F. Sudden and unexpected death in epilepsy (SUDEP): Evidence of acute neuronal injury using HSP-70 and c-Jun immunohistochemistry. Neuropathol. Appl. Neurobiol. 2003, 29, 132–143. [Google Scholar] [CrossRef]
- Dericioglu, N.; Soylemezoglu, F.; Gursoy-Ozdemir, Y.; Akalan, N.; Saygi, S.; Dalkara, T. Cell death and survival mechanisms are concomitantly active in the hippocampus of patients with mesial temporal sclerosis. Neuroscience 2013, 237, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Li, W.X.; Chen, C.H.; Ling, C.C.; Li, G.C. Apoptosis in heat induced cell killing: The protective role of hsp-70 and the sensitization eVect of the c-myc gene. Radiat. Res. 1996, 145, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.G.; Ma, Q.Z.; Xu, C.X. Abrogation of heat-shock protein (HSP) 70 expression induced cell growth inhibition and apoptosis in human androgen-independent prostate cancer cell line PC-3m. Asian J. Androl. 2004, 6, 319–324. [Google Scholar] [PubMed]
- Lee, C.S.; Montebello, J.; Rush, M.; Georgiou, T.; Wawryk, S.; Rode, J. Overexpression of heat shock protein (HSP) 70 associated with abnormal p53 expression in cancer of the pancreas. Zentralbl. Pathol. 1994, 140, 259–264. [Google Scholar] [PubMed]
- Merrick, B.A.; He, C.; Witcher, L.L.; Patterson, R.M.; Reid, J.J.; Pence-Pawlowski, P.M.; Selkirk, J.K. HSP binding and mitochondrial localization of p53 protein in human HT1080 and mouse C3H10T1/2 cell lines. Biochim. Biophys. Acta 1996, 1297, 57–68. [Google Scholar] [CrossRef]
- Kanitkara, M.; Bhonde, R.R. Curcumin treatment enhances islet recovery by induction of heat shock response proteins, Hsp70 and heme oxygenase-1, during cryopreservation. Life Sci. 2008, 82, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Ayala, G.X.; Tapia, R. HSP70 expression protects against hippocampal neurodegeneration induced by endogenous glutamate in vivo. Neuropharmacology 2008, 55, 1383–1390. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.M.; Huang, C.C.; Li, L.H.; Tsai, J.J.; Hsu, K.S. The Chinese herbal medicine Chai-Hu-Long-Ku-Mu-Li-Tan (TW-001) exerts anticonvulsant effects against different experimental models of seizure in rats. Jpn. J. Pharmacol. 2000, 82, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Lie, D.C.; Colamarino, S.A.; Song, H.J.; Desire, L.; Mira, H.; Consiglio, A.; Lein, E.S.; Jessberger, S.; Lansford, H.; Dearie, A.R.; et al. Wnt signalling regulates adult hippocampal neurogenesis. Nature 2005, 437, 1370–1375. [Google Scholar] [CrossRef] [PubMed]
- Wisniewska, M.B. Physiological role of beta-catenin/TCF signaling in neurons of the adult brain. Neurochem. Res. 2013, 38, 1144–1155. [Google Scholar] [CrossRef] [PubMed]
- De Ferrari, G.V.; Papassotiropoulos, A.; Biechele, T.; Wavrant De-Vrieze, F.; Avila, M.E.; Major, M.B.; Myers, A.; Sáez, K.; Henríquez, J.P.; Zhao, A.; et al. Common genetic variation within the low-density lipoprotein receptor-related protein 6 and late-onset Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2007, 104, 9434–9439. [Google Scholar] [CrossRef] [PubMed]
- Lovestone, S.; Guntert, A.; Hye, A.; Lynham, S.; Thambisetty, M.; Ward, M. Proteomics of Alzheimer’s disease: Understanding mechanisms and seeking biomarkers. Expert Rev. Proteom. 2007, 4, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yuan, X.; Wang, Z.; Li, R. The canonical Wnt signaling pathway in autism. CNS Neurol. Disord. Drug Targets 2014, 13, 765–770. [Google Scholar] [CrossRef] [PubMed]
- Fasen, K.; Beck, H.; Elger, C.E.; Lie, A.A. Differential regulation of cadherins and catenins during axonal reorganization in the adult rat CNS. J. Neuropathol. Exp. Neurol. 2002, 61, 903–913. [Google Scholar] [CrossRef] [PubMed]
- Madsen, T.M.; Newton, S.S.; Eaton, M.E.; Russell, D.S.; Duman, R.S. Chronic electroconvulsive seizure up-regulates betacatenin expression in rat hippocampus: Role in adult neurogenesis. Biol. Psychiatry 2003, 54, 1006–1014. [Google Scholar] [CrossRef]
- Theilhaber, J.; Rakhade, S.N.; Sudhalter, J.; Kothari, N.; Klein, P.; Pollard, J.; Jensen, F.E. Gene expression profiling of a hypoxic seizure model of epilepsy suggests a role for mTOR and Wnt signaling in epileptogenesis. PLoS ONE 2013, 8, e74428. [Google Scholar] [CrossRef] [PubMed]
- Xing, X.L.; Sha, L.Z.; Zhang, D.; Shen, Y.; Wu, L.W.; Xu, Q. Role of beta-catenin in the pathogenesis of mesial temporal lobe epilepsy. Zhongguo Yi Xue Ke Xue Yuan Xue Bao 2011, 33, 659–662. [Google Scholar] [PubMed]
- Busceti, C.L.; Biagioni, F.; Aronica, E.; Riozzi, B.; Storto, M.; Battaglia, G.; Giorgi, F.S.; Gradini, R.; Fornai, F.; Caricasole, A.; et al. Induction of the Wnt inhibitor, Dickkopf-1, is associated with neurodegeneration related to temporal lobe epilepsy. Epilepsia 2007, 48, 694–705. [Google Scholar] [CrossRef] [PubMed]
- Goodenough, S.; Schleusner, D.; Pietrzik, C.; Skutella, T.; Behl, C. Glycogen synthase kinase 3beta links neuroprotection by 17 beta-estradiol to key Alzheimer processes. Neuroscience 2005, 132, 581–589. [Google Scholar] [CrossRef] [PubMed]
- Campos, V.E.; Du, M.; Li, Y. Increased seizure susceptibility and cortical malformation in beta-catenin mutant mice. Biochem. Biophys. Res. Commun. 2004, 320, 606–614. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Stage of Seizure | MDA | GSH | CAT | Caspase-3 | β-Catenin | Hsp70 | LC3 | |
|---|---|---|---|---|---|---|---|---|---|
| Seizure stage | r | 0.78 | −0.74 | −0.34 | 0.65 | 0.63 | −0.61 | 0.67 | |
| p | 0.001 | 0.02 | 0.092 | 0.005 | 0.03 | 0.04 | 0.011 | ||
| MDA | r | −0.811 | −0.44 | 0.88 | 0.67 | −0.74 | 0.75 | ||
| p | 0.01 | 0.067 | 0.005 | 0.01 | 0.01 | 0.01 | |||
| GSH | r | 0.32 | −0.82 | 0.85 | 0.67 | −0.71 | |||
| p | 0.21 | 0.005 | 0.001 | 0.004 | 0.003 | ||||
| CAT | r | −0.13 | −0.32 | 0.64 | −0.67 | ||||
| p | 0.62 | 0.21 | 0.01 | 0.009 | |||||
| Caspase-3 | r | 0.68 | −0.62 | 0.68 | |||||
| p | 0.01 | 0.04 | 0.03 | ||||||
| β-catenin | r | 0.63 | 0.62 | ||||||
| p | 0.01 | 0.01 | |||||||
| Hsp70 | r | 0.62 | |||||||
| p | 0.011 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hussein, A.M.; Adel, M.; El-Mesery, M.; Abbas, K.M.; Ali, A.N.; Abulseoud, O.A. l-Carnitine Modulates Epileptic Seizures in Pentylenetetrazole-Kindled Rats via Suppression of Apoptosis and Autophagy and Upregulation of Hsp70. Brain Sci. 2018, 8, 45. https://doi.org/10.3390/brainsci8030045
Hussein AM, Adel M, El-Mesery M, Abbas KM, Ali AN, Abulseoud OA. l-Carnitine Modulates Epileptic Seizures in Pentylenetetrazole-Kindled Rats via Suppression of Apoptosis and Autophagy and Upregulation of Hsp70. Brain Sciences. 2018; 8(3):45. https://doi.org/10.3390/brainsci8030045
Chicago/Turabian StyleHussein, Abdelaziz M., Mohamed Adel, Mohamed El-Mesery, Khaled M. Abbas, Amr N. Ali, and Osama A. Abulseoud. 2018. "l-Carnitine Modulates Epileptic Seizures in Pentylenetetrazole-Kindled Rats via Suppression of Apoptosis and Autophagy and Upregulation of Hsp70" Brain Sciences 8, no. 3: 45. https://doi.org/10.3390/brainsci8030045