Neuroprotective Role of N-acetylcysteine against Learning Deficits and Altered Brain Neurotransmitters in Rat Pups Subjected to Prenatal Stress


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Animal Groups
- Group 1.
- (Control (CON)) Pups of dams who received normal saline intra-peritoneally in a dose of 10 mL/kg body weight during the entire course of pregnancy.
- Group 2.
- (NAC) Pups of dams who received NAC alone during the entire course of pregnancy.
- Group 3.
- (EGS) Pups of dams subjected to restrain stress from day 1 to 10 of pregnancy.
- Group 4.
- (LGS) Pups of dams subjected to restrain stress from day 11 of pregnancy until delivery.
- Group 5.
- (EGS + NAC) Pups of dams subjected to restrain stress from day 1 to 10 of pregnancy along with NAC treatment throughout pregnancy.
- Group 6.
- (LGS + NAC) Pups of dams subjected to restrain stress from day 11 of pregnancy until delivery along with the NAC treatment throughout pregnancy.
2.3. Stressing Procedure
2.4. Chemicals
2.5. Neonatal Study Parameters
2.6. Learning and Memory Test: Condition Avoidance Test (Shuttle Box Test)
2.7. Sacrifice and Tissue Processing for Estimating Whole Brain Neurotransmitter Levels
2.8. Statistical Analysis
3. Results
3.1. Effect of Prenatal Stress and NAC on Neonatal Parameters
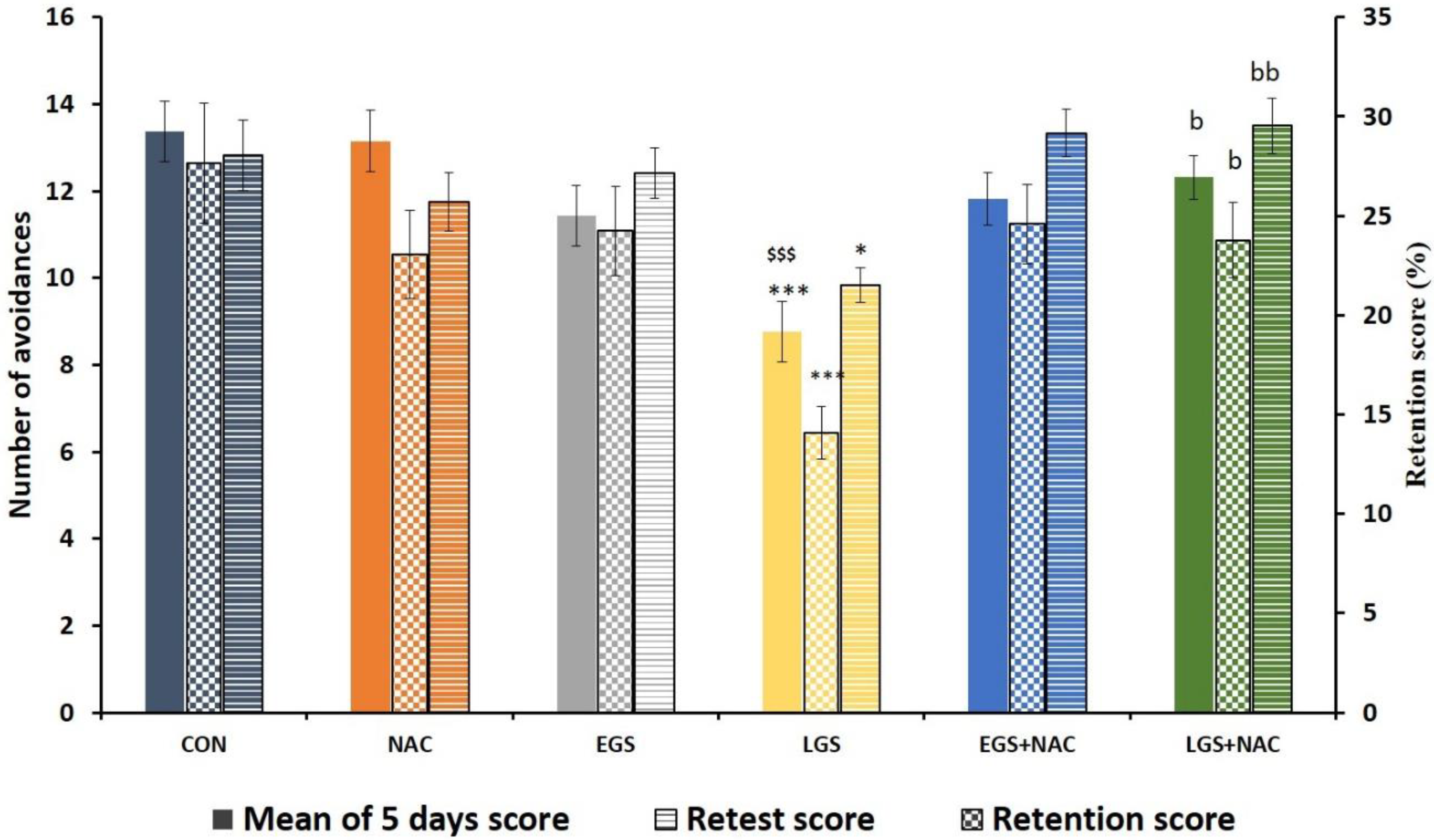
3.2. Effect of Prenatal Stress and NAC on Learning and Memory (PND 28)
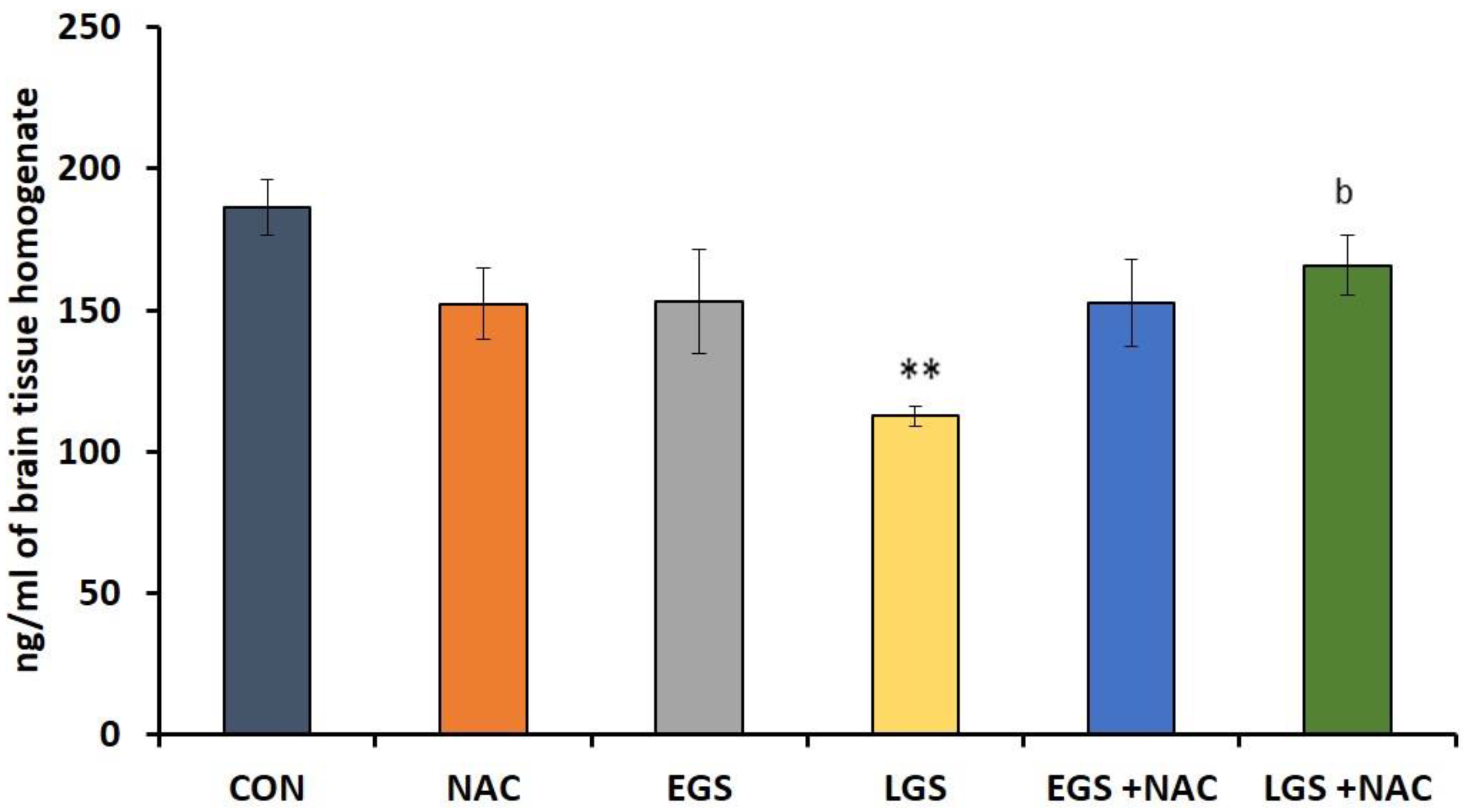
3.3. Effect of Prenatal Stress and NAC on Whole Brain Dopamine Levels
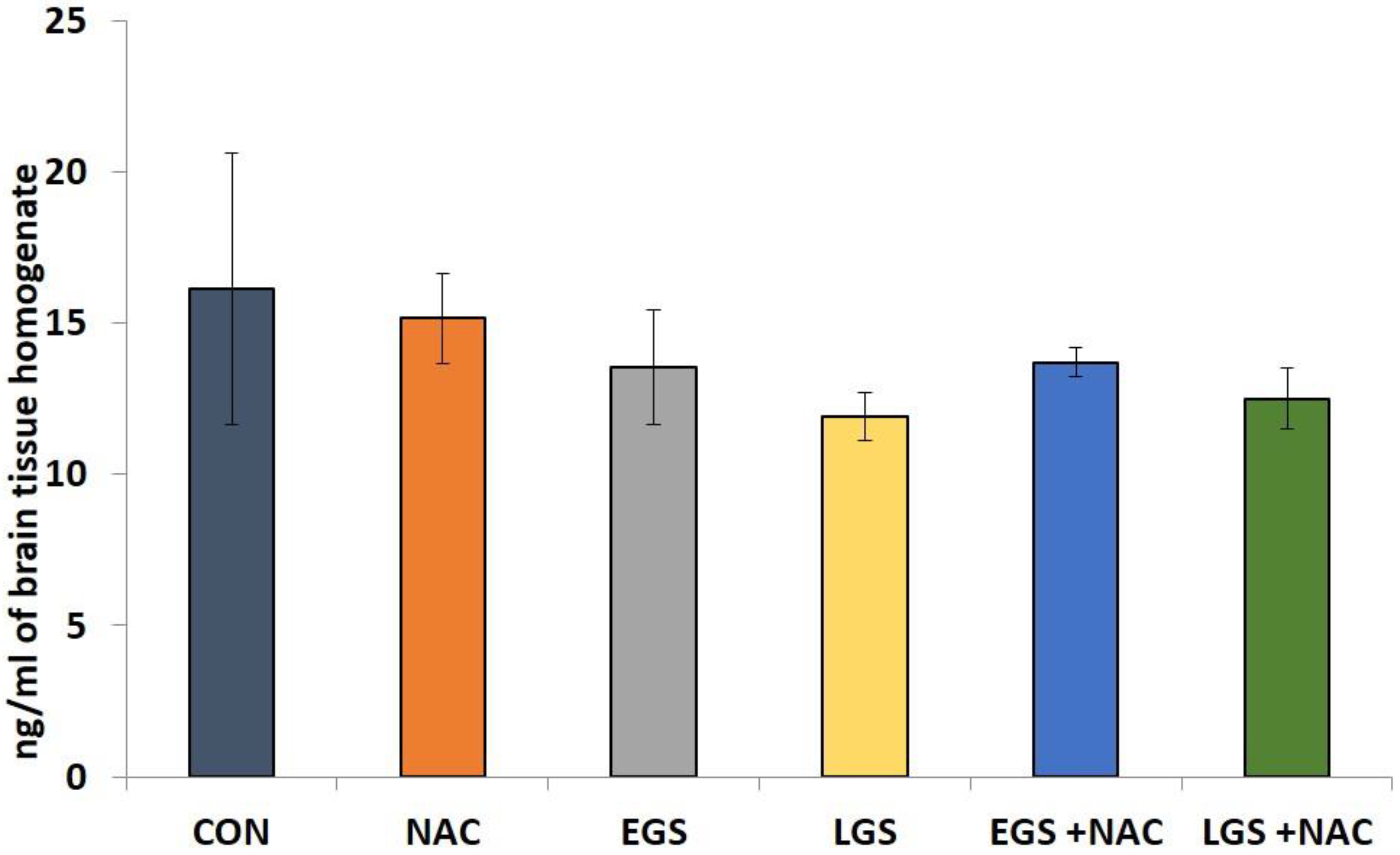
3.4. Effect of Prenatal Stress and NAC on Brain Norepinephrine Levels
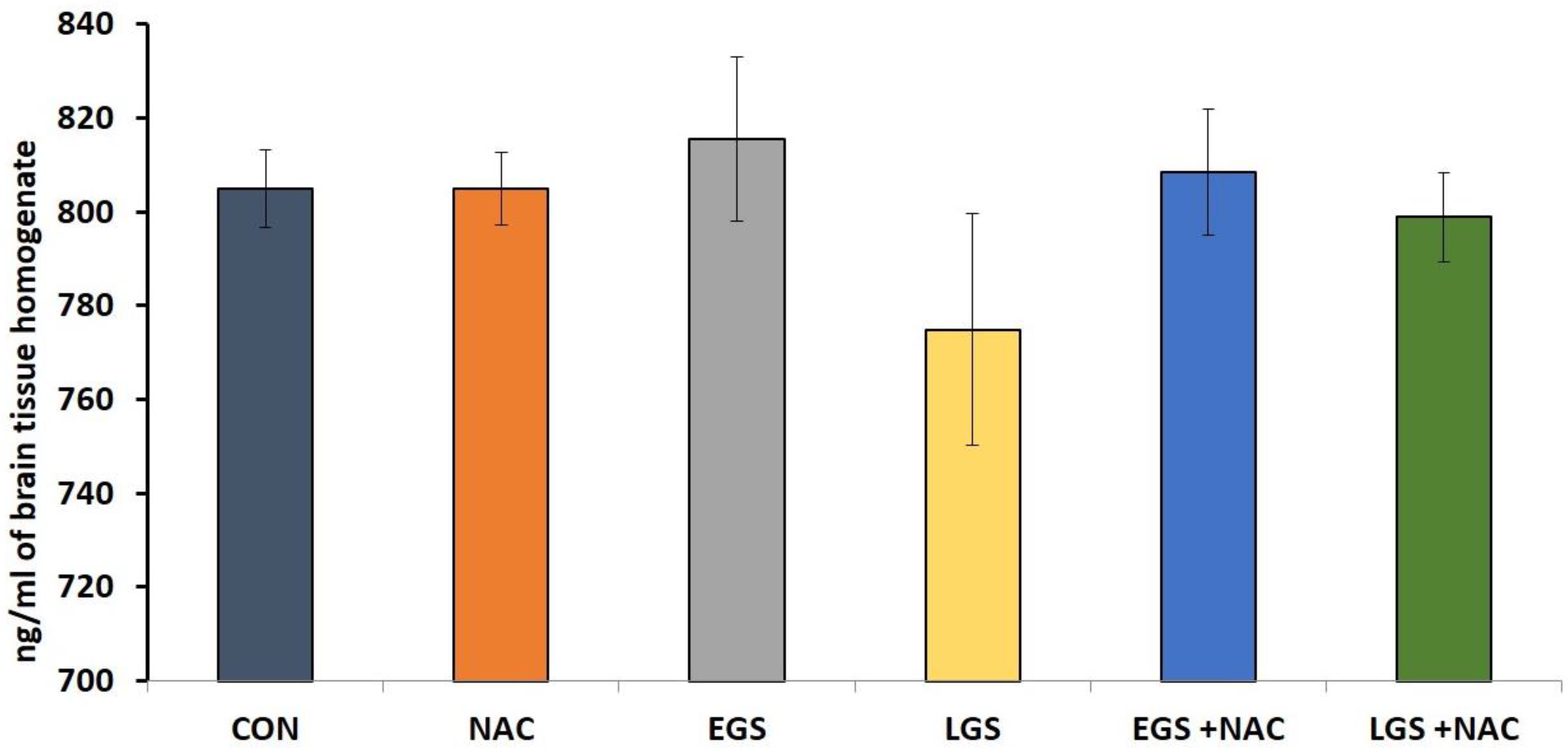
3.5. Effect of Prenatal Stress and NAC on Brain Serotonin (5HT) Levels
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Weinstock, M.; Matlina, E.; Maor, G.I.; Rosen, H.; McEwen, B.S. Prenatal stress selectively alters the reactivity of the hypothalamic-pituitary adrenal system in the female rat. Brain Res. 1992, 595, 195–200. [Google Scholar] [CrossRef]
- Weinstock, M. Alterations induced by gestational stress in brain morphology and behaviour of the offspring. Prog. Neurobiol. 2001, 65, 427–451. [Google Scholar] [CrossRef]
- Darnaudery, M.; Maccari, S. Epigenetic programming of the stress response in male and female rats by prenatal restraint stress. Brain Res. Rev. 2008, 57, 571–585. [Google Scholar] [CrossRef] [PubMed]
- Madrigal, J.L.; Olivenza, R.; Moro, M.A.; Lizasoain, I.; Lorenzo, P.; Rodrigo, J.; Leza, J.C. Glutathione depletion, lipid peroxidation and mitochondrial dysfunction are induced by chronic stress in rat brain. Neuropsychopharmacology 2001, 24, 420–429. [Google Scholar] [CrossRef]
- Yuen, E.Y.; Wei, J.; Liu, W.; Zhong, P.; Li, X.; Yan, Z. Repeated stress causes cognitive impairment by suppressing glutamate receptor expression and function in prefrontal cortex. Neuron 2012, 73, 962–977. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, T.; Hossain, M.; Sanin, K.I. Global burden of maternal and child undernutrition and micronutrient deficiencies. Ann. Nutr. Metab. 2012, 61, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, T.; Chen, J.; Takahashi, T.; Ichitani, Y.; Nakahara, D. Prenatal stress suppresses cell proliferation in the early developing brain. Neuroreport 2006, 17, 1515–1518. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Mori, A. Stress, aging, and brain oxidative damage. Neurochem. Res. 1999, 24, 1479–1497. [Google Scholar] [CrossRef] [PubMed]
- Weinstock, M. The potential influence of maternal stress hormones on development and mental health of the offspring. Brain Behav. Immun. 2005, 19, 296–308. [Google Scholar] [CrossRef] [PubMed]
- Bowman, R.E.; MacLusky, N.J.; Sarmiento, Y.; Frankfurt, M.; Gordon, M.; Luine, V.N. Sexually dimorphic effects of prenatal stress on cognition, hormonal responses, and central neurotransmitters. Endocrinology 2004, 145, 3778–3787. [Google Scholar] [CrossRef] [PubMed]
- Felten, D.L.; Hallman, H.; Jonsson, G. Evidence for a neurotrophic role of noradrenaline neurons in the postnatal development of rat cerebral cortex. J. Neurocytol. 1982, 11, 119–135. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, L.K.; Turner, J.G.; Kalin, N.H. Prenatal stress alters brain catecholaminergic activity and potentiates stress-induced behavior in adult rats. Brain Res. 1992, 574, 131–137. [Google Scholar] [CrossRef]
- Rinne, J.O.; Portin, R.; Ruottinen, H.; Nurmi, E.; Bergman, J.; Haaparanta, M.; Solin, O. Cognitive impairment and the brain dopaminergic system in parkinson disease: [18f] fluorodopa positron emission tomographic study. Arch. Neurol. 2000, 57, 470–475. [Google Scholar] [CrossRef] [PubMed]
- Nieoullon, A. Dopamine and the regulation of cognition and attention. Prog. Neurobiol. 2002, 67, 53–83. [Google Scholar] [CrossRef]
- Cools, R. Role of dopamine in the motivational and cognitive control of behavior. Neuroscientist 2008, 14, 381–395. [Google Scholar] [CrossRef] [PubMed]
- Peters, D.A. Maternal stress increases fetal brain and neonatal cerebral cortex 5-hydroxytryptamine synthesis in rats: A possible mechanism by which stress influences brain development. Pharmacol. Biochem. Behav. 1990, 35, 943–947. [Google Scholar] [CrossRef]
- Madhyastha, S.K.; Prabhu, L.V.; Nayak, S.; Rai, R.; Pai, M.M.; Madhyastha, P.S. Effect of prenatal stress and serotonin depletion on postnatal serotonin metabolism in wistar rats. Iran. J. Pharmacol. Ther. 2008, 7, 71–77. [Google Scholar]
- Madhyastha, S.; Sahu, S.S.; Rao, G. Resveratrol for prenatal-stress-induced oxidative damage in growing brain and its consequences on survival of neurons. J. Basic Clin. Physiol. Pharmacol. 2014, 25, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Paintlia, M.K.; Paintlia, A.S.; Barbosa, E.; Singh, I.; Singh, A.K. N-acetylcysteine prevents endotoxin-induced degeneration of oligodendrocyte progenitors and hypomyelination in developing rat brain. J. Neurosci. Res. 2004, 78, 347–361. [Google Scholar] [CrossRef] [PubMed]
- Lante, F.; Meunier, J.; Guiramand, J.; Maurice, T.; Cavalier, M.; de Jesus Ferreira, M.-C.; Aimar, R.; Cohen-Solal, C.; Vignes, M.; Barbanel, G. Neurodevelopmental damage after prenatal infection: Role of oxidative stress in the fetal brain. Free Radic. Biol. Med. 2007, 42, 1231–1245. [Google Scholar] [CrossRef] [PubMed]
- Lante, F.; Meunier, J.; Guiramand, J.; De Jesus Ferreira, M.-C.L.; Cambonie, G.; Aimar, R.; Cohen-Solal, C.; Maurice, T.; Vignes, M.; Barbanel, G.R. Late N-acetylcysteine treatment prevents the deficits induced in the offspring of dams exposed to an immune stress during gestation. Hippocampus 2008, 18, 602–609. [Google Scholar] [CrossRef] [PubMed]
- Falluel-Morel, A.; Lin, L.; Sokolowski, K.; McCandlish, E.; Buckley, B.; DiCicco-Bloom, E. N-acetyl cysteine treatment reduces mercury-induced neurotoxicity in the developing rat hippocampus. J. Neurosci. Res. 2012, 90, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Neuwelt, E.A.; Pagel, M.A.; Hasler, B.P.; Deloughery, T.G.; Muldoon, L.L. Therapeutic efficacy of aortic administration of N-acetylcysteine as a chemoprotectant against bone marrow toxicity after intracarotid administration of alkylators, with or without glutathione depletion in a rat model. Cancer Res. 2001, 61, 7868–7874. [Google Scholar] [PubMed]
- Farr, S.A.; Poon, H.F.; Dogrukol-Ak, D.; Drake, J.; Banks, W.A.; Eyerman, E.; Butterfield, D.A.; Morley, J.E. The antioxidants α-lipoic acid and N-acetylcysteine reverse memory impairment and brain oxidative stress in aged samp8 mice. J. Neurochem. 2003, 84, 1173–1183. [Google Scholar] [CrossRef] [PubMed]
- Dean, O.M.; van den Buuse, M.; Berk, M.; Copolov, D.L.; Mavros, C.; Bush, A.I. N-acetyl cysteine restores brain glutathione loss in combined 2-cyclohexene-1-one and d-amphetamine-treated rats: Relevance to schizophrenia and bipolar disorder. Neurosci. Lett. 2011, 499, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Holson, R.; Pearce, B. Principles and pitfalls in the analysis of prenatal treatment effects in multiparous species. Neurotoxicol. Teratol. 1992, 14, 221–228. [Google Scholar] [CrossRef]
- Ampuero, E.; Luarte, A.; Santibañez, M.; Varas-Godoy, M.; Toledo, J.; Diaz-Veliz, G.; Cavada, G.; Rubio, F.J.; Wyneken, U. Two chronic stress models based on movement restriction in rats respond selectively to antidepressant drugs: Aldolase c as a potential biomarker. Int. J. Neuropsychopharmacol. 2015, 18, pyv038. [Google Scholar] [CrossRef] [PubMed]
- Amin, A.F.; Shaaban, O.M.; Bediawy, M.A. N-acetyl cysteine for treatment of recurrent unexplained pregnancy loss. Reprod. Biomed. Online 2008, 17, 722–726. [Google Scholar] [CrossRef]
- Basyigit, I.; Tugay, M.; Dilioglugil, M.; Yildiz, F.; Maral, H.; Sozubir, S. Protective effects of N-acetylcysteine on peroxidative changes of the fetal rat lungs whose mothers were exposed to cigarette smoke. Hum. Exp. Toxicol. 2007, 26, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Madhyastha, S.; Somayaji, S.; Nalini, K.; Nayak, B.; Bairy, K. Intraventricular methotrexate: Motor deficit in male wistar rats. Indian J. Pharmacol. 2002, 34, 350–355. [Google Scholar]
- Ambrosi, G.; Armentero, M.-T.; Levandis, G.; Bramanti, P.; Nappi, G.; Blandini, F. Effects of early and delayed treatment with an mGluR5 antagonist on motor impairment, nigrostriatal damage and neuroinflammation in a rodent model of parkinson’s disease. Brain Res. Bull. 2010, 82, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Kvarik, T.; Mammel, B.; Reglodi, D.; Antonelli, M.C.; Farkas, J.; Tamas, A.; Ertl, T.; Atlasz, T.; Bodzai, G.; Kiss, P. Effects of maternal stress during different periods of pregnancy on the early neurobehavioral response of rats. J. Neurol. Neurosci. 2016, 7, 1–8. [Google Scholar] [CrossRef]
- Sanchez, M.M. The impact of early adverse care on HPA axis development: Nonhuman primate models. Horm. Behav. 2006, 50, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Glutatyon, P.S.N.S.B. Effect of prenatal stress on expression of glutathione system in neonatal rat brain. Turk. Neurosurg. 2012, 22, 576–582. [Google Scholar]
- Weinstock, M. Sex-dependent changes induced by prenatal stress in cortical and hippocampal morphology and behaviour in rats: An update. Stress 2011, 14, 604–613. [Google Scholar] [CrossRef] [PubMed]
- Palacios-García, I.; Lara-Vásquez, A.; Montiel, J.F.; Díaz-Véliz, G.F.; Sepúlveda, H.; Utreras, E.; Montecino, M.; González-Billault, C.; Aboitiz, F. Prenatal stress down-regulates reelin expression by methylation of its promoter and induces adult behavioral impairments in rats. PLoS ONE 2015, 10, e0117680. [Google Scholar] [CrossRef] [PubMed]
- Zuena, A.R.; Mairesse, J.; Casolini, P.; Cinque, C.; Alemà, G.S.; Morley-Fletcher, S.; Chiodi, V.; Spagnoli, L.G.; Gradini, R.; Catalani, A. Prenatal restraint stress generates two distinct behavioral and neurochemical profiles in male and female rats. PLoS ONE 2008, 3, e2170. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, P.; Kus, K.; Murawiecka, P.; Słodzińska, I.; Giermaziak, W.; Nowakowska, E. Biochemical and cognitive impairments observed in animal models of schizophrenia induced by prenatal stress paradigm or methylazoxymethanol acetate administration. Acta Neurobiol. Exp. 2015, 75, 314–325. [Google Scholar]
- Barzegar, M.; Sajjadi, F.S.; Talaei, S.A.; Hamidi, G.; Salami, M. Prenatal exposure to noise stress: Anxiety, impaired spatial memory, and deteriorated hippocampal plasticity in postnatal life. Hippocampus 2015, 25, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Paris, J.J.; Frye, C.A. Juvenile offspring of rats exposed to restraint stress in late gestation have impaired cognitive performance and dysregulated progestogen formation. Stress 2011, 14, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Mina, T.H.; Reynolds, R.M. Mechanisms Linking in Utero Stress to Altered Offspring Behaviour. In Behavioral Neurobiology of Stress-Related Disorders; Springer: Berlin, Germany, 2014; pp. 93–122. [Google Scholar]
- Kim, Y.; Chung, C.W. Factors of prenatal depression by stress-vulnerability and stress-coping models. Korean J. Women Health Nurs. 2014, 20, 38–47. [Google Scholar] [CrossRef]
- Huizink, A.C.; Robles de Medina, P.G.; Mulder, E.J.; Visser, G.H.; Buitelaar, J.K. Stress during pregnancy is associated with developmental outcome in infancy. J. Child Psychol. Psychiatry 2003, 44, 810–818. [Google Scholar] [CrossRef] [PubMed]
- Andersen, S.L. Trajectories of brain development: Point of vulnerability or window of opportunity? Neurosci. Biobehav. Rev. 2003, 27, 3–18. [Google Scholar] [CrossRef]
- Polman, J.A.E.; Hunter, R.G.; Speksnijder, N.; van den Oever, J.M.; Korobko, O.B.; McEwen, B.S.; de Kloet, E.R.; Datson, N.A. Glucocorticoids modulate the mtor pathway in the hippocampus: Differential effects depending on stress history. Endocrinology 2012, 153, 4317–4327. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Li, X.; Chen, W.; Zhao, Y.; Li, H.; Qing, C.; Jia, N.; Bai, Z.; Liu, J. Prenatal stress causes gender-dependent neuronal loss and oxidative stress in rat hippocampus. J. Neurosci. Res. 2004, 78, 837–844. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, L.K.; Madhyastha, S.; Bairy, L.; Kishore, A. Status of the brain antioxidant system at different growing periods after prenatal stress and N-acetyl cysteine administration. Folia Neuropathol. 2017, 55, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Adair, J.C.; Knoefel, J.E.; Morgan, N. Controlled trial of n-acetylcysteine for patients with probable alzheimer’s disease. Neurology 2001, 57, 1515–1517. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.-J.; Leão, P.; Carvalho, M.; Almeida, O.F.; Sousa, N. Potential programming of dopaminergic circuits by early life stress. Psychopharmacology 2011, 214, 107–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voorn, P.; Kalsbeek, A.; Jorritsma-Byham, B.; Groenewegen, H. The pre-and postnatal development of the dopaminergic cell groups in the ventral mesencephalon and the dopaminergic innervation of the striatum of the rat. Neuroscience 1988, 25, 857–887. [Google Scholar] [CrossRef]
- Prakash, N.; Wurst, W. Development of dopaminergic neurons in the mammalian brain. Cell. Mol. Life Sci. CMLS 2006, 63, 187–206. [Google Scholar] [CrossRef] [PubMed]
- Davis, K.L.; Kahn, R.S.; Ko, G.; Davidson, M. Dopamine in schizophrenia: A review and reconceptualization. Am. J. Psychiatry 1991, 148, 1414. [Google Scholar]
- Holsboer, F. Stress, hypercortisolism and corticosteroid receptors in depression: Implicatons for therapy. J. Affect. Disord. 2001, 62, 77–91. [Google Scholar] [CrossRef]
- Heim, C.; Nemeroff, C.B. The role of childhood trauma in the neurobiology of mood and anxiety disorders: Preclinical and clinical studies. Biol. Psychiatry 2001, 49, 1023–1039. [Google Scholar] [CrossRef]
- Baier, C.J.; Katunar, M.R.; Adrover, E.; Pallarés, M.E.; Antonelli, M.C. Gestational restraint stress and the developing dopaminergic system: An overview. Neurotox. Res. 2012, 22, 16–32. [Google Scholar] [CrossRef] [PubMed]
- Son, G.H.; Chung, S.; Geum, D.; Kang, S.S.; Choi, W.S.; Kim, K.; Choi, S. Hyperactivity and alteration of the midbrain dopaminergic system in maternally stressed male mice offspring. Biochem. Biophys. Res. Commun. 2006, 352, 823–829. [Google Scholar] [CrossRef] [PubMed]
- De Souza, M.A.; Szawka, R.E.; Centenaro, L.A.; Diehl, L.A.; Lucion, A.B. Prenatal stress produces sex differences in nest odor preference. Physiol. Behav. 2012, 105, 850–855. [Google Scholar] [CrossRef] [PubMed]
- Chandramani Shivalingappa, P.; Jin, H.; Anantharam, V.; Kanthasamy, A.; Kanthasamy, A. N-acetyl cysteine protects against methamphetamine-induced dopaminergic neurodegeneration via modulation of redox status and autophagy in dopaminergic cells. Parkinson’s Dis. 2012, 2012, 424285. [Google Scholar] [CrossRef] [PubMed]
- Bharath, S.; Hsu, M.; Kaur, D.; Rajagopalan, S.; Andersen, J.K. Glutathione, iron and parkinson’s disease. Biochem. Pharmacol. 2002, 64, 1037–1048. [Google Scholar] [CrossRef]
- Berman, A.E.; Chan, W.Y.; Brennan, A.M.; Reyes, R.C.; Adler, B.L.; Suh, S.W.; Kauppinen, T.M.; Edling, Y.; Swanson, R.A. N-acetylcysteine prevents loss of dopaminergic neurons in the eaac1−/− mouse. Ann. Neurol. 2011, 69, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Gere-Paszti, E.; Jakus, J. The effect of n-acetylcysteine on amphetamine-mediated dopamine release in rat brain striatal slices by ion-pair reversed-phase high performance liquid chromatography. Biomed. Chromatogr. 2009, 23, 658–664. [Google Scholar] [CrossRef] [PubMed]
- Spencer, R.C.; Devilbiss, D.M.; Berridge, C.W. The cognition-enhancing effects of psychostimulants involve direct action in the prefrontal cortex. Biol. Psychiatry 2015, 77, 940–950. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, K.; Tsukada, H.; Nishiyama, S.; Fukumoto, D.; Kakiuchi, T.; Shimizu, E.; Iyo, M. Effects of N-acetyl-l-cysteine on the reduction of brain dopamine transporters in monkey treated with methamphetamine. Ann. N. Y. Acad. Sci. 2004, 1025, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Janáky, R.; Dohovics, R.; Saransaari, P.; Oja, S.S. Modulation of [3 h] dopamine release by glutathione in mouse striatal slices. Neurochem. Res. 2007, 32, 1357–1364. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CON | NAC | EGS | LGS | EGS + NAC | LGS + NAC | |
|---|---|---|---|---|---|---|
| Gestational length (n = 12) | 21.66 ± 0.14 | 21.41 ± 0.15 | 21.66 ± 0.14 | 21.5 ± 0.19 | 21.58 ± 0.14 | 21.5 ± 0.15 |
| Litter size | 9.66 ± 0.22 | 9.25 ± 0.65 | 8.5 ± 0.65 | 9.08 ± 0.65 | 9.66 ± 0.22 | 9.58 ± 0.22 |
| Day of upper incisor eruption (n = 24) | 7.95 ± 0.35 | 7.70 ± 0.55 | 8.29 ± 0.46 | 8.08 ± 0.82 | 7.95 ± 0.55 | 8.04 ± 0.55 |
| Day of eye opening (n = 24) | 8.20 ± 0.72 | 8.16 ± 0.13 | 8.41 ± 0.71 | 8.62 ± 0.57 | 8.12 ± 0.67 | 8.12 ± 0.74 |
| Animal Groups | Mean Number of Pups | Still Born Pups | Until Post-Natal Day 7 | Post-Natal Day 7 to 21 |
|---|---|---|---|---|
| CON | 9.6 | 8.3% | 0% | 0% |
| NAC | 9.2 | 16% | 0% | 0% |
| EGS | 8.5 | 5.8% | 0% | 0% |
| LGS | 9.08 | 5.5% | 0% | 0% |
| EGS + NAC | 9.6 | 9.75% | 0% | 0% |
| LGS + NAC | 9.6 | 6.09% | 0% | 0% |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bernhardt, L.K.; Bairy, K.L.; Madhyastha, S. Neuroprotective Role of N-acetylcysteine against Learning Deficits and Altered Brain Neurotransmitters in Rat Pups Subjected to Prenatal Stress. Brain Sci. 2018, 8, 120. https://doi.org/10.3390/brainsci8070120
Bernhardt LK, Bairy KL, Madhyastha S. Neuroprotective Role of N-acetylcysteine against Learning Deficits and Altered Brain Neurotransmitters in Rat Pups Subjected to Prenatal Stress. Brain Sciences. 2018; 8(7):120. https://doi.org/10.3390/brainsci8070120
Chicago/Turabian StyleBernhardt, Liegelin Kavitha, K. Lakshminarayana Bairy, and Sampath Madhyastha. 2018. "Neuroprotective Role of N-acetylcysteine against Learning Deficits and Altered Brain Neurotransmitters in Rat Pups Subjected to Prenatal Stress" Brain Sciences 8, no. 7: 120. https://doi.org/10.3390/brainsci8070120