
Non-Nutrient, Naturally Occurring Phenolic Compounds with Antioxidant Activity for the Prevention and Treatment of Periodontal Diseases


Abstract
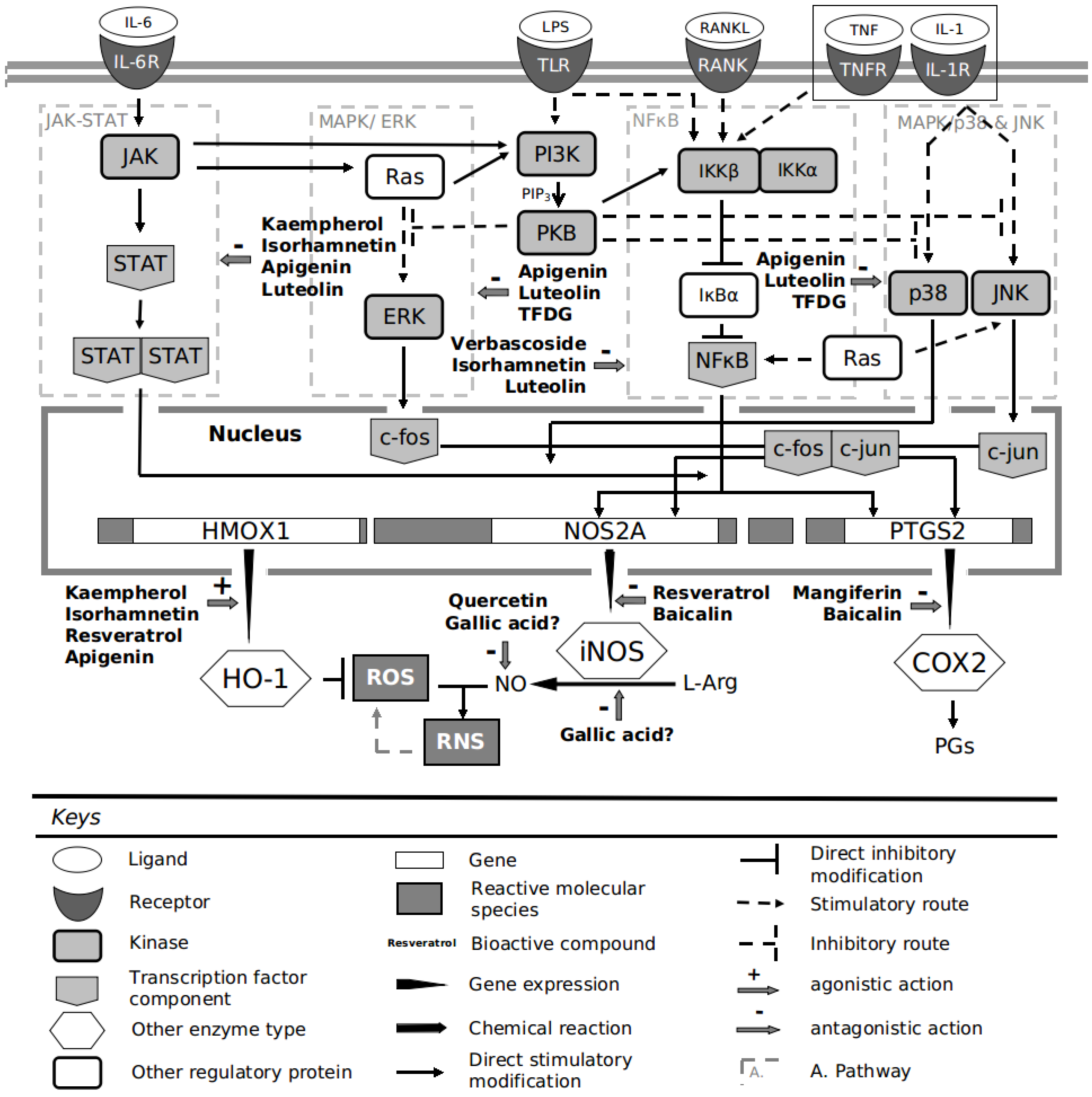
:1. Introduction
2. Thymoquinone
3. Coenzyme Q
4. Gallic Acid
5. Hydroxytyrosol
6. Mangiferin
7. Resveratrol
8. Verbascoside
9. Flavonoids
9.1. Flavonols
9.1.1. Quercetin
9.1.2. Kaempferol
9.1.3. Isorhamnetin
9.2. Flavones
9.2.1. Luteolin

9.2.2. Baicalin
9.2.3. Apigenin
9.2.4. Nobiletin and Tangeretin
9.3. Flavans
9.3.1. Catechins
9.3.2. Theaflavins
9.3.3. Proanthocyanidins
10. Conclusions

{kind=link}
| Compound | Study Type | Sample Studied (age/weight), n | Adminitration (Dosage, Frecuency and Duration) | Main Effect/s | Reference |
|---|---|---|---|---|---|
| Thymoquinone | Animal | Male Wistar rats with ligature-induced periodontitis (300 ± 10 g), n = 8 | Intragastric (10 mg/kg, daily for 11 days) | ABL, inflammatory infiltrate and osteoclasts n° were reduced; osteoblasts activity was maintained | [25] |
| Coenzyme Q | NCT | Patients under routine care for periodontitis (N/A) n = 8 | Oral (N/A) | Periodontal inflammation and periodontal pocket depth decreased | [46] |
| RCT | Chronic periodontitis patients (20–55 year), n = 3–6 | Topical (CoQ10 in a vegetable oil base in ratio of 1:9 in gel-form for 2/4 weeks) | Plaque index, gingival index, gingival bleeding index, periodontal probing pocket depth, and clinical attachment level were improved | [38,51] | |
| Animal | Male Fischer rats without additional treatments (8/16 weeks) n = 24 | Topical (ointment with 1% rCoQ10 daily for 2/4 months) | Osteoclast differentiation associated to aging decreased. The same conditions lowered gene periodontal expression of caspase-1 and IL-1β | [50] | |
| Gallic acid | In vitro | RAW 264.7 cells stimulated with LPS of A. actinomycetemcomitans or F. nucleatum | Preincubation (4 mg/ml for 2 h) | NO production was strongly inhibited | [60] |
| Hidroxytyrosol | Animal | New Zealand White rabbits with dietary-induced atherosclerosis (2.5 kg), n = 8 | Dietary supplementation (25 mg/kg daily for 30 days) | Endothelial activation was decreased | [66] |
| Mangiferin | Animal | Male Wistar rats with ligature-induced periodontitis (180 g), n = 4–7 | Oral (100 mg/kg daily for 7 days) | ABL and cellularity were reduced. COX-2 expression and the inhibition of rolling and adhesion of leukocyte | [82] |
| Resveratrol | Animal | Male Wistar rats with ligature-induced periodontitis (8 weeks), n = 6 | Oral supplementation (10 mg/kg daily melinjo resveratrol in drinking water for 20 days) | ABL was prevented and systemic levels of 8OH-dG, dityrosine, nitrotyrosine and proinflammatory cytokines (IL-1β IL-6, and TNF-α) were reduced | [92] |
| Animal | Male Wistar ratswith ligature-induced periodontitis (12 weeks), n = 12 | Intragastric (10 mg/kg daily for 30 days) | ABL and gingival IL-17 were reduced, but L-1β and IL-4 levels were unaffected | [28] | |
| Verbascoside | Animal | Male Sprague–Dawley rats with ligature-induced periodontitis (280–400 g), n = 10 | Oral (2 mg/kg daily for 8 days) | Gingivomucosal tissue injury and multiple inflammation and oxidative stress markers (MPO activity, activation of poly(ADP-ribose) polymerase TBARS, nitration of tyrosine residues, expression of NF-κB , iNOS, Bax and Bcl-2) were decreased | [98] |
| Quercetin | Animal | Male Sprague–Dawley rats LPS-injected (6 weeks), n = 4 | Oral (75 mg/kg, daily for 8/12 days) | Osteoclasts induction and inflammatory infiltrate were reduced. Bone crest levels were more apical and coronal alveolar level was higher than in the controls | [116] |
| Animal | Male Balb/c mice with A. actinomycetemcomitans infection (6–7 weeks), n = 5 | Subcutaneous injections (100 mg/kg daily for 15 days) | ABL and gingival levels of IL-1β, TNF-α, IL-17, RANKL, and ICAM-1 were reduced | [109] | |
| Kaempferol | In vitro | RAW246.7 cell stimulated by LPS from P. intermedia | Coincubation (10–100 μM for 24 h) | NO production was strongly inhibited | [124] |
| Isorhamnetin | In vitro | RAW264.7 cells stimulated by LPS from P. intermedia | Coincubation (12.5–50 μM for 24 h) | IL-6 mRNA and protein levels were down-regulated | [129] |
| Luteolin | In vitro | RAW264.7 cells stimulated by LPS from E. coli | Preincubation (10 μM for 30 min/60 min) | Activation of several MAPK family members and TNF-a release were reduced | [131] |
| In vitro | Human gingival fibroblasts stimulated by LPS from Salmonella enteritidis | Preincubation (10 μM for 30 min) | NO synthesis and cellular proliferation inhibition were suppressed | [132] | |
| Baicalin | Animal | Male Sprague–Dawley rats with ligature-induced periodontitis (8 weeks), n = 9 groups | Intragastric (50, 100 or 200 mg/kg, daily for 7 days) | ABL and collagen fibers loss were reduced; which was associated with COX-2 and iNOS levels down-regulation | [134] |
| In vitro | Human periodontal ligament cells and gingival fibroblats stimulated by IL-1β | Coincubation (N/A for 1 h) | Pro-MMP-1 secretion and MMP-3 expression were reduced | [136] | |
| In vitro | RAW 264.7 cells stimulated by LPS | Coincubation (5–40 μM for 24 h) | iNOS expression was reduced | [141] | |
| Apigenin | In vitro | Human periodontal ligament cells stimulated by nicotine and LPS from P. gingivalis | Preincubation (40 μM for 4 h) | NO production and up-regulation of iNOS, COX-2 and several proinflammatory factors (PGE2, IL-1β, TNF-α, IL-6, and IL-12) were inhibited | [144] |
| Nobiletin and tangeretin | In vitro | RAW264.7 cells stimulated by LPS | (30 μM for 5 days) in the presence of sRANKL (100 ng/mL) | Osteoclast formation and bone resorption and RANKL-induced osteoclastogenesis was reduced | [150] |
| Catechins | RCT | Volunteers with advanced periodontitis but with no systemic disorders (41–64 years), n = 6 | Topical (HPC strips containing GTC, once a w for 8 weeks) | Pocket depth and the proportion of BPR were decreased in combination with mechanical treatment | [185] |
| Animal | Male Sprague–Dawley Rats with ligature-induced periodontitis (150–180 g), n = 24 | Intragastric (200 mg/kg EGCG daily for 1/2/4 weeks) | ABL and collagen destruction were decreased which was associated to a reduction of IL-6 and TNF-α expression | [183] | |
| Animal | Male Wistar rats with periodontal inflammation induced by LPS from E. coli and proteases from S. griseus (8 weeks), n = 11 | Topical (dentifrice with 1.0% GTC daily for 4 weeks) | Inflammatory infiltrate was reduced which was accompanied by lower gingival hexanoyl-lysine, nitrotyrosine, and TNF-α | [180] | |
| Animal | Male BALB/c mice receiving gingival injections of LPS from E. coli (7 weeks), n = 6 | Oral supplementation (in drinking water with 1% Sunphenon BG 1 free acess for 20 days) | ABL and IL-1β expression were decreased | [182] | |
| Animal | Male BALB/c mice receiving gingival injections of LPS from E. coli (7 weeks), n = 6 | Gingival injection (1% Sunphenon BG 1 every 48 h for 20 days) | ABL and IL-1β expression were decreased | [182] | |
| In vitro | Osteoclasts from mouse bone marrow macrophagesstimulated by RANKL | Coincubation (2–10 μg/mL Sunphenon BG 1 for 4 days) | Osteoclasts n° decreased in a dose-dependent manner | [182] | |
| In vitro | RAW264.7 cells stimulated by LPS from E. coli | Preincubation (50 mmol/L of EGCG for 2 h) | TNFα production and mRNA expression were decreased as well as nuclear NF-κB–binding activity | [195] | |
| In vitro | Human gingival fibroblasts stimulated by oncostatin M | Preincubation (5/50 μg/mL EGCG or ECG, for 1 h) | CXCL10 production was prevented | [193] | |
| In vitro | Human gingival fibroblast stimulated by tumor necrosis factor superfamily 14 | Preincubation (5/50 mg/mL EGCG or ECG for 1 h) | IL-6 production was prevented | [199] | |
| Theaflavins | In vitro | Human gingival fibroblast stimulated by tumor necrosis factor superfamily 14 | Preincubation (5/50 mg/mL TFDG for 1 h) | IL-6 production was prevented | [199] |
| Proanthocyanidins | In vitro | Human macrophages stimulated by LPS from different periodontopathogens 2 | Preincubation (10–50 μg/mL of a Proanthocyanidin-rich cranberry fraction for 2 h) | IL-1β, IL-6, IL-8, TNF -α, and RANTES production was reduced | [206] |
| In vitro | Human gingival fibroblasts stimulated by LPS from A. actinomycetemcomitans | Preincubation (10-50 μg/mL of a Proanthocyanidin-rich cranberry fraction for 2 h) | IL-6, IL-8, and PGE2 production and increase of COX-2 expression were inhibited | [207] | |
| In vitro | Human macrophages and gingival fibroblasts stimulated by LPS from A. actinomycetemcomitans | Preicubation (10–150 μg/mL of a proanthocyanidin-rich cranberry fraction for 2 h) | MMP-3 and MMP-9 production and activity were inhibited | [208] | |
| In vitro | RAW 264.7 cells, simulated by LPS from A. actinomycetemcomitans or F. nucleatum | Grape seed proanthocyanidin extract (4 mg/mL for 2 h) | NO and ROS production and iNOS l levels decreased | [60] |
Acknowledgments
Conflicts of Interest
References
- Williams, R.C. Periodontal disease. N. Engl. J. Med. 1990, 322, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Page, R.C.; Kornman, K.S. The pathogenesis of human periodontitis: An introduction. Periodontology 2000 1997, 14, 9–11. [Google Scholar] [CrossRef] [PubMed]
- Bullon, P.; Newman, H.N.; Battino, M. Obesity, diabetes mellitus, atherosclerosis and chronic periodontitis: A shared pathology via oxidative stress and mitochondrial dysfunction? Periodontology 2000 2014, 64, 139–153. [Google Scholar] [CrossRef] [PubMed]
- Paquette, D.W.; Brodala, N.; Nichols, T.C. Cardiovascular disease, inflammation, and periodontal infection. Periodontology 2000 2007, 44, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Bullon, P.; Cordero, M.D.; Quiles, J.L.; Ramirez-Tortosa, M.C.; Gonzalez-Alonso, A.; Alfonsi, S.; García-Marín, R.; de Miguel, M.; Battino, M. Autophagy in periodontitis patients and gingival fibroblasts: Unraveling the link between chronic diseases and inflammation. BMC Med. 2012, 10, 122. [Google Scholar] [CrossRef] [PubMed]
- Battino, M.; Bullon, P.; Wilson, M.; Newman, H. Oxidative injury and inflammatory periodontal diseases: The challenge of anti-oxidants to free radicals and reactive oxygen species. Crit. Rev. Oral Biol. Med. 1999, 10, 458–476. [Google Scholar] [CrossRef] [PubMed]
- Chapple, I.L.; Milward, M.R.; Dietrich, T. The prevalence of inflammatory periodontitis is negatively associated with serum antioxidant concentrations. J. Nutr. 2007, 137, 657–664. [Google Scholar] [PubMed]
- D’aiuto, F.; Nibali, L.; Parkar, M.; Patel, K.; Suvan, J.; Donos, N. Oxidative stress, systemic inflammation, and severe periodontitis. J. Dent. Res. 2010, 89, 1241–1246. [Google Scholar] [CrossRef] [PubMed]
- Matthews, J.B.; Wright, H.J.; Roberts, A.; Cooper, P.R.; Chapple, I.L. Hyperactivity and reactivity of peripheral blood neutrophils in chronic periodontitis. Clin. Exp. Immunol. 2007, 147, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Matthews, J.B.; Wright, H.J.; Roberts, A.; Ling-Mountford, N.; Cooper, P.R.; Chapple, I.L. Neutrophil hyper-responsiveness in periodontitis. J. Dent. Res. 2007, 86, 718–722. [Google Scholar] [CrossRef] [PubMed]
- Bullon, P.; Cordero, M.D.; Quiles, J.L.; Morillo, J.M.; del Carmen Ramirez-Tortosa, M.; Battino, M. Mitochondrial dysfunction promoted by porphyromonas gingivalis lipopolysaccharide as a possible link between cardiovascular disease and periodontitis. Free Radic. Biol. Med. 2011, 50, 1336–1343. [Google Scholar] [CrossRef] [PubMed]
- Battino, M.; Ferreiro, M.-S.; Bompadre, S.; Leone, L.; Mosca, F.; Bullon, P. Elevated hydroperoxide levels and antioxidant patterns in Papillon-Lefèvre syndrome. J. Periodontol. 2001, 72, 1760–1766. [Google Scholar] [CrossRef] [PubMed]
- Baltacioğlu, E.; Akalin, F.A.; Alver, A.; Değer, O.; Karabulut, E. Protein carbonyl levels in serum and gingival crevicular fluid in patients with chronic periodontitis. Arch. Oral Biol. 2008, 53, 716–722. [Google Scholar] [CrossRef] [PubMed]
- Uno, K.; Nicholls, S.J. Biomarkers of inflammation and oxidative stress in atherosclerosis. Biomark. Med. 2010, 4, 361–373. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Tortosa, M.C.; Quiles, J.L.; Battino, M.; Granados, S.; Morillo, J.M.; Bompadre, S.; Newman, H.N.; Bullon, P. Periodontitis is associated with altered plasma fatty acids and cardiovascular risk markers. Nutr. Metab. Cardiovasc. Dis. NMCD 2010, 20, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Battino, M.; Ferreiro, M.S.; Quiles, J.L.; Bompadre, S.; Leone, L.; Bullon, P. Alterations in the oxidation products, antioxidant markers, antioxidant capacity and lipid patterns in plasma of patients affected by Papillon-Lefèvre syndrome. Free Radic. Res. 2003, 37, 603–609. [Google Scholar] [CrossRef] [PubMed]
- Chapple, I.L. Reactive oxygen species and antioxidants in inflammatory diseases. J. Clin. Periodontol. 1997, 24, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.V.; Puleo, D.A. Infection, inflammation, and bone regeneration. J. Dent. Res. 2011, 90, 1052–1061. [Google Scholar] [CrossRef] [PubMed]
- Kendall, H.; Marshall, R.; Bartold, P. Nitric oxide and tissue destruction. Oral Dis. 2001, 7, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.P.; McAndrew, J.; Sellak, H.; White, C.R.; Jo, H.; Freeman, B.A.; Darley-Usmar, V.M. Biological aspects of reactive nitrogen species. Biochim. Biophys. Acta 1999, 1411, 385–400. [Google Scholar] [CrossRef]
- Janssen-Heininger, Y.M.; Poynter, M.E.; Baeuerle, P.A. Recent advances towards understanding redox mechanisms in the activation of nuclear factor kappaB. Free Radic. Biol. Med. 2000, 28, 1317–1327. [Google Scholar] [CrossRef]
- Brock, G.R.; Butterworth, C.J.; Matthews, J.B.; Chapple, I.L. Local and systemic total antioxidant capacity in periodontitis and health. J. Clin. Periodontol. 2004, 31, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Panjamurthy, K.; Manoharan, S.; Ramachandran, C.R. Lipid peroxidation and antioxidant status in patients with periodontitis. Cell. Mol. Biol. Lett. 2005, 10, 255–264. [Google Scholar] [PubMed]
- Palmer, R.M.; Wilson, R.F.; Hasan, A.S.; Scott, D.A. Mechanisms of action of environmental factors--tobacco smoking. J. Clin. Periodontol. 2005, 32 (Suppl. 6), 180–195. [Google Scholar] [CrossRef] [PubMed]
- Ozdemir, H.; Kara, M.I.; Erciyas, K.; Ozer, H.; Ay, S. Preventive effects of thymoquinone in a rat periodontitis model: A morphometric and histopathological study. J. Periodontal Res. 2012, 47, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.-Y.; Jin, J.-Y.; Choi, J.-I.; Choi, I.S.; Kim, S.-J. Effects of luteolin on the release of nitric oxide and interleukin-6 by macrophages stimulated with lipopolysaccharide from Prevotella intermedia. J. Periodontol. 2011, 82, 1509–1517. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, A.; Bevilacqua, N.; Guida, L.; Annunziata, M.; Romano Carratelli, C.; Paolillo, R. Effect of resveratrol and modulation of cytokine production on human periodontal ligament cells. Cytokine 2012, 60, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Casati, M.Z.; Algayer, C.; Cardoso da Cruz, G.; Ribeiro, F.V.; Casarin, R.C.V.; Pimentel, S.P.; Cirano, F.R. Resveratrol decreases periodontal breakdown and modulates local levels of cytokines during periodontitis in rats. J. Periodontol. 2013, 84, e58–e64. [Google Scholar] [CrossRef] [PubMed]
- Nader, M.A.; El-Agamy, D.S.; Suddek, G.M. Protective effects of propolis and thymoquinone on development of atherosclerosis in cholesterol-fed rabbits. Arch. Pharm. Res. 2010, 33, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Tekeoglu, I.; Dogan, A.; Ediz, L.; Budancamanak, M.; Demirel, A. Effects of thymoquinone (volatile oil of black cumin) on rheumatoid arthritis in rat models. Phytother. Res. 2007, 21, 895–897. [Google Scholar] [CrossRef] [PubMed]
- Cluis, C.P.; Burja, A.M.; Martin, V.J.J. Current prospects for the production of coenzyme Q10 in microbes. Trends Biotechnol. 2007, 25, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Bhagavan, H.N.; Chopra, R.K. Coenzyme Q10: Absorption, tissue uptake, metabolism and pharmacokinetics. Free Radic. Res. 2006, 40, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Gaby, A.R. The role of coenzyme Q10 in clinical medicine: Part I. Altern. Med. Rev. 1996, 1, 11–17. [Google Scholar]
- Crane, F.L. Biochemical functions of coenzyme Q10. J. Am. Coll. Nutr. 2001, 20, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Shahla, A.H. Coenzyme Q10: A review. Hosp. Pharm. 2000, 35, 51–55. [Google Scholar]
- Tomasetti, M.; Alleva, R.; Borghi, B.; Collins, A.R. In vivo supplementation with coenzyme Q10 enhances the recovery of human lymphocytes from oxidative DNA damage. FASEB J. 2001, 15, 1425–1427. [Google Scholar] [CrossRef] [PubMed]
- Littarru, G.P.; Tiano, L. Bioenergetic and antioxidant properties of coenzyme Q10: Recent developments. Mol. Biotechnol. 2007, 37, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Hans, M.; Prakash, S.; Gupta, S. Clinical evaluation of topical application of Perio-Q gel coenzyme Q10 in chronic periodontitis patients. J. Indian Soc. Periodontol. 2012, 16, 193–199. [Google Scholar] [PubMed]
- Blatt, T.; Littarru, G.P. Biochemical rationale and experimental data on the antiaging properties of CoQ10 at skin level. BioFactors 2011, 37, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Weant, K.A.; Smith, K.M. The role of coenzyme Q10 in heart failure. Ann. Pharmacother. 2005, 39, 1522–1526. [Google Scholar] [CrossRef] [PubMed]
- Dallner, G.; Brismar, K.; Chojnacki, T.; Swiezewska, E. Regulation of coenzyme Q biosynthesis and breakdown. Biofactors 2003, 18, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, R.; Littarru, G.P.; Folkers, K.; Wilkinson, E.G. Deficiency of coenzyme Q in gingiva of patients with periodontal disease. Int. J. Vitam. Nutr. Res. 1973, 43, 84–92. [Google Scholar] [PubMed]
- Littarru, G.P.; Nakamura, R.; Ho, L.; Folkers, K.; Kuzell, W.C. Deficiency of coenzyme Q 10 in gingival tissue from patients with periodontal disease. Proc. Natl. Acad. Sci. USA 1971, 68, 2332–2335. [Google Scholar] [CrossRef] [PubMed]
- Bentinger, M.; Tekle, M.; Dallner, G. Coenzyme Q—Biosynthesis and functions. Biochem. Biophys. Res. Commun. 2010, 396, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Battino, M.; Bompadre, S.; Politi, A.; Fioroni, M.; Rubini, C.; Bullon, P. Antioxidant status CoQ10 and Vit. E Levels and immunohistochemical analysis of soft tissues in periodontal diseases. BioFactors 2005, 25, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, E.G.; Arnold, R.M.; Folkers, K.; Hansen, I.; Kishi, H. Bioenergetics in clinical medicine. II. Adjunctive treatment with coenzyme Q in periodontal therapy. Res. Commun. Chem. Pathol .Pharmacol. 1975, 12, 111–123. [Google Scholar] [PubMed]
- Shizukuishi, S.; Hanioka, T.; Tsunemitsu, A.; Fukunaga, Y.; Kishi, T.; Sato, N. Clinical effect of coenzyme Q10 on periodontal disease; Evaluation of Oxygen Utilisation in Gingiva by Tissue Reflectance Spectrophotometry. Biomed. Clin. Asp. Coenzyme Q 1986, 5, 359–368. [Google Scholar]
- Denny, N.; Chapple, I.L.; Matthews, J.B. Antioxidant and anti-inflammatory effects of coenzyme Q10: A preliminary study. J. Dent. Res. 1999, 78, 543. [Google Scholar]
- Matthews-Brzozowska, T.; Kurhañska-Flisykowska, A.; Stopa, J. Healing of periodontal tissue assisted by coenzyme Q10 with vitamin E—Clinical and laboratory evaluation. Pharmacol Rep. 2007, 59, 257–260. [Google Scholar]
- Yoneda, T.; Tomofuji, T.; Ekuni, D.; Azuma, T.; Endo, Y.; Kasuyama, K.; Machida, T.; Morita, M. Anti-aging effects of co-enzyme Q10 on periodontal tissues. J. Dent. Res. 2013, 92, 735–739. [Google Scholar] [CrossRef] [PubMed]
- Sale, S.T.; Parvez, H.; Yeltiwar, R.K.R.; Vivekanandan, G.; Pundir, A.J.; Jain, P. A Comparative evaluation of topical and intrasulcular application of coenzyme Q10 Perio Q™ gel in chronic periodontitis patients: A clinical study. J. Indian Soc. Periodontol. 2014, 18, 461–465. [Google Scholar] [PubMed]
- Hanioka, T.; Tanaka, M.; Ojima, M.; Shizukuishi, S.; Folkers, K. Effect of topical application of coenzyme Q10 on adult periodontitis. Mol. Asp. Med. 1994, 15 (Suppl. 1), s241–s248. [Google Scholar] [CrossRef]
- Singh, J.; Rai, G.K.; Upadhyay, A.K.; Kumar, R.; Singh, K.P. Antioxidant phytochemicals in tomato lycopersicon esculentum. Indian J. Agric. Sci. 2004, 74, 3–5. [Google Scholar]
- Ma, J.; Luo, X.-D.; Protiva, P.; Yang, H.; Ma, C.; Basile, M.J.; Weinstein, I.B.; Kennelly, E.J. Bioactive novel polyphenols from the fruit of Manilkara zapota (Sapodilla). J. Nat. Prod. 2003, 66, 983–986. [Google Scholar] [CrossRef] [PubMed]
- Shahrzad, S.; Bitsch, I. Determination of some pharmacologically active phenolic acids in juices by high-performance liquid chromatography. J. Chromatogr. A 1996, 741, 223–231. [Google Scholar] [CrossRef]
- Eyles, A.; Davies, N.W.; Mitsunaga, T.; Mihara, R.; Mohammed, C. Role of eucalyptus globulus wound wood extractives: Evidence of superoxide dismutase-like activity. For. Pathol. 2004, 34, 225–232. [Google Scholar] [CrossRef]
- Murugananthan, M.; Raju, G.B.; Prabhakar, S. Removal of tannins and polyhydroxy phenols by electro-chemical techniques. J. Chem. Technol. Biotechnol. 2005, 80, 1188–1197. [Google Scholar] [CrossRef]
- Kim, D.-O.; Lee, K.W.; Lee, H.J.; Lee, C.Y. Vitamin C Equivalent antioxidant capacity VCEAC of phenolic phytochemicals. J. Agric. Food Chem. 2002, 50, 3713–3717. [Google Scholar] [CrossRef] [PubMed]
- Kroes, B.H.; van Den Berg, A.J.J.; Quarles van Ufford, H.C.; van Dijk, H.; Labadie, R.P. Anti-inflammatory activity of gallic acid. Planta Med. 1992, 58, 499–504. [Google Scholar] [CrossRef] [PubMed]
- Houde, V.; Grenier, D.; Chandad, F. Protective effects of grape seed proanthocyanidins against oxidative stress induced by lipopolysaccharides of periodontopathogens. J. Periodontol. 2006, 77, 1371–1379. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Tortosa, M.C.; Granados, S.; Quiles, J.L. Chemical composition, types and characteristics of olive oil. In Olive Oil and Health; Quiles, J.L., Ramirez-Tortosa, M.C., Yaqoob, P., Eds.; CABI Publishing: Oxford, UK, 2006; pp. 45–62. [Google Scholar]
- Brenes, M.; García, A.; García, P.; Garrido, A. Rapid and complete extraction of phenols from olive oil and determination by means of a coulometric electrode array system. J. Agric. Food Chem. 2000, 48, 5178–5183. [Google Scholar] [CrossRef] [PubMed]
- Owen, R.W.; Giacosa, A.; Hull, W.E.; Haubner, R.; Spiegelhalder, B.; Bartsch, H. The antioxidant/anticancer potential of phenolic compounds isolated from olive oil. Eur. J. Cancer 2000, 36, 1235–1247. [Google Scholar] [CrossRef]
- Briante, R.; Patumi, M.; Terenziani, S.; Bismuto, E.; Febbraio, F.; Nucci, R. Olea europaea L. leaf extract and derivatives: antioxidant properties. J. Agric. Food Chem. 2002, 50, 4934–4940. [Google Scholar] [CrossRef] [PubMed]
- Granados-Principal, S.; Quiles, J.L.; Ramirez-Tortosa, C.L.; Sanchez-Rovira, P.; Ramirez-Tortosa, M.C. Hydroxytyrosol: From laboratory investigations to future clinical trials. Nutr. Rev. 2010, 68, 191–206. [Google Scholar] [CrossRef] [PubMed]
- Bullon, P.; Quiles, J.L.; Morillo, J.M.; Rubini, C.; Goteri, G.; Granados-Principal, S.; Battino, M.; Ramirez-Tortosa, M. Gingival vascular damage in atherosclerotic rabbits: hydroxytyrosol and squalene benefits. Food Chem. Toxicol. 2009, 47, 2327–2331. [Google Scholar] [CrossRef] [PubMed]
- Bullon, P.; Battino, M.; Varela-Lopez, A.; Perez-Lopez, P.; Granados-Principal, S.; Ramirez-Tortosa, M.C.; Ochoa, J.J.; Cordero, M.D.; Gonzalez-Alonso, A.; Ramirez-Tortosa, C.L.; et al. Diets based on virgin olive oil or fish oil but not on sunflower oil prevent age-related alveolar bone resorption by mitochondrial-related mechanisms. PLoS ONE 2013, 8, e74234. [Google Scholar] [CrossRef] [PubMed]
- Quiles, J.L.; Farquharson, A.J.; Simpson, D.K.; Grant, I.; Wahle, K.W.J. Olive oil phenolics: Effects on DNA oxidation and redox enzyme mRNA in prostate cells. Br. J. Nutr. 2002, 88, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Quiles, J.L.; Ochoa, J.J.; Huertas, J.R.; López-Frías, M.; Mataix, J. Olive oil and mitochondrial oxidative stress: studies on adriamycin toxicity, physical exercise and ageing. In Olive Oil and Health; Quiles, J.L., Ramirez-Tortosa, M.C., Yaqoob, P., Eds.; CABI Publishing: Oxford, UK, 2006; pp. 119–151. [Google Scholar]
- Ochoa-Herrera, J.J.; Huertas, J.R.; Quiles, J.L.; Mataix, J. Dietary oils high in oleic acid, but with different non-glyceride contents, have different effects on lipid profiles and peroxidation in rabbit hepatic mitochondria. J. Nutr. Biochem. 2001, 12, 357–364. [Google Scholar] [CrossRef]
- Visioli, F.; Galli, C. Biological properties of olive oil phytochemicals. Crit. Rev. Food Sci. Nutr. 2002, 42, 209–221. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Jiménez, F.; Ruano, J.; Perez-Martinez, P.; Lopez-Segura, F.; Lopez-Miranda, J. The influence of olive oil on human health: Not a question of fat alone. Mol. Nutr. Food Res. 2007, 51, 1199–1208. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, G.M.; Re, L.; Giuliani, A.; Núñez-Sellés, A.J.; Davison, G.P.; León-Fernández, O.S. Protective effects of Mangifera indica L. extract, Mangiferin and selected antioxidants against TPA-induced biomolecules oxidation and peritoneal macrophage activation in mice. Pharmacol. Res. 2000, 42, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Andreu, G.P.; Delgado, R.; Velho, J.A.; Curti, C.; Vercesi, A.E. Iron complexing activity of mangiferin, a naturally occurring glucosylxanthone, inhibits mitochondrial lipid peroxidation induced by Fe 2+-citrate. Eur. J. Pharmacol. 2005, 513, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Coe, F.G.; Anderson, G.J. Screening of medicinal plants used by the garífuna of Eastern Nicaragua for bioactive compounds. J. Ethnopharmacol. 1996, 53, 29–50. [Google Scholar] [CrossRef]
- Dar, A.; Faizi, S.; Naqvi, S.; Roome, T.; Zikr-Ur-Rehman, S.; Ali, M.; Firdous, S.; Moin, S.T. Analgesic and antioxidant activity of mangiferin and its derivatives: The structure activity relationship. Biol. Pharm. Bull. 2005, 28, 596–600. [Google Scholar] [CrossRef] [PubMed]
- Ojewole, J.A.O. Antiinflammatory, analgesic and hypoglycemic effects of Mangifera indica Linn. (Anacardiaceae) stem-bark aqueous extract. Methods Find. Exp. Clin. Pharmacol. 2005, 27, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Ichiki, H.; Miura, T.; Kubo, M.; Ishihara, E.; Komatsu, Y.; Tanigawa, K.; Okada, M. New antidiabetic compounds, mangiferin and its glucoside. Biol. Pharm. Bull. 1998, 21, 1389–1390. [Google Scholar] [CrossRef] [PubMed]
- Guha, S.; Ghosal, S.; Chattopadhyay, U. Antitumor, immunomodulatory and anti-HIV effect of mangiferin, a naturally occurring glucosylxanthone. Chemotherapy 1996, 42, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Duang, X.; Wang, Q.; Zhou, X.; Huang, D. Mangiferin: A possible strategy for periodontal disease to therapy. Med. Hypotheses 2011, 76, 486–488. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Miyahara, T.; Tezuka, Y.; Namba, T.; Nemoto, N.; Tonami, S.; Seto, H.; Tada, T.; Kadota, S. The effect of Kampo formulae on bone resorption in vitro and in vivo. I. Active constituents of Tsu-Kan-Gan. Biol. Pharm. Bull. 1998, 21, 1322–1326. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, R.R.; Pellizzon, C.H.; Justulin, L.; Felisbino, S.L.; Vilegas, W.; Bruni, F.; Lopes-Ferreira, M.; Hiruma-Lima, C.A. Effect of Mangiferin on the development of periodontal disease: Involvement of lipoxin A4, anti-chemotaxic action in leukocyte rolling. Chem. Biol. Interact. 2009, 179, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Baur, J.A.; Sinclair, D.A. Therapeutic potential of resveratrol: The in vivo evidence. Nat. Rev. Drug Discov. 2006, 5, 493–506. [Google Scholar] [CrossRef] [PubMed]
- Pervaiz, S. Resveratrol: From grapevines to mammalian biology. FASEB J. 2003, 17, 1975–1985. [Google Scholar] [CrossRef] [PubMed]
- Bhat, K.P.L.; Pezzuto, J.M. Cancer chemopreventive activity of resveratrol. Ann. N. Y. Acad. Sci. 2002, 957, 210–229. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.U.; Casper, R.F.; Fritz, P.C.; Sukhu, B.; Ganss, B.; Girard, B.; Savouret, J.F.; Tenenbaum, H.C. Inhibition of dioxin effects on bone formation in vitro by a newly described aryl hydrocarbon receptor antagonist, resveratrol. J. Endocrinol. 2000, 167, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Soufi, F.G.; Sheervalilou, R.; Vardiani, M.; Khalili, M.; Alipour, M.R. Chronic resveratrol administration has beneficial effects in experimental model of type 2 diabetic rats. Endocr. Regul. 2012, 46, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.; Cai, L.; Udeani, G.O.; Slowing, K.V.; Thomas, C.F.; Beecher, C.W.; Fong, H.H.; Farnsworth, N.R.; Kinghorn, A.D.; Mehta, R.G.; et al. Cancer chemopreventive activity of resveratrol, a natural product derived from grapes. Science 1997, 275, 218–220. [Google Scholar] [CrossRef] [PubMed]
- Whitlock, N.C.; Baek, S.J. The anticancer effects of resveratrol: Modulation of transcription factors. Nutr. Cancer 2012, 64, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Breen, D.M.; Dolinsky, V.W.; Zhang, H.; Ghanim, H.; Guo, J.; Mroziewicz, M.; Tsiani, E.L.; Bendeck, M.P.; Dandona, P.; Dyck, J.R.B.; et al. Resveratrol inhibits neointimal formation after arterial injury through an endothelial nitric oxide synthase-dependent mechanism. Atherosclerosis 2012, 222, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Tseng, P.-C.; Hou, S.-M.; Chen, R.-J.; Peng, H.-W.; Hsieh, C.-F.; Kuo, M.-L.; Yen, M.-L. Resveratrol promotes osteogenesis of human mesenchymal stem cells by upregulating RUNX2 gene expression via the SIRT1/FOXO3A axis. J. Bone Miner. Res. 2011, 26, 2552–2563. [Google Scholar] [CrossRef] [PubMed]
- Tamaki, N.; Cristina Orihuela-Campos, R.; Inagaki, Y.; Fukui, M.; Nagata, T.; Ito, H.-O. Resveratrol improves oxidative stress and prevents the progression of periodontitis via the activation of the Sirt1/AMPK and the Nrf2/antioxidant defense pathways in a rat periodontitis model. Free Radic. Biol. Med. 2014, 75, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, S.R.; Lokhandwala, M.F.; Banday, A.A. Resveratrol prevents endothelial nitric oxide synthase uncoupling and attenuates development of hypertension in spontaneously hypertensive rats. Eur. J. Pharmacol. 2011, 667, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Huang, J.; Xie, X.; Wang, S.; Chen, C.; Shen, X.; Liu, P.; Huang, H. Sirt1 resists advanced glycation end products-induced expressions of fibronectin and TGF-β1 by activating the Nrf2/ARE pathway in glomerular mesangial cells. Free Radic. Biol. Med. 2013, 65, 528–540. [Google Scholar] [CrossRef] [PubMed]
- Howitz, K.T.; Bitterman, K.J.; Cohen, H.Y.; Lamming, D.W.; Lavu, S.; Wood, J.G.; Zipkin, R.E.; Chung, P.; Kisielewski, A.; Zhang, L.-L.; et al. Small molecule activators of sirtuins extend Saccharomyces cerevisiae lifespan. Nature 2003, 425, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Zong, Y.; Sun, L.; Liu, B.; Deng, Y.-S.; Zhan, D.; Chen, Y.-L.; He, Y.; Liu, J.; Zhang, Z.-J.; Sun, J.; et al. Resveratrol inhibits LPS-Induced MAPKs activation via activation of the phosphatidylinositol 3-kinase pathway in murine RAW 264.7 macrophage cells. PLoS ONE 2012, 7, e44107. [Google Scholar] [CrossRef] [PubMed]
- Bi, X.L.; Yang, J.Y.; Dong, Y.X.; Wang, J.M.; Cui, Y.H.; Ikeshima, T.; Zhao, Y.Q.; Wu, C.F. Resveratrol inhibits nitric oxide and TNF-alpha production by lipopolysaccharide-activated microglia. Int. Immunopharmacol. 2005, 5, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Paola, R.D.; Oteri, G.; Mazzon, E.; Crisafulli, C.; Galuppo, M.; Toso, R.D.; Pressi, G.; Cordasco, G.; Cuzzocrea, S. Effects of verbascoside, biotechnologically purified by Syringa vulgaris plant cell cultures, in a rodent model of periodontitis. J. Pharm. Pharmacol. 2011, 63, 707–717. [Google Scholar] [CrossRef] [PubMed]
- Akbay, P.; Calis, I.; Ündeger, Ü.; Basaran, N.; Basaran, A.A. In vitro immunomodulatory activity of verbascoside from Nepeta ucrainica L. Phytother. Res. 2002, 16, 593–595. [Google Scholar] [CrossRef] [PubMed]
- Feng, R.; Lu, Y.; Bowman, L.L.; Qian, Y.; Castranova, V.; Ding, M. Inhibition of activator protein-1, NF-κB, and MAPKs and induction of phase 2 detoxifying enzyme activity by chlorogenic acid. J. Biol. Chem. 2005, 280, 27888–27895. [Google Scholar] [CrossRef] [PubMed]
- Servili, M.; Esposto, S.; Fabiani, R.; Urbani, S.; Taticchi, A.; Mariucci, F.; Selvaggini, R.; Montedoro, G.F. Phenolic compounds in olive oil: Antioxidant, health and organoleptic activities according to their chemical structure. Inflammopharmacology 2009, 17, 76–84. [Google Scholar] [CrossRef] [PubMed]
- De Groot, H.; Rauen, U. Tissue injury by reactive oxygen species and the protective effects of flavonoids. Fundam. Clin. Pharmacol. 1998, 12, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Florit, M.; Monjo, M.; Ramis, J.M. Identification of quercitrin as a potential therapeutic agent for periodontal applications. J. Periodontol. 2014, 85, 966–974. [Google Scholar] [CrossRef] [PubMed]
- Graziani, Y.; Erikson, E.; Erikson, R.L. The effect of quercetin on the phosphorylation activity of the Rous sarcoma virus transforming gene product in vitro and in vivo. Eur. J. Biochem. 1983, 135, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Ferriola, P.C.; Cody, V.; Middleton, E., Jr. Protein kinase c inhibition by plant flavonoids. Kinetic mechanisms and structure-activity relationships. Biochem. Pharmacol. 1989, 38, 1617–1624. [Google Scholar] [CrossRef]
- Formica, J.V.; Regelson, W. Review of the biology of quercetin and related bioflavonoids. Food Chem. Toxicol. 1995, 33, 1061–1080. [Google Scholar] [CrossRef]
- Fotsis, T.; Pepper, M.S.; Aktas, E.; Breit, S.; Rasku, S.; Adlercreutz, H.; Wähälä, K.; Montesano, R.; Schweigerer, L. Flavonoids, dietary-derived inhibitors of cell proliferation and in vitro angiogenesis. Cancer Res. 1997, 57, 2916–2921. [Google Scholar] [PubMed]
- Middleton, E.; Kandaswami, C.; Theoharides, T.C. The effects of plant flavonoids on mammalian cells: Implications for inflammation, heart disease, and cancer. Pharmacol. Rev. 2000, 52, 673–751. [Google Scholar] [PubMed]
- Napimoga, M.H.; Clemente-Napimoga, J.T.; Macedo, C.G.; Freitas, F.F.; Stipp, R.N.; Pinho-Ribeiro, F.A.; Casagrande, R.; Verri, W.A. Quercetin inhibits inflammatory bone resorption in a mouse periodontitis model. J. Nat. Prod. 2013, 76, 2316–2321. [Google Scholar] [CrossRef] [PubMed]
- Geoghegan, F.; Wong, R.W.K.; Rabie, A.B.M. Inhibitory effect of quercetin on periodontal pathogens in vitro. Phytother. Res. PTR 2010, 24, 817–820. [Google Scholar] [PubMed]
- Häkkinen, S.H.; Kärenlampi, S.O.; Heinonen, I.M.; Mykkänen, H.M.; Törrönen, A.R. Content of the flavonols quercetin, myricetin, and kaempferol in 25 edible berries. J. Agric. Food Chem. 1999, 47, 2274–2279. [Google Scholar] [CrossRef] [PubMed]
- Boots, A.W.; Haenen, G.R.M.M.; Bast, A. Health effects of quercetin: From antioxidant to nutraceutical. Eur. J. Pharmacol. 2008, 585, 325–337. [Google Scholar] [CrossRef] [PubMed]
- Yu, E.S.; Min, H.J.; An, S.Y.; Won, H.Y.; Hong, J.H.; Hwang, E.S. Regulatory mechanisms of IL-2 and IFNgamma suppression by quercetin in T helper cells. Biochem. Pharmacol. 2008, 76, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, S.C. Quercetin: Potentials in the prevention and therapy of disease. Curr. Opin. Clin. Nutr. Metab. Care 2008, 11, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.W.K.; Rabie, A.B.M. Effect of quercetin on bone formation. J. Orthop. Res. 2008, 26, 1061–1066. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.-C.; Huang, R.-Y.; Chiang, C.-Y.; Chen, J.-K.; Liu, C.-H.; Chu, C.-L.; Fu, E. Ameliorative effect of quercetin on the destruction caused by experimental periodontitis in rats. J. Periodontal Res. 2010, 45, 788–795. [Google Scholar] [CrossRef] [PubMed]
- Takahama, U.; Hirota, S.; Oniki, T. Quercetin-dependent scavenging of reactive nitrogen species derived from nitric oxide and nitrite in the human oral cavity: Interaction of quercetin with salivary redox components. Arch. Oral Biol. 2006, 51, 629–639. [Google Scholar] [CrossRef] [PubMed]
- Guazelli, C.F.S.; Fattori, V.; Colombo, B.B.; Georgetti, S.R.; Vicentini, F.T.M.C.; Casagrande, R.; Baracat, M.M.; Verri, W.A. Quercetin-loaded microcapsules ameliorate experimental colitis in mice by anti-inflammatory and antioxidant mechanisms. J. Nat. Prod. 2013, 76, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Maciel, R.M.; Costa, M.M.; Martins, D.B.; França, R.T.; Schmatz, R.; Graça, D.L.; Duarte, M.M.M.F.; Danesi, C.C.; Mazzanti, C.M.; Schetinger, M.R.C.; et al. Antioxidant and anti-inflammatory effects of quercetin in functional and morphological alterations in streptozotocin-induced diabetic rats. Res. Vet. Sci. 2013, 95, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Calderon-Montano, J.M.; Burgos-Moron, E.; Pérez-Guerrero, C.; López-Lázaro, M. A review on the dietary flavonoid kaempferol. Mini Rev. Med. Chem. 2011, 11, 298–344. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.-K.; Yu, Y.-L.; Chen, K.-C.; Chang, W.-T.; Lee, M.-S.; Yang, M.-J.; Cheng, H.-C.; Liu, C.-H.; Chen, D.-C.; Chu, C.-L. Kaempferol from semen cuscutae attenuates the immune function of dendritic cells. Immunobiology 2011, 216, 1103–1109. [Google Scholar] [CrossRef] [PubMed]
- Park, S.E.; Sapkota, K.; Kim, S.; Kim, H.; Kim, S.J. Kaempferol acts through mitogen-activated protein kinases and protein kinase B/AKT to elicit protection in a model of neuroinflammation in BV2 microglial cells. Br. J. Pharmacol. 2011, 164, 1008–1025. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, J.; Samojedny, A.; Paul, M.; Pietsz, G.; Wilczok, T. Effect of kaempferol on the production and gene expression of monocyte chemoattractant protein-1 in J774.2 macrophages. Pharmacol. Rep. 2005, 57, 107–112. [Google Scholar] [PubMed]
- Choi, I.S.; Choi, E.-Y.; Jin, J.-Y.; Park, H.R.; Choi, J.-I.; Kim, S.-J. Kaempferol inhibits P. Intermedia lipopolysaccharide-induced production of nitric oxide through translational regulation in murine macrophages: Critical role of heme oxygenase-1-mediated ROS reduction. J. Periodontol. 2013, 84, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Pennington, J.A.T. Food composition databases for bioactive food components. J. Food Compos. Anal. 2002, 15, 419–434. [Google Scholar] [CrossRef]
- Morand, C.; Crespy, V.; Manach, C.; Besson, C.; Demigné, C.; Rémésy, C. Plasma metabolites of quercetin and their antioxidant properties. Am. J. Physiol. 1998, 275, R212–R219. [Google Scholar] [PubMed]
- Bao, M.; Lou, Y. Isorhamnetin prevent endothelial cell injuries from oxidized LDL via activation of p38MAPK. Eur. J. Pharmacol. 2006, 547, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Boesch-Saadatmandi, C.; Loboda, A.; Wagner, A.E.; Stachurska, A.; Jozkowicz, A.; Dulak, J.; Döring, F.; Wolffram, S.; Rimbach, G. Effect of quercetin and its metabolites isorhamnetin and quercetin-3-glucuronide on inflammatory gene expression: Role of MIR-155. J. Nutr. Biochem. 2011, 22, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.Y.; Choi, E.Y.; Park, H.R.; Choi, J.I.; Choi, I.S.; Kim, S.J. Isorhamnetin inhibits Prevotella intermedia lipopolysaccharide-induced production of interleukin-6 in murine macrophages via anti-inflammatory heme oxygenase-1 induction and inhibition of nuclear factor-κB and signal transducer and activator of transcription 1 activation. J. Periodontal Res. 2013, 48, 687–695. [Google Scholar] [PubMed]
- López-Lázaro, M. Distribution and biological activities of the flavonoid luteolin. Mini Rev. Med. Chem. 2009, 9, 31–59. [Google Scholar] [CrossRef] [PubMed]
- Xagorari, A.; Roussos, C.; Papapetropoulos, A. Inhibition of LPS-stimulated pathways in macrophages by the flavonoid luteolin. Br. J. Pharmacol. 2002, 136, 1058–1064. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Venegas, G.; Kawasaki-Cárdenas, P.; Arroyo-Cruz, S.R.; Maldonado-Frías, S. Luteolin inhibits lipopolysaccharide actions on human gingival fibroblasts. Eur. J. Pharmacol. 2006, 541, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Chiang, L.-C.; Ng, L.T.; Chiang, W.; Chang, M.-Y.; Lin, C.-C. Immunomodulatory activities of flavonoids, monoterpenoids, triterpenoids, iridoid glycosides and phenolic compounds of Plantago species. Planta Med. 2003, 69, 600–604. [Google Scholar] [PubMed]
- Ciesielska, E.; Gwardys, A.; Metodiewa, D. Anticancer, antiradical and antioxidative actions of novel Antoksyd S and its major components, baicalin and baicalein. Anticancer Res. 2002, 22, 2885–2891. [Google Scholar] [PubMed]
- Cai, X.; Li, C.; Du, G.; Cao, Z. Protective effects of baicalin on ligature-induced periodontitis in rats. J. Periodontal Res. 2008, 43, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Cao, Z.; Yang, R.; Shang, Z.; Jin, L.; Cobert, E.F. Effects of baicalin on the expression of pro-MMP-1 and MMP-3 in human gingival fibroblasts and periodontal ligament cells. Zhonghua Kou Qiang Yi Xue Za Zhi 2004, 39, 197–200. [Google Scholar] [PubMed]
- Hwang, J.-M.; Wang, C.-J.; Chou, F.-P.; Tseng, T.-H.; Hsieh, Y.-S.; Hsu, J.-D.; Chu, C.-Y. Protective effect of baicalin on tert-butyl hydroperoxide-induced rat hepatotoxicity. Arch. Toxicol. 2005, 79, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Krakauer, T.; Li, B.Q.; Young, H.A. The flavonoid baicalin inhibits superantigen-induced inflammatory cytokines and chemokines. FEBS Lett. 2001, 500, 52–55. [Google Scholar] [CrossRef]
- Kubo, M.; Matsuda, H.; Tanaka, M.; Kimura, Y.; Okuda, H.; Higashino, M.; Tani, T.; Namba, K.; Arichi, S. Studies on Scutellariae radix. VII. Anti-arthritic and anti-inflammatory actions of methanolic extract and flavonoid components from Scutellariae radix. Chem. Pharm. Bull. Tokyo 1984, 32, 2724–2729. [Google Scholar] [CrossRef] [PubMed]
- Kase, Y.; Saitoh, K.; Makino, B.; Hashimoto, K.; Ishige, A.; Komatsu, Y. Relationship between the antidiarrhoeal effects of hange-shashin-to and its active components. Phytother. Res. 1999, 13, 468–473. [Google Scholar] [CrossRef]
- Chen, Y.C.; Shen, S.C.; Chen, L.G.; Lee, T.J.; Yang, L.L. Wogonin, baicalin, and baicalein inhibition of inducible nitric oxide synthase and cyclooxygenase-2 gene expressions induced by nitric oxide synthase inhibitors and lipopolysaccharide. Biochem. Pharmacol. 2001, 61, 1417–1427. [Google Scholar] [CrossRef]
- Yano, S.; Umeda, D.; Yamashita, S.; Yamada, K.; Tachibana, H. Dietary apigenin attenuates the development of atopic dermatitis-like skin lesions in NC/Nga mice. J. Nutr. Biochem. 2009, 20, 876–881. [Google Scholar] [CrossRef] [PubMed]
- Conforti, F.; Rigano, D.; Menichini, F.; Loizzo, M.R.; Senatore, F. Protection against neurodegenerative diseases of Iris pseudopumila extracts and their constituents. Fitoterapia 2009, 80, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Jeong, G.-S.; Lee, S.-H.; Jeong, S.-N.; Kim, Y.-C.; Kim, E.-C. Anti-inflammatory effects of apigenin on nicotine- and lipopolysaccharide-stimulated human periodontal ligament cells via heme oxygenase-1. Int. Immunopharmacol. 2009, 9, 1374–1380. [Google Scholar] [CrossRef] [PubMed]
- Rezai-Zadeh, K.; Ehrhart, J.; Bai, Y.; Sanberg, P.R.; Bickford, P.; Tan, J.; Shytle, R.D. Apigenin and luteolin modulate microglial activation via inhibition of STAT1-induced CD40 expression. J. Neuroinflamm. 2008, 5, 41. [Google Scholar] [CrossRef] [PubMed]
- Ha, S.K.; Lee, P.; Park, J.A.; Oh, H.R.; Lee, S.Y.; Park, J.-H.; Lee, E.H.; Ryu, J.H.; Lee, K.R.; Kim, S.Y. Apigenin Inhibits the Production of NO and PGE2 in microglia and inhibits neuronal cell death in a middle cerebral artery occlusion-induced focal ischemia mice model. Neurochem. Int. 2008, 52, 878–886. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Zhang, L.; Bertucci, A.M.; Pope, R.M.; Datta, S.K. Apigenin, a dietary flavonoid, sensitizes human T cells for activation-induced cell death by inhibiting PKB/Akt and NF-κB activation pathway. Immunol. Lett. 2008, 121, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Nicholas, C.; Batra, S.; Vargo, M.A.; Voss, O.H.; Gavrilin, M.A.; Wewers, M.D.; Guttridge, D.C.; Grotewold, E.; Doseff, A.I. Apigenin blocks lipopolysaccharide-induced lethality in vivo and proinflammatory cytokines expression by inactivating NF-κB through the suppression of p65 phosphorylation. J. Immunol. 2007, 179, 7121–7127. [Google Scholar] [CrossRef] [PubMed]
- Akao, Y.; Itoh, T.; Ohguchi, K.; Iinuma, M.; Nozawa, Y. Interactive effects of polymethoxy flavones from citrus on cell growth inhibition in human neuroblastoma SH-SY5Y cells. Bioorg. Med. Chem. 2008, 16, 2803–2810. [Google Scholar] [CrossRef] [PubMed]
- Tominari, T.; Hirata, M.; Matsumoto, C.; Inada, M.; Miyaura, C. Polymethoxy flavonoids, nobiletin and tangeretin, prevent lipopolysaccharide-induced inflammatory bone loss in an experimental model for periodontitis. J. Pharmacol. Sci. 2012, 119, 390–394. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.-Q.; Zhou, C.-H.; Tao, J.; Li, S.-N. Antagonistic effects of nobiletin, a polymethoxyflavonoid, on eosinophilic airway inflammation of asthmatic rats and relevant mechanisms. Life Sci. 2006, 78, 2689–2696. [Google Scholar] [CrossRef] [PubMed]
- Harada, S.; Tominari, T.; Matsumoto, C.; Hirata, M.; Takita, M.; Inada, M.; Miyaura, C. Nobiletin, a polymethoxy flavonoid, suppresses bone resorption by inhibiting NF-κB-dependent prostaglandin E synthesis in osteoblasts and prevents bone loss due to estrogen deficiency. J. Pharmacol. Sci. 2011, 115, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.S.; Wang, Z.Y. Tea and cancer. J. Natl. Cancer Inst. 1993, 85, 1038–1049. [Google Scholar] [CrossRef] [PubMed]
- Haqqi, T.M.; Anthony, D.D.; Gupta, S.; Ahmad, N.; Lee, M.S.; Kumar, G.K.; Mukhtar, H. Prevention of collagen-induced arthritis in mice by a polyphenolic fraction from green tea. Proc. Natl. Acad. Sci. USA 1999, 96, 4524–4529. [Google Scholar] [CrossRef] [PubMed]
- Katiyar, S.K.; Matsui, M.S.; Elmets, C.A.; Mukhtar, H. Polyphenolic antioxidant (−)-Epigallocatechin-3-gallate from green tea reduces UVB-induced inflammatory responses and infiltration of leukocytes in human skin. Photochem. Photobiol. 1999, 69, 148–153. [Google Scholar] [CrossRef]
- Basu, A.; Lucas, E.A. Mechanisms and effects of green tea on cardiovascular health. Nutr. Rev. 2007, 65, 361–375. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.L.; Lane, J.; Coverly, J.; Stocks, J.; Jackson, S.; Stephen, A.; Bluck, L.; Coward, A.; Hendrickx, H. Effects of dietary supplementation with the green tea polyphenol epigallocatechin-3-gallate on insulin resistance and associated metabolic risk factors: randomized controlled trial. Br. J. Nutr. 2009, 101, 886–894. [Google Scholar] [CrossRef] [PubMed]
- McKay, D.L.; Blumberg, J.B. The role of tea in human health: An update. J. Am. Coll. Nutr. 2002, 21, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Leenen, R.; Roodenburg, A.J.; Tijburg, L.B.; Wiseman, S.A. A Single dose of tea with or without milk increases plasma antioxidant activity in humans. Eur. J. Clin. Nutr. 2000, 54, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.M.-Y.; Fong, D.; Ho, C.-T.; Huang, H.-I. Inhibition of inducible nitric oxide synthase gene expression and enzyme activity by epigallocatechin gallate, a natural product from green tea. Biochem. Pharmacol. 1997, 54, 1281–1286. [Google Scholar] [CrossRef]
- Yen, G.C.; Chen, H.Y. Scavenging effect of various tea extracts on superoxide derived from the metabolism of mutagens. Biosci. Biotechnol. Biochem. 1998, 62, 1768–1770. [Google Scholar] [CrossRef] [PubMed]
- Rice-Evans, C.A.; Miller, N.J.; Bolwell, P.G.; Bramley, P.M.; Pridham, J.B. The relative antioxidant activities of plant-derived polyphenolic flavonoids. Free Radic. Res. 1995, 22, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Balentine, D.A.; Wiseman, S.A.; Bouwens, L.C. The chemistry of tea flavonoids. Crit. Rev. Food Sci. Nutr. 1997, 37, 693–704. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.Y.; Park, J.O.; Phyu, H.; Dong, Z.; Yang, C.S. Mechanisms of inhibition of the Ras-MAP Kinase signaling pathway in 30.7b Ras 12 cells by tea polyphenols (−)-epigallocatechin-3-gallate and theaflavin-3,3ʹ-digallate. FASEB J. 2001, 15, 2022–2024. [Google Scholar] [PubMed]
- Chung, J.H.; Han, J.H.; Hwang, E.J.; Seo, J.Y.; Cho, K.H.; Kim, K.H.; Youn, J.I.; Eun, H.C. Dual mechanisms of green tea extract EGCG.-induced cell survival in human epidermal keratinocytes. FASEB J. 2003, 17, 1913–1915. [Google Scholar] [PubMed]
- Pianetti, S.; Guo, S.; Kavanagh, K.T.; Sonenshein, G.E. Green tea polyphenol epigallocatechin-3 gallate inhibits Her-2/neu signaling, proliferation, and transformed phenotype of breast cancer cells. Cancer Res. 2002, 62, 652–655. [Google Scholar] [PubMed]
- Ikigai, H.; Nakae, T.; Hara, Y.; Shimamura, T. Bactericidal catechins damage the lipid bilayer. Biochim. Biophys. Acta 1993, 1147, 132–136. [Google Scholar] [CrossRef]
- Ho, C.T.; Chen, Q.; Shi, H.; Zhang, K.Q.; Rosen, R.T. Antioxidative effect of polyphenol extract prepared from various Chinese teas. Prev. Med. 1992, 21, 520–525. [Google Scholar] [CrossRef]
- Fujiki, H.; Suganuma, M.; Imai, K.; Nakachi, K. Green tea: Cancer preventive beverage and/or drug. Cancer Lett. 2002, 188, 9–13. [Google Scholar] [CrossRef]
- Khan, N.; Mukhtar, H. Tea polyphenols for health promotion. Life Sci. 2007, 81, 519–533. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.P.; Schell, J.B.; Ho, C.-T.; Chen, K.Y. Green tea epigallocatechin gallate shows a pronounced growth inhibitory effect on cancerous cells but not on their normal counterparts. Cancer Lett. 1998, 129, 173–179. [Google Scholar] [CrossRef]
- Yang, C.S.; Maliakal, P.; Meng, X. Inhibition of carcinogenesis by tea. Annu. Rev. Pharmacol. Toxicol. 2002, 42, 25–54. [Google Scholar] [CrossRef] [PubMed]
- Bode, A.M.; Dong, Z. Targeting signal transduction pathways by chemopreventive agents. Mutat. Res. Mol. Mech. Mutagen. 2004, 555, 33–51. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, H.; Wachi, M.; Woo, J.-T.; Kato, M.; Kasai, S.; Takahashi, F.; Lee, I.-S.; Nagai, K. Fenton reaction is primarily involved in a mechanism of (−)-epigallocatechin-3-gallate to induce osteoclastic cell death. Biochem. Biophys. Res. Commun. 2002, 292, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.; Islam, N.; Kermode, T.; Johnstone, B.; Mukhtar, H.; Moskowitz, R.W.; Goldberg, V.M.; Malemud, C.J.; Haqqi, T.M. Involvement of caspase-3 in epigallocatechin-3-gallate-mediated apoptosis of human chondrosarcoma cells. Biochem. Biophys. Res. Commun. 2000, 270, 793–797. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, S.; Saeki, K.; Sazuka, M.; Suzuki, Y.; Shoji, Y.; Ohta, T.; Kaji, K.; Yuo, A.; Isemura, M. Apoptosis Induction by epigallocatechin gallate involves its binding to Fas. Biochem. Biophys. Res. Commun. 2001, 285, 1102–1106. [Google Scholar] [CrossRef] [PubMed]
- Oka, Y.; Iwai, S.; Amano, H.; Irie, Y.; Yatomi, K.; Ryu, K.; Yamada, S.; Inagaki, K.; Oguchi, K. Tea polyphenols inhibit rat osteoclast formation and differentiation. J. Pharmacol. Sci. 2012, 118, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Kamon, M.; Zhao, R.; Sakamoto, K. Green tea polyphenol (−)-epigallocatechin gallate suppressed the differentiation of murine osteoblastic MC3T3-E1 cells. Cell Biol. Int. 2010, 34, 109–116. [Google Scholar]
- Yun, J.-H.; Kim, C.-S.; Cho, K.-S.; Chai, J.-K.; Kim, C.-K.; Choi, S.-H. (−)-Epigallocatechin gallate induces apoptosis, via caspase activation, in osteoclasts differentiated from RAW 264.7 Cells. J. Periodontal Res. 2007, 42, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, T.; Tomofuji, T.; Endo, Y.; Irie, K.; Azuma, T.; Ekuni, D.; Tamaki, N.; Yamamoto, T.; Morita, M. Supplementation of green tea catechins in dentifrices suppresses gingival oxidative stress and periodontal inflammation. Arch. Oral Biol. 2011, 56, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.F.; Andrian, E.; Rowland, E.; Marquez, I.C. Immune response and alveolar bone resorption in a mouse model of treponema denticola infection. Infect. Immun. 2009, 77, 694–698. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, H.; Ukai, T.; Yoshimura, A.; Kozuka, Y.; Yoshioka, H.; Yoshinaga, Y.; Abe, Y.; Hara, Y. Green tea catechin inhibits lipopolysaccharide-induced bone resorption in vivo. J. Periodontal Res. 2010, 45, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Cho, A.-R.; Kim, J.-H.; Lee, D.-E.; Lee, J.-S.; Jung, U.-W.; Bak, E.-J.; Yoo, Y.-J.; Chung, W.-G.; Choi, S.-H. The effect of orally administered epigallocatechin-3-gallate on ligature-induced periodontitis in rats. J. Periodontal Res. 2013, 48, 781–789. [Google Scholar] [CrossRef] [PubMed]
- Kushiyama, M.; Shimazaki, Y.; Murakami, M.; Yamashita, Y. Relationship between intake of green tea and periodontal disease. J. Periodontol. 2009, 80, 372–377. [Google Scholar] [CrossRef] [PubMed]
- Hirasawa, M.; Takada, K.; Makimura, M.; Otake, S. Improvement of periodontal status by green tea catechin using a local delivery system: A clinical pilot study. J. Periodontal Res. 2002, 37, 433–438. [Google Scholar] [CrossRef] [PubMed]
- Makimura, M.; Hirasawa, M.; Kobayashi, K.; Indo, J.; Sakanaka, S.; Taguchi, T.; Otake, S. Inhibitory effect of tea catechins on collagenase activity. J. Periodontol. 1993, 64, 630–636. [Google Scholar] [CrossRef] [PubMed]
- Demeule, M.; Brossard, M.; Pagé, M.; Gingras, D.; Béliveau, R. Matrix metalloproteinase inhibition by green tea catechins. Biochim. Biophys. Acta 2000, 1478, 51–60. [Google Scholar] [CrossRef]
- Garbisa, S.; Biggin, S.; Cavallarin, N.; Sartor, L.; Benelli, R.; Albini, A. Tumor invasion: Molecular shears blunted by green tea. Nat. Med. 1999, 5, 1216–1216. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.-H.; Pang, E.-K.; Kim, C.-S.; Yoo, Y.-J.; Cho, K.-S.; Chai, J.-K.; Kim, C.-K.; Choi, S.-H. Inhibitory effects of green tea polyphenol (−)-epigallocatechin gallate on the expression of matrix metalloproteinase-9 and on the formation of osteoclasts. J. Periodontal Res. 2004, 39, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Maeda-Yamamoto, M.; Suzuki, N.; Sawai, Y.; Miyase, T.; Sano, M.; Hashimoto-Ohta, A.; Isemura, M. Association of suppression of extracellular signal-regulated kinase phosphorylation by epigallocatechin gallate with the reduction of matrix metalloproteinase activities in human fibrosarcoma HT1080 cells. J. Agric. Food Chem. 2003, 51, 1858–1863. [Google Scholar] [CrossRef] [PubMed]
- Isemura, M.; Saeki, K.; Minami, T.; Hayakawa, S.; Kimura, T.; Shoji, Y.; Sazuka, M. Inhibition of matrix metalloproteinases by tea catechins and related polyphenols. In Inhibition of Matrix Metalloproteinases: Therapeutic Applications; Greenwald, R.A., Zucker, S., Golub, L.M., Eds.; New York Academy Sciences: New York, NY, USA, 1999; Volume 878, pp. 629–631. [Google Scholar]
- Gillespie, K.; Kodani, I.; Dickinson, D.P.; Ogbureke, K.U.E.; Camba, A.M.; Wu, M.; Looney, S.; Chu, T.-C.; Qin, H.; Bisch, F.; et al. Effects of oral consumption of the green tea polyphenol EGCG in a murine model for human Sjogren’s syndrome, an autoimmune disease. Life Sci. 2008, 83, 581–588. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, Y.; Hosokawa, I.; Ozaki, K.; Nakanishi, T.; Nakae, H.; Matsuo, T. Catechins inhibit CXCL10 production from oncostatin M-stimulated human gingival fibroblasts. J. Nutr. Biochem. 2010, 21, 659–664. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, N.; Haqqi, T.M. Epigallocatechin-3-gallate suppresses the global interleukin-1beta-induced inflammatory response in human chondrocytes. Arthritis Res. Ther. 2011, 13, R93. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; de Villiers, W.J.; McClain, C.J.; Varilek, G.W. Green tea polyphenols block endotoxin-induced tumor necrosis factor-production and lethality in a murine model. J. Nutr. 1998, 128, 2334–2340. [Google Scholar] [PubMed]
- Crouvezier, S.; Powell, B.; Keir, D.; Yaqoob, P. The effects of phenolic components of tea on the production of pro- and anti-inflammatory cytokines by human leukocytes in vitro. Cytokine 2001, 13, 280–286. [Google Scholar] [CrossRef] [PubMed]
- Rogers, J.; Perkins, I.; van Olphen, A.; Burdash, N.; Klein, T.W.; Friedman, H. Epigallocatechin gallate modulates cytokine production by bone marrow-derived dendritic cells stimulated with lipopolysaccharide or muramyldipeptide, or infected with Legionella pneumophila. Exp. Biol. Med. Maywood 2005, 230, 645–651. [Google Scholar] [PubMed]
- Kojima-Yuasa, A.; Hua, J.J.; Kennedy, D.O.; Matsui-Yuasa, I. Green tea extract inhibits angiogenesis of human umbilical vein endothelial cells through reduction of expression of VEGF receptors. Life Sci. 2003, 73, 1299–1313. [Google Scholar] [CrossRef]
- Leung, L.K.; Su, Y.; Chen, R.; Zhang, Z.; Huang, Y.; Chen, Z.-Y. Theaflavins in black tea and catechins in green tea are equally effective antioxidants. J. Nutr. 2001, 131, 2248–2251. [Google Scholar] [PubMed]
- Hosokawa, Y.; Hosokawa, I.; Ozaki, K.; Nakanishi, T.; Nakae, H.; Matsuo, T. Tea polyphenols inhibit IL-6 production in tumor necrosis factor superfamily 14-stimulatedhuman gingival fibroblasts. Mol. Nutr. Food Res. 2010, 54, S151–S158. [Google Scholar] [CrossRef] [PubMed]
- Pace-Asciak, C.R.; Hahn, S.; Diamandis, E.P.; Soleas, G.; Goldberg, D.M. The red wine phenolics trans-resveratrol and quercetin block human platelet aggregation and eicosanoid synthesis: implications for protection against coronary heart disease. Clin. Chim. Acta Int. J. Clin. Chem. 1995, 235, 207–219. [Google Scholar] [CrossRef]
- Hayek, T.; Fuhrman, B.; Vaya, J.; Rosenblat, M.; Belinky, P.; Coleman, R.; Elis, A.; Aviram, M. Reduced progression of atherosclerosis in apolipoprotein E-deficient mice following consumption of red wine, or its polyphenols quercetin or catechin, is associated with reduced susceptibility of LDL to oxidation and aggregation. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 2744–2752. [Google Scholar] [CrossRef] [PubMed]
- Maffei Facinó, R.; Carini, M.; Aldini, G.; Berti, F.; Rossoni, G.; Bombardelli, E.; Morazzoni, P. Procyanidines from Vitis vinifera seeds protect rabbit heart from ischemia/reperfusion injury: Antioxidant intervention and/or iron and copper sequestering ability. Planta Med. 1996, 62, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Wang, J.; Chen, Y.; Agarwal, R. Anti-tumor-promoting activity of a polyphenolic fraction isolated from grape seeds in the mouse skin two-stage initiation–promotion protocol and identification of procyanidin B5-3′-gallate as the most effective antioxidant constituent. Carcinogenesis 1999, 20, 1737–1745. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.P.; Lee, J.Y.; Jeong, J.K.; Bae, S.W.; Lee, H.K.; Jo, I. Nongenomic stimulation of nitric oxide release by estrogen is mediated by estrogen receptor alpha localized in caveolae. Biochem. Biophys. Res. Commun. 1999, 263, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Bagchi, D.; Garg, A.; Krohn, R.L.; Bagchi, M.; Tran, M.X.; Stohs, S.J. Oxygen free radical scavenging abilities of vitamins C and E, and a grape seed proanthocyanidin extract in vitro. Res. Commun. Mol. Pathol. Pharmacol. 1997, 95, 179–189. [Google Scholar] [PubMed]
- Bodet, C.; Chandad, F.; Grenier, D. Anti-inflammatory activity of a high-molecular-weight cranberry fraction on macrophages stimulated by lipopolysaccharides from periodontopathogens. J. Dent. Res. 2006, 85, 235–239. [Google Scholar] [CrossRef] [PubMed]
- Bodet, C.; Chandad, F.; Grenier, D. Cranberry components inhibit interleukin-6, interleukin-8, and prostaglandin E-2 production by lipopolysaccharide-activated gingival fibroblasts. Eur. J. Oral Sci. 2007, 115, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Bodet, C.; Chandad, F.; Grenier, D. Inhibition of host extracellular matrix destructive enzyme production and activity by a high-molecular-weight cranberry fraction. J. Periodontal Res. 2007, 42, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Labrecque, J.; Bodet, C.; Chandad, F.; Grenier, D. Effects of a high-molecular-weight cranberry fraction on growth, biofilm formation and adherence of Porphyromonas gingivalis. J. Antimicrob. Chemother. 2006, 58, 439–443. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Varela-López, A.; Bullón, P.; Giampieri, F.; Quiles, J.L. Non-Nutrient, Naturally Occurring Phenolic Compounds with Antioxidant Activity for the Prevention and Treatment of Periodontal Diseases. Antioxidants 2015, 4, 447-481. https://doi.org/10.3390/antiox4030447
Varela-López A, Bullón P, Giampieri F, Quiles JL. Non-Nutrient, Naturally Occurring Phenolic Compounds with Antioxidant Activity for the Prevention and Treatment of Periodontal Diseases. Antioxidants. 2015; 4(3):447-481. https://doi.org/10.3390/antiox4030447
Chicago/Turabian StyleVarela-López, Alfonso, Pedro Bullón, Francesca Giampieri, and José L. Quiles. 2015. "Non-Nutrient, Naturally Occurring Phenolic Compounds with Antioxidant Activity for the Prevention and Treatment of Periodontal Diseases" Antioxidants 4, no. 3: 447-481. https://doi.org/10.3390/antiox4030447