
Equine PBMC Cytokines Profile after In Vitro α- and γ-EHV Infection: Efficacy of a Parapoxvirus Ovis Based-Immunomodulator Treatment

 , , ,
, , , 
Abstract
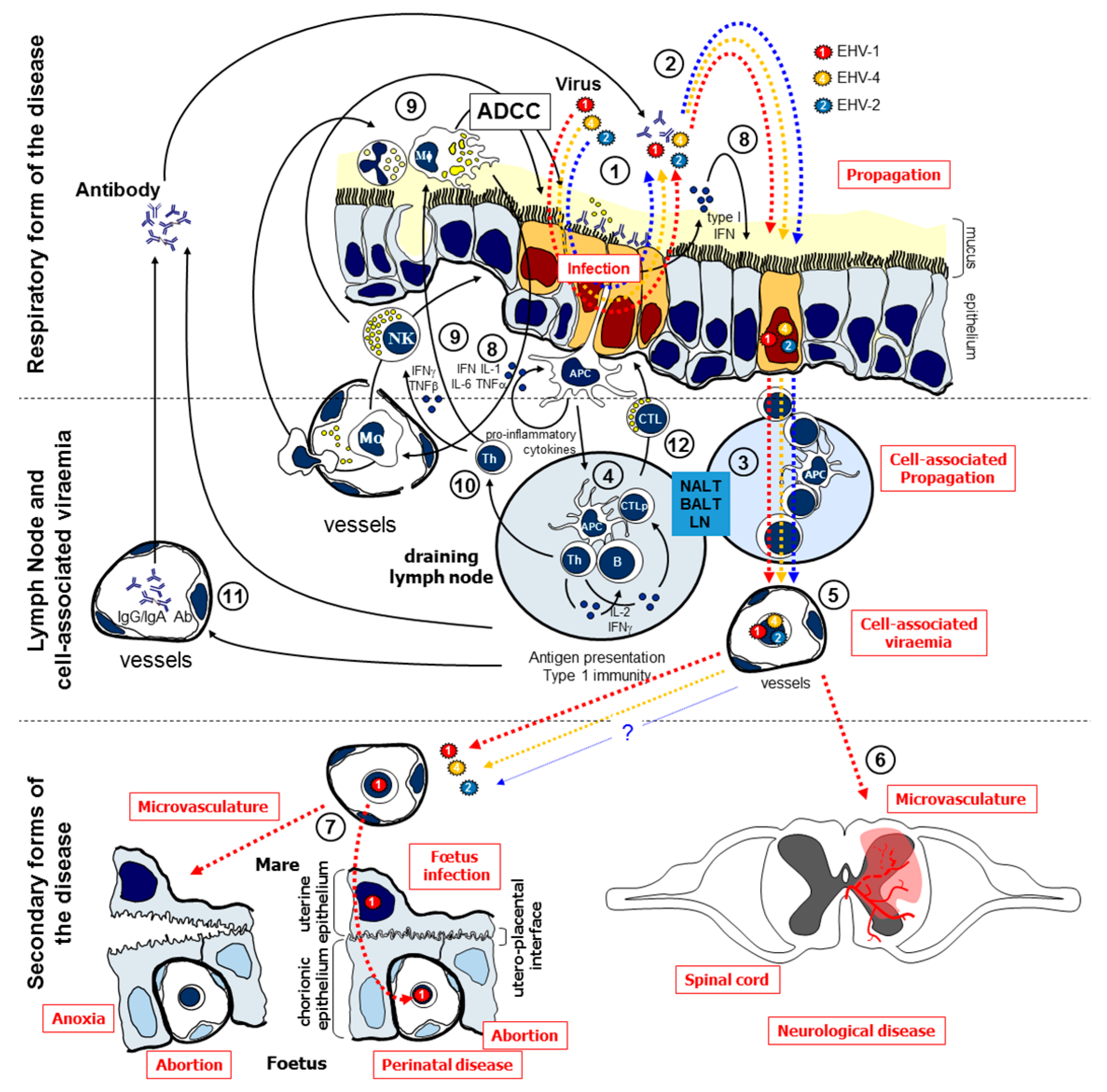
:1. Introduction
2. Materials and Methods
2.1. Virus Strains and Immunomodulator
2.2. Horses
2.3. Culture of Equine PBMC
2.4. mRNA Isolation and Reverse Transcription
2.5. Cytokines’ Gene Expression Analysis
2.6. Quantitation of EHV by qPCR
2.7. Statistical Analyses
3. Results
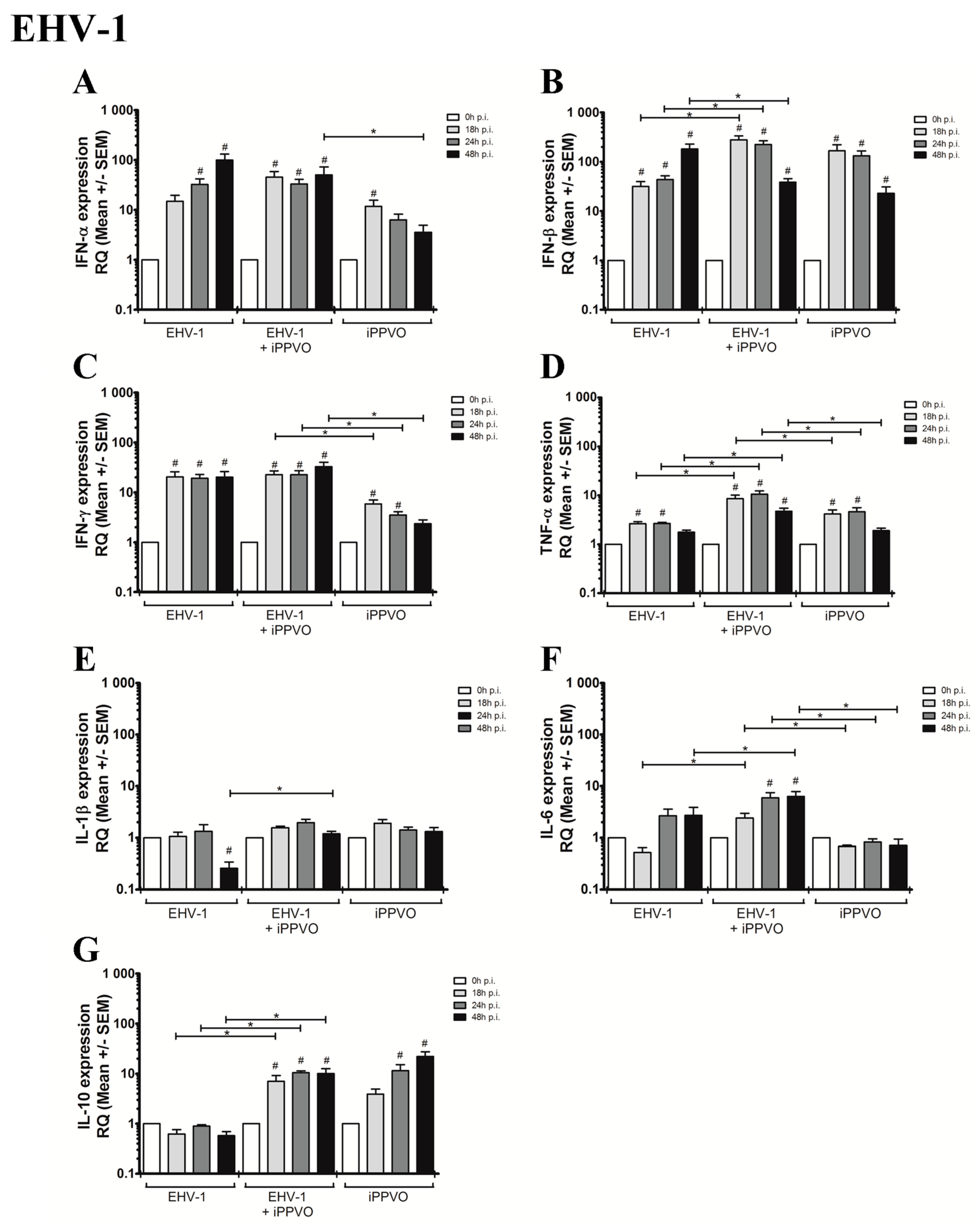
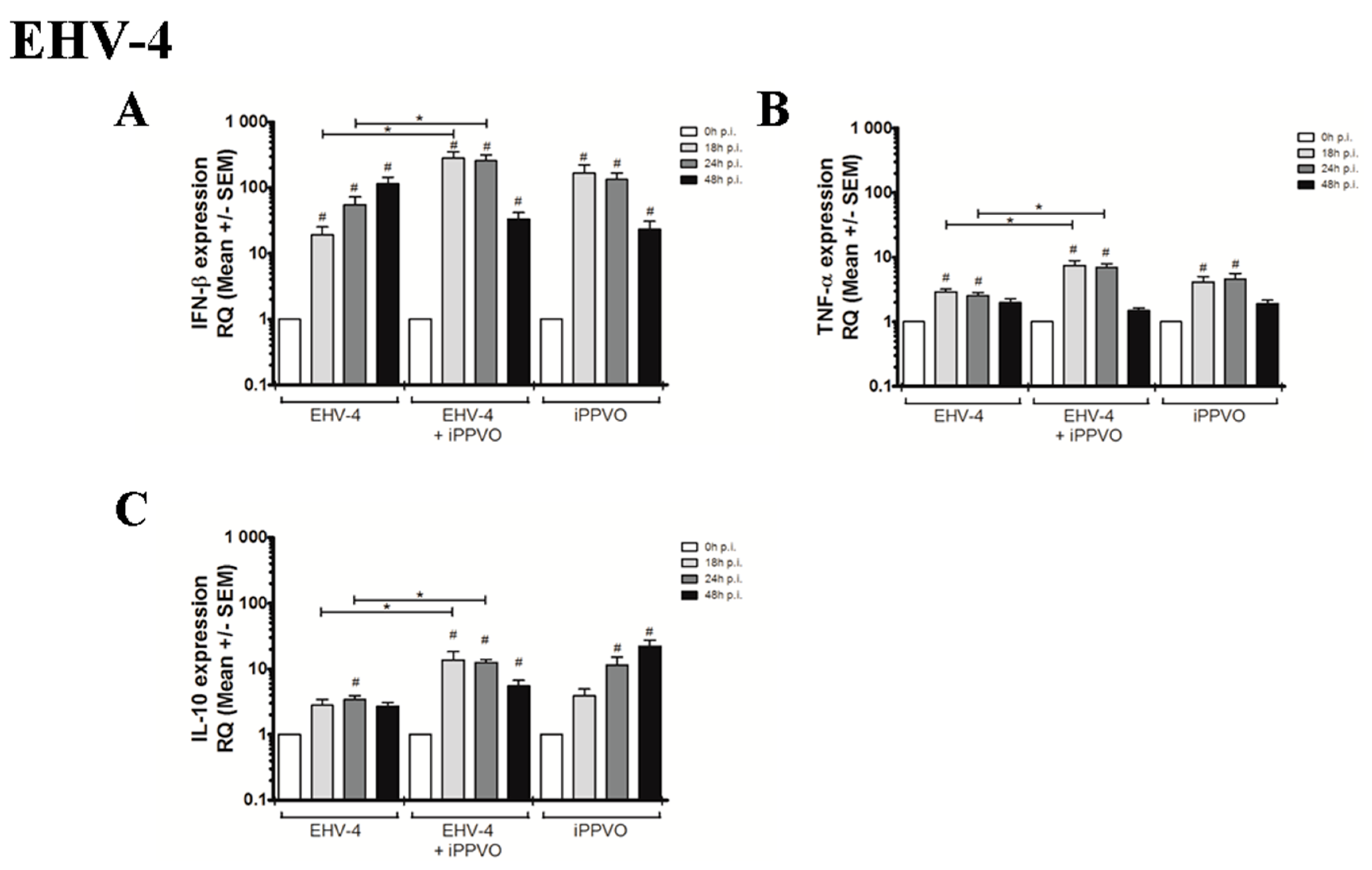
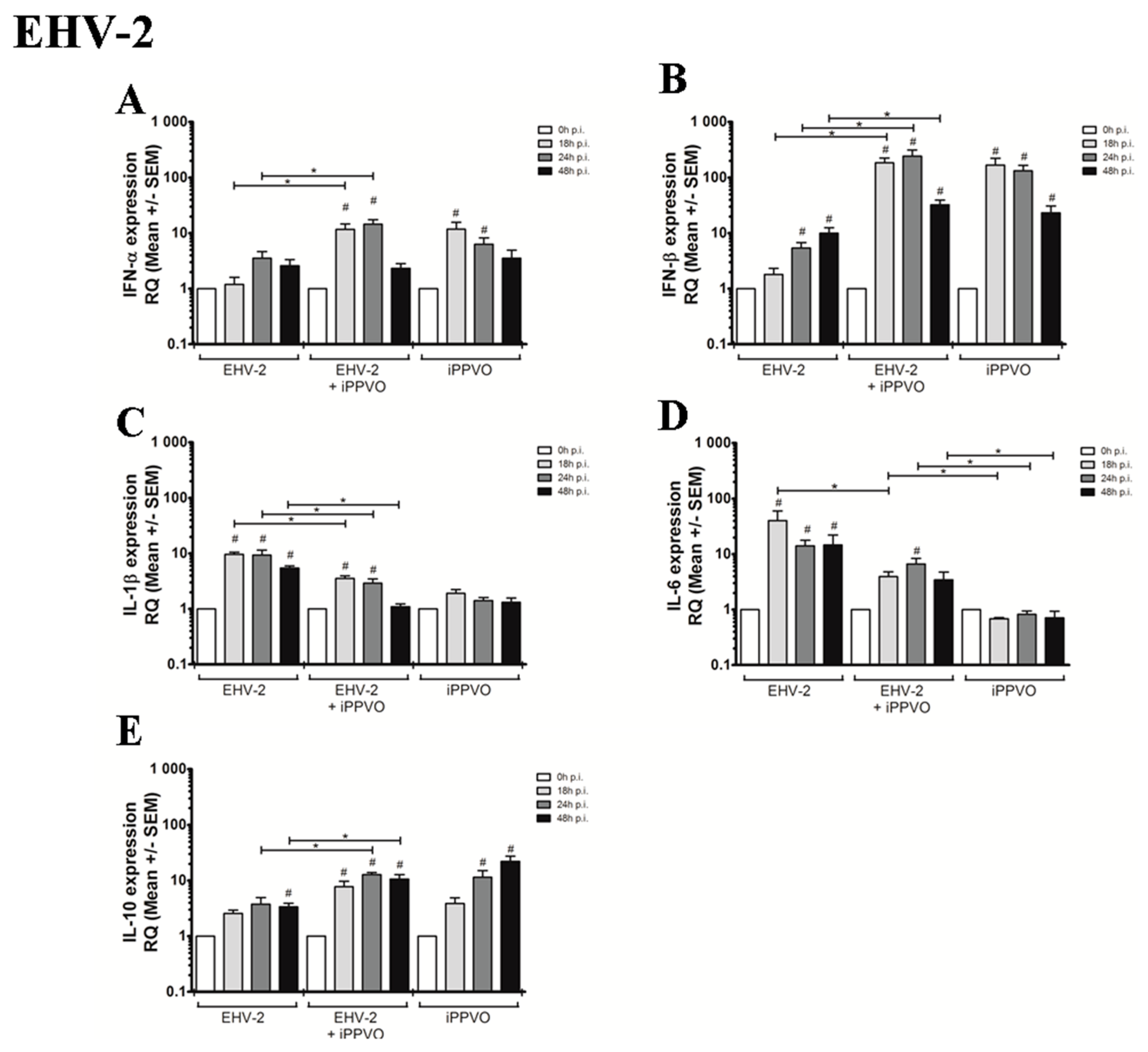
3.1. Cytokines Profiles Following PBMC Infection with EHVs
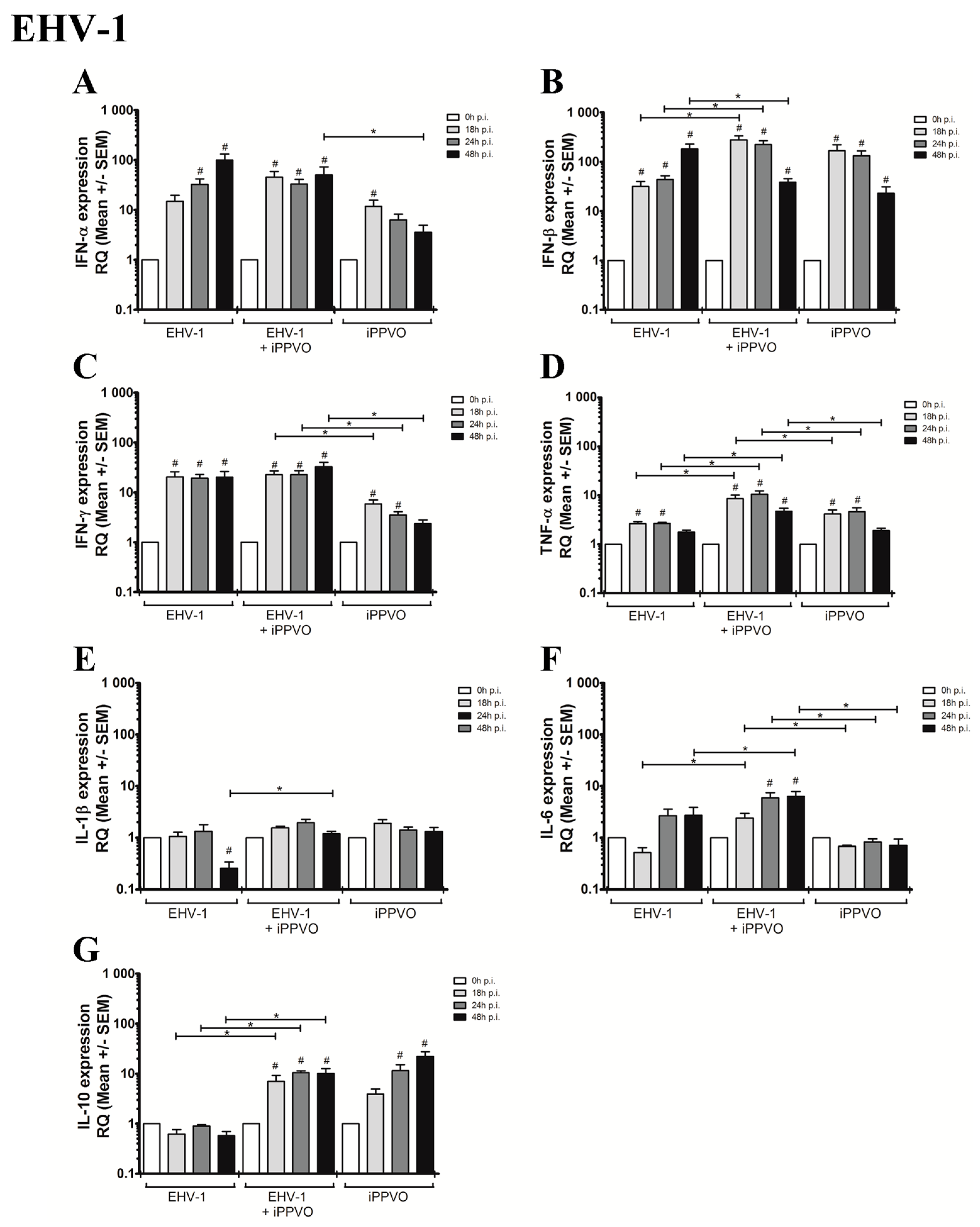
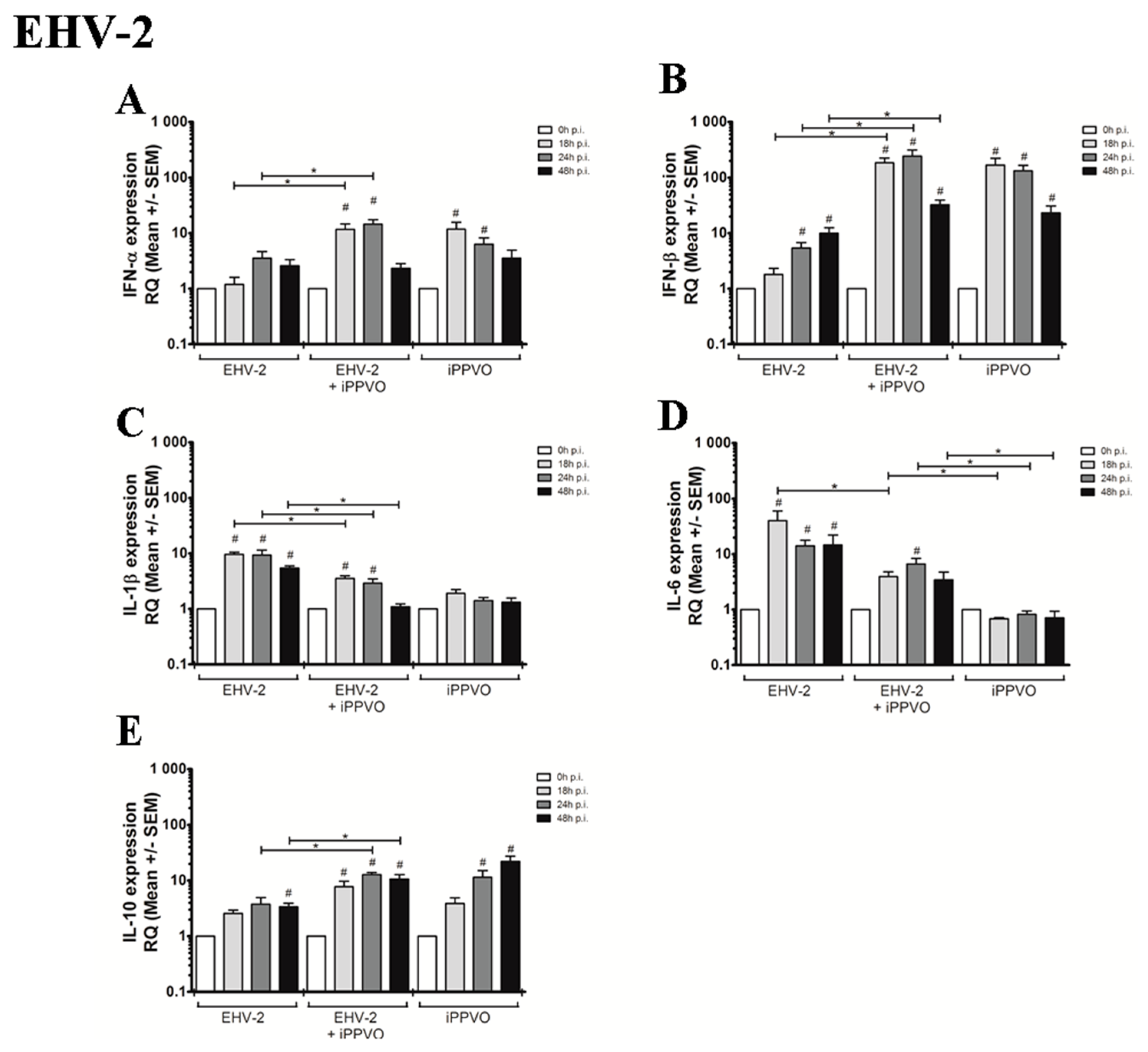
3.2. Cytokines Profiles Following EHV Infection and iPPVO Co-Treatment of PBMC
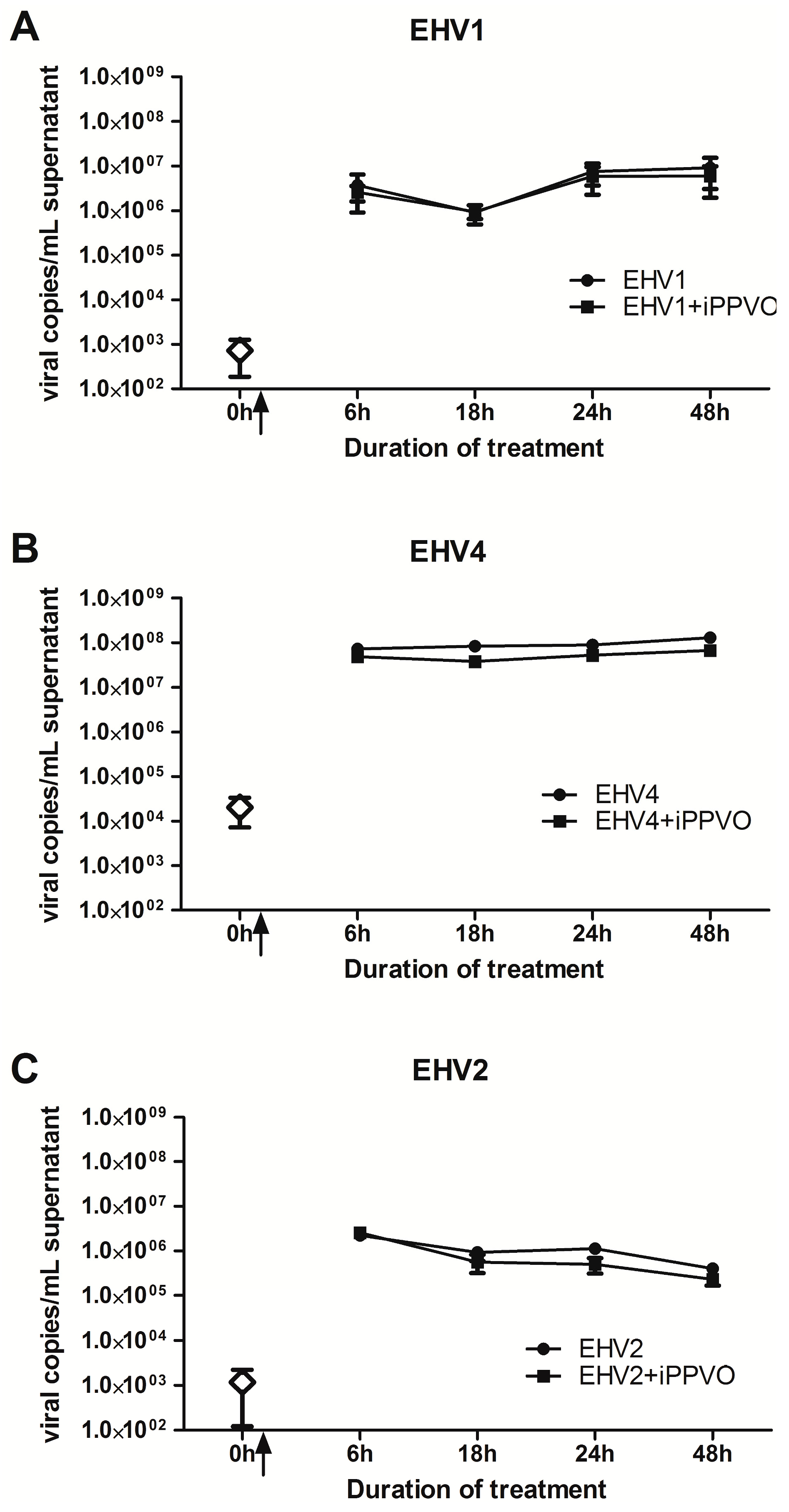
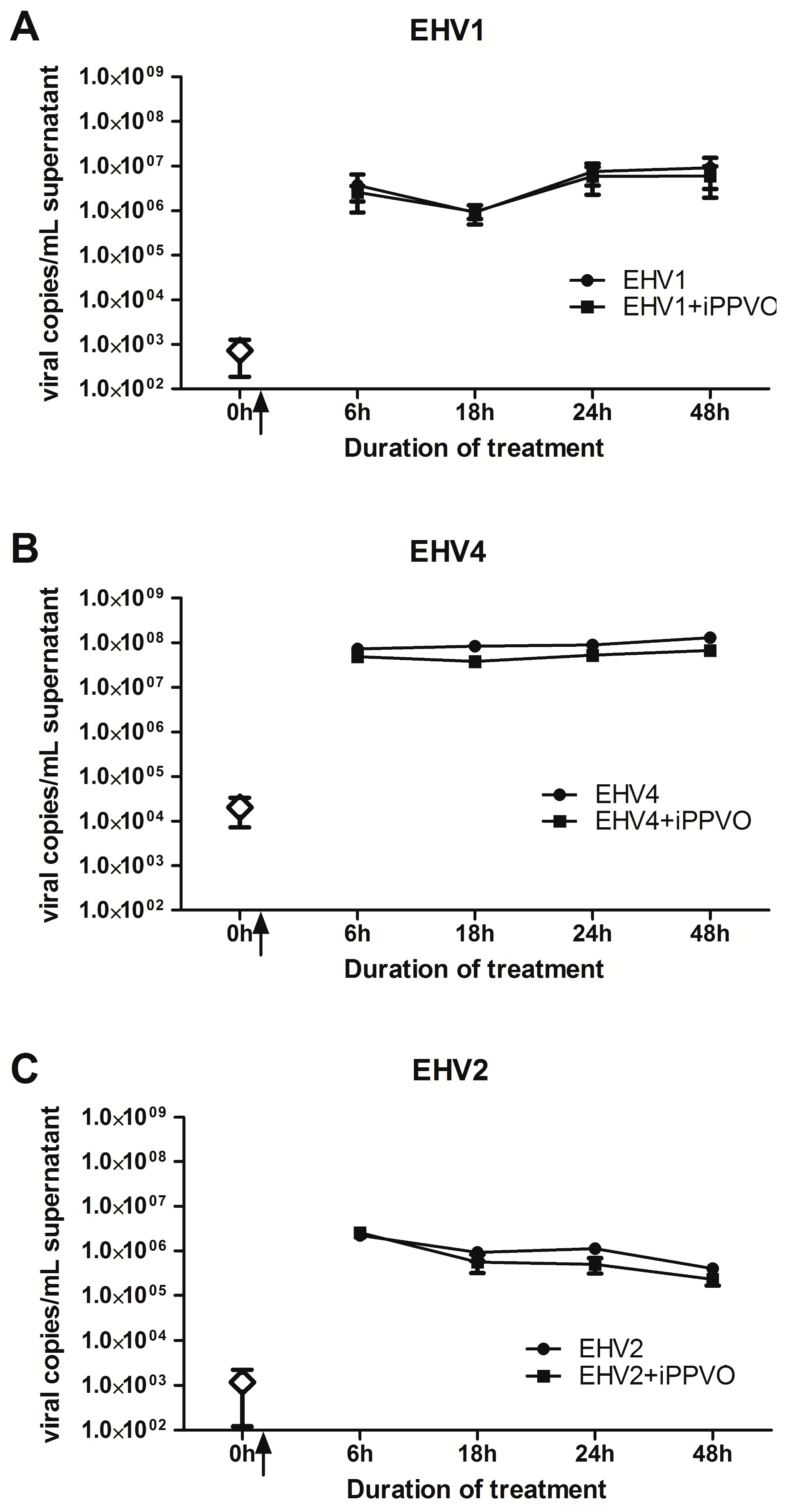
3.3. Growth Kinetics of EHV +/− iPPVO in PBMC
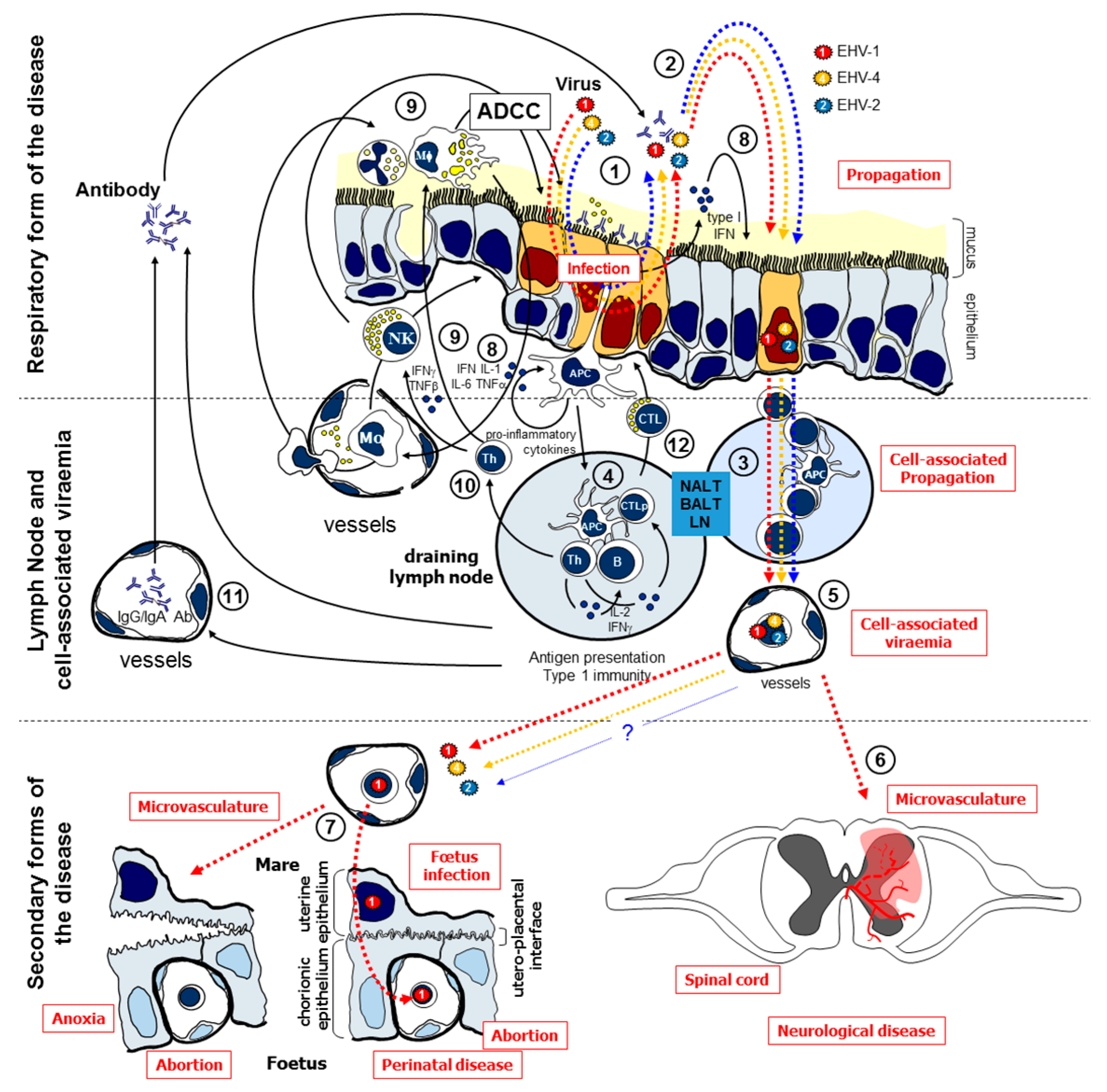
4. Discussion
4.1. EHV Replication and Modulation of Cytokine Responses Following PBMC Infection
4.2. Efficacy of iPPVO against EHV Infections in Equine PBMC
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Davison, A.J.; Eberle, R.; Ehlers, B.; Hayward, G.S.; McGeoch, D.J.; Minson, A.C.; Pellett, P.E.; Roizman, B.; Studdert, M.J.; Thiry, E. The order Herpesvirales. Arch. Virol. 2009, 154, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Allen, G.P. Development of a real-time polymerase chain reaction assay for rapid diagnosis of neuropathogenic strains of equine herpesvirus-1. J. Vet. Diagn. Investig. 2007, 19, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Pusterla, N.; Wilson, W.D.; Mapes, S.; Finno, C.; Isbell, D.; Arthur, R.M.; Ferraro, G.L. Characterization of viral loads, strain and state of equine herpesvirus-1 using real-time PCR in horses following natural exposure at a racetrack in California. Vet. J. 2009, 179, 230–239. [Google Scholar] [CrossRef] [PubMed]
- Fortier, G.; van Erck, E.; Pronost, S.; Lekeux, P.; Thiry, E. Equine gammaherpesviruses: Pathogenesis, epidemiology and diagnosis. Vet. J. 2010, 186, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Slater, J.D.; Borchers, K.; Thackray, A.M.; Field, H.J. The trigeminal ganglion is a location for equine herpesvirus 1 latency and reactivation in the horse. J. Gen. Virol. 1994, 75, 2007–2016. [Google Scholar] [CrossRef] [PubMed]
- Bell, S.A.; Balasuriya, U.B.R.; Gardner, I.A.; Barry, P.A.; Wilson, W.D.; Ferraro, G.L.; MacLachlan, N.J. Temporal detection of equine herpesvirus infections of a cohort of mares and their foals. Vet. Microbiol. 2006, 116, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Van der Meulen, K.M.; Nauwynck, H.J.; Buddaert, W.; Pensaert, M.B. Replication of equine herpesvirus type 1 in freshly isolated equine peripheral blood mononuclear cells and changes in susceptibility following mitogen stimulation. J. Gen. Virol. 2000, 81, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Breathnach, C.C.; Soboll, G.; Suresh, M.; Lunn, D.P. Equine herpesvirus-1 infection induces IFN-γ production by equine T lymphocyte subsets. Vet. Immunol. Immunopathol. 2005, 103, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Taktaz Hafshejani, T.; Nekoei, S.; Vazirian, B.; Doosti, A.; Khamesipour, F.; Anyanwu, M.U. Molecular Detection of Equine Herpesvirus Types 1 and 4 Infection in Healthy Horses in Isfahan Central and Shahrekord Southwest Regions, Iran. BioMed Res. Int. 2015, 2015, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Fortier, G.; Richard, E.; Hue, E.; Fortier, C.; Pronost, S.; Pottier, D.; Lemaitre, L.; Lekeux, P.; Borchers, K.; Thiry, E. Long-lasting airway inflammation associated with equid herpesvirus-2 in experimentally challenged horses. Vet. J. 2013, 197, 492–495. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.M.; Zhang, Y.; Jennings, S.R.; O’Callaghan, D.J. Characterization of the cytolytic T-lymphocyte response to a candidate vaccine strain of equine herpesvirus 1 in CBA mice. J. Virol. 1998, 72, 5366–5372. [Google Scholar] [PubMed]
- Paillot, R.; Case, R.; Ross, J.; Newton, R.; Nugent, J. Equine herpes virus-1: Virus, immunity and vaccines. Open Vet. Sci. J. 2008, 2, 68–91. [Google Scholar] [CrossRef]
- Lunn, D.P.; Davis-Poynter, N.; Flaminio, M.J.B.F.; Horohov, D.W.; Osterrieder, K.; Pusterla, N.; Townsend, H.G.G. Equine Herpesvirus-1 Consensus Statement. J. Vet. Intern. Med. 2009, 23, 450–461. [Google Scholar] [CrossRef] [PubMed]
- Garré, B.; Gryspeerdt, A.; Croubels, S.; De Backer, P.; Nauwynck, H. Evaluation of orally administered valacyclovir in experimentally EHV1-infected ponies. Vet. Microbiol. 2009, 135, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Paillot, R.; Daly, J.M.; Luce, R.; Montesso, F.; Davis-Poynter, N.; Hannant, D.; Kydd, J.H. Frequency and phenotype of EHV-1 specific, IFN-γ synthesising lymphocytes in ponies: The effects of age, pregnancy and infection. Dev. Comp. Immunol. 2007, 31, 202–214. [Google Scholar] [CrossRef] [PubMed]
- Soboll Hussey, G.; Ashton, L.V.; Quintana, A.M.; Lunn, D.P.; Goehring, L.S.; Annis, K.; Landolt, G. Innate immune responses of airway epithelial cells to infection with Equine herpesvirus-1. Vet. Microbiol. 2014, 170, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Svansson, V.; Roelse, M.; Ólafsdóttir, G.; Thorsteinsdóttir, L.; Torfason, E.G.; Torsteinsdóttir, S. Immune response against equine gammaherpesvirus in Icelandic horses. Vet. Microbiol. 2009, 137, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Yan, N.; Chen, Z.J. Intrinsic antiviral immunity. Nat. Immunol. 2012, 13, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Paillot, R. A systematic review of the immune-modulators Parapoxvirus ovis and Propionibacterium acnes for the prevention of respiratory disease and other infections in the horse. Vet. Immunol. Immunopathol. 2013, 153, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ziebell, K.-L.; Steinmann, H.; Kretzdorn, D.; Schlapp, T.; Failing, K.; Schmeer, N. The use of Baypamun N in crowding associated infectious respiratory disease: Efficacy of Baypamun N (freeze dried product) in 4–10 month old horses. J. Vet. Med. Ser. B 1997, 44, 529–536. [Google Scholar] [CrossRef]
- Horohov, D.W.; Breathnach, C.C.; Sturgill, T.L.; Rashid, C.; Stiltner, J.L.; Strong, D.; Nieman, N.; Holland, R.E. In vitro and in vivo modulation of the equine immune response by parapoxvirus ovis. Equine Vet. J. 2008, 40, 468–472. [Google Scholar] [CrossRef] [PubMed]
- Weber, O.; Siegling, A.; Friebe, A.; Limmer, A.; Schlapp, T.; Knolle, P.; Mercer, A.; Schaller, H.; Volk, H.-D. Inactivated parapoxvirus ovis (Orf virus) has antiviral activity against hepatitis B virus and herpes simplex virus. J. Gen. Virol. 2003, 84, 1843–1852. [Google Scholar] [CrossRef] [PubMed]
- Donecker, J.M.; Holland, R.E., Jr. Efficacy of the Immunomodulator Zylexis in Horses Challenged with Equine Herpesvirus under Field Conditions. Available online: https://www.zoetisus.com/_locale-assets/mcm-portal-assets/widgets/le/advantage/pdf/equine/eqb06020-zylexis.pdf (accessed on 29 October 2015).
- Ons, E.; Van Brussel, L.; Lane, S.; King, V.; Cullinane, A.; Kenna, R.; Lyons, P.; Hammond, T.-A.; Salt, J.; Raue, R. Efficacy of a Parapoxvirus ovis-based immunomodulator against equine herpesvirus type 1 and Streptococcus equi equi infections in horses. Vet. Microbiol. 2014, 173, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Coombs, D.K.; Patton, T.; Kohler, A.K.; Soboll, G.; Breathnach, C.; Townsend, H.G.G.; Lunn, D.P. Cytokine responses to EHV-1 infection in immune and non-immune ponies. Vet. Immunol. Immunopathol. 2006, 111, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Colahan, P.T.; Kollias-Baker, C.; Leutenegger, C.M.; Jones, J.H. Does training affect mRNA transciption for cytokine production in circulating leucocytes? Equine Vet. J. 2002, 34, 154–158. [Google Scholar] [CrossRef] [PubMed]
- Saulez, M.N.; Godfroid, J.; Bosman, A.; Stiltner, J.L.; Breathnach, C.C.; Horohov, D.W. Cytokine mRNA expressions after racing at a high altitude and at sea level in horses with exercise-induced pulmonary hemorrhage. Am. J. Vet. Res. 2010, 71, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Diallo, I.S.; Hewitson, G.; Wright, L.; Rodwell, B.J.; Corney, B.G. Detection of equine herpesvirus type 1 using a real-time polymerase chain reaction. J. Virol. Methods 2006, 131, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Diallo, I.S.; Hewitson, G.; Wright, L.L.; Kelly, M.A.; Rodwell, B.J.; Corney, B.G. Multiplex real-time PCR for the detection and differentiation of equid herpesvirus 1 (EHV-1) and equid herpesvirus 4 (EHV-4). Vet. Microbiol. 2007, 123, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Hue, E.S.; Fortier, C.I.; Laurent, A.M.; Quesnelle, Y.F.; Fortier, G.D.; Legrand, L.J.; Pronost, S.L. Development and Validation of a Quantitative PCR Method for Equid Herpesvirus-2 Diagnostics in Respiratory Fluids. J. Vis. Exp. JoVE 2016. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Soboll Hussey, G.; Ashton, L.V.; Quintana, A.M.; Van de Walle, G.R.; Osterrieder, N.; Lunn, D.P. Equine herpesvirus type 1 pUL56 modulates innate responses of airway epithelial cells. Virology 2014, 464–465, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Paillot, R.; Daly, J.M.; Juillard, V.; Minke, J.M.; Hannant, D.; Kydd, J.H. Equine interferon gamma synthesis in lymphocytes after in vivo infection and in vitro stimulation with EHV-1. Vaccine 2005, 23, 4541–4551. [Google Scholar] [CrossRef] [PubMed]
- Wagner, B.; Wimer, C.; Freer, H.; Osterrieder, N.; Erb, H.N. Infection of peripheral blood mononuclear cells with neuropathogenic equine herpesvirus type-1 strain Ab4 reveals intact interferon-α induction and induces suppression of anti-inflammatory interleukin-10 responses in comparison to other viral strains. Vet. Immunol. Immunopathol. 2011, 143, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Balasuriya, U.B.R.; Horohov, D.W.; Chambers, T.M. Equine herpesvirus-1 suppresses type-I interferon induction in equine endothelial cells. Vet. Immunol. Immunopathol. 2016, 1–9. [Google Scholar] [CrossRef] [PubMed]
- McNab, F.; Mayer-Barber, K.; Sher, A.; Wack, A.; O’Garra, A. Type I interferons in infectious disease. Nat. Rev. Immunol. 2015, 15, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Redpath, S.; Angulo, A.; Gascoigne, N.R.; Ghazal, P. Murine cytomegalovirus infection down-regulates MHC class II expression on macrophages by induction of IL-10. J. Immunol. 1999, 162, 6701–6707. [Google Scholar] [PubMed]
- Salek-Ardakani, S.; Stuart, A.D.; Arrand, J.E.; Lyons, S.; Arrand, J.R.; Mackett, M. High level expression and purification of the Epstein-Barr virus encoded cytokine viral interleukin 10: Efficient removal of endotoxin. Cytokine 2002, 17, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Friebe, A.; Siegling, A.; Friederichs, S.; Volk, H.-D.; Weber, O. Immunomodulatory Effects of Inactivated Parapoxvirus Ovis (Orf Virus) on Human Peripheral Immune Cells: Induction of Cytokine Secretion in Monocytes and Th1-Like Cells. J. Virol. 2004, 78, 9400–9411. [Google Scholar] [CrossRef] [PubMed]
- Ryan, C.; Giguère, S.; Fultz, L.; Long, M.T.; Crawford, P.C. Effects of two commercially available immunostimulants on leukocyte function of foals following ex vivo exposure to Rhodococcus equi. Vet. Immunol. Immunopathol. 2010, 138, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Schütze, N.; Raue, R.; Büttner, M.; Alber, G. Inactivated parapoxvirus ovis activates canine blood phagocytes and T lymphocytes. Vet. Microbiol. 2009, 137, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Anziliero, D.; Weiblen, R.; Kreutz, L.C.; Spilki, F.; Flores, E.F. Inactivated Parapoxvirus ovis induces a transient increase in the expression of proinflammatory, Th1-related, and autoregulatory cytokines in mice. Braz. J. Med. Biol. Res. 2014. [Google Scholar] [CrossRef] [PubMed]
- Fachinger, V.; Schlapp, T.; Strube, W.; Schmeer, N.; Saalmüller, A. Poxvirus-induced immunostimulating effects on porcine leukocytes. J. Virol. 2000, 74, 7943–7951. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Oligo | Sequences (5′–3′) | PCR Product Size (bp) | Efficiency (%) | Sequence Accession Number | Reference |
|---|---|---|---|---|---|---|
| ACTB | Forward | AGCGAAATCGTGCGTGACA | 70 | 81 | NM_001081838 AF035774 | [25] |
| Reverse | GCCATCTCCTGCTCGAAGT | |||||
| Probe | VIC-CAAGGAGAAGCTCTGCTATGTCGCCCT-MGB | |||||
| GAPDH | Forward | AAGTGGATATTGTCGCCATCAAT | 88 | 90 | NM_001163856 AF157626 AF035774 AF097179 | [26] |
| Reverse | AACTTGCCATGGGTGGAATC | |||||
| Probe | VIC-ACTACATGGTCTACATGTTTCAGTA-MGB | |||||
| IFN-α | Forward | CCTTACTGATGGCCCTGGTG | 124 | 87 | NM_001099441; EU682378 | adapted from [21] |
| Reverse | ATTCTCCTCATTTGTCCCAGGA | |||||
| Probe | FAM-CCTGCCTCACACCCATAG-MGB | |||||
| IFN-β | Forward | ACACCTGGCGTATTTTCAGAAGA | 74 | 93 | NM_001099440 | [21] |
| Reverse | CACAAGGAGGTTCTTAACGATGGT | |||||
| Probe | FAM-CTAGCACTGGCTGGAATGA-MGB | |||||
| IFN-γ | Forward | AGCAGCACCAGCAAGCT | 72 | 100 | U04050; D28520; NM_001081949 | [21] |
| Reverse | TTTGCGCTGGACCTTCAGA | |||||
| Probe | FAM-ATTCAGATTCCGGTAAATGA-MGB | |||||
| TNF-α | Forward | TTACCGAATGCCTTCCAGTCAAT | 85 | 89 | M64087 NM_001081819 | [21] |
| Reverse | GGGCTACAGGCTTGTCACTT | |||||
| Probe | FAM-CCAGACACTCAGATCAT-MGB | |||||
| IL-1β | Forward | CCGACACCAGTGACATGATGA | 64 | 89 | NM_001082526; D42147; U92481; D42165 | adapted from [27] |
| Reverse | TCCTCCTCAAAGAACAGGTCATTC | |||||
| Probe | FAM-CTTACTGCAGCGGCAAT-MGB | |||||
| IL-6 | Forward | GGATGCTTCCAATCTGGGTTCAAT | 65 | 86 | NM_001082496; AF005227; U64794; EU438770 | [27] |
| Reverse | TCCGAAAGACCAGTGGTGATTTT | |||||
| Probe | FAM-ATCAGGCAGGTCTCCTG-MGB | |||||
| IL-10 | Forward | GACATCAAGGAGCACGTGAACTC | 113 | 100 | U38200; EU438771; NM_001082490 | [26] |
| Reverse | TGCTCCACTGCCTTGCTCTT | |||||
| Probe | FAM-TGCGGCGCTGTCATCGATTTCTG-MGB | |||||
| EHV-1 gB | Forward | CATGTCAACGCACTCCCA | 63 | 93 | AY665713.1 | adapted from [28] |
| Reverse | GGGTCGGGCGTTTCTGT | |||||
| Probe | FAM-CCCTACGCTGCTCC-MGB | |||||
| EHV-4 gB | Forward | GGGCTATTGGATTACAGCGAGAT | 86 | 84 | NC_001844.1 | adapted from [29] |
| Reverse | TAGAATCGGAGGGCGTGAAG | |||||
| Probe | VIC-CAGCGCCGTAACCAG-MGB | |||||
| EHV-2 gB | Forward | GTGGCCAGCGGGGTGTTC | 78 | 97 | NC_001650 | [30] |
| Reverse | CCCCCAAAGGGATTYTTGAA | |||||
| Probe | FAM-CCCTCTTTGGGAGCATAGTCTCGGGG-TAMRA |
| Response | Cytokine | EHV-1 | EHV-4 | EHV-2 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 h | 18 h | 24 h | 48 h | 0 h | 18 h | 24 h | 48 h | 0 h | 18 h | 24 h | 48 h | ||
| IFN response | IFN-α | 1 | 14,9 (+/− 4,7) | 32,5 (+/− 9,5) | 99,7 (+/− 31,8) | 1 | 5,4 (+/− 1,9) | 9,8 (+/− 2,9) | 8,6 (+/− 3,4) | 1 | 1,2 (+/− 0,4) | 3,5 (+/− 1,1) | 2,6 (+/− 0,8) |
| IFN-β | 1 | 31,8 (+/− 8,1) | 43,7 (+/− 8,5) | 182,9 (+/− 45,4) | 1 | 19,1 (+/− 6,2) | 54,8 (+/− 17,5) | 115,7 (+/− 27,1) | 1 | 1,8 (+/− 0,5) | 5,4 (+/− 1,4) | 10,0 (+/− 2,6) | |
| IFN-γ | 1 | 20,4 (+/− 5,4) | 19,3 (+/− 3,5) | 20,1 (+/− 6,0) | 1 | 22,0 (+/− 4,3) | 25,2 (+/− 4,7) | 40,3 (+/− 8,3) | 1 | 25,0 (+/− 5,1) | 25,9 (+/− 6,5) | 18,2 (+/− 4,4) | |
| Inflammatory response | TNF-α | 1 | 2,6 (+/− 0,3) | 2,6 (+/− 0,2) | 1,8 (+/− 0,2) | 1 | 2,9 (+/− 0,3) | 2,5 (+/− 0,3) | 2,0 (+/− 0,3) | 1 | 3,7 (+/− 0,5) | 3,0 (+/−0,3) | 2,5 (+/− 0,3) |
| IL-1β | 1 | 1,1 (+/− 0,2) | 1,3 (+/− 0,5) | 0,3 (+/− 0,1) | 1 | 2,6 (+/− 0,5) | 2,1 (+/− 0,6) | 1,5 (+/− 0,3) | 1 | 9,6 (+/− 0,8) | 9,4 (+/− 2,1) | 5,4 (+/− 0,5) | |
| IL-6 | 1 | 0,5 (+/− 0,1) | 2,7 (+/− 0,9) | 2,7 (+/− 1,1) | 1 | 2,9 (+/− 0,6) | 5,7 (+/− 2,0) | 2,9 (+/− 1,3) | 1 | 39,9 (+/− 19,8) | 14,1 (+/− 3,7) | 14,7 (+/− 7,5) | |
| Treg response | IL-10 | 1 | 0,6 (+/− 0,1) | 0,9 (+/− 0,1) | 0,6 (+/− 0,1) | 1 | 2,8 (+/− 0,6) | 3,4 (+/− 0,5) | 2,7 (+/− 0,4) | 1 | 2,6 (+/− 0,4) | 3,8 (+/− 1,2) | 3,4 (+/− 0,5) |
| Response | Cytokine | iPPVO | EHV + iPPVO | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EHV-1 + iPPVO | EHV-4 + iPPVO | EHV-2 + iPPVO | |||||||||||||||
| 0 h | 18 h | 24 h | 48 h | 0 h | 18 h | 24 h | 48 h | 0 h | 18 h | 24 h | 48 h | 0 h | 18 h | 24 h | 48 h | ||
| IFN response | IFN-α | 1 | 11,8 (+/− 3,8) | 6,3 (+/− 1,9) | 3,6 (+/− 1,4) | 1 | 45,5 (+/− 13,4) | 33,2 (+/− 7,8) | 50,4 (+/− 22,7) | 1 | 27,4 (+/− 8,4) | 13,5 (+/− 2,8) | 4,6 (+/− 1,2) | 1 | 11,7 (+/− 3,0) | 14,5 (+/− 3,02) | 2,3 (+/− 0,5) |
| IFN-β | 1 | 167,1 (+/− 55,4) | 132,3 (+/− 33,7) | 23,1 (+/− 7,8) | 1 | 277,1 (+/− 56,8) | 223,4 (+/− 45,2) | 38,4 (+/− 7,02) | 1 | 278,7 (+/− 70,6) | 256,3 (+/− 59,4) | 33,2 (+/− 8,5) | 1 | 184,0 (+/− 40,6) | 242,9 (+/− 69,7) | 32,4 (+/− 6,9) | |
| IFN-γ | 1 | 5,8 (+/− 1,2) | 3,5 (+/− 0,6) | 2,4 (+/− 0,4) | 1 | 22,8 (+/− 4,2) | 22,5 (+/− 4,6) | 32,5 (+/− 7,6) | 1 | 32,7 (+/− 7,6) | 26,8 (+/− 3,8) | 30,4 (+/− 7) | 1 | 28,5 (+/− 4,3) | 32,1 (+/− 7,7) | 27,6 (+/− 6,3) | |
| Inflammatory response | TNF-α | 1 | 4,2 (+/− 0,9) | 4,6 (+/− 1,0) | 1,9 (+/− 0,2) | 1 | 8,6 (+/− 1,5) | 10,5 (+/− 1,8) | 4,7 (+/− 0,7) | 1 | 7,4 (+/− 1,4) | 6,9 (+/− 0,9) | 1,5 (+/− 0,1) | 1 | 5,3 (+/− 0,7) | 7,0 (+/− 1,6) | 1,6 (+/− 0,2) |
| IL-1β | 1 | 1,9 (+/− 0,3) | 1,4 (+/− 0,2) | 1,3 (+/− 0,3) | 1 | 1,6 (+/− 0,1) | 2,0 (+/− 0,3) | 1,2 (+/− 0,1) | 1 | 3,5 (+/− 0,8) | 2,9 (+/− 0,4) | 1,2 (+/− 0,2) | 1 | 3,5 (+/− 0,4) | 2,9 (+/− 0,6) | 1,1 (+/− 0,1) | |
| IL-6 | 1 | 0,7 (+/− 0,04) | 0,8 (+/− 0,1) | 0,7 (+/− 0,2) | 1 | 2,4 (+/− 0,5) | 6,0 (+/− 1,5) | 6,3 (+/− 1,5) | 1 | 4,1 (+/− 0,8) | 6,1 (+/− 2,1) | 1,1 (+/− 0,3) | 1 | 4,0 (+/− 0,8) | 6,6 (+/− 1,72) | 3,5 (+/− 1,3) | |
| Treg response | IL-10 | 1 | 3,9 (+/− 1,0) | 11,5 (+/− 3,6) | 22,1 (+/− 5,2) | 1 | 7,02 (+/− 2,1) | 10,5 (+/− 0,9) | 10,0 (+/− 2,5) | 1 | 13,7 (+/− 4,7) | 12,6 (+/− 1,3) | 5,5 (+/− 1,2) | 1 | 7,7 (+/− 1,9) | 12,7 (+/− 1,1) | 10,6 (+/− 2,1) |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hue, E.S.; Richard, E.A.; Fortier, C.I.; Fortier, G.D.; Paillot, R.; Raue, R.; Pronost, S.L. Equine PBMC Cytokines Profile after In Vitro α- and γ-EHV Infection: Efficacy of a Parapoxvirus Ovis Based-Immunomodulator Treatment. Vaccines 2017, 5, 28. https://doi.org/10.3390/vaccines5030028
Hue ES, Richard EA, Fortier CI, Fortier GD, Paillot R, Raue R, Pronost SL. Equine PBMC Cytokines Profile after In Vitro α- and γ-EHV Infection: Efficacy of a Parapoxvirus Ovis Based-Immunomodulator Treatment. Vaccines. 2017; 5(3):28. https://doi.org/10.3390/vaccines5030028
Chicago/Turabian StyleHue, Erika S., Eric A. Richard, Christine I. Fortier, Guillaume D. Fortier, Romain Paillot, Rudiger Raue, and Stéphane L. Pronost. 2017. "Equine PBMC Cytokines Profile after In Vitro α- and γ-EHV Infection: Efficacy of a Parapoxvirus Ovis Based-Immunomodulator Treatment" Vaccines 5, no. 3: 28. https://doi.org/10.3390/vaccines5030028