
Retromer-Mediated Trafficking of Transmembrane Receptors and Transporters

Abstract
:1. Introduction

2. The Retromer Complex and Trafficking
2.1. Retrograde Transport from Endosomes towards the TGN

2.2. Transcytosis
3. Retromer-Binding Receptors and Transporters
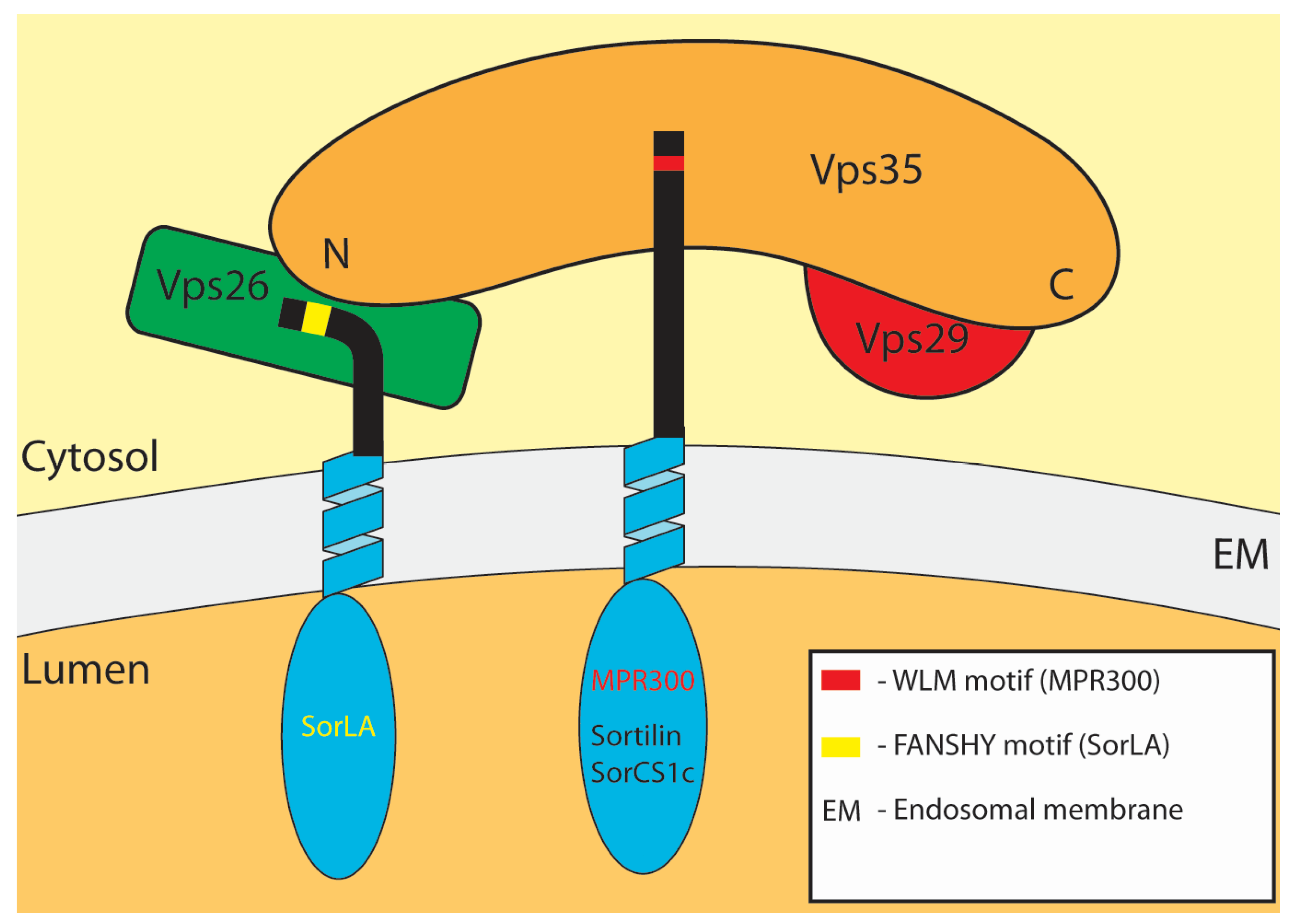
3.1. Cation-Independent Mannose-6-Phosphate Receptor (MPR300)

{kind=link}
{kind=link}
| Receptors | Type | Ligands | Retromer sorting | Interacting retromer subunit | Receptor binding motif | References |
|---|---|---|---|---|---|---|
| CI-MPR (MPR300) | Type-I | M6P containing ligands: acidic hydrolases, TGF-β1, proliferin, granzyme B, thyroglobulin; Non-M6P containing ligands: IGF-II, retinoic acid, uPAR, plasminogen, serglycin | EE to TGN | Vps35 | WLM | 26 |
| SorLA (LR11) | Type-I | LpL, apoE, apoA-V-DMPC, GDNF, GFRα1, uPA-PAI1 PAI-1, tPA-PAI-1, PDGF-BB, APP, SPAK, TrkB | EE to TGN | Vps26 | FANSHY | 30, 107 |
| Sortilin | Type-I | LpL, pro-NGF, neurotensin, progranulin PGRN, IL6, apoE, PCSK9, delta-like homologue receptor, IL6, IFN-gamma, CNTF, apoAV, apoB100, APP | EE to TGN | Vps35 | Not known | 29,105 |
| SorCS-1 | Type-I | PDGF-BB, APP, sortilin | Not known | Vps35 | Not known | 108,109 |
| pIgR | Type-I | IgA | Transcytosis | Vps35 | Not known | 65 |
| Wntless (GPR177) | 7TM | Wtn family proteins | EE to TGN | Vps35 | FLM | 120, 124 |
| Transporters | ||||||
| Menkes protein (ATP7A) | ATPase (8TM) | Cu++ | Recycling to cell surface | Vps26 | Not known | 45, 132 |
| Glut4 | 12 TM | Glucose | EE to TGN | Not known | Not known | 128, 130 |
| DMT1-II | 12 TM | Divalent cation transporter | Recycling to cell surface | Not known | YLL | 135, 136 |
3.2. Sortilins
3.3. The Wntless Receptor
3.4. Other Receptors and Transporters
4. Concluding Remarks
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bonifacino, J.S.; Rojas, R. Retrograde transport from endosomes to the trans-Golgi network. Nat. Rev. Mol. Cell Biol. 2006, 7, 568–579. [Google Scholar] [CrossRef] [PubMed]
- Spang, A. Retrograde traffic from the Golgi to the endoplasmic reticulum. Cold Spring Harb. Perspect. Biol. 2013, 5. [Google Scholar] [CrossRef] [PubMed]
- Johannes, L.; Wunder, C. Retrograde transport: Two (or more) roads diverged in an endosomal tree? Traffic 2011, 12, 956–962. [Google Scholar] [CrossRef] [PubMed]
- Chia, P.Z.; Gunn, P.; Gleeson, P.A. Cargo trafficking between endosomes and the trans-Golgi network. Histochem. Cell Biol. 2013, 140, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, T. Emerging roles of recycling endosomes. J. Biochem. 2013, 153, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Sheff, D.R.; Daro, E.A.; Hull, M.; Mellman, I. The receptor recycling pathway contains two distinct populations of early endosomes with different sorting functions. J. Cell Biol. 1999, 145, 123–139. [Google Scholar] [CrossRef] [PubMed]
- Sonnichsen, B.; De Renzis, S.; Nielsen, E.; Rietdorf, J.; Zerial, M. Distinct membrane domains on endosomes in the recycling pathway visualized by multicolor imaging of Rab4, Rab5, and Rab11. J. Cell Biol. 2000, 149, 901–914. [Google Scholar] [CrossRef] [PubMed]
- Johannes, L.; Popoff, V. Tracing the Retrograde Route in Protein Trafficking. Cell 2008, 135, 1175–1187. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, L.S.; Yang, Z. Microtubule-based transport systems in neurons: the roles of kinesins and dyneins. Annu. Rev. Neurosci. 2000, 23, 39–71. [Google Scholar] [CrossRef] [PubMed]
- Horton, A.C.; Ehlers, M.D. Neuronal polarity and trafficking. Neuron 2003, 40, 277–295. [Google Scholar] [CrossRef]
- Lasiecka, Z.M.; Winckler, B. Mechanisms of polarized membrane trafficking in neurons—Focusing in on endosomes. Mol. Cell. Neurosci. 2011, 48, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Small, S.A.; Petsko, G.A. Retromer in Alzheimer disease, Parkinson disease and other neurological disorders. Nat. Rev. Neurosci. 2015, 16, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Vilarino-Guell, C.; Wider, C.; Ross, O.A.; Dachsel, J.C.; Kachergus, J.M.; Lincoln, S.J.; Soto-Ortolaza, A.I.; Cobb, S.A.; Wilhoite, G.J.; Bacon, J.A.; et al. VPS35 mutations in Parkinson disease. Am. J. Hum. Genet. 2011, 89, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Yu, S.; Sakamori, R.; Stypulkowski, E.; Gao, N. Wntless in Wnt secretion: Molecular, cellular and genetic aspects. Front. Biol. 2012, 7, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Sandvig, K.; Garred, O.; Prydz, K.; Kozlov, J.V.; Hansen, S.H.; van Deurs, B. Retrograde transport of endocytosed Shiga toxin to the endoplasmic reticulum. Nature 1992, 358, 510–512. [Google Scholar] [CrossRef] [PubMed]
- Blagoveshchenskaya, A.D.; Thomas, L.; Feliciangeli, S.F.; Hung, C.H.; Thomas, G. HIV-1 Nef downregulates MHC-I by a PACS-1- and PI3K-regulated ARF6 endocytic pathway. Cell 2002, 111, 853–866. [Google Scholar] [CrossRef]
- Chinnapen, D.J.; Chinnapen, H.; Saslowsky, D.; Lencer, W.I. Rafting with cholera toxin: Endocytosis and trafficking from plasma membrane to ER. FEMS Microbiol. Lett. 2007, 266, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Gonatas, N.K.; Steiber, A.; Kim, S.U.; Graham, D.I.; Avrameas, S. Internalization of neuronal plasma membrane ricin receptors into the Golgi apparatus. Exp. Cell Res. 1975, 94, 426–431. [Google Scholar] [CrossRef]
- Kerr, M.; Teasdale, R.D. Live imaging of endosome dynamics. Semin. Cell Dev. Biol. 2014, 31, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Scott, C.C.; Vacca, F.; Gruenberg, J. Endosome maturation, transport and functions. Semin. Cell Dev. Biol. 2014, 31, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Tooze, J.; Hollinshead, M. Tubular early endosomal networks in AtT20 and other cells. J. Cell Biol. 1991, 115, 635–653. [Google Scholar] [CrossRef] [PubMed]
- Seaman, M.N.; McCaffery, J.M.; Emr, S.D. A membrane coat complex essential for endosome-to-Golgi retrograde transport in yeast. J. Cell Biol. 1998, 142, 665–681. [Google Scholar] [CrossRef] [PubMed]
- Nothwehr, S.F.; Bruinsma, P.; Strawn, L.A. Distinct domains within Vps35p mediate the retrieval of two different cargo proteins from the yeast prevacuolar/endosomal compartment. Mol. Biol. Cell 1999, 10, 875–890. [Google Scholar] [CrossRef] [PubMed]
- Horazdovsky, B.F.; Davies, B.A.; Seaman, M.N.; McLaughlin, S.A.; Yoon, S.; Emr, S.D. A sorting nexin-1 homologue, Vps5p, forms a complex with Vps17p and is required for recycling the vacuolar protein-sorting receptor. Mol. Biol. Cell 1997, 8, 1529–1541. [Google Scholar] [CrossRef] [PubMed]
- Norwood, S.J.; Shaw, D.J.; Cowieson, N.P.; Owen, D.J.; Teasdale, R.D.; Collins, B.M. Assembly and solution structure of the core retromer protein complex. Traffic 2011, 12, 56–71. [Google Scholar] [CrossRef] [PubMed]
- Rojas, R.; Kametaka, S.; Haft, C.R.; Bonifacino, J.S. Interchangeable but essential functions of SNX1 and SNX2 in the association of retromer with endosomes and the trafficking of mannose 6-phosphate receptors. Mol. Cell. Biol. 2007, 27, 1112–1124. [Google Scholar] [CrossRef] [PubMed]
- Wassmer, T.; Attar, N.; Bujny, M.V.; Oakley, J.; Traer, C.J.; Cullen, P.J. A loss-of-function screen reveals SNX5 and SNX6 as potential components of the mammalian retromer. J. Cell Sci. 2007, 120, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Hierro, A.; Rojas, A.L.; Rojas, R.; Murthy, N.; Effantin, G.; Kajava, A.V.; Steven, A.C.; Bonifacino, J.S.; Hurley, J.H. Functional architecture of the retromer cargo-recognition complex. Nature 2007, 449, 1063–1067. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Lee, Y.; Lee, H.J.; Kim, J.S.; Song, B.S.; Huh, J.W.; Lee, S.R.; Kim, S.U.; Kim, S.H.; Hong, Y.; et al. Implication of mouse Vps26b-Vps29-Vps35 retromer complex in sortilin trafficking. Biochem. Biophys. Res. Commun. 2010, 403, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Fjorback, A.W.; Seaman, M.; Gustafsen, C.; Mehmedbasic, A.; Gokool, S.; Wu, C.; Militz, D.; Schmidt, V.; Madsen, P.; Nyengaard, J.R.; et al. Retromer binds the FANSHY sorting motif in SorLA to regulate amyloid precursor protein sorting and processing. J. Neurosci. 2012, 32, 1467–1480. [Google Scholar] [CrossRef] [PubMed]
- Gallon, M.; Cullen, P.J. Retromer and sorting nexins in endosomal sorting. Biochem. Soc. Trans. 2015, 43, 33–47. [Google Scholar] [CrossRef] [PubMed]
- Bugarcic, A.; Zhe, Y.; Kerr, M.C.; Griffin, J.; Collins, B.M.; Teasdale, R.D. Vps26A and Vps26B subunits define distinct retromer complexes. Traffic 2011, 12, 1759–1773. [Google Scholar] [CrossRef] [PubMed]
- Seaman, M.N.; Harbour, M.E.; Tattersall, D.; Read, E.; Bright, N. Membrane recruitment of the cargo-selective retromer subcomplex is catalysed by the small GTPase Rab7 and inhibited by the Rab-GAP TBC1D5. J. Cell Sci. 2009, 122, 2371–2382. [Google Scholar] [CrossRef] [PubMed]
- Rojas, R.; van Vlijmen, T.; Mardones, G.A.; Prabhu, Y.; Rojas, A.L.; Mohammed, S.; Heck, A.J.; Raposo, G.; van der Sluijs, P.; Bonifacino, J.S. Sorting nexin-1 mediates tubular endosome-to-TGN transport through coincidence sensing of high- curvature membranes and 3-phosphoinositides. Curr. Biol. 2004, 14, 1791–1800. [Google Scholar]
- Carlton, J.; Bujny, M.; Peter, B.J.; Oorschot, V.M.; Rutherford, A.; Mellor, H.; Klumperman, J.; McMahon, H.T.; Cullen, P.J. Sorting nexin-1 mediates tubular endosome-to-tgn transport through coincidence sensing of high- curvature membranes and 3-phosphoinositides. Current biology : CB 2004, 14, 1791–1800. [Google Scholar] [CrossRef] [PubMed]
- Zimmerberg, J.; McLaughlin, S. Membrane curvature: How BAR domains bend bilayers. Curr. Biol. 2004, 14, R250–R252. [Google Scholar] [CrossRef] [PubMed]
- Teasdale, R.D.; Collins, B.M. Insights into the PX (phox-homology) domain and SNX (sorting nexin) protein families: Structures, functions and roles in disease. Biochem. J. 2012, 441, 39–59. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, V.K.; Madsen, K.L.; Bolinger, P.Y.; Kunding, A.; Hedegard, P.; Gether, U.; Stamou, D. Amphipathic motifs in BAR domains are essential for membrane curvature sensing. EMBO J. 2009, 28, 3303–3314. [Google Scholar] [CrossRef] [PubMed]
- Peter, B.J.; Kent, H.M.; Mills, I.G.; Vallis, Y.; Butler, P.J.; Evans, P.R.; McMahon, H.T. BAR domains as sensors of membrane curvature: The amphiphysin BAR structure. Science 2004, 303, 495–499. [Google Scholar] [CrossRef] [PubMed]
- Priya, A.; Kalaidzidis, I.V.; Kalaidzidis, Y.; Lambright, D.; Datta, S. Molecular insights into Rab7-mediated endosomal recruitment of core retromer: deciphering the role of Vps26 and Vps35. Traffic 2015, 16, 68–84. [Google Scholar] [CrossRef] [PubMed]
- Harterink, M.; Port, F.; Lorenowicz, M.J.; McGough, I.J.; Silhankova, M.; Betist, M.C.; van Weering, J.R.T.; van Heesbeen, R.G.H.P.; Middelkoop, T.C.; Basler, K.; et al. A SNX3-dependent retromer pathway mediates retrograde transport of the Wnt sorting receptor Wntless and is required for Wnt secretion. Nat. Cell Biol. 2011, 13, 914–923. [Google Scholar] [CrossRef] [PubMed]
- Strochlic, T.I.; Setty, T.G.; Sitaram, A.; Burd, C.G. Grd19/Snx3p functions as a cargo-specific adapter for retromer-dependent endocytic recycling. J. Cell Biol. 2007, 177, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Temkin, P.; Lauffer, B.; Jager, S.; Cimermancic, P.; Krogan, N.J.; von Zastrow, M. SNX27 mediates retromer tubule entry and endosome-to-plasma membrane trafficking of signalling receptors. Nat. Cell Biol. 2011, 13, 715–721. [Google Scholar] [CrossRef] [PubMed]
- Lauffer, B.E.; Melero, C.; Temkin, P.; Lei, C.; Hong, W.; Kortemme, T.; von Zastrow, M. SNX27 mediates PDZ-directed sorting from endosomes to the plasma membrane. J. Cell Biol. 2010, 190, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, F.; Gallon, M.; Winfield, M.; Thomas, E.C.; Bell, A.J.; Heesom, K.J.; Tavare, J.M.; Cullen, P.J. A global analysis of SNX27-retromer assembly and cargo specificity reveals a function in glucose and metal ion transport. Nat. Cell Biol. 2013, 15, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Wu, Y.; Belenkaya, T.Y.; Lin, X. SNX3 controls Wingless/Wnt secretion through regulating retromer-dependent recycling of Wntless. Cell Res. 2011, 21, 1677–1690. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Garcia-Santos, D.; Ishikawa, Y.; Seguin, A.; Li, L.; Fegan, K.H.; Hildick-Smith, G.J.; Shah, D.I.; Cooney, J.D.; Chen, W.; et al. Snx3 regulates recycling of the transferrin receptor and iron assimilation. Cell Metab. 2013, 17, 343–352. [Google Scholar] [CrossRef] [PubMed]
- McGough, I.J.; Cullen, P.J. Recent advances in retromer biology. Traffic 2011, 12, 963–971. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Hong, W. From endosomes to the trans-Golgi network. Semin. Cell Dev. Biol. 2014, 31, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Burke, B.; Griffiths, G.; Reggio, H.; Louvard, D.; Warren, G. A monoclonal antibody against a 135-K Golgi membrane protein. EMBO J. 1982, 1, 1621–1628. [Google Scholar] [PubMed]
- Saint-Pol, A.; Yelamos, B.; Amessou, M.; Mills, I.G.; Dugast, M.; Tenza, D.; Schu, P.; Antony, C.; McMahon, H.T.; Lamaze, C.; et al. Clathrin adaptor epsinR is required for retrograde sorting on early endosomal membranes. Dev. Cell. 2004, 6, 525–538. [Google Scholar] [CrossRef]
- Stoorvogel, W.; Oorschot, V.; Geuze, H.J. A novel class of clathrin-coated vesicles budding from endosomes. J. Cell Biol. 1996, 132, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Draper, R.K.; Goda, Y.; Brodsky, F.M.; Pfeffer, S.R. Antibodies to clathrin inhibit endocytosis but not recycling to the trans Golgi network in vitro. Science 1990, 248, 1539–1541. [Google Scholar] [CrossRef] [PubMed]
- McGough, I.J.; Cullen, P.J. Clathrin is not required for SNX-BAR-retromer-mediated carrier formation. J. Cell Sci. 2013, 126, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Popoff, V.; Mardones, G.A.; Bai, S.K.; Chambon, V.; Tenza, D.; Burgos, P.V.; Shi, A.; Benaroch, P.; Urbe, S.; Lamaze, C.; et al. Analysis of articulation between clathrin and retromer in retrograde sorting on early endosomes. Traffic 2009, 10, 1868–1880. [Google Scholar] [CrossRef] [PubMed]
- Shi, A.; Sun, L.; Banerjee, R.; Tobin, M.; Zhang, Y.; Grant, B.D. Regulation of endosomal clathrin and retromer-mediated endosome to Golgi retrograde transport by the J-domain protein RME-8. EMBO J. 2009, 28, 3290–3302. [Google Scholar] [CrossRef] [PubMed]
- Gomez, T.S.; Billadeau, D.D. A FAM21-containing WASH complex regulates retromer-dependent sorting. Dev. Cell 2009, 17, 699–711. [Google Scholar] [CrossRef] [PubMed]
- Freeman, C.L.; Hesketh, G.; Seaman, M.N. RME-8 coordinates the activity of the WASH complex with the function of the retromer SNX dimer to control endosomal tubulation. J. Cell Sci. 2014, 127, 2053–2070. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Zhang, C.; Sun, Z.; Hong, Z.; Li, K.; Sun, D.; Yang, Y.; Tian, C.; Gong, W.; Liu, J.J. PtdIns(4)P regulates retromer-motor interaction to facilitate dynein-cargo dissociation at the trans-Golgi network. Nat. Cell Biol. 2013, 15, 417–429. [Google Scholar] [CrossRef] [PubMed]
- Granger, E.; McNee, G.; Allan, V.; Woodman, P. The role of the cytoskeleton and molecular motors in endosomal dynamics. Semin. Cell Dev. Biol. 2014, 31, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Wassmer, T.; Attar, N.; Harterink, M.; van Weering, J.R.; Traer, C.J.; Oakley, J.; Goud, B.; Stephens, D.J.; Verkade, P.; Korswagen, H.C.; et al. The retromer coat complex coordinates endosomal sorting and dynein-mediated transport, with carrier recognition by the trans-Golgi network. Dev. Cell 2009, 17, 110–122. [Google Scholar] [CrossRef] [PubMed]
- Chia, P.Z.; Gasnereau, I.; Lieu, Z.Z.; Gleeson, P.A. Rab9-dependent retrograde transport and endosomal sorting of the endopeptidase furin. J. Cell Sci. 2011, 124, 2401–2413. [Google Scholar] [CrossRef] [PubMed]
- Ramanujam, K.S.; Seetharam, S.; Dahms, N.M.; Seetharam, B. Effect of processing inhibitors on cobalamin (vitamin B12) transcytosis in polarized opossum kidney cells. Arch. Biochem. Biophys. 1994, 315, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Tuma, P.; Hubbard, A.L. Transcytosis: Crossing cellular barriers. Physiol. Rev. 2003, 83, 871–932. [Google Scholar] [CrossRef] [PubMed]
- Verges, M.; Luton, F.; Gruber, C.; Tiemann, F.; Reinders, L.G.; Huang, L.; Burlingame, A.L.; Haft, C.R.; Mostov, K.E. The mammalian retromer regulates transcytosis of the polymeric immunoglobulin receptor. Nat. Cell Biol. 2004, 6, 763–769. [Google Scholar] [CrossRef] [PubMed]
- Mellado, M.; Cuartero, Y.; Brugada, R.; Verges, M. Subcellular localisation of retromer in post-endocytic pathways of polarised Madin-Darby canine kidney cells. Biol. Cell 2014, 106, 377–393. [Google Scholar] [CrossRef] [PubMed]
- Sahagian, G.G.; Distler, J.; Jourdian, G.W. Characterization of a membrane-associated receptor from bovine liver that binds phosphomannosyl residues of bovine testicular beta-galactosidase. Proc. Natl. Acad. Sci. USA 1981, 78, 4289–4293. [Google Scholar] [CrossRef] [PubMed]
- Hancock, M.K.; Yammani, R.D.; Dahms, N.M. Localization of the carbohydrate recognition sites of the insulinlike growth factor II/mannose 6-phosphate receptor to domains 3 and 9 of the extracytoplasmic region. J. Biol. Chem. 2002, 277, 47205–47212. [Google Scholar] [CrossRef] [PubMed]
- Hawkes, C.; Kar, S. The insulin-like growth factor-II/mannose-6-phosphate receptor: Structure, distribution and function in the central nervous system. Brain Res. Rev. 2004, 44, 117–140. [Google Scholar] [CrossRef] [PubMed]
- Lobel, P.; Dahms, N.M.; Kornfeld, S. Cloning and sequence analysis of the cation-independent mannose 6-phosphate receptor. J. Biol. Chem. 1988, 263, 2563–2570. [Google Scholar] [PubMed]
- Olson, L.J.; Castonguay, A.C.; Lasanajak, Y.; Peterson, F.C.; Cummings, R.D.; Smith, D.F.; Dahms, N.M. Identification of a fourth mannose 6-phosphate binding site in the cation-independent mannose 6-phosphate receptor. Glycobiology 2015, 25, 591–606. [Google Scholar] [CrossRef] [PubMed]
- Oshima, A.; Nolan, C.M.; Kyle, J.W.; Grubb, J.H.; Sly, W.S. The human cation-independent mannose 6-phosphate receptor. Cloning and sequence of the full-length cDNA and expression of functional receptor in cos cells. J. Biol. Chem. 1988, 263, 2553–2562. [Google Scholar] [PubMed]
- Reddy, S.T.; Chai, W.; Childs, R.A.; Page, J.D.; Feizi, T.; Dahms, N.M. Identification of a low affinity mannose 6-phosphate-binding site in domain 5 of the cation-independent mannose 6-phosphate receptor. J. Biol. Chem. 2004, 279, 38658–38667. [Google Scholar] [CrossRef] [PubMed]
- Braulke, T.; Bonifacino, J.S. Sorting of lysosomal proteins. Biochim. Biophys. Acta Mol. Cell Res. 2009, 1793, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, P.; Kornfeld, S. The cytoplasmic tail of the cation-independent mannose 6-phosphate receptor contains four binding sites for AP-1. Arch. Biochem. Biophys. 2004, 426, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Puertollano, R.; Aguilar, R.C.; Gorshkova, I.; Crouch, R.J.; Bonifacino, J.S. Sorting of mannose 6-phosphate receptors mediated by the GGAs. Science 2001, 292, 1712–1716. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Doray, B.; Poussu, A.; Lehto, V.P.; Kornfeld, S. Binding of GGA2 to the lysosomal enzyme sorting motif of the mannose 6-phosphate receptor. Science 2001, 292, 1716–1718. [Google Scholar] [CrossRef] [PubMed]
- Doray, B.; Ghosh, P.; Griffith, J.; Geuze, H.J.; Kornfeld, S. Cooperation of GGAs and AP-1 in packaging MPRs at the trans-Golgi network. Science (N. Y.) 2002, 297, 1700–1703. [Google Scholar] [CrossRef] [PubMed]
- Kollmann, K.; Pohl, S.; Marschner, K.; Encarnacao, M.; Sakwa, I.; Tiede, S.; Poorthuis, B.J.; Lubke, T.; Muller-Loennies, S.; Storch, S.; et al. Mannose phosphorylation in health and disease. Eur. J. Cell Biol. 2010, 89, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Arighi, C.N.; Harmell, L.M.; Aguilar, R.C.; Haft, C.R.; Bonifacino, J.S. Role of the mammalian retromer in sorting of the cation-independent mannose 6-phosphate receptor. J. Cell Biol. 2004, 165, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Riederer, M.A.; Soldati, T.; Shapiro, A.D.; Lin, J.; Pfeffer, S.R. Lysosome biogenesis requires rab9 function and receptor recycling from endosomes to the trans-Golgi network. J. Cell Biol. 1994, 125, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, D.; Soldati, T.; Riederer, M.A.; Goda, Y.; Zerial, M.; Pfeffer, S.R. Rab9 functions in transport between late endosomes and the trans Golgi network. EMBO J. 1993, 12, 677–682. [Google Scholar] [PubMed]
- Mari, M.; Bujny, M.V.; Zeuschner, D.; Geerts, W.J.C.; Griffith, J.; Petersen, C.M.; Cullen, P.J.; Klumperman, J.; Geuze, H.J. SNX1 defines an early endosomal recycling exit for sortilin and mannose 6-phosphate receptors. Traffic 2008, 9, 380–393. [Google Scholar] [CrossRef] [PubMed]
- Umeda, A.; Fujita, H.; Kuronita, T.; Hirosako, K.; Himeno, M.; Tanaka, Y. Distribution and trafficking of MPR300 is normal in cells with cholesterol accumulated in late endocytic compartments: Evidence for early endosome-to-TGN trafficking of MPR300. J. Lipid Res. 2003, 44, 1821–1832. [Google Scholar] [CrossRef] [PubMed]
- Díaz, E.; Pfeffer, S.R. TIP47: A cargo selection device for mannose 6-phosphate receptor trafficking. Cell 1998, 93, 433–443. [Google Scholar] [CrossRef]
- Bulankina, A.V.; Deggerich, A.; Wenzel, D.; Mutenda, K.; Wittmann, J.G.; Rudolph, M.G.; Burger, K.N.; Honing, S. TIP47 functions in the biogenesis of lipid droplets. J. Cell Biol. 2009, 185, 641–655. [Google Scholar] [CrossRef] [PubMed]
- Crump, C.M.; Xiang, Y.; Thomas, L.; Gu, F.; Austin, C.; Tooze, S.A.; Thomas, G. PACS-1 binding to adaptors is required for acidic cluster motif-mediated protein traffic. EMBO J. 2001, 20, 2191–2201. [Google Scholar] [CrossRef] [PubMed]
- Wan, L.; Molloy, S.S.; Thomas, L.; Liu, G.; Xiang, Y.; Rybak, S.L.; Thomas, G. PACS-1 defines a novel gene family of cytosolic sorting proteins required for trans-Golgi network localization. Cell 1998, 94, 205–216. [Google Scholar] [CrossRef]
- Itin, C.; Rancaño, C.; Nakajima, Y.; Pfeffer, S.R. A novel assay reveals a role for soluble N-ethylmaleimide-sensitive fusion attachment protein in mannose 6-phosphate receptor transport from endosomes to the trans Golgi network. J. Biol. Chem. 1997, 272, 27737–27744. [Google Scholar] [CrossRef] [PubMed]
- Seaman, M.N.J. Identification of a novel conserved sorting motif required for retromer-mediated endosome-to-TGN retrieval. J. Cell Sci. 2007, 120, 2378–2389. [Google Scholar] [CrossRef] [PubMed]
- Petersen, C.M.; Nielsen, M.S.; Nykjaer, A.; Jacobsen, L.; Tommerup, N.; Rasmussen, H.H.; Roigaard, H.; Gliemann, J.; Madsen, P.; Moestrup, S.K. Molecular identification of a novel candidate sorting receptor purified from human brain by receptor-associated protein affinity chromatography. J. Biol. Chem. 1997, 272, 3599–3605. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, L.; Madsen, P.; Moestrup, S.K.; Lund, A.H.; Tommerup, N.; Nykjaer, A.; Sottrup-Jensen, L.; Gliemann, J.; Petersen, C.M. Molecular characterization of a novel human hybrid-type receptor that binds the alpha2-macroglobulin receptor-associated protein. J. Biol. Chem. 1996, 271, 31379–31383. [Google Scholar] [CrossRef] [PubMed]
- Hermey, G.; Riedel, I.B.; Hampe, W.; Schaller, H.C.; Hermans-Borgmeyer, I. Identification and characterization of SorCS, a third member of a novel receptor family. Biochem. Biophys. Res. Commun. 1999, 266, 347–351. [Google Scholar] [CrossRef] [PubMed]
- Hermey, G.; Sjogaard, S.S.; Petersen, C.M.; Nykjaer, A.; Gliemann, J. Tumour necrosis factor alpha-converting enzyme mediates ectodomain shedding of Vps10p-domain receptor family members. Biochem. J. 2006, 395, 285–293. [Google Scholar] [PubMed]
- Glerup, S.; Nykjaer, A.; Vaegter, C.B. Sortilins in neurotrophic factor signaling. Handb. Exp. Pharmacol. 2014, 220, 165–189. [Google Scholar] [PubMed]
- Lefrancois, S.; Zeng, J.; Hassan, A.J.; Canuel, M.; Morales, C.R. The lysosomal trafficking of sphingolipid activator proteins (SAPs) is mediated by sortilin. EMBO J. 2003, 22, 6430–6437. [Google Scholar] [CrossRef] [PubMed]
- Mazella, J.; Petrault, O.; Lucas, G.; Deval, E.; Beraud-Dufour, S.; Gandin, C.; El-Yacoubi, M.; Widmann, C.; Guyon, A.; Chevet, E.; et al. Spadin, a sortilin-derived peptide, targeting rodent TREK-1 channels: A new concept in the antidepressant drug design. PLoS Biol. 2010, 8, e1000355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, E.N.; Melbin, J.; Spear, J.F.; Hill, J.D. Sequence of atrial excitation in the dog during antegrade and retrograde activation. J. Electrocardiol. 1971, 4, 283–290. [Google Scholar] [CrossRef]
- Andersen, O.M.; Reiche, J.; Schmidt, V.; Gotthardt, M.; Spoelgen, R.; Behlke, J.; von Arnim, C.A.; Breiderhoff, T.; Jansen, P.; Wu, X.; et al. Neuronal sorting protein-related receptor sorLA/LR11 regulates processing of the amyloid precursor protein. Proc. Natl. Acad. Sci. USA 2005, 102, 13461–13466. [Google Scholar] [CrossRef] [PubMed]
- Klinger, S.C.; Glerup, S.; Raarup, M.K.; Mari, M.C.; Nyegaard, M.; Koster, G.; Prabakaran, T.; Nilsson, S.K.; Kjaergaard, M.M.; Bakke, O.; et al. SorLA regulates the activity of lipoprotein lipase by intracellular trafficking. J. Cell Sci. 2011, 124, 1095–1105. [Google Scholar] [CrossRef] [PubMed]
- Gustafsen, C.; Glerup, S.; Pallesen, L.T.; Olsen, D.; Andersen, O.M.; Nykjaer, A.; Madsen, P.; Petersen, C.M. Sortilin and SorLA display distinct roles in processing and trafficking of amyloid precursor protein. J. Neurosci. 2013, 33, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, M.B.; Kjolby, M.; Gunnersen, S.; Larsen, J.V.; Palmfeldt, J.; Falk, E.; Nykjaer, A.; Bentzon, J.F. Targeting sortilin in immune cells reduces proinflammatory cytokines and atherosclerosis. J. Clin. Investig. 2014, 124, 5317–5322. [Google Scholar] [CrossRef] [PubMed]
- Kjolby, M.; Nielsen, M.S.; Petersen, C.M. Sortilin, encoded by the cardiovascular risk gene SORT1, and its suggested functions in cardiovascular disease. Curr. Atheroscler. Rep. 2015, 17, 496. [Google Scholar] [CrossRef] [PubMed]
- Kebede, M.A.; Oler, A.T.; Gregg, T.; Balloon, A.J.; Johnson, A.; Mitok, K.; Rabaglia, M.; Schueler, K.; Stapleton, D.; Thorstenson, C.; et al. SORCS1 is necessary for normal insulin secretory granule biogenesis in metabolically stressed beta cells. J. Clin. Investig. 2014, 124, 4240–4256. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, M.S.; Madsen, P.; Christensen, E.I.; Nykjaer, A.; Gliemann, J.; Kasper, D.; Pohlmann, R.; Petersen, C.M. The sortilin cytoplasmic tail conveys Golgi-endosome transport and binds the VHS domain of the GGA2 sorting protein. EMBO J. 2001, 20, 2180–2190. [Google Scholar] [CrossRef] [PubMed]
- Baltes, J.; Larsen, J.V.; Radhakrishnan, K.; Geumann, C.; Kratzke, M.; Petersen, C.M.; Schu, P. Sigma1b adaptin regulates adipogenesis by mediating the sorting of sortilin in adipose tissue. J. Cell Sci. 2014, 127, 3477–3487. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, M.S.; Gustafsen, C.; Madsen, P.; Nyengaard, J.R.; Hermey, G.; Bakke, O.; Mari, M.; Schu, P.; Pohlmann, R.; Dennes, A.; et al. Sorting by the cytoplasmic domain of the amyloid precursor protein binding receptor SorLA. Mol. Cell Biol. 2007, 27, 6842–6851. [Google Scholar] [CrossRef] [PubMed]
- Lane, R.F.; Raines, S.M.; Steele, J.W.; Ehrlich, M.E.; Lah, J.A.; Small, S.A.; Tanzi, R.E.; Attie, A.D.; Gandy, S. Diabetes-associated SorCS1 regulates Alzheimer’s amyloid-beta metabolism: evidence for involvement of SorL1 and the retromer complex. J. Neurosci. 2010, 30, 13110–13115. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, M.S.; Keat, S.J.; Hamati, J.W.; Madsen, P.; Gutzmann, J.J.; Engelsberg, A.; Pedersen, K.M.; Gustafsen, C.; Nykjaer, A.; Gliemann, J.; et al. Different motifs regulate trafficking of SorCS1 isoforms. Traffic 2008, 9, 980–994. [Google Scholar] [CrossRef] [PubMed]
- Bänziger, C.; Soldini, D.; Schütt, C.; Zipperlen, P.; Hausmann, G.; Basler, K. Wntless, a Conserved Membrane Protein Dedicated to the Secretion of Wnt Proteins from Signaling Cells. Cell 2006, 125, 509–522. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, B.T.; Tamai, K.; He, X. Wnt/beta-Catenin Signaling: Components, Mechanisms, and Diseases. Dev. Cell 2009, 17, 9–26. [Google Scholar] [CrossRef] [PubMed]
- Port, F.; Basler, K. Wnt Trafficking: New Insights into Wnt Maturation, Secretion and Spreading. Traffic 2010, 11, 1265–1271. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Kittanakom, S.; Wong, V.; Reyes, B.A.S.; Van Bockstaele, E.J.; Stagljar, I.; Berrettini, W.; Levenson, R. Interaction of the mu-opioid receptor with GPR177 (Wntless) inhibits Wnt secretion: potential implications for opioid dependence. BMC Neurosci. 2010, 11, 33. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Jiang, M.; Mirando, A.J.; Yu, H.-M.I.; Hsu, W. Reciprocal regulation of Wnt and Gpr177/mouse Wntless is required for embryonic axis formation. Proc. Natl. Acad. Sci. USA 2009, 106, 18598–18603. [Google Scholar] [CrossRef] [PubMed]
- Coombs, G.S.; Yu, J.; Canning, C.A.; Veltri, C.A.; Covey, T.M.; Cheong, J.K.; Utomo, V.; Banerjee, N.; Zhang, Z.H.; Jadulco, R.C.; et al. WLS-dependent secretion of WNT3A requires Ser209 acylation and vacuolar acidification. J. Cell Sci. 2010, 123, 3357–3367. [Google Scholar] [CrossRef] [PubMed]
- Komekado, H.; Yamamoto, H.; Chiba, T.; Kikuchi, A. Glycosylation and palmitoylation of Wnt-3a are coupled to produce an active form of Wnt-3a. Genes Cells 2007, 12, 521–534. [Google Scholar] [CrossRef] [PubMed]
- Kurayoshi, M.; Yamamoto, H.; Izumi, S.; Kikuchi, A. Post-translational palmitoylation and glycosylation of Wnt-5a are necessary for its signalling. Biochem. J. 2007, 402, 515–523. [Google Scholar] [PubMed]
- van den Heuvel, M.; Harryman-Samos, C.; Klingensmith, J.; Perrimon, N.; Nusse, R. Mutations in the segment polarity genes wingless and porcupine impair secretion of the wingless protein. EMBO J. 1993, 12, 5293–5302. [Google Scholar] [PubMed]
- Bartscherer, K.; Pelte, N.; Ingelfinger, D.; Boutros, M. Secretion of Wnt Ligands Requires Evi, a Conserved Transmembrane Protein. Cell 2006, 125, 523–533. [Google Scholar] [CrossRef] [PubMed]
- Belenkaya, T.Y.; Wu, Y.; Tang, X.; Zhou, B.; Cheng, L.; Sharma, Y.V.; Yan, D.; Selva, E.M.; Lin, X. The Retromer Complex Influences Wnt Secretion by Recycling Wntless from Endosomes to the Trans-Golgi Network. Dev. Cell 2008, 14, 120–131. [Google Scholar] [CrossRef] [PubMed]
- Franch-Marro, X.; Wendler, F.; Guidato, S.; Griffith, J.; Baena-Lopez, A.; Itasaki, N.; Maurice, M.M.; Vincent, J.-P. Wingless secretion requires endosome-to-Golgi retrieval of Wntless/Evi/Sprinter by the retromer complex. Nat. Cell Biol. 2008, 10, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Port, F.; Kuster, M.; Herr, P.; Furger, E.; Bänziger, C.; Hausmann, G.; Basler, K. Wingless secretion promotes and requires retromer-dependent cycling of Wntless. Nat. Cell Biol. 2008, 10, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.T.; Lorenowicz, M.J.; Silhankova, M.; Coudreuse, D.Y.M.; Betist, M.C.; Korswagen, H.C. Wnt Signaling Requires Retromer-Dependent Recycling of MIG-14/Wntless in Wnt-Producing Cells. Dev. Cell 2008, 14, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Gasnereau, I.; Herr, P.; Chia, P.Z.C.; Basler, K.; Gleeson, P.A. Identification of an endocytosis motif in an intracellular loop of Wntless protein, essential for its recycling and the control of Wnt protein signaling. J. Biol. Chem. 2011, 286, 43324–43333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, C.L.; Baum, P.D.; Gu, M.; Jorgensen, E.M.; Clark, S.G.; Garriga, G. C. elegans AP-2 and Retromer Control Wnt Signaling by Regulating MIG-14/Wntless. Dev. Cell 2008, 14, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Chia, J.; Canning, C.A.; Jones, C.M.; Bard, F.A.; Virshup, D.M. WLS Retrograde transport to the endoplasmic reticulum during Wnt secretion. Dev. Cell 2014, 29, 277–291. [Google Scholar] [CrossRef] [PubMed]
- Sivitz, W.I.; DeSautel, S.L.; Kayano, T.; Bell, G.I.; Pessin, J.E. Regulation of glucose transporter messenger RNA in insulin-deficient states. Nature 1989, 340, 72–74. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Holman, G.D. Comparison of GLUT4 and GLUT1 subcellular trafficking in basal and insulin-stimulated 3T3-L1 cells. J. Biol. Chem. 1993, 268, 4600–4603. [Google Scholar] [PubMed]
- Shewan, A.M.; van Dam, E.M.; Martin, S.; Luen, T.B.; Hong, W.; Bryant, N.J.; James, D.E. GLUT4 recycles via a trans-Golgi network (TGN) subdomain enriched in Syntaxins 6 and 16 but not TGN38: Involvement of an acidic targeting motif. Mol. Biol. Cell. 2003, 14, 973–986. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Nakagawa, Y.; Kojima, I.; Shibata, H. Prolonged insulin stimulation down-regulates GLUT4 through oxidative stress-mediated retromer inhibition by a protein kinase CK2-dependent mechanism in 3T3-L1 adipocytes. J. Biol. Chem. 2014, 289, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hodgkinson, V.; Zhu, S.; Weisman, G.A.; Petris, M.J. Advances in the understanding of mammalian copper transporters. Adv. Nutr. 2011, 2, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Petris, M.J.; Camakaris, J.; Greenough, M.; LaFontaine, S.; Mercer, J.F. A C-terminal di-leucine is required for localization of the Menkes protein in the trans-Golgi network. Hum. Mol. Genet. 1998, 7, 2063–2071. [Google Scholar] [CrossRef] [PubMed]
- Vulpe, C.; Levinson, B.; Whitney, S.; Packman, S.; Gitschier, J. Isolation of a candidate gene for Menkes disease and evidence that it encodes a copper-transporting ATPase. Nat. Genet. 1993, 3, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Yi, L.; Kaler, S.G. Direct interactions of adaptor protein complexes 1 and 2 with the copper transporter ATP7A mediate its anterograde and retrograde trafficking. Hum. Mol. Genet. 2015, 24, 2411–2425. [Google Scholar] [CrossRef] [PubMed]
- Tabuchi, M.; Yanatori, I.; Kawai, Y.; Kishi, F. Retromer-mediated direct sorting is required for proper endosomal recycling of the mammalian iron transporter DMT1. J. Cell Sci. 2010, 123, 756–766. [Google Scholar] [CrossRef] [PubMed]
- Touret, N.; Furuya, W.; Forbes, J.; Gros, P.; Grinstein, S. Dynamic traffic through the recycling compartment couples the metal transporter Nramp2 (DMT1) with the transferrin receptor. J. Biol. Chem. 2003, 278, 25548–25557. [Google Scholar] [CrossRef] [PubMed]
- Verges, M.; Sebastian, I.; Mostov, K.E. Phosphoinositide 3-kinase regulates the role of retromer in transcytosis of the polymeric immunoglobulin receptor. Exp. Cell Res. 2007, 313, 707–718. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klinger, S.C.; Siupka, P.; Nielsen, M.S. Retromer-Mediated Trafficking of Transmembrane Receptors and Transporters. Membranes 2015, 5, 288-306. https://doi.org/10.3390/membranes5030288
Klinger SC, Siupka P, Nielsen MS. Retromer-Mediated Trafficking of Transmembrane Receptors and Transporters. Membranes. 2015; 5(3):288-306. https://doi.org/10.3390/membranes5030288
Chicago/Turabian StyleKlinger, Stine C., Piotr Siupka, and Morten S. Nielsen. 2015. "Retromer-Mediated Trafficking of Transmembrane Receptors and Transporters" Membranes 5, no. 3: 288-306. https://doi.org/10.3390/membranes5030288