Prostate-Associated Gene 4 (PAGE4): Leveraging the Conformational Dynamics of a Dancing Protein Cloud as a Therapeutic Target

 ,
,  ,
, 
Abstract
:1. Introduction
1.1. Prostate Cancer
1.2. Benign Prostatic Hyperplasia
1.3. Coincidence of BPH and Prostate Cancer
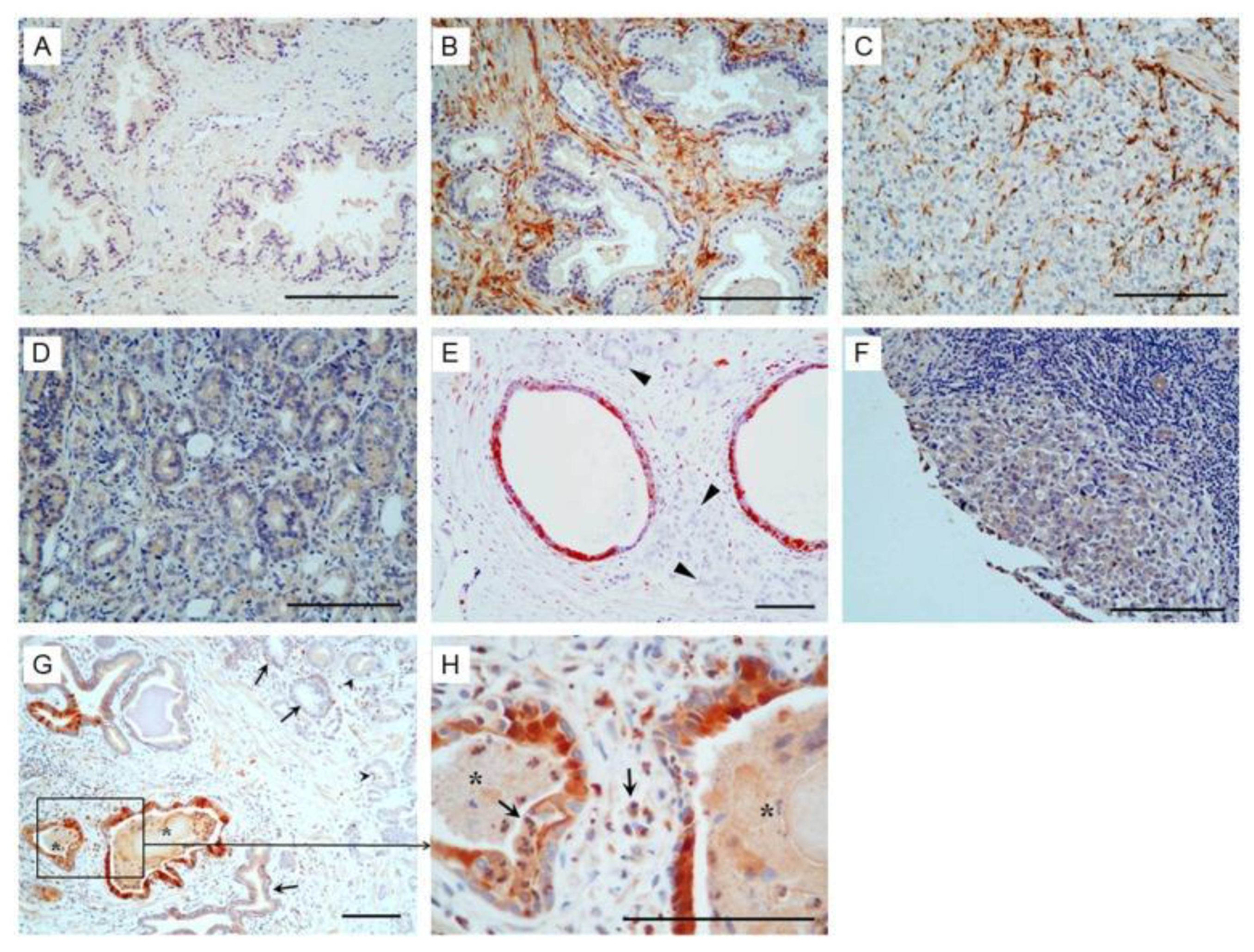
1.4. PAGE4, a Factor Common to Both BPH and PCa
2. Targeting PAGE4 in Symptomatic BPH and Early-Stage ‘Low Risk’ PCa
2.1. Small Molecule Inhibitors (Disrupt PAGE4/AP-1 Interaction)
2.2. Immunotherapy
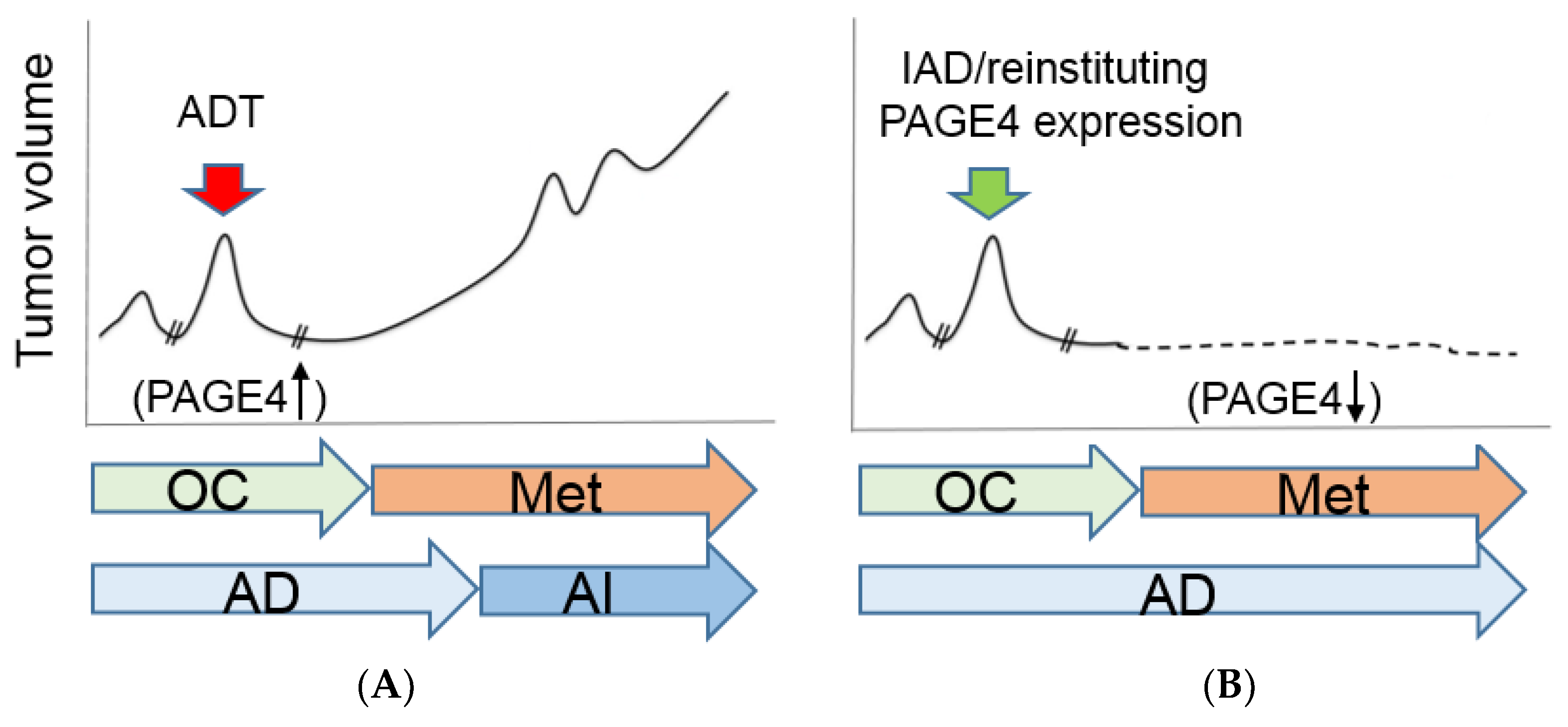
3. Reinstituting Sustained PAGE4 Expression in Advanced PCa
4. Conclusions and Outlook
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Barry, M.J. Screening for prostate cancer—The controversy that refuses to die. N. Engl. J. Med. 2009, 360, 1351–1354. [Google Scholar] [CrossRef] [PubMed]
- Klotz, L.H.; Higano, C.S. Intermittent androgen deprivation therapy for prostate cancer. JAMA Oncol. 2016, 2, 1531–1532. [Google Scholar] [CrossRef] [PubMed]
- Walsh, P.C. Prostate cancer screening. N. Engl. J. Med. 2017, 376, 2401–2402. [Google Scholar] [PubMed]
- Catalona, W.J. Prostate cancer screening. Med. Clin. N. Am. 2018, 102, 199–214. [Google Scholar] [CrossRef] [PubMed]
- Keetch, D.W.; Catalona, W.J.; Smith, D.S. Serial prostatic biopsies in men with persistently elevated serum prostate specific antigen values. J. Urol. 1994, 151, 1571–1574. [Google Scholar] [CrossRef]
- Ellis, W.J.; Brawer, M.K. Repeat prostate needle biopsy: Who needs it? J. Urol. 1995, 153, 1496–1498. [Google Scholar] [CrossRef]
- Djavan, B.; Milani, S.; Remzi, M. Prostate biopsy: Who, how and when. An update. Can. J. Urol. 2005, 1, 44–48. [Google Scholar]
- Cooperberg, M.R.; Moul, J.W.; Carroll, P.R. The changing face of prostate cancer. J. Clin. Oncol. 2005, 23, 8146–8151. [Google Scholar] [CrossRef] [PubMed]
- Holmberg, L.; Bill-Axelson, A.; Garmo, H.; Palmgren, J.; Norlén, B.J.; Adami, H.O.; Johansson, J.E. Prognostic Markers Under Watchful Waiting and Radical Prostatectomy. Hematol. Oncol. Clin. N. Am. 2006, 20, 845–855. [Google Scholar] [CrossRef] [PubMed]
- Cooperberg, M.R.; Carroll, P.R.; Klotz, L. Active surveillance for prostate cancer: Progress and promise. J. Clin. Oncol. 2011, 29, 3669–3676. [Google Scholar] [CrossRef] [PubMed]
- Singer, E.A.; Kaushal, A.; Turkbey, B.; Couvillon, A.; Pinto, P.A.; Parnes, H.L. Active surveillance for prostate cancer: Past, present and future. Curr. Opin. Oncol. 2012, 24, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.H.; Ollendorf, D.A.; Pearson, S.D.; Barry, M.J.; Kantoff, P.W.; Stewart, S.T.; Sweeney, C.J.; Stahl, J.E.; Mcmahon, P.M. Active Surveillance compared with initial treatment for men with low-risk prostate cancer. JAMA 2010, 304, 2373–2380. [Google Scholar] [CrossRef] [PubMed]
- Tosoian, J.J.; Trock, B.J.; Landis, P.; Feng, Z.; Epstein, J.I.; Partin, A.W.; Walsh, P.C.; Carter, H.B. Active surveillance program for prostate cancer: An update of the Johns Hopkins experience. J. Clin. Oncol. 2011, 29, 2185–2190. [Google Scholar] [CrossRef] [PubMed]
- Carter, H.B.; Walsh, P.C.; Landis, P.; Epstein, J.I. Expectant management of nonpalpable prostate cancer with curative intent: Preliminary results. J. Urol. 2002, 167, 1231–1234. [Google Scholar] [CrossRef]
- Azzouzi, A.R.; Vincendeau, S.; Barret, E.; Cicco, A.; Kleinclauss, F.; van der Poel, H.G.; Stief, C.G.; Rassweiler, J.; Salomon, G.; Solsona, E.; et al. Padeliporfin vascular-targeted photodynamic therapy versus active surveillance in men with low-risk prostate cancer (CLIN1001 PCM301): An open-label, phase 3, randomised controlled trial. Lancet Oncol. 2017, 18, 181–191. [Google Scholar] [CrossRef]
- Hamdy, F.C.; Donovan, J.L.; Lane, J.A.; Mason, M.; Metcalfe, C.; Holding, P.; Davis, M.; Peters, T.J.; Turner, E.L.; Martin, R.M.; et al. 10-Year outcomes after monitoring, surgery, or radiotherapy for localized prostate cancer. N. Engl. J. Med. 2016, 375, 1415–1424. [Google Scholar] [CrossRef] [PubMed]
- Corcoran, A.T.; Smaldone, M.C.; Egleston, B.L.; Simhan, J.; Ginzburg, S.; Morgan, T.M.; Walton, J.; Chen, D.Y.T.; Viterbo, R.; Greenberg, R.E.; et al. Comparison of prostate cancer diagnosis in patients receiving unrelated urological and non-urological cancer care. BJU Int. 2013, 112, 161–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulkarni, P.; Uversky, V.N. Cancer/testis antigens: “smart” biomarkers for diagnosis and prognosis of prostate and other cancers. Int. J. Mol. Sci. 2017, 18, 740. [Google Scholar] [CrossRef] [PubMed]
- Wilt, T.J.; Jones, K.M.; Barry, M.J.; Andriole, G.L.; Culkin, D.; Wheeler, T.; Aronson, W.J.; Brawer, M.K. Follow-up of prostatectomy versus observation for early prostate cancer. N. Engl. J. Med. 2017, 377, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Scher, H.I.; Buchanan, G.; Gerald, W.; Butler, L.M.; Tilley, W.D. Targeting the androgen receptor: Improving outcomes for castration-resistant prostate cancer. Endocr. Relat. Cancer 2004, 11, 459–476. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, K.E.; Scher, H.I. Starving the addiction: New opportunities for durable suppression of AR signaling in prostate cancer. Clin. Cancer Res. 2009, 15, 4792–4798. [Google Scholar] [CrossRef] [PubMed]
- Rathkopf, D.; Scher, H.I. Androgen Receptor Aatagonists in castration-resistant prostate Ccncer. Cancer J. 2013, 19, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Pham, S.; Deb, S.; Ming, D.S.; Adomat, H.; Hosseini-Beheshti, E.; Zoubeidi, A.; Gleave, M.; Guns, E.S.T. Next-generation steroidogenesis inhibitors, dutasteride and abiraterone, attenuate but still do not eliminate androgen biosynthesis in 22RV1 cells in vitro. J. Steroid Biochem. Mol. Biol. 2014, 144, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Bambury, R.M.; Scher, H.I. Enzalutamide: Development from bench to bedside. Urol. Oncol. Semin. Orig. Investig. 2015, 33, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Wolin, K.Y.; Grubb, R.L.; Pakpahan, R.; Ragard, L.; Mabie, J.; Andriole, G.L.; Sutcliffe, S. Physical activity and benign prostatic hyperplasia-related outcomes and nocturia. Med. Sci. Sports Exerc. 2014, 47, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Oesterling, J.E. Benign prostatic hyperplasia: A review of its histogenesis and natural history. Prostate Suppl. 1996, 6, 67–73. [Google Scholar] [CrossRef]
- Lepor, H. Pathophysiology of lower urinary tract symptoms in the aging male population. Rev. Urol. 2005, 7 (Suppl. 7), S3–S11. [Google Scholar] [PubMed]
- Bostwick, D.G.; Cooner, W.H.; Denis, L.; Jones, G.W.; Scardino, P.T.; Murphy, G.P. The association of benign prostatic hyperplasia and cancer of the prostate. Cancer 1992, 70, 291–301. [Google Scholar] [CrossRef]
- Haas, G.P.; Delongchamps, N.; Brawley, O.W.; Wang, C.Y.; de la Roza, G. The worldwide epidemiology of prostate cancer: Perspectives from autopsy studies. Can. J. Urol. 2008, 15, 3866–3871. [Google Scholar] [PubMed]
- Sommers, S.C. Endocrine changes with prostatic carcinoma. Cancer 1957, 10, 345–358. [Google Scholar] [CrossRef] [Green Version]
- Ørsted, D.D.; Bojesen, S.E.; Nielsen, S.F.; Nordestgaard, B.G. Association of clinical benign prostate hyperplasia with prostate cancer incidence and mortality revisited: A nationwide cohort study of 3,009,258 men. Eur. Urol. 2011, 60, 691–698. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, P.; Getzenberg, R.H. Disorder, promiscuous interactions, and stochasticity Rrgulate state switching in the unstable prostate. J. Cell. Biochem. 2016, 2240, 2235–2240. [Google Scholar] [CrossRef] [PubMed]
- Orsted, D.D.; Bojesen, S.E. The link between benign prostatic hyperplasia and prostate cancer. Nat. Rev. Urol. 2013, 10, 49–54. [Google Scholar] [CrossRef] [PubMed]
- De Nunzio, C.; Kramer, G.; Marberger, M.; Montironi, R.; Nelson, W.; Schröder, F.; Sciarra, A.; Tubaro, A. The controversial relationship between benign prostatic hyperplasia and prostate cancer: The role of inflammation. Eur. Urol. 2011, 60, 106–117. [Google Scholar] [CrossRef] [PubMed]
- Sfanos, K.; Marzo, A. Prostate cancer and inflammation. Histopathologyogy 2012, 60, 199–215. [Google Scholar] [CrossRef] [PubMed]
- Gurel, B.; Lucia, M.S.; Thompson, I.M.; Goodman, P.J.; Tangen, C.M.; Kristal, A.R.; Parnes, H.L.; Hoque, A.; Lippman, S.M.; Sutcliffe, S.; et al. Chronic inflammation in benign prostate tissue is associated with high-grade prostate cancer in the placebo arm of the prostate cancer prevention trial. Cancer Epidemiol. Biomarkers Prev. 2014, 23, 847–856. [Google Scholar] [CrossRef] [PubMed]
- Sfanos, K.S.; Yegnasubramaniam, S.; Nelson, W.G.; Marzo, A.M. De The inflammatory microenvironment and microbiome in prostate cancer development. Nat. Rev. Urol. 2018, 15, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Miah, S.; Catto, J. BPH and prostate cancer risk. Indian J. Urol. 2014, 30, 214–218. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Fang, X.; Ma, Y.; Xianyu, J. Benign Ppostatic Hhperplasia and the Rrsk of Ppostate Ccncer and bladder cancer. Medicine 2016, 95, e3493. [Google Scholar] [CrossRef] [PubMed]
- Old, L.J.; Chen, Y.-T. New paths in human cancer serology. J. Exp. Med. 1998, 187, 1163–1167. [Google Scholar] [CrossRef] [PubMed]
- Scanlan, M.J.; Simpson, A.J.G.; Old, L.J. The cancer/testis genes: Review, standardization, and commentary. Cancer Immun. 2004, 4, 1. [Google Scholar]
- Simpson, A.J.G.; Caballero, O.L.; Jungbluth, A.; Chen, Y.T.; Old, L.J. Cancer/testis antigens, gametogenesis and cancer. Nat. Rev. Cancer 2005, 5, 615–625. [Google Scholar] [CrossRef] [PubMed]
- Rajagopalan, K.; Qiu, R.; Mooney, S.M.; Rao, S.; Shiraishi, T.; Sacho, E.; Huang, H.; Shapiro, E.; Weninger, K.R.; Kulkarni, P. The Stress-response protein prostate-associated gene 4, interacts with c-Jun and potentiates its transactivation. Biochim. Biophys. Acta 2014, 1842, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Gao, D.; Kim, J.J.; Shiraishi, T.; Terada, N.; Kakehi, Y.; Kong, C.; Getzenberg, R.; Kulkarni, P. Prostate-associated gene 4 (PAGE4) protects cells against stress by elevating p21 and suppressing reactive oxygen species production. Am. J. Clin. Exp. Urol. 2013, 1, 39–52. [Google Scholar] [PubMed]
- Nelson, W.G.; De Marzo, A.M.; Isaacs, W.B. Prostate cancer. N. Engl. J. Med. 2003, 349, 366–381. [Google Scholar] [CrossRef] [PubMed]
- Suyama, T.; Shiraishi, T.; Zeng, Y.; Yu, W.; Parekh, N.; Vessella, R.L.; Luo, K.; Getzenberg, R.; Kulkarni, P. Expression of cancer/testis antigens in prostate cancer is associated with disease progression. Prostate 2010, 70, 1778–1787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
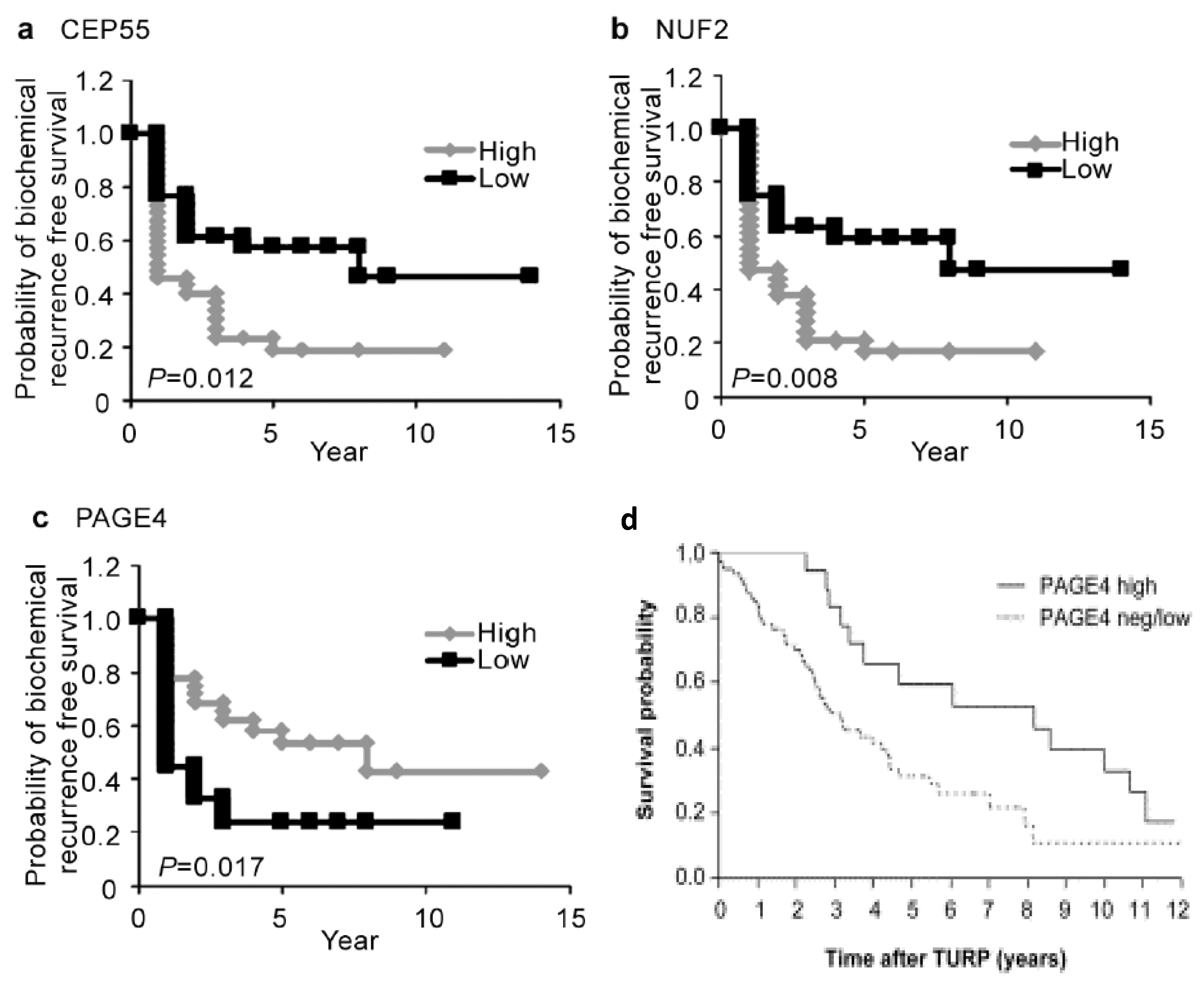
- Shiraishi, T.; Terada, N.; Zeng, Y.; Suyama, T.; Luo, J.; Trock, B.; Kulkarni, P.; Getzenberg, R. Cancer/Testis Antigens as potential predictors of biochemical recurrence of prostate cancer following radical prostatectomy. J. Transl. Med. 2011, 14, 153. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, P.; Dunker, K.A.; Weninger, K.R.; Orban, J. Prostate-associated gene 4 (PAGE4), an intrinsically disordered cancer/testis antigen, is a novel therapeutic target for prostate cancer. Asian J. Androl. 2016, 18, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; He, Y.; Yang, F.; Mooney, S.M.; Getzenberg, R.H.; Orban, J.; Kulkarni, P. The cancer/testis antigen prostate-associated gene 4 (PAGE4) is a highly intrinsically disordered protein. J. Biol. Chem. 2011, 286, 13985–13994. [Google Scholar] [CrossRef] [PubMed]
- Prakash, K.; Pirozzi, G.; Elashoff, M.; Munger, W.; Waga, I.; Dhir, R.; Kakehi, Y.; Getzenberg, R.H. Symptomatic and asymptomatic benign prostatic hyperplasia: Molecular differentiation by using microarrays. Proc. Natl. Acad. Sci. USA 2002, 99, 7598–7603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin-Tsai, O.; Clark, P.; Miller, N.; Fowke, J.; Hameed, O.; Hayward, S.; Strand, D. Surgical intervention for symptomatic benign prostatic hyperplasia is correlated with expression of the AP-1 transcription factor network. Prostate 2014, 74, 669–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgos, J. Inflammation and prostatic diseases: From bench to bedside. Eur. Urol. Suppl. 2015, 14, e1453–e1454. [Google Scholar] [CrossRef]
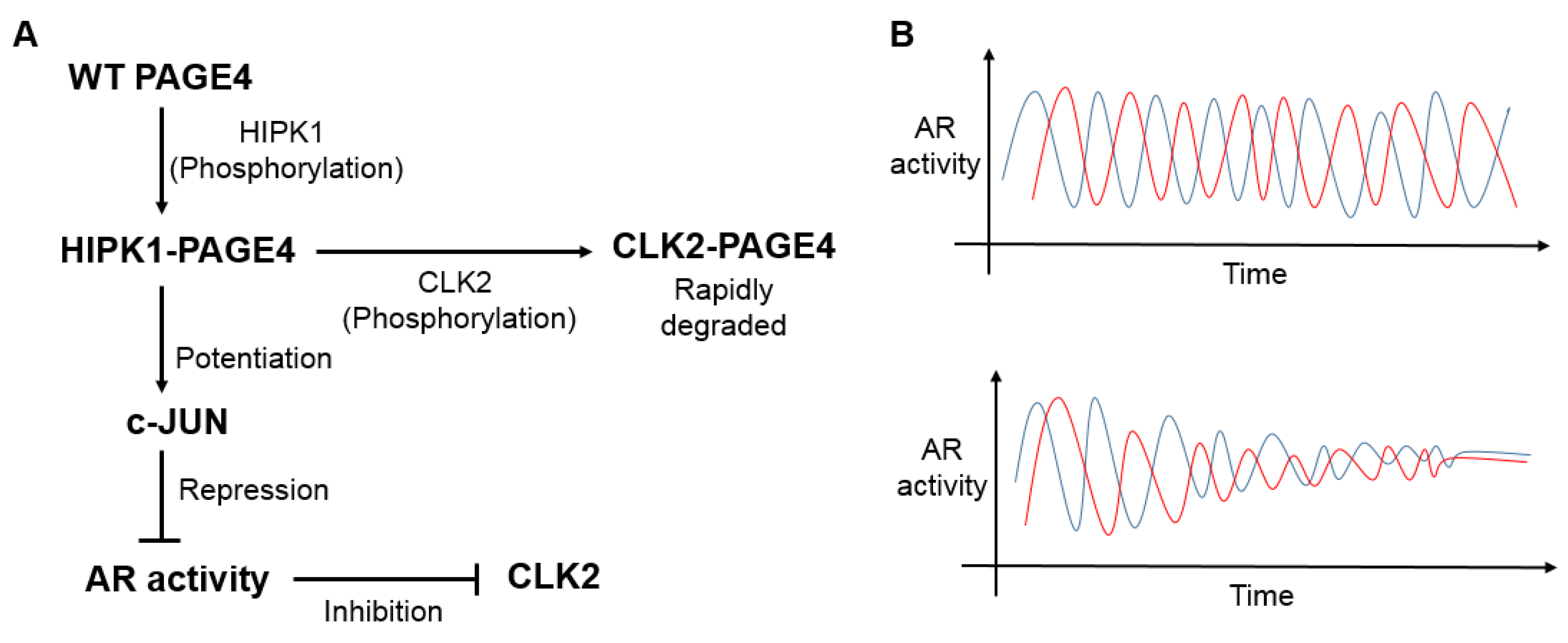
- Mooney, S.M.; Qiu, R.; Kim, J.J.; Sacho, E.J.; Rajagopalan, K.; Johng, D.; Shiraishi, T.; Kulkarni, P.; Weninger, K.R. Cancer/testis antigen PAGE4, a regulator of c-Jun transactivation, is phosphorylated by homeodomain-interacting protein kinase 1, a component of the stress-response pathway. Biochemistry 2014, 53, 1670–1679. [Google Scholar] [CrossRef] [PubMed]
- Leppa, S.; Bohmann, D. Diverse functions of JNK signaling and c-Jun in stress response and apoptosis. Oncogene 1999, 18, 6158–6162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouyang, X.; Jessen, W.J.; Al-Ahmadie, H.; Serio, A.M.; Lin, Y.; Shih, W.-J.; Reuter, V.E.; Scardino, P.T.; Shen, M.M.; Aronow, B.J.; et al. Activator protein-1 transcription factors are associated with progression and recurrence of prostate cancer. Cancer Res. 2008, 68, 2132–2144. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, P.; Jolly, M.K.; Jia, D.; Mooney, S.M.; Bhargava, A.; Kagohara, L.T.; Chen, Y.; Hao, P.; He, Y.; Veltri, R.W.; et al. Phosphorylation-induced conformational dynamics in an intrinsically disordered protein and potential role in phenotypic heterogeneity. Proc. Natl. Acad. Sci. USA 2017, 114, E2644–E2653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajagopalan, K.; Mooney, S.M.; Parekh, N.; Getzenberg, R.H.; Kulkarni, P. A majority of the Cancer/Testis Antigens are intrinsically disordered proteins. J. Cell. Biochem. 2011, 112, 3256–3267. [Google Scholar] [CrossRef] [PubMed]
- Uversky, V.N. Dancing protein clouds: The strange biology and chaotic physics of intrinsically disordered proteins. J. Biol. Chem. 2016, 291, 6681–6688. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudabadi, G.; Rajagopalan, K.; Getzenberg, R.H.; Hannenhalli, S.; Rangarajan, G.; Kulkarni, P. Intrinsically disordered proteins and conformational noise Implications in cancer. Cell Cycle 2013, 12, 26–31. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Chen, Y.; Mooney, S.M.; Rajagopalan, K.; Bhargava, A.; Sacho, E.; Weninger, K.; Bryan, P.N.; Kulkarni, P.; Orban, J. Phosphorylation-induced conformational ensemble switching in an intrinsically disordered Cancer/Testis Antigen. J. Biol. Chem. 2015, 290, 25090–25102. [Google Scholar] [CrossRef] [PubMed]
- Heller, G.T.; Aprile, F.A.; Vendruscolo, M. Methods of probing the interactions between small molecules and disordered proteins. Cell. Mol. Life Sci. 2017, 74, 3225–3243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammoudeh, D.I.; Follis, A.V.; Prochownik, E.V.; Metallo, S.J. Multiple independent binding sites for small-molecule inhibitors on the oncoprotein c-Myc. J. Am. Chem. Soc. 2009, 131, 7390–7401. [Google Scholar] [CrossRef] [PubMed]
- Berg, T. Small-molecule modulators of c-Myc/Max and Max/Max interactions. Curr. Top. Microbiol. Immunol. 2010, 348, 139–149. [Google Scholar]
- Iconaru, L.I.; Ban, D.; Bharatham, K.; Ramanathan, A.; Zhang, W.; Shelat, A.A.; Zuo, J.; Kriwacki, R.W. Discovery of small molecules that inhibit the disordered protein, p27Kip1. Sci. Rep. 2015, 5, 15686. [Google Scholar] [CrossRef] [PubMed]
- Neira, J.L.; Bintz, J.; Arruebo, M.; Rizzuti, B.; Bonacci, T.; Vega, S.; Lanas, A.; Velázquez-Campoy, A.; Iovanna, J.L.; Abián, O. Identification of a drug targeting an intrinsically disordered protein involved in pancreatic adenocarcinoma. Sci. Rep. 2017, 7, 39732. [Google Scholar] [CrossRef] [PubMed]
- Sumanasuriya, S.; De Bono, J. Treatment of advanced prostate cancer—A review of current therapies and future promise. Cold Spring Harb. Perspect. Med. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Handy, C.E.; Antonarakis, E.S. Sipuleucel-T for the treatment of prostate cancer: Novel insights and future directions. Future Oncol. 2017, 14, 907–917. [Google Scholar] [CrossRef] [PubMed]
- Komura, K.; Sweeney, C.J.; Inamoto, T.; Ibuki, N.; Azuma, H.; Kantoff, P.W. Current treatment strategies for advanced prostate cancer. Int. J. Urol. 2018, 25, 220–231. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, P.; Shiraishi, T.; Rajagopalan, K.; Kim, R.; Mooney, S.M.; Getzenberg, R.H. Cancer/testis antigens and urological malignancies. Nat. Rev. Urol. 2012, 9, 386–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokokawa, J.; Bera, T.K.; Palena, C.; Cereda, V.; Remondo, C.; Gulley, J.L.; Arlen, P.M.; Pastan, I.; Schlom, J.; Tsang, K.Y. Identification of cytotoxic T-lymphocyte epitope(s) and its agonist epitope(s) of a novel target for vaccine therapy (PAGE4). Int. J. Cancer 2007, 121, 595–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geldmacher, A.; Freier, A.; Losch, F.O.; Walden, P. Therapeutic vaccination for cancer immunotherapy: Antigen selection and clinical responses. Hum. Vaccines 2011, 7, 115–119. [Google Scholar] [CrossRef]
- Jolly, M.K.; Kulkarni, P.; Weninger, K.; Orban, J.; Levine, H. Phenotypic plasticity, bet-hedging, and androgen independence in prostate cancer: Role of non-genetic heterogeneity. Front. Oncol. 2018, 8, 50. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Roy, S.; Jolly, M.K.; Bocci, F.; Schafer, N.; Tsai, M.-Y.; Kulkarni, P.; Chen, Y.; He, Y.; Orban, J.; et al. PAGE4 and conformational switching: Insights from molecular dynamics simulations and implications for prostate cancer. J. Mol. Biol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Beyzavi, A.; Krakowiak, J.; Patel, N.; Khalil, A.S.; Pincus, D. Hsf1 Phosphorylation generates cell-to-cell variation in Hsp90 levels and promotes phenotypic plasticity. Cell Rep. 2018, 22, 3099–3106. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.M.; Vessella, R.L.; Morrissey, C.; Brown, L.G.; Coleman, I.M.; Higano, C.S.; Mostaghel, E.A.; Zhang, X.; True, L.D.; Lam, H.M.; et al. LuCaP prostate cancer patient-derived xenografts reflect the molecular heterogeneity of advanced disease and serve as models for evaluating cancer Therapeutics. Prostate 2017, 77, 654–671. [Google Scholar] [CrossRef] [PubMed]
- Isaacs, J.T.; Coffey, D.S. Adaptation versus selection as the mechanism responsible for the relapse of prostatic cancer to androgen ablation therapy as studied in the Dunning R-3327-H adenocarcinoma. Cancer Res. 1981, 41, 5070–5075. [Google Scholar] [PubMed]
- Gallaher, J.A.; Enriquez-Navas, P.M.; Luddy, K.A.; Gatenby, R.A.; Anderson, A.R.A. Heterogeneity and evolutionary dynamics modulate time to recurrence in continuous and adaptive cancer therapies. Cancer Res. 2018, 78, 2127–2139. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.B.; Zhao, L.; Zhang, L.; Zhao, K.N. Virus, oncolytic virus and human prostate cancer. Curr. Cancer Drug Targets 2017, 17, 522–533. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.; Gujar, S. Potentiating prostate cancer immunotherapy with oncolytic viruses. Nat. Rev. Urol. 2018, 15, 235–250. [Google Scholar] [CrossRef] [PubMed]
- Barry, J.N.; Vertegel, A.A. Nanomaterials for protein-mediated therapy and delivery. Nano Life 2013, 3, 1343001. [Google Scholar] [CrossRef] [PubMed]
- Lohcharoenkal, W.; Wang, L.; Chen, Y.C.; Rojanasaku, Y. Protein nanoparticles as drug delivery carriers for cancer therapy. BioMed Res. Int. 2014, 2014, 180549. [Google Scholar] [CrossRef] [PubMed]
- Sampson, N.; Ruiz, C.; Zenzmaier, C.; Bubendorf, L.; Berger, P. PAGE4 positivity is associated with attenuated AR signaling and predicts patient survival in hormone-naive prostate cancer. Am. J. Pathol. 2012, 181, 1443–1454. [Google Scholar] [CrossRef] [PubMed]
- Cannon, G.W.; Mullins, C.; Lucia, M.S.; Hayward, S.W.; Lin, V.; Liu, B.C.S.; Slawin, K.; Rubin, M.A.; Getzenberg, R.H. A preliminary study of JM-27: A serum marker that can specifically identify men with symptomatic benign prostatic hyperplasia. J. Urol. 2007, 177, 610–614. [Google Scholar] [CrossRef] [PubMed]
- Bechis, S.K.; Otsetov, A.G.; Ge, R.; Olumi, A.F. Personalized medicine for the management of benign prostatic hyperplasia. J. Urol. 2014, 192, 16–23. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CTA | BPH | Primary PCa | Met PCa |
|---|---|---|---|
| PAGE4 (205564_at) | 6732.5 | 5702.8 | 56.6 |
| CSAG2 (220445_s_at) | 16.6 | 20.6 | 1026.4 |
| MAGEA2 (214603_at) | 46.7 | 42.7 | 1343.9 |
| MAGEA6 (214612_x_at) | 30.5 | 16.2 | 2785.1 |
| MAGEA12 (210467_x_at) | 34.9 | 52.9 | 1168.4 |
| TBP (203135_at) | 608.66 | 577.58 | 882.2 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salgia, R.; Jolly, M.K.; Dorff, T.; Lau, C.; Weninger, K.; Orban, J.; Kulkarni, P. Prostate-Associated Gene 4 (PAGE4): Leveraging the Conformational Dynamics of a Dancing Protein Cloud as a Therapeutic Target. J. Clin. Med. 2018, 7, 156. https://doi.org/10.3390/jcm7060156
Salgia R, Jolly MK, Dorff T, Lau C, Weninger K, Orban J, Kulkarni P. Prostate-Associated Gene 4 (PAGE4): Leveraging the Conformational Dynamics of a Dancing Protein Cloud as a Therapeutic Target. Journal of Clinical Medicine. 2018; 7(6):156. https://doi.org/10.3390/jcm7060156
Chicago/Turabian StyleSalgia, Ravi, Mohit Kumar Jolly, Tanya Dorff, Clayton Lau, Keith Weninger, John Orban, and Prakash Kulkarni. 2018. "Prostate-Associated Gene 4 (PAGE4): Leveraging the Conformational Dynamics of a Dancing Protein Cloud as a Therapeutic Target" Journal of Clinical Medicine 7, no. 6: 156. https://doi.org/10.3390/jcm7060156