Conditions Affecting Shelf-Life of Inoculated Legume Seed

Abstract
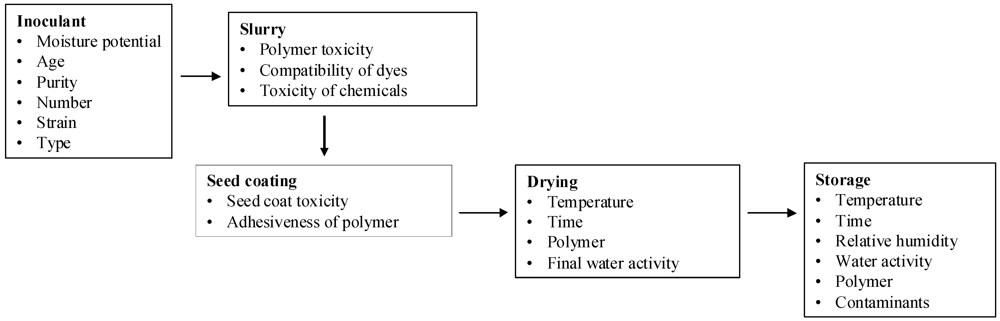
:1. Introduction

2. Results and Discussion
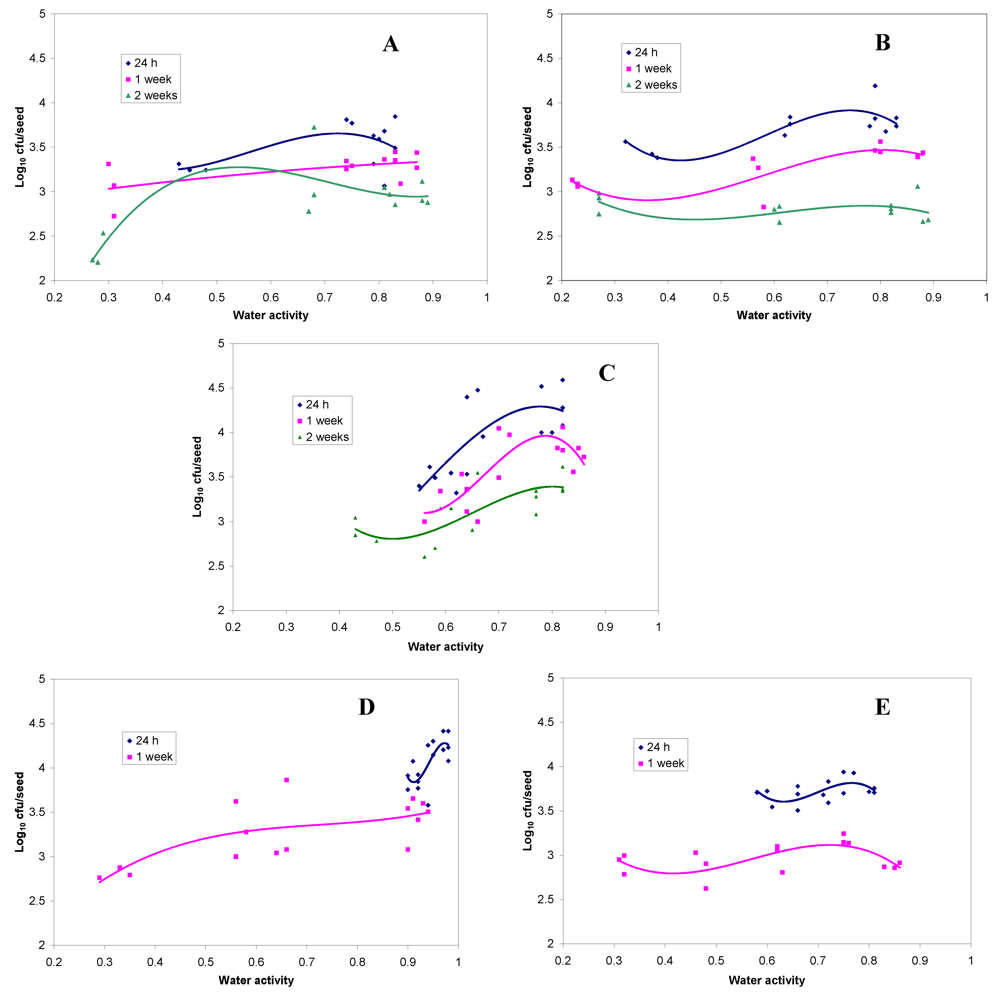
2.1. Survival of Rhizobia on Laboratory Coated Seed after Storage at Different Relative Humidity

2.2. Survival of Rhizobia on Commercially Coated Legume Seed

{kind=link}
{kind=link}
{kind=link}
| Sample | Seed | Seed company | Time since coating | Log10 no./seed | Contaminants |
|---|---|---|---|---|---|
| A | Lucerne | 1 | 2 weeks | 1.10 | No |
| B | Lucerne | 1 | 2 weeks | 1.93 | No |
| C | Lucerne | 1 | 2 weeks | 2.10 | No |
| D | Lucerne | 1 | 2 weeks | 2.17 | No |
| E | Lucerne | 2 | Unknown | TCTC | Yes |
| F | Lucerne | 2 | 1 week | 4.12 | Yes |
| G | Subteranean clover | 2 | 1 week | TCTC | Yes |
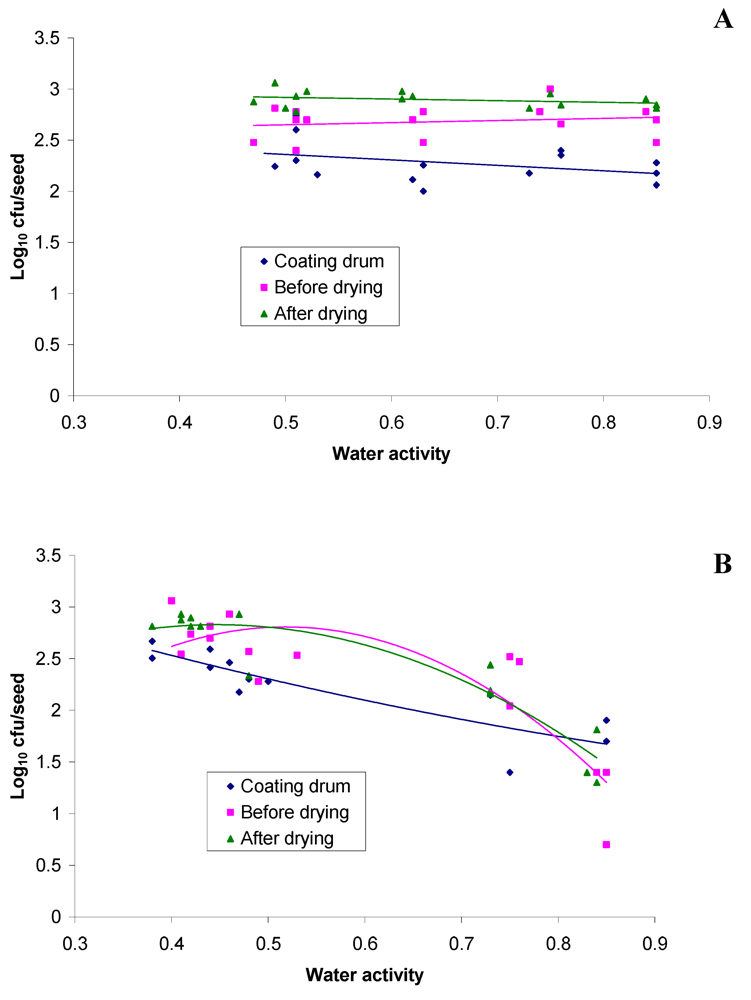
2.3. Survival of RRI128 on Commercially Coated Lucerne after Storage at Different Relative Humidity
| Seed sample | Log10 cfu/seed | Log10 cfu/seed | Percent | Water activity |
|---|---|---|---|---|
| 3 days after coating | 16 days after coating | survival (%) | 16 days after coating | |
| Coating drum | 4.08a | 3.11a | 12.3 | 0.84a |
| Before drying | 4.03a | 2.91a | 9.6 | 0.79b |
| After drying | 3.87b | 3.24a | 24.3 | 0.59c |

| Seed sample | Recovery at highest RH (%) | Change in Aw at highest RH | Recovery at lowest RH (%) | Change in Aw at lowest RH |
|---|---|---|---|---|
| Coating drum | 0.49 | +0.01 | 3.07 | −0.42 |
| Before drying | 0.18 | +0.06 | 8.29 | −0.36 |
| After drying | 0.51 | +0.25 | 10.95 | −0.18 |
3. Experimental Section
3.1. Survival of Rhizobia on Coated Seed Stored at Different Relative Humidities
3.2. Inoculation of Seed in the Laboratory
3.3. Seed Collected from Commercial Seed Companies
3.4. Storage of Inoculated Seed at Different Relative Humidities
| Saturated salt solution (g/L) | Relative humidity (%) |
|---|---|
| Potassium hydroxide (2000) | 10 |
| Potassium acetate (2500) | 23 |
| Magnesium chloride (2800) | 33 |
| Magnesium nitrate (2200) | 54 |
| Sodium chloride (500) | 75 |
| Potassium chloride (500) | 85 |
3.5. Determination of Viable Numbers of Rhizobia and Measurement of Water Activity
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
References
- Hynes, R.K.; Boyetchko, S.M. Research initiatives in the art and science of biopesticide formulations. Soil Biol. Biochem. 2006, 38, 845–849. [Google Scholar] [CrossRef]
- Thompson, J.A. Survival of root-nodule bacteria on inoculated seed. In Microbiology in Action; Murrell, W.G., Kennedy, I.R., Eds.; Research Studies Press: Letchworth, UK, 1988; pp. 67–80. [Google Scholar]
- Roughley, R.J.; Gemell, L.G.; Thompson, J.A.; Brockwell, J. The number of Bradyrhizobium sp. (lupinus) applied to seed and its effect on rhizosphere colonisation, nodulation and yield of lupin. Soil Biol. Biochem. 1993, 25, 1453–1458. [Google Scholar] [CrossRef]
- Howieson, J.; Herridge, D. Forward special issue. Aust. J. Exp. Agric. 2005, 56, 45. [Google Scholar]
- Howieson, J.; Ballard, R. Optimising the legume symbiosis in stressful and competitive environments within southern Australia—Some contemporary thoughts Soil Biol. Biochem. 2004, 36, 1261–1273. [Google Scholar]
- Ireland, J.A.; Vincent, J.M. A quantitative study of competition for nodule formation. In Proceedings of the Ninth International Congress of Soil Science Transactions, Adelaide; International Society of Soil Science and Angus and Robertson: Sydney, Australia, 1968; Volume 2, pp. 85–93. [Google Scholar]
- Nandasena, K.G.; O’Hara, G.W.; Tiwari, R.P.; Sezmis, E.; Howieson, J.G. In situ lateral transfer of symbiosis islands results in rapid evolution of diverse competitive strains of mesorhizobia suboptimal in symbiotic nitrogen fixation on the pasture legume Biserrula pelecinus L. Environ. Microbiol. 2007, 9, 2496–2511. [Google Scholar] [CrossRef]
- Brockwell, J.; Herridge, D.F.; Roughley, R.J.; Thompson, J.A.; Gault, R.R. Studies on seed pelleting as an aid to legume seed inoculation: Examination of preinoculated seed. Aust. J. Exp. Agric. Anim. Husb. 1975, 15, 780–787. [Google Scholar] [CrossRef]
- Gemell, L.G.; Hartley, E.J.; Herridge, D.F. Point-of-sale evaluation of preinoculated and custom-inoculated pasture legume seed. Aust. J. Exp. Agric. 2005, 45, 161–169. [Google Scholar] [CrossRef]
- Brockwell, J. Application of legume seed inoculants. In A Treatise on Dinitrogen Fixation; Hardy, R.W.F., Gibson, A.H., Eds.; Wiley: Sydney, Australia, 1977; pp. 277–309. [Google Scholar]
- Deaker, R.; Roughley, R.J.; Kennedy, I.R. Desiccation tolerance of rhizobia when protected by synthetic polymers. Soil Biol. Biochem. 2007, 39, 573–580. [Google Scholar] [CrossRef]
- Vincent, J.M.; Thompson, J.A.; Donovan, K.O. Death of root nodule bacteria on drying. Aust. J. Agric. Res. 1962, 13, 258–270. [Google Scholar] [CrossRef]
- Deaker, R.; Roughley, R.J.; Kennedy, I.R. Legume seed inoculation technology—A review. Soil Biol. Biochem. 2004, 36, 1275–1288. [Google Scholar] [CrossRef]
- Mary, P.; Ochin, D.; Tailliez, R. Rates of drying and survival of Rhizobium meliloti strains during storage at different relative humidities. Appl. Environ. Microbiol. 1985, 50, 207–211. [Google Scholar]
- Mary, P.; Dupuy, N.; Dolhem-Biremon, C.; Defives, C.; Tailliez, R. Differences among Rhizobium meliloti and Bradyrhizobium japonicum strains in tolerance to desiccation and storage at different relative humidities. Soil Biol. Biochem. 1994, 26, 1125–1132. [Google Scholar] [CrossRef]
- Kosanke, J.W.; Osburn, R.M.; Shuppe, G.I.; Smith, R.S. Slow rehydration improves the recovery of dried bacterial population. Can. J. Microbiol. 1992, 38, 520–525. [Google Scholar] [CrossRef]
- Mugnier, J.; Jung, G. Survival of bacteria and fungi in relation to water activity and the solvent properties of water in biopolymer. Appl. Environ. Microbiol. 1985, 50, 108–114. [Google Scholar]
- McInnes, A.; Date, R.A. Improving the survival of rhizobia on Desmanthus and Stylosanthes seed at high temperature. Aust. J. Exp. Agric. 2005, 45, 171–182. [Google Scholar] [CrossRef]
- Dart, P.J.; Roughley, R.J.; Chandler, M.R. Peat culture of Rhizobium trifolii—An examination by electron microscopy. J. Appl. Bacteriol. 1969, 32, 352–357. [Google Scholar] [CrossRef]
- Materon, L.A.; Weaver, R.W. Inoculant maturity influences survival of rhizobia on seed. Appl. Environ. Microbiol. 1985, 49, 465–467. [Google Scholar]
- Feng, L.; Roughley, R.J.; Copeland, L. Morphological changes of rhizobia in peat cultures. Appl. Environ. Microbiol. 2002, 68, 1064–1070. [Google Scholar] [CrossRef]
- Gemell, L.G.; Hartley, E.; Hartley, J.; Deaker, R. Compatibility with rhizobia of polymer adhesives and colourants used in preinoculated legume pasture seed. In Proceedings of the 17th International Congress on Nitrogen Fixation, Fremantle, Australia, 17 November–1 December 2011.
- Thompson, J.A. Inhibition of nodule bacteria by an antibiotic from legume seed coats. Nature 1960, 187, 619–620. [Google Scholar] [CrossRef]
- Roughley, R.J.; Vincent, J.M. Growth and survival of Rhizobium spp. in peat culture. J. Appl. Bacteriol. 1967, 30, 362–376. [Google Scholar] [CrossRef]
- Bell, L.N.; Labuza, T.P. Moisture Sorption: Practical Aspects of Isotherm Measurement and Use; American Association of Cereal Chemists, Inc.: St. Paul, MN, USA, 2000. [Google Scholar]
- Kremer, R.J.; Peterson, H.L. Effect of inoculant carrier on survival of Rhizobium on inoculated seed. Soil Sci. 1982, 134, 117–124. [Google Scholar]
- Date, R.A. Growth and Survival of Root Nodule Bacteria. Unpublished MSc Thesis, Department of Microbiology, University of Sydney, Sydney, Australia, 1959. [Google Scholar]
- Pixton, S.W.; Warburton, S. Moisture content/relative humidity equilibrium of some cereal grains at different temperatures. J. Stored Prod. Res. 1971, 6, 283–293. [Google Scholar] [CrossRef]
- Vincent, J.M. A Manual for the Practical Study of Root Nodule Bacteria; International Biological Program Handbook number 15; Blackwell Scientific Publications: Oxford, UK, 1970. [Google Scholar]
- Greenspan, L. Humidity fixed points of binary saturated aqueous solutions. J. Res. Nat. Bur. Stand. 1977, 81A, 89–96. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Deaker, R.; Hartley, E.; Gemell, G. Conditions Affecting Shelf-Life of Inoculated Legume Seed. Agriculture 2012, 2, 38-51. https://doi.org/10.3390/agriculture2010038
Deaker R, Hartley E, Gemell G. Conditions Affecting Shelf-Life of Inoculated Legume Seed. Agriculture. 2012; 2(1):38-51. https://doi.org/10.3390/agriculture2010038
Chicago/Turabian StyleDeaker, Rosalind, Elizabeth Hartley, and Greg Gemell. 2012. "Conditions Affecting Shelf-Life of Inoculated Legume Seed" Agriculture 2, no. 1: 38-51. https://doi.org/10.3390/agriculture2010038